Introduction
Pulmonary fibrosis occurs in various clinical
conditions, including chronic obstructive pulmonary disease,
asthma, idiopathic pulmonary fibrosis, interstitial pneumonia,
radiation-induced lung injury and end-stage lung failure (1–3).
Pulmonary fibrosis is characterized by destruction of the lung
architecture, and the production and excessive accumulation of a
collagen-rich extracellular matrix (ECM), eventually leading to
respiratory insufficiency. The primary etiology of fibrosis remains
poorly understood, and there is a lack of therapeutic interventions
and novel therapies for pulmonary fibrosis (4,5).
Previous studies have demonstrated that fibroblast
proliferation and activation serve a role in the pathogenic
fibrotic process. When stimulated by transforming growth factor
(TGF)-β1 or other local cytokines, fibroblasts proliferate,
migrate, and differentiate into myofibroblasts, which drive lung
fibrogenesis (6). Fibroblasts and
myofibroblasts produce numerous cytokines and growth factors in
addition to the ECM, which are the hallmarks of fibrotic pulmonary
disease (7). A previous study
demonstrated that dysregulation of fibroblast activation and
differentiation into myofibroblasts may be a therapeutic strategy
for pulmonary fibrosis (8).
A family of endogenous specialized pro-resolving
mediators, including lipoxins, resolvins, protectins and maresins,
serve a role in the regulation of tissue inflammation, organ
protection and fibrotic diseases (9,10).
Maresin 1 is biosynthesized via lipoxygenase by docosahexaenoic
acid to generate 14S-hydroperoxydocosa-4Z, 7Z, 10Z, 12E, 16Z,
19Z-hexaenoic acid, which undergoes further conversion via
epoxidation in macrophages and is subsequently converted to 7R,
14S-dihydroxydocosa-4Z, 8Z, 10E, 12Z, 16Z, 19Z-hexaenoic acid
(Fig. 1A), known as maresin 1 (MaR
1) (11). Previous studies in
vitro and in vivo demonstrated that MaR 1 exhibited
homeostatic activity in models, including colitis, tissue
generation, liver injury, lung injury and intimal hyperplasia
(12–16). A recent study reported that MaR 1
ameliorated bleomycin-induced pulmonary fibrosis in mice and
suppressed epithelial-mesenchymal transition in the mouse type II
alveolar epithelial cell line MLE-12 (17). Due to the important role of
fibroblasts in lung fibrogenesis, it was hypothesized that MaR 1
may directly influence lung fibroblast activation.
The aim of the present study was to evaluate the
impact of MaR 1 on TGF-β1-induced human lung fibroblast (MRC-5)
proliferation, migration and differentiation in vitro. In
order to obtain an increased understanding of the underlying
mechanisms, the effects of MaR 1 on mothers against decapentaplegic
homolog 2/3 (Smad2/3) and extracellular signal-regulated kinase
(ERK) 1/2 phosphorylation signaling pathways were investigated in
TGF-β1-treated MRC5 cells.
Materials and methods
Cell culture and stimulation
MRC5 lung fibroblasts (human fetal lung fibroblasts;
American Type Culture Collection, Manassas, VA, USA) were grown in
Dulbecco's modified Eagle medium (Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) with 4.5 g/l glucose, 10% fetal bovine
serum (Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1%
penicillin/streptomycin solution. Cells were maintained at 37°C in
a humidified incubator, in the presence of 5% CO2.
Following incubation for 3 days, the cells were cultured to ~80%
confluence.
MRC5 cells were pretreated with MaR 1 (Cayman
Chemical Company, Ann Arbor, MI, USA) at 1, 10 or 100 nM, or
vehicle (0.035% ethanol) for 30 min. Subsequently, TGF-β1 (10
ng/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added and
co-incubated for 24 h.
Cell viability assay
Cell viability was assessed using an MTT assay
according to the manufacturer's protocol (Cayman Chemical Company).
MRC5 cells (2.5×103/100 µl) were seeded into the wells of 96-well
culture plates and treated as described above. A total of 24 h
subsequent to treatment, cells were incubated with 50 µg MTT
solution for 4 h. The supernatants were removed and dimethyl
sulfoxide was added to each well. The quantity of aqueous soluble
formazan, which was produced by viable cells from tetrazolium salt,
was measured using spectrophotometry at absorbance (A) 570 nm, and
was equal to the number of living cells. Optical density
A570 was measured in six samples of each group.
