Introduction
The intervertebral disc (IVD) consists of three
distinct sections: The outer annulus fibrosus (AF) which contains
abundant type I collagens; the inner nucleus pulposus (NP) which is
a gelatinous substance containing type II collagen and
proteoglycans; and the cartilage end plates that connect the
adjacent vertebral bodies (1–3).
With age, morphological and molecular alterations in the IVD may
induce progressive degeneration and pathologic alterations of this
tissue (4). IVD degeneration may
eventually result in conditions including disc herniation, facet
joint osteoarthritis, spinal canal stenosis, segmental instability
and spondylolisthesis. The mechanisms underlying IVD degeneration
are considered to be associated with decreased cell viability and
adhesion, reduced diffusion of nutrients and proteoglycan synthesis
(5,6). Accelerated aging of the IVD due to
abnormal apoptosis of IVD cells is a primary cytological process
(7–9), and apoptosis of IVD cells serves a
key role in the process of IVD degeneration.
Tumor necrosis factor (TNF)-α is an important member
of the TNF superfamily, and is involved in immunity, cellular
remodeling, apoptosis and cell survival (10). It acts primarily via TNF receptor-1
to induce apoptosis by activating caspases through
mitochondria-dependent and -independent pathways (11). In addition to secretion by
infiltrating immune cells, resident NP and AF cells produce high
levels of TNF-α and interleukin (IL)-1β to promote inflammation
(12–14). TNF-α has been confirmed to have a
central role in degenerative osteoarthropathy (15). Additionally, it has been reported
that TNF-α may induce apoptosis of NP cells in vitro
(16). Furthermore, TNF-α released
during the inflammatory process induced by the herniated IVD serves
a fundamental role in the development of mechanical and thermal
hyperalgesia (16–18). The inhibition of TNF-α-induced
abnormal apoptosis of human NP cells may delay the degeneration of
the IVD, preventing degenerative scoliosis and other spinal
degenerative diseases.
17β-estradiol (17β-E2) has been extensively
investigated due to its anti-apoptotic activity (19–22).
Bozzo et al (23) reported
that 17β-E2 protects nerve cells from β-amyloid peptide-induced
apoptosis, and that 17β-E2 exerts anti-apoptotic effects by
upregulating the expression of essential cell membrane components
including α1β1 integrin, and affecting cell cycle progression. In
addition, 17β-E2 has been reported to inhibit caspase-3/9 activity
and increase B-cell lymphoma 2, cyclin D1 and survivin expression
(24–26). Wang et al (6) and Yang et al (22) demonstrated that 17β-E2 protects rat
IVD cells from apoptosis induced by IL-1β and levofloxacin.
However, whether 17β-E2 inhibits TNF-α-induced apoptosis in human
NP cells, and the concentration-response effect of 17β-E2 on
TNF-α-induced human NP cell apoptosis, remains unclear.
Therefore, the aim of the present study was to
investigate whether 17β-E2 modulates apoptosis induced by TNF-α in
human NP cells, the concentration-response effect and whether
17β-E2 exerts protective effects via the caspase-3/poly
(ADP-ribose) polymerase (PARP) pathway.
Materials and methods
Reagents
Human NP cells, NP Cell Growth Supplement and NP
cell medium (NPCM) were purchased from ScienCell Research
Laboratories, Inc. (Carlsbad, CA, USA), and Dulbecco's modified
Eagle's medium (DMEM)/F-12 and fetal bovine serum (FBS) were
obtained from HyClone; Thermo Fisher Scientific, Inc. (Waltham, MA,
USA). Trypsin, Cell Counting kit-8 (CCK-8) and 17β-E2 were
purchased from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). PBS
(phosphate buffered saline) was obtained from Gibco; Thermo Fisher
Scientific, Inc. The Annexin V-Fluorescein Isothiocyanate
(FITC)/Propidium Iodide (PI) kit was purchased from BD Biosciences
(Franklin Lakes, NJ, USA), and the caspase-3 activity kit, Hoechst
staining kit and cell lysis buffer for western and
immunoprecipitation were obtained from Beyotime Institute of
Biotechnology (Shanghai, China). Z-DEVD-FMK was purchased from
MedChem Express (Monmouth Junction, NJ, USA), ICI 182,780 was
obtained from Sigma-Aldrich, Merck KGaA; and TNF-α was purchased
from PeproTech, Inc. (Rocky Hill, NJ, USA).
Initiation of human NP cell
culture
The purchased cryopreserved human NP cells were
originally isolated from the NP of human IVD. NPCM (500 ml) was
supplemented with 10 ml FBS, 5 ml NP Cell Growth Supplement and 5
ml penicillin/streptomycin solution (P/S) to make complete human NP
cell medium prior to cell recovery. Subsequently, cells were thawed
in their original vials in a 37°C water bath and placed in a 50-ml
culture flask containing 15 ml complete HNPC as soon as possible
and with minimal handling. Cells were cultured in a humidified
atmosphere of 5% CO2 at 37°C. The culture medium was
replaced with fresh the next day to remove residual dimethyl
sulfoxide and non-adherent cells. The medium was replaced every
three days thereafter. Once the culture reached 70% confluence, the
medium was replaced every other day until the culture was ~90%
confluent.
Subculture and drug treatment
When the culture reached ≥90% confluence, the cells
were digested, plated into appropriate culture plates and cultured
as described in the previous section. When the culture reached
80–90% confluence, cells were washed twice with PBS and the medium
was replaced with DMEM/F-12 without phenol red, FBS and P/S. Cells
were divided into 7 groups according to treatment, the conditions
of which are listed in Table I.
All groups were incubated for 48 h in serum-free medium without
phenol red.
 | Table I.Experimental groups. |
Table I.
Experimental groups.
| Group | Treatment |
|---|
| Control | Ethanol
(<0.1%) |
| TNF | 100 ng/ml TNF for
48 h |
| TNF + 0.1 E2 | 100 ng/ml TNF for
48 h, with pretreatment of 0.1 µmol/l E2 for 30 min |
| TNF + 1 E2 | 100 ng/ml TNF for
48 h, with pretreatment of 1 µmol/l E2 for 30 min |
| TNF + 10 E2 | 100 ng/ml TNF for
48 h, with pretreatment of 10 µmol/l E2 for 30 min |
| TNF + 10 E2 +
ICI | 100 ng/ml TNF for
48 h, with pretreatment of 10 µmol/l E2 and 10 µmol/l ICI for 30
min |
| TNF + FMK | 100 ng/ml TNF for
48 h, with pretreatment of 20 µmol/l FMK for 2 h |
Morphological observations
Human NP cells were plated in 6-well plates at a
density of 2×105 cells/well in NPCM. A total of 48 h
after treatment, the apoptotic morphological alterations in the
human NP cells were observed under an inverted phase-contrast
microscope (Olympus Corporation, Tokyo, Japan) and imaged using a
digital camera (Nikon Corporation, Tokyo, Japan). Cells were
subsequently washed twice with PBS and stained with Hoechst 33258
staining solution according to the manufacturer's protocol. Stained
nuclei were detected under a fluorescence microscope (Olympus
Corporation) with an excitation wavelength of 350 nm and an
emission wavelength of 460 nm.
Proliferation assay
The effects of 17β-E2 on cell viability and
proliferation were evaluated using CCK-8. Human NP cells were
seeded into 96-well plates at a density of 2×104
cells/well (100 µl/well). Following treatment for 48 h, 10 µl CCK-8
solution was added to each well of the plate, and the plates were
incubated for an additional 3 h in a 5% CO2 incubator at
37°C. The proliferation of cells was measured using a microplate
reader at a wavelength of 450 nm.
Caspase-3 activity assay
Caspase-3 activity was measured using a Caspase-3
Activity kit, which measures the conversion by caspase-3 of
N-acetyl-Asp-Glu-Val-Asp-p-nitroanilide into a yellow formazan
product, p-nitroaniline. Human NP cells were cultured in 6-well
plates as aforementioned, and the assay was performed according to
the manufacturer's protocol. Briefly, the treated NP cells were
washed twice with cold PBS and lysed with lysis buffer (100
µl/2×106 cells) for 15 min on ice. Following incubation
of 10 µl cell lysates, 80 µl reaction buffer and 10 µl caspase-3
substrate (2 mM) in 96-well microtiter plates at 37°C for 2 h,
caspase-3 activity was quantified at a wavelength of 405 nm using a
microplate spectrophotometer (BioTek Instruments, Inc., Winooski,
VT, USA). Caspase-3 activity is expressed as the fold change in
enzyme activity over control.
Flow cytometric analysis of
apoptosis
Human NP cells were seeded into 6-well plates at a
density of 2×105 cells/well and cultured as
aforementioned. Following 48 h treatment, adherent cells and cells
in suspension were collected and washed twice with cold PBS.
Apoptotic cells were detected by staining with annexin V-FITC and
PI according to the manufacturer's protocol. Briefly, cells were
resuspended in 100 µl 1X binding buffer at a concentration of
1×106 cells/ml, and 100 µl (1×105 cells) were
transferred to 5 ml culture tubes. PI (5 µl) and 5 µl annexin
V-FITC was added to each tube and incubated for 15 min at room
temperature in the dark, followed by addition of 400 ml 1X binding
buffer. Samples were analyzed on a flow cytometer (BD Biosciences)
with CellQuest software (version 5.2, BD Biosciences).
Western blot analysis
Protein expression levels of PARP and caspase-3 were
determined by western blot analysis. Cells were incubated in 100 µl
lysis buffer containing 1% protease inhibitors. The cell lysate was
collected by centrifugation at 4°C for 10 min at 16,000 × g. The
protein concentrations were determined using a Bicinchoninic Acid
Protein assay kit (Beyotime Institute of Biotechnology). Proteins
(50 µg) from each sample were separated on 12% SDS-polyacrylamide
gels according to molecular weight and transferred onto
polyvinylidene difluoride membranes (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The membranes were blocked with 5% non-fat milk
powder in TBS for 1 h at room temperature and incubated with
primary antibodies against caspase-3 p17 (#9664; rabbit; 1:1,000),
PARP (#9532; rabbit; 1:1,000) (both from Cell Signaling Technology,
Inc., Danvers, MA, USA), and GAPDH (10494–1-AP; rabbit; 1:5,000;
Wuhan Sanying Biotechnology, Wuhan, China) diluted in 5% non-fat
milk powder in PBS containing Tween 20 (PBST) at 4°C overnight.
Membranes were washed 3 times for 15 min in PBST and incubated with
a peroxidase labeled anti-rabbit IgG (H+L) antibody (074–1506;
goat; 1:1,000; KPL, Inc., Gaithersburg, MD, USA) with agitation for
1 h at room temperature. Following washing 3 times for 15 min with
PBST, protein bads were detected using Enhanced Chemiluminescence
reagent (Amercontrol Biosciences, San Francisco, CA, USA) and
quantified with Quantity One imaging software version 4.6.2
(Bio-Rad Laboratories, Inc.).
Statistical analysis
Statistical analyses were performed using SPSS
software version 16.0 (SPSS, Inc., Chicago, IL, USA). Data are
presented as the mean ± standard deviation. Groups were compared by
one-way analysis of variance followed by pairwise comparison using
the Student-Newman-Keuls post hoc test. All statistical tests were
two-sided. P<0.01 was considered to indicate a statistically
significant difference.
Results
Morphology of apoptotic human NP
cells
The morphological characteristics of human NP cells
were observed using inverted phase-contrast microscopy (Fig. 1). Cell nuclei were stained using
Hoechst 33258 and detected under a fluorescence microscope with an
excitation wavelength of 350 nm and an emission wavelength of 460
nm (Fig. 2). Control untreated
cells were well-adhered, exhibiting typical human NP cell
morphology. By contrast, treatment with TNF-α induced apoptosis, as
demonstrated by detachment from the plate, cell shrinkage, and
nuclei that were compact, condensed and brightly stained with
Hoechst 33258. Following pretreatment with various concentrations
of 17β-E2, the number of apoptotic cells markedly decreased
compared with those treated with TNF-α only. The effect of 17β-E2
was blocked by the estrogen receptor (ER) antagonist ICI
182,780.
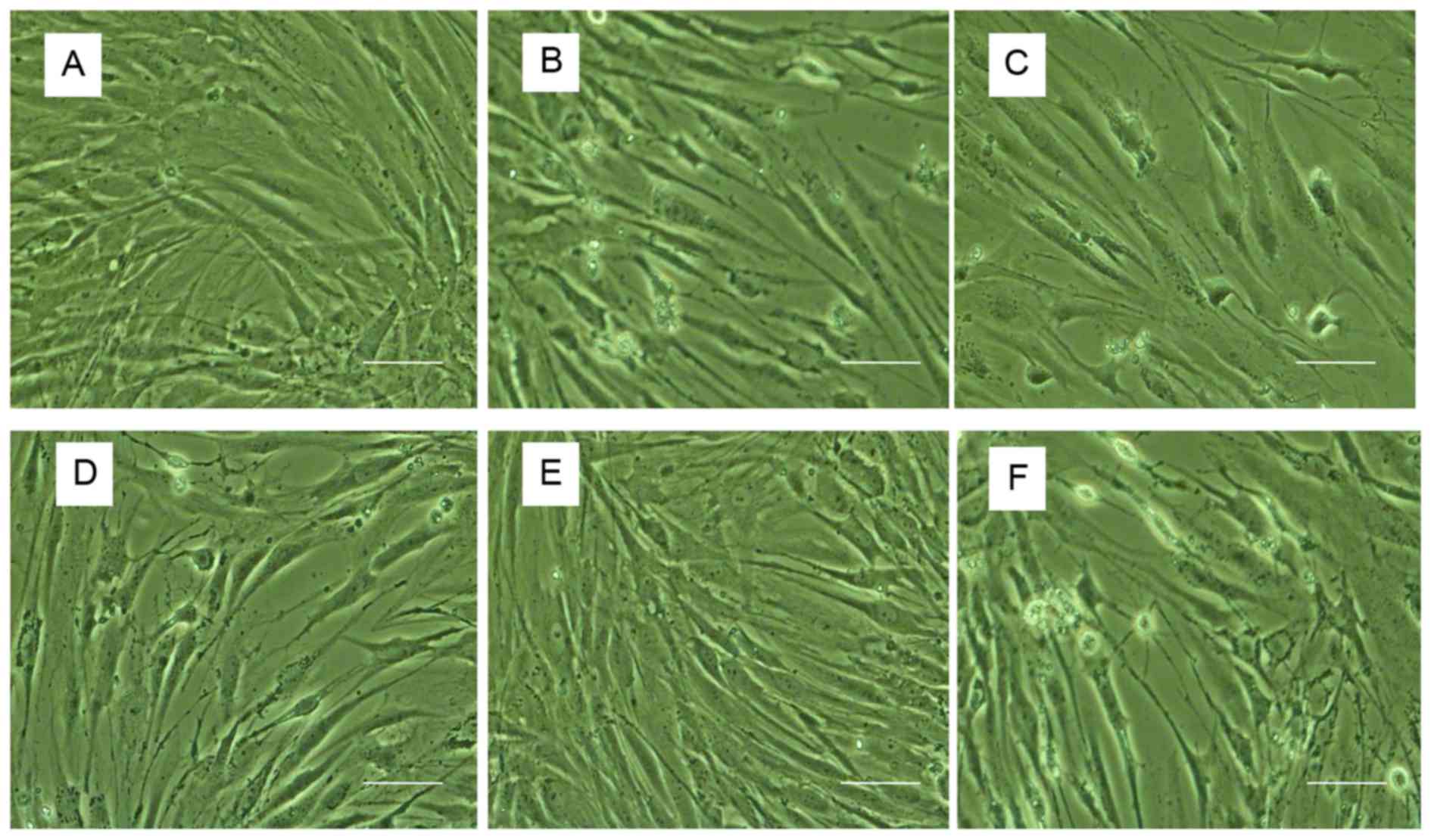 | Figure 1.Morphological observations by
phase-contrast microscopy. (A) Control untreated human NP cells.