Cell migration
In order to evaluate the migratory response of MaR 1
pretreated cells following TGF-β1 exposure, a scratch wound healing
assay was performed. Cells were seeded at confluent status for 24 h
and pre-incubated with MaR 1 (10 nM) for 30 min prior to the
addition of TGF-β1 (10 ng/ml) as a chemotactic factor. A cell-free
area (900-µm scratch wound) in each well was created using a 200-µl
pipette tip. A total of 24 h subsequently, the migratory cells in
the gap were counted using a light microscope at ×200
magnification. All of the data were obtained in six independent
experiments.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Cells were pretreated with or without MaR 1 (10 nM)
for 30 min, followed by the addition of TGF-β1 (10 ng/ml) for 24 h.
Total cellular RNA was extracted using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), and 2 µg RNA was reverse
transcribed into cDNA using the RevertAid™ First Strand
cDNA Synthesis kit (Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. The specific primer sequences
(Invitrogen; Thermo Fisher Scientific, Inc.) used were as follows
(5′-3′): α-SMA forward, GAC AAT GGC TCT GGG CTC TGT AA and reverse,
ATG CCA TGT TCT ATC GGG TAC TT; collagen α-1(I) chain (COL1A1)
forward, GAG GGC CAA GAC GAA GAC ATC and reverse, CAG ATC ACG TCA
TCG CAC AAC; and GAPDH forward, AGT GCC AGC CTC GTC TCA TAG and
reverse, CGT TGA ACT TGC CGT GGG TAG. Each specific gene product
was amplified by real-time PCR using SYBR-Green Master Mix in the
StepOne™ Real-Time PCR system (Thermo Fisher Scientific, Inc.). PCR
thermal cycling was conducted at 95°C for 15 sec, followed by
denaturing at 95°C for 5 sec, annealing at 60°C for 60 sec and a
final extension step at 72°C for 10 sec, for a total of 40 cycles.
The reaction was duplicated for each sample. The relative
expression levels of each transcript were normalized to GAPDH
expression. The quantitative fold changes in gene expression were
calculated using the 2−ΔΔCq (18) method following normalization to
GAPDH.
Immunofluorescence assay
Cells were fixed with 4% paraformaldehyde for 30 min
and permeabilized using 0.5% Triton X-100 for 5 min, followed by
blocking with 5% bull serum albumin (Dingguo Changsheng
Biotechnology Co., Ltd., Beijing, China) at room temperature for 60
min. The fixed cells were incubated with specific primary α-SMA
monoclonal antibody (1:200, cat no. AA132; Beyotime Institute of
Biotechnology, Haimen, China) for 12 h at room temperature, washed
with PBS, and incubated with streptavidin biotin
complex-fluorescein isothiocyanate (Boster Biological Technology,
Pleasanton, CA, USA) at 37°C for 30 min. The cells were incubated
with DAPI (BestBio, Shanghai, China) at room temperature for 1 min,
immunofluorescence was observed and images were captured using an
Olympus immunofluorescence microscope (Olympus Corporation, Tokyo,
Japan) at ×40 magnification.
Western blot analysis
Cells were washed off the plates with PBS at 4°C and
total protein was extracted using radioimmunoprecipitation assay
lysis buffer (Boster Biological Technology) supplemented with
protease inhibitors (Nanjing KeyGen Biotech Co., Ltd., Nanjing,
China) for 30 min. The lysates were centrifuged at 14,000 × g for
15 min at 4°C. The protein concentration of lysates was measured
using a bicinchoninic acid protein assay (Beyotime Institute of
Biotechnology). Equal amounts of protein from each sample (40 µg)
were separated using 10% SDS-PAGE and transferred to a
nitrocellulose membrane (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The membranes were blocked with 5% non-fat dry milk in TBS
for 1 h at room temperature and incubated overnight at 4°C with the
following primary antibodies: Mouse monoclonal collagen type I
antibody (1:1,000; cat no. sc-59772; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), mouse anti-rat α-SMA monoclonal antibody
(1:1,000; cat no. AA132; Beyotime Institute of Biotechnology),
rabbit anti-rat phosphorylated (p)ERK1/2 (1:500; cat no. 4370L) and
ERK1/2 (1:500; cat no. 4695S) monoclonal antibodies, rabbit
anti-rat p-Smad2 (1:500; cat no. 3108P) and Smad2 (1:500; cat no.
3122S) monoclonal antibodies, rabbit anti-rat p-Smad3 (1:500; cat
no. 9520S) and Smad3 (1:500; cat no. 9523P) monoclonal antibodies
(all from Cell Signaling Technology, Inc., Danvers, MA, USA), and
rabbit anti-rat GAPDH polyclonal antibody (1:1,000; cat no. 25778;
Santa Cruz Biotechnology, Inc.), for 12 h at 4°C. Membranes were
washed three times and subsequently incubated with horseradish
peroxidase-conjugated goat anti-rabbit (1:2,000; cat no. A0208) or
goat anti-mouse (1:2,000; cat no. A0216) immunoglobulin G secondary
antibodies (Beyotime Institute of Biotechnology) at 37°C for 2 h.