Human NP cells treated with (B) 100 ng/ml TNF-α, (C) 100 ng/ml
TNF-α following pretreatment with 0.1 µmol/l 17β-E2, (D) 100 ng/ml
TNF-α following pretreatment with 1 µmol/l 17β-E2, (E) 100 ng/ml
TNF-α following pretreatment with 10 µmol/l 17β-E2 and (F) 100
ng/ml TNF-α following pretreatment with 10 µmol/l 17β-E2 and 10
µmol/l ICI 182,780. Apoptotic cells were characterized by shedding
of smaller fragments from the cells, cell shrinkage and condensed
nuclei. Original magnification, ×200; scale bar, 200 nm. NP,
nucleus pulposus; TNF-α, tumor necrosis factor-α; 17β-E2,
17β-estradiol. |
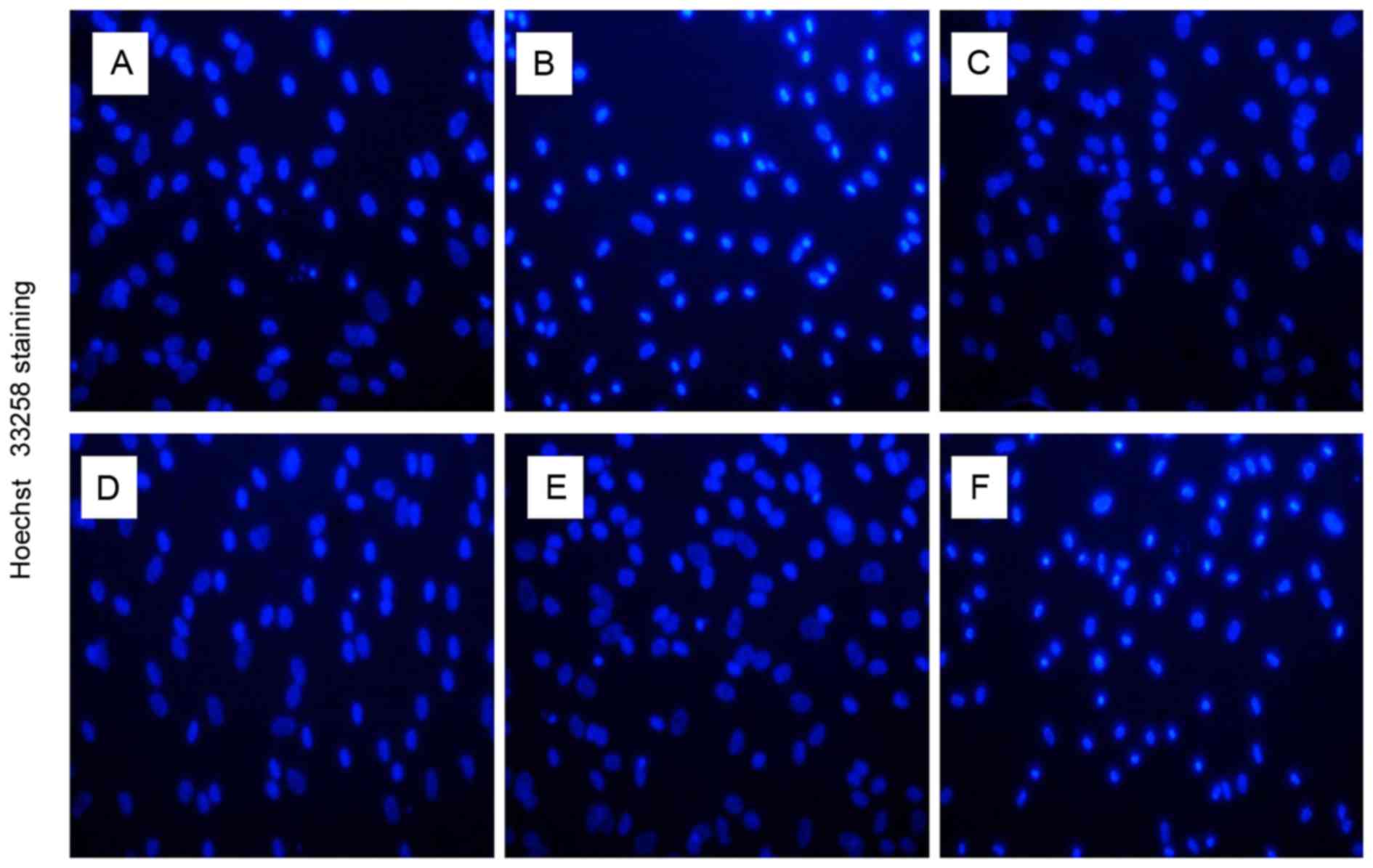 | Figure 2.Hoechst 33258 staining. (A) Control
untreated human NP cells. Human NP cells treated with (B) 100 ng/ml
TNF-α, (C) 100 ng/ml TNF-α following pretreatment with 0.1 µmol/l
17β-E2, (D) 100 ng/ml TNF-α following pretreatment with 1 µmol/l
17β-E2, (E) 100 ng/ml TNF-α following pretreatment with 10 µmol/l
17β-E2 and (F) 100 ng/ml TNF-α following pretreatment with 10
µmol/l 17β-E2 and 10 µmol/l ICI 182,780. The nuclei of living cells
were a homogeneous blue, whereas apoptotic nuclei were compact,
condensed and brightly stained with Hoechst 33258. Original
magnification, ×200. NP, nucleus pulposus; TNF-α, tumor necrosis
factor-α; 17β-E2, 17β-estradiol. |
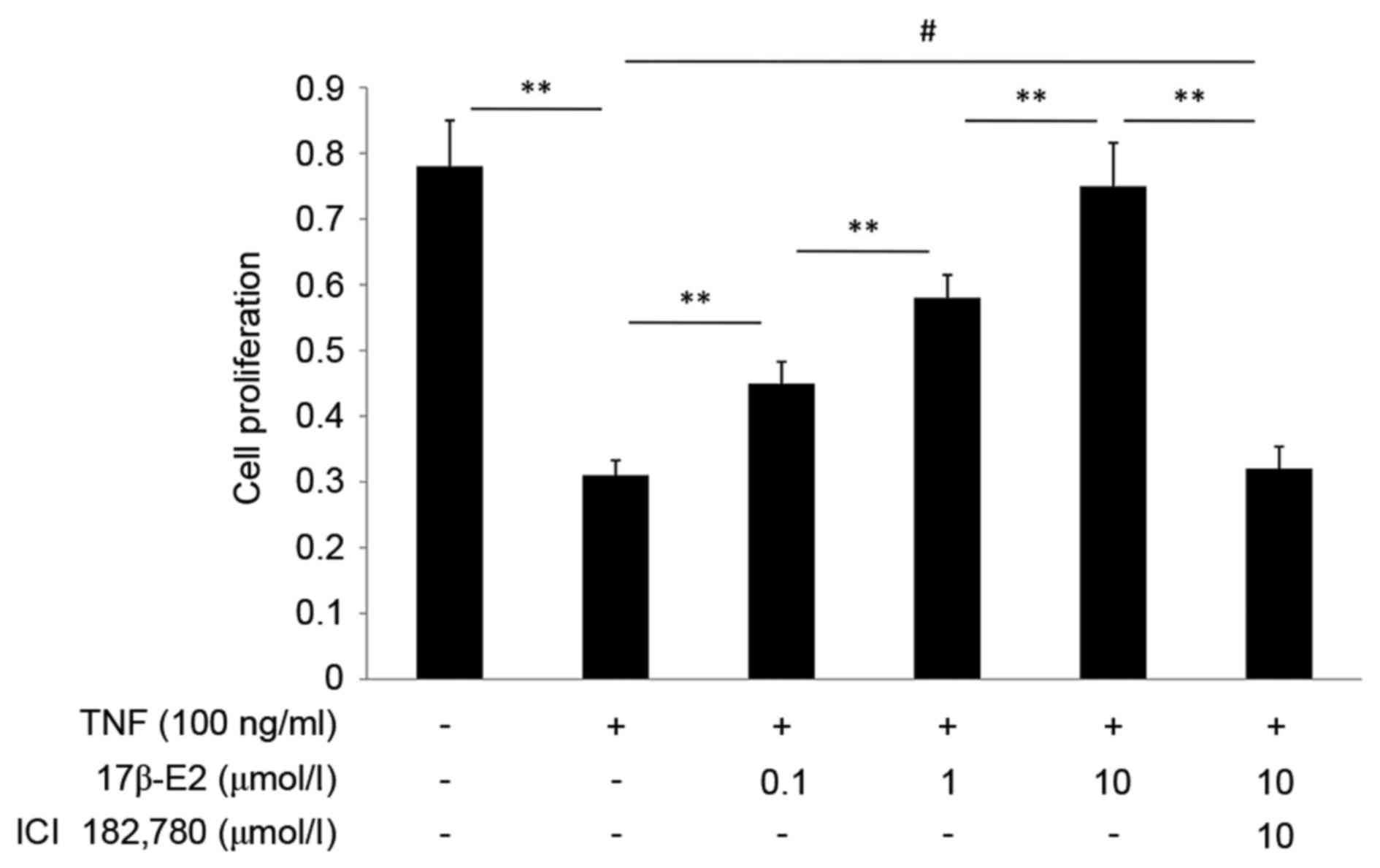
17β-E2 promotes proliferation of human
NP cells damaged by TNF-α
The effect of 17β-E2 on proliferation of human NP
cells was detected by CCK-8 assay. As presented in Fig. 3, TNF-α inhibited the proliferation
of human NP cells (P<0.001). This effect was abrogated when the
cells were pretreated with 17β-E2 (P<0.001). The effect of
17β-E2 was blocked by ICI 182,780 (P<0.001). 17β-E2 inhibited
the effect of TNF-α in a concentration-dependent manner.
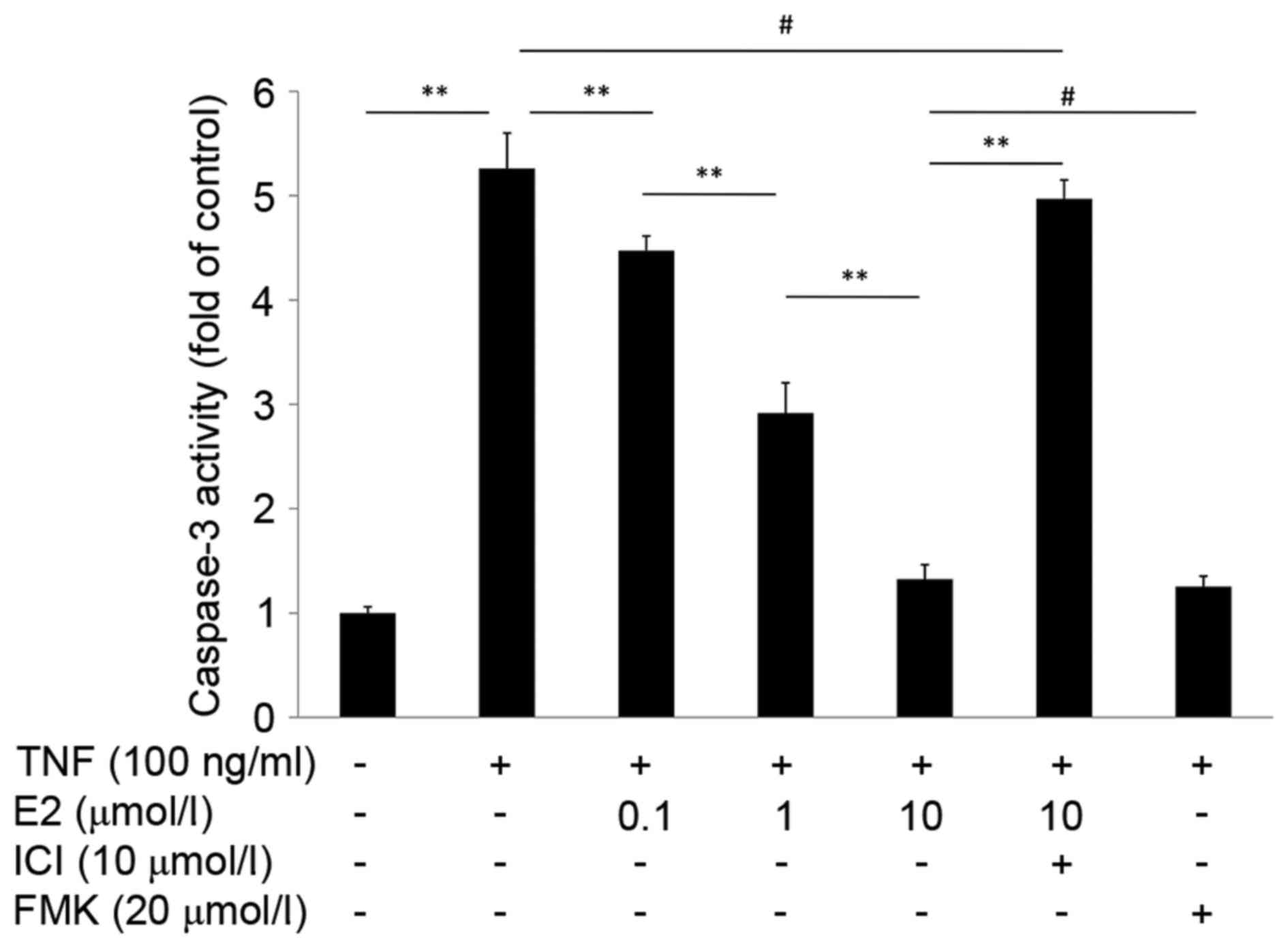
17β-E2 reduces caspase-3 activity
The effect of 17β-E2 on caspase-3 activity in human
NP cells was measured using a caspase-3 activity assay. As
presented in Fig. 4, the caspase-3
activity of cells treated with TNF-α alone was 5-fold greater
compared with the control untreated group (P<0.001). However,
cells pretreated with 17β-E2 demonstrated reduced caspase-3
activity compared with cells treated with TNF-α only; the effect of