The membranes were washed with PBS and were exposed to an enhanced
chemiluminescence reagent (Beyotime Institute of Biotechnology).
Protein bands were quantified using Quantity One software (version
4.6.3; Bio-Rad Laboratories, Inc.) and normalized against the
loading control, GAPDH.
Statistical analysis
All experimental data are presented as the mean ±
standard error of the mean and were analyzed using SPSS software
(version 17.0; SPSS Inc., Chicago, IL, USA). Statistical analysis
was performed using one-way analysis of variance and
Student-Newman-Keuls post hoc analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
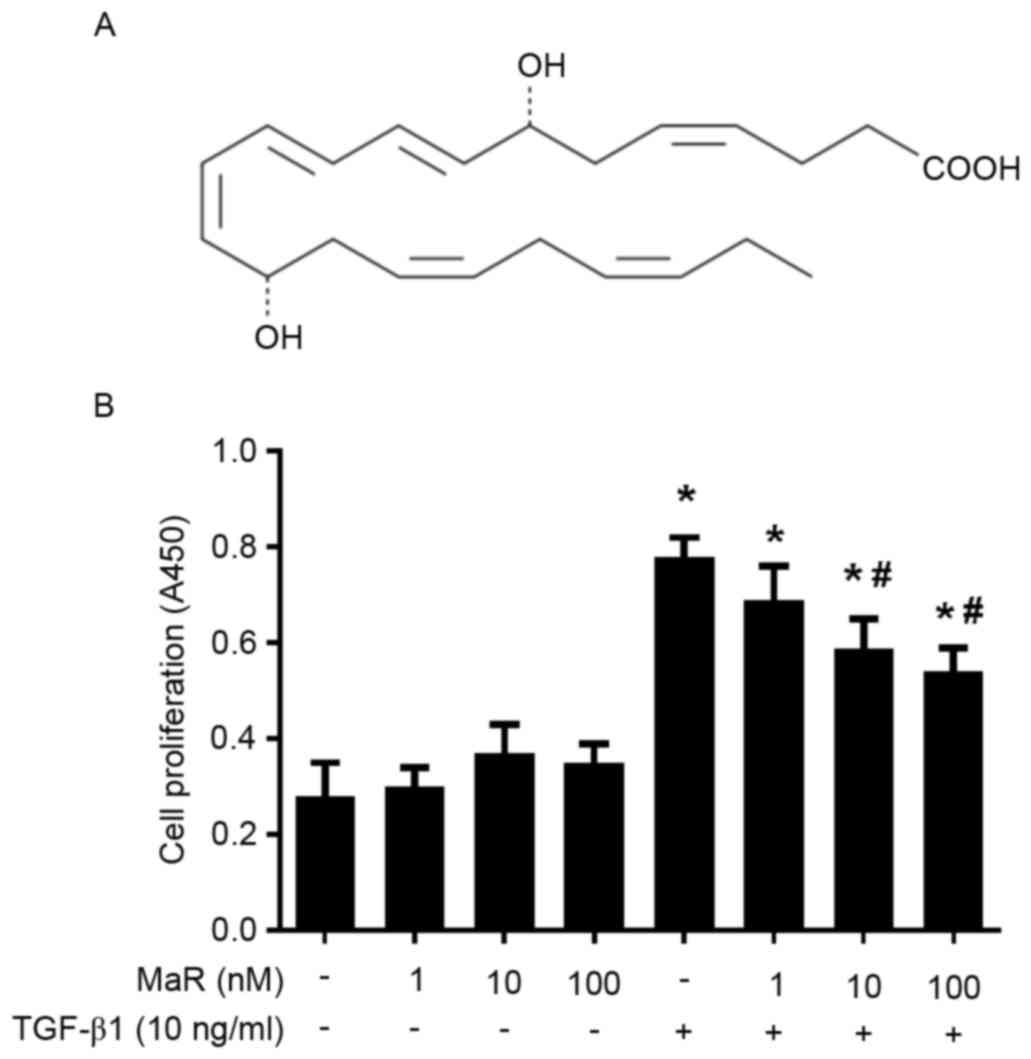
MaR 1 inhibits TGF-β1-induced
proliferation in MRC5 cells
As presented in Fig.