17β-E2 was concentration-dependent. The effect of pretreatment with
10 µmol/l 17β-E2 for 30 min was similar to pretreatment with 20
µmol/l Z-DEVD-FMK (a specific, irreversible caspase-3 inhibitor)
for 2 h (P>0.05). Addition of the ER antagonist ICI 182,780
abolished the protective effects of 17β-E2 (P<0.001).
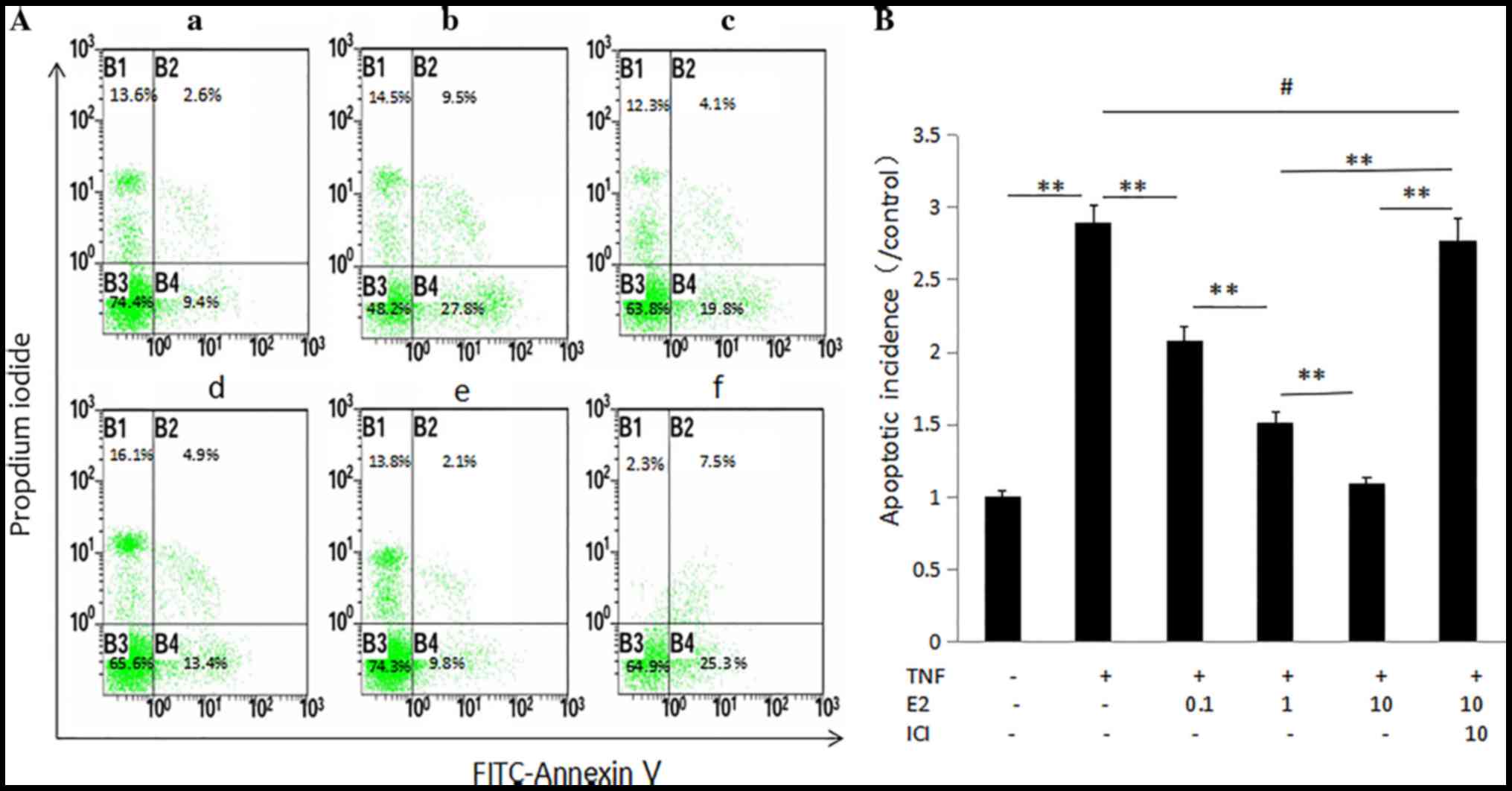
17β-E2 reduces TNF-α-induced human NP
cell apoptosis
The effect of 17β-E2 on apoptosis of human NP cells
was examined by flow cytometry following annexin V-FITC and PI
double staining. The apoptotic rate is presented as apoptotic cells
in the experimental group/apoptotic cells in the control group. As
presented in Fig. 5, an increase
in the apoptotic rate was observed following treatment with TNF-α.
Pretreatment with 17β-E2 significantly decreased the rate of
apoptosis in a concentration-dependent manner compared with cells
treated with TNF-α only (P<0.001). Addition of the ER antagonist
ICI 182,780 blocked the effect of 17β-E2 on TNF-α treatment,
further demonstrating the potential contribution of 17β-E2 to
TNF-α-induced human NP cell apoptosis.
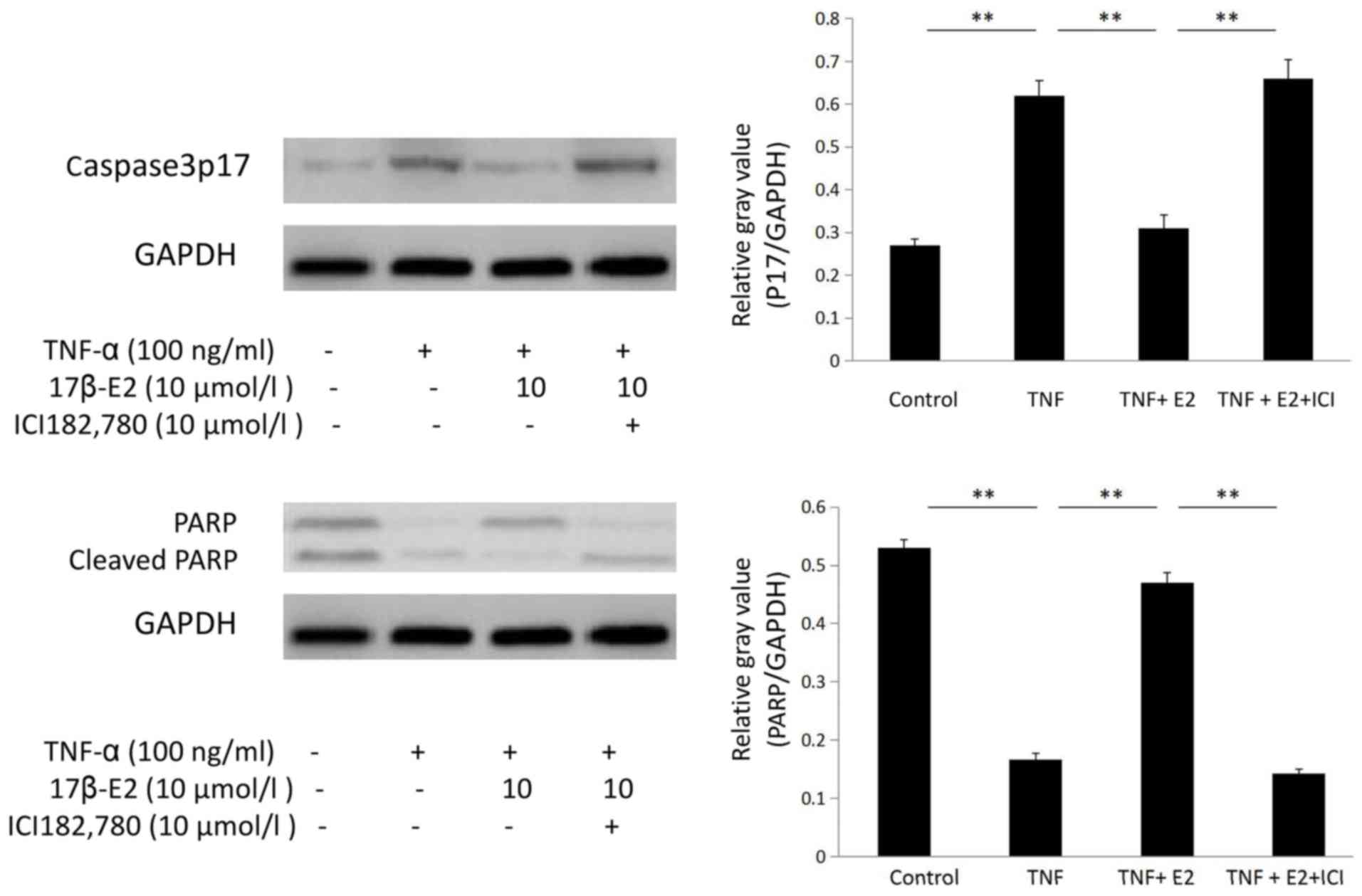
Western blot analysis
Human NP cells were divided into the following four
groups: Control untreated, 100 ng/ml TNF-α, 100 ng/ml TNF-α
following pretreatment with 10 µmol/l 17β-E2 for 30 min and 100
ng/ml TNF-α following pretreatment with 10 µmol/l 17β-E2 and 10
µmol/l ICI 182,780 for 30 min. Following treatment, protein
expression levels of PARP, caspase-3p17 and GAPDH were determined
by western blot analysis. Caspase-3p17 protein expression levels
were increased in human NP cells following treatment with TNF-α
compared with the control cells (P<0.001); this increased was
inhibited by pretreatment with 17β-E2 (P<0.001). By contrast,
PARP protein expression levels were decreased in cells subjected to
TNF-α treatment (P<0.001); this decrease was abrogated by
pretreatment with 17β-E2 (P<0.001). These effects of 17β-E2 were
blocked by ICI 182,780 treatment (P<0.001; Fig. 6).
Discussion
One of the primary causes of spinal degeneration is
IVD degeneration. Apoptosis of NP cells and the loss of ECM are the
central features of the aged and degenerated IVD (7,8,27,28).
Gruber et al (29) reported
that the ER gene is expressed in human IVD cells in vivo and
in vitro, and further demonstrated that 17β-E2 significantly
increased proliferation of annulus cells. Previous studies have
revealed that TNF-α may induce NP cell apoptosis (16,30),
and various in vitro studies have demonstrated that 17β-E2
protects against apoptosis in IVD cells (6,22,24).
Therefore, the hypothesis of the present study was that 17β-E2
would attenuate TNF-α-induced apoptosis in human NP cells.
The results of the present study suggested that
17β-E2 abolished TNF-α-induced apoptosis in human NP cells by
increasing proliferation and reducing apoptosis. A CCK-8 assay
revealed that 17β-E2 enhanced the proliferative capacity of human
NP cells. Morphological observations and flow cytometric analysis
demonstrated that TNF-α-induced apoptosis in human NP cells was
prevented by 17β-E2. Western blot analysis was used to detect
alterations in the protein expression levels of PARP and caspase-3
in human NP cells. Caspase-3 is a crucial executioner protease,
which activates a caspase-activated DNase and serves an important
role in apoptosis (27,31–33).
Previous studies have reported that caspase-3 may be a therapeutic
target for regulation of IVD degeneration (34). Pro-caspase-3 may be sheared into
two fragments, p17 and p10, upon activation. The protein expression
levels of caspase-3 p17 are positively correlated with caspase-3
activity. PARP is a substrate of caspase-3, which may trigger
inflammation and cell death (35).
Shakibaei et al (36)
demonstrated that the cleavage of PARP was activated in
vitro in apoptotic human articular chondrocytes.
The present study investigated whether 17β-E2
inhibited human NP cell apoptosis induced by TNF-α. Estrogen has
positive effects on bone structure, chondrocyte and nerve cells.
Various recent studies have revealed the anti-apoptotic effect of
17β-E2 in rat IVD cells (6,22,37).
However, whether 17β-E2 may prevent TNF-α-induced apoptosis in
human NP cells remains to be investigated. In the present study,
human NP cells were used to provide an environment approaching the
physiological state for the in vitro test of the effect of
17β-E2 on human NP. Phenol red may increase cell proliferation
(38); therefore, medium without
phenol red was used in the present study.
IVD degeneration is demonstrated by alterations in
cell proliferation and cell matrix metabolism, which includes
inhibition of nuclear proteoglycan synthesis and enhanced matrix
degradation. IVD cells have been reported to produce inflammatory
cytokines, including IL-1β, IL-6, TNF-α, matrix metalloproteinases
(MMPs), nitric oxide and prostaglandin E2 (39). As an important inflammatory
molecule, TNF-α may induce the cellular and matrix alterations of
IVD degeneration (30), and
activate crosstalk between signaling pathways including nuclear
factor-κB and phosphoinositide 3-kinase (PI3K)/protein kinase B
(Akt) (40,41).
The mechanism underlying the protection of 17β-E2
against TNF-α-induced apoptosis in human NP cells is complex.
Various potential mechanisms may be involved in the anti-apoptotic
effect of 17β-E2 in human NP cells. Firstly, integrin production
was induced in NP cells by 17β-E2. It has been reported that 17β-E2
may protect rat IVD cells by upregulating integrin α and β
(22,42). Secondly, 17β-E2 may inhibit human
NP cell apoptosis via the mitochondrial pathway. Yang et al
(43) demonstrated that 17β-E2
protects against apoptosis by downregulating MMP-3 and −13 via a
mitochondrial pathway in rat NP cells. Finally, the PI3K/Akt
signaling pathway may be involved in the effect of 17β-E2. A
further study by Yang et al (37) revealed that 17β-E2 protects against
apoptosis via the activation of PI3K/Akt signaling pathway. This
signaling pathway is associated with cell migration and invasion
(44,45).
The present study investigated the effect of 17β-E2
on human NP cells; however, the findings of the present study are
limited. The results of the present study were used as preliminary
data for our subsequent study, which demonstrated that 17β-E2
inhibits TNF-α-induced apoptosis in human NP cells via the PI3K/Akt
pathway (46). Further signaling
mechanisms underlying the anti-apoptotic effects of 17β-E2 in human
NP cells will require further investigation. In addition, 17β-E2 is
a systemic hormone, and therefore further studies are required to
determine the systemic effect of 17β-E2 in humans, to assess the
suitability of 17β-E2 as a therapeutic strategy for the clinical
treatment of IVD degeneration.
In conclusion, the results of the present study
revealed that 17β-E2 protected against TNF-α-induced apoptosis in
human NP cells in a concentration-dependent manner, and that the
activation of caspase-3 and inhibition of PARP may participate in
this process. The ability of 17β-E2 to protect human NP cells from
apoptosis was blocked by ICI 172,780, an ER antagonist. The results
of the present study may present a novel route for the prevention
and therapy of IVD degeneration.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Hebei Province (grant no. H2014206075).
References
|
1
|
Zhao CQ, Jiang LS and Dai LY: Programmed
cell death in intervertebral disc degeneration. Apoptosis.
11:2079–2088. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buckwalter JA: Aging and degeneration
human intervertebral disc. Spine (Phila Pa 1976). 20:1307–1314.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferguson SJ, Ito K and Nolte LP: Fluid
flow and convection transport of solutes within the intervertebral
disc. J Biomech. 37:213–221. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Anderson DG, Li X and Balian G: A
fibronectin fragment alters the metabolism by rabbit intervertebral
disc cells in vitro. Spine (Phila Pa 1976). 30:1242–1246. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Podichetty VK: The aging spine: The role
of inflammatory mediators in intervertebral disc degeneration. Cell
Mol Biol (Noisy-le-grand). 53:4–18. 2007.PubMed/NCBI
|
|
6
|
Wang H, Ding W, Yang D, Gu T, Yang S and
Bai Z: Different concentrations of 17β-estradiol modulates
apoptosis induced by interleukin-1β in rat annulus firosus cells.
Mol Med Rep. 10:2745–2751. 2014.PubMed/NCBI
|
|
7
|
Zhou GQ, Yang F, Leung VVL and Cheung KMC:
Molecular and cellular biology of the intervertebral disc and the
use of animal models. Current Orthopaedics. 22:267–273. 2008.
View Article : Google Scholar
|
|
8
|
Le Maitre CL, Freemont AJ and Hoyland JA:
Accelerated cellular senescence in degenerate intervertebral discs:
A possible role in the pathogenesis of intervertebral disc
degeneration. Arthritis Res Ther. 9:R452007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Boos N, Weissbach S, Rohrbach H, Weiler C,
Spratt KF and Nerlich AG: Classification of age-related changes in
lumbar intervertebral discs: 2002 Volvo Award in basic science.
Spine (Phila Pa 1976). 27:2631–2644. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Szlosarek P, Charles KA and Balkwill FR:
Tumour necrosis factor-alpha as atumour promoter. Eur J Cancer.
42:745–750. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bharadwaj U, Marin-Muller C, Li M, Chen C
and Yao Q: Mesothelin confers pancreatic cancer cell resistance to
TNF-a-induced apoptosis through Akt/PI3K/NF-κB activation and
IL-6/Mcl-1 overexpression. Mol Cancer. 10:1062011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Le Maitre CL, Freemont AJ and Hoyland JA:
The role of interleukin-1 in the pathogenesis of human
Intervertebral disc degeneration. Arthritis Res Ther. 7:R732–R745.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Markova D, Anderson DG, Zheng Z,
Shapiro IM and Risbud MV: TNF-α and IL-1β promote a
disintegrin-like and metalloprotease with thrombospondin type I
motif-5-mediated aggrecan degradation through syndecan-4 in
intervertebral disc. J Biol Chem. 286:39738–39749. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee S, Moon CS, Sul D, Lee J, Bae M, Hong
Y, Lee M, Choi S, Derby R, Kim BJ, et al: Comparison of growth
factor and cytokine expression in patients with degenerated disc
disease and herniated nucleus pulposus. Clin Biochem. 42:1504–1511.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mourão AF, Caetano-Lopes J, Costa P,
Canhão H, Santos MJ, Pinto P, Brito I, Nicola P, Cavaleiro J, Teles
J, et al: Tumor necrosis factor-alpha-308 genotypes influence
inflammatory activity and TNF-alpha serum concentrations in
children with juvenile idiopathic arthritis. J Rheumatol.
36:837–842. 2009.PubMed/NCBI
|
|
16
|
Zhang CC, Cui GP, Hu JG, Xiao YZ, Zhou XS,
Shao C, Lin Q and Zhou JS: Effects of adenoviral vector expressing
hIGF-1 on apoptosis in nucleus pulposus cells in vitro. Int J Mol
Med. 33:401–405. 2014.PubMed/NCBI
|
|
17
|
de Souza Grava AL, Ferrari LF and Defino
HL: Cytokine inhibition and time-related influence of inflammatory
stimuli on the hyperalgesia induced by the nucleus pulposus. Eur
Spine J. 21:537–545. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Le Maitre CL, Hoyland JA and Freemont AJ:
Catabolic cytokine expression in degenerate and herniated human
intervertebral discs: IL-1beta and TNFalpha expression profile.
Arthritis Res Ther. 9:R772007. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ackermann S, Hiller S, Osswald H, Lösle M,
Grenz A and Hambrock A: 17beta-Estradiol modulates apoptosis in
pancreatic beta-cells by specific involvement of the sulfonylurea
receptor (SUR) isoform SUR1. J Biol Chem. 284:4905–4913. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamaguchi A, Nozawa K, Fujishiro M,
Kawasaki M, Takamori K, Ogawa H, Sekigawa I and Takasaki Y:
Estrogen inhibits apoptosis and promotes CC motif chemokine ligand
13 expression on synovial fibroblasts in rheumatoid arthritis.
Immunopharmacol Immunotoxicol. 34:852–857. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hattori Y, Kojima T, Kato D, Matsubara H,
Takigawa M and Ishiguro N: A selective estrogen receptor modulator
inhibits tumor necrosis factor-α-induced apoptosis through the
ERK1/2 signaling pathway in human chondrocytes. Biochem Biophys Res
Commun. 421:418–424. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang SD, Ma L, Gu TX, Ding WY, Zhang F,
Shen Y, Zhang YZ, Yang DL, Zhang D, Sun YP and Song YL:
17β-Estradiol protects against apoptosis induced by levofloxacin in
rat nucleus pulposus cells by upregulating integrin α2β1.
Apoptosis. 19:789–800. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bozzo C, Graziola F, Chiocchetti A and
Canonico PL: Estrogen and beta-amyloid toxicity: Role of integrin
and PI3-K. Mol Cell Neurosci. 45:85–91. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nilsen J and Brinton R Diaz: Mechanism of
estrogen-mediated neuroprotection: Regulation of mitochondrial
calcium and Bcl-2 expression. Proc Natl Acad Sci USA.
100:2842–2847. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu X, Zhang X, Dhakal IB, Beggs M,
Kadlubar S and Luo D: Induction of cell proliferation and survival
genes by estradiol-repressed microRNAs in breast cancer cells. BMC
Cancer. 12:292012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ge C, Yu M and Zhang C: G Protein-Coupled
Receptor 30 mediates estrogen-induced proliferation of primordial
germ cells via EGFR/Akt/β-catenin signaling pathway. Endocrinology.
153:3504–3516. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ding F, Shao ZW, Yang SH, Wu Q, Gao F and
Xiong LM: Role of mitochondrial pathway in compression-induced
apoptosis of nucleus pulposus cells. Apoptosis. 17:579–90. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nardinocchi L, Puca R, Sacchi A and
D'Orazi G: Inhibition of HIF-1alpha activity by
homeodomain-interacting protein kinase-2 correlates with
sensitization of chemoresistant cells to undergo apoptosis. Mol
Cancer. 8:12009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gruber HE, Yamaguchi D, Ingram J, Leslie
K, Huang W, Miller TA and Hanley EN Jr: Expression and localization
of estrogen receptor-beta in annulus cells of the human
intervertebral disc and the mitogenic effect of 17-beta-estradiol
in vitro. BMC Musculoskelet Disord. 3:42002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Weber A, Wandinger KP, Mueller W, Aktas O,
Wengert O, Grundström E, Ehrlich S, Windemuth C, Kuhlmann T,
Wienker T, et al: Identification and functional characterization of
a highly polymorphic region in the human TRAIL promoter in multiple
sclerosis. J Neuroimmunol. 149:195–201. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Singh M, Sharma H and Singh N: Hydrogen
peroxide induces apoptosis in HeLa cells through mitochondrial
pathway. Mitochondrion. 7:367–373. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He B, Tao H, Liu S and Wei A: Protective
effect of carboxymethylated chitosan on hydrogen peroxide-induced
apoptosis in nucleus pulposus cells. Mol Med Rep. 11:1629–1638.
2015.PubMed/NCBI
|
|
33
|
Li Z, Jo J, Jia JM, Lo SC, Whitcomb DJ,
Jiao S, Cho K and Sheng M: Caspase-3 activation via mitochondria is
required for long-term depression and AMPA receptor
internalization. Cell. 141:859–871. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sudo H and Minami A: Caspase 3 as a
therapeutic target for regulation of intervertebral disc
degeneration in rabbits. Arthritis Rheum. 63:1648–1657. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao H, Yenari MA, Cheng D, Sapolsky RM
and Steinberg GK: Bcl-2 overexpression protects against neuron loss
within the ischemic margin following experimental stroke and
inhibits cytochrome c translocation and caspase-3 activity. J
Neurochem. 85:1026–1036. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shakibaei M, John T, Seifarth C and
Mobasheri A: Resveratrol inhibits IL-1 beta-induced stimulation of
caspase-3 and cleavage of PARP in human articular chondrocytes in
vitro. Ann N Y Acad Sci. 1095:554–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang SD, Ma L, Yang DL and Ding WY:
Combined effect of 17β-estradiol and resveratrol against apoptosis
induced by interleukin-1β in rat nucleus pulposus cells via
PI3K/Akt/caspase-3 pathway. PeerJ. 4:e16402016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wesierska-Gadek J, Schreiner T,
Gueorguieva M and Ranftler C: Phenol red reduces ROSC mediated cell
cycle arrest and apoptosis in human MCF-7 cells. J Cell Biochem.
98:1367–1379. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Weiler C, Nerlich AG, Bachmeier BE and
Boos N: Expression and distribution of tumor necrosis factor alpha
in human lumbar intervertebral discs: A study in surgical specimen
and autopsy controls. Spine (Phila Pa 1976). 30:44–54. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu Y and Zhou BP:
TNF-alpha/NF-kappaB/Snail pathway in cancer cell migration and
invasion. Br J Cancer. 102:639–644. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sandra F, Matsuki NA, Takeuchi H, Ikebe T,
Kanematsu T, Ohishi M and Hirata M: TNF inhibited the apoptosis by
activation of Akt serine/threonine kinase in the human head and
neck squamous cell carcinoma. Cell Signal. 14:771–778. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao CM, Chen Q, Zhang WJ, Huang AB, Zhang
W, Yang HL and Zhang ZM: 17β-estradiol protects rat annulus
fibrosus cells against apoptosis via α1 integrin-mediated adhesion
to type I collagen: An in-vitro study. Med Sci Monit. 22:1375–1383.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang SD, Yang DL, Sun YP, Wang BL, Ma L,
Feng SQ and Ding WY: 17β-estradiol protects against apoptosis
induced by interleukin-1β in rat nucleus pulposus cells by
down-regulating MMP-3 and MMP-13. Apoptosis. 20:348–357. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Aggarwal BB, Shishodia S, Sandur SK,
Pandey MK and Sethi G: Inflammation and cancer: How hot is the
link? Biochem Pharmacol. 72:1605–1621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Schmiegel W, Roeder C, Schmielau J, Rodeck
U and Kalthoff H: Tumor necrosis factor alpha induces the
expression of transforming growth factor alpha and the epidermal
growth factor receptor in human pancreatic cancer cells. Proc Natl
Acad Sci USA. 90:863–867. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang T, Yang SD, Liu S, Wang H, Liu H and
Ding WY: 17β-estradiol inhibites tumor necrosis factor-a induced
apoptosis of human nucleus pulposus cells via the PI3K/Akt pathway.
Med Sci Monit. 22:4312–4322. 2016. View Article : Google Scholar : PubMed/NCBI
|