1B, TGF-β1 significantly increased MRC5 cell viability measured
by MTT assay (P<0.05). Incubation with MaR 1 alone exerted no
effects on MRC5 cell viability compared with the untreated group
(P>0.05). Pretreated with MaR 1, the TGF-β1-induced
proliferation of MRC5 cells was significantly decreased at 10 and
100 nM (P<0.05). The lower concentration of MaR 1 (1 nM) exerted
no effect on viability. As presented in Fig. 1B, the TGF-β1-induced proliferative
ability decreased in a dose-dependent manner. The concentrations of
10 ng/ml and 10 nM were selected to be the working concentration of
TGF-β1 and MaR 1, respectively.
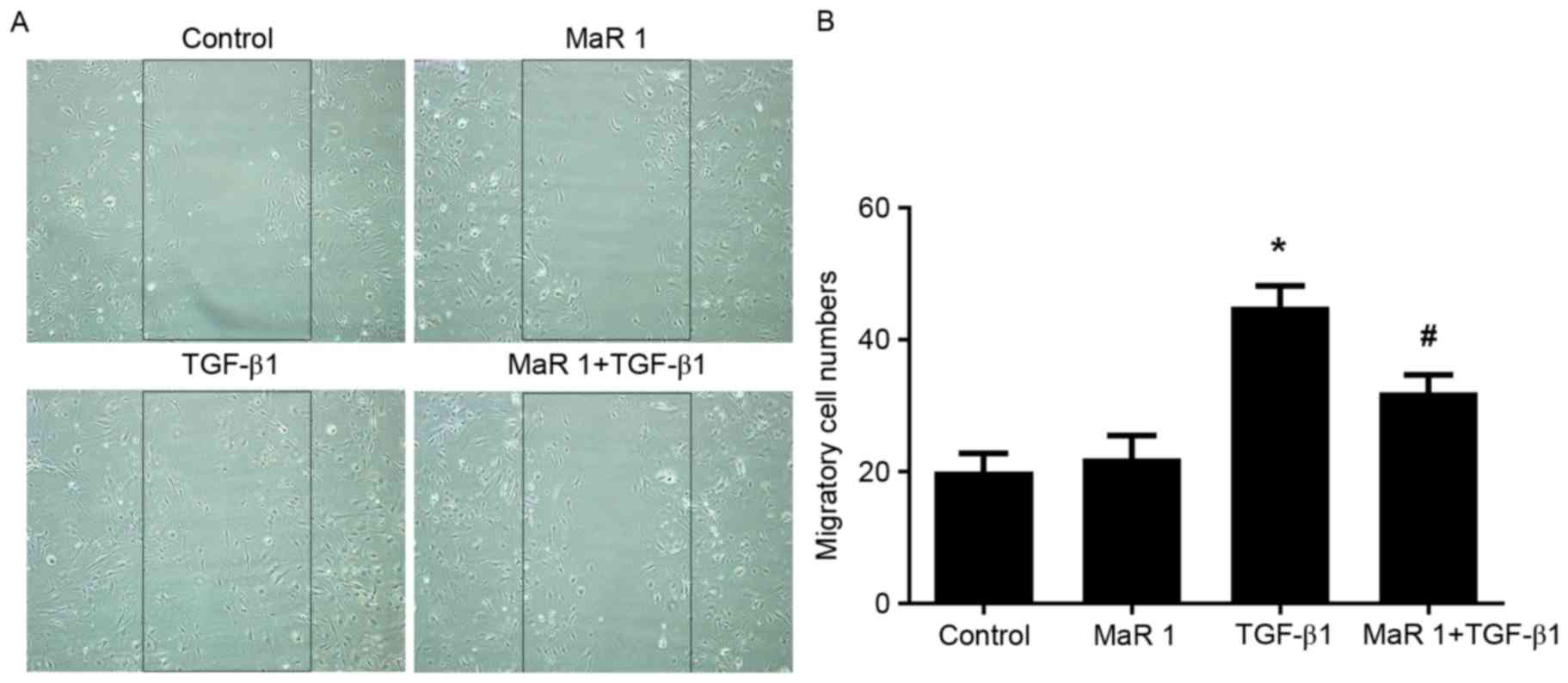
MaR 1 attenuates TGF-β1-induced
migration in MRC5 cells
The scratch assay was used to measure the effect of
MaR 1 on TGF-β1-induced fibroblast migration. TGF-β1 significantly
increased the migration of fibroblasts compared with the control
and MaR 1-only treated groups (P<0.05). MaR 1 alone exerted no
apparent influence on the migration of fibroblasts. However,
administration of MaR 1 significantly decreased fibroblast
migration and scratch closure induced by TGF-β1 (P<0.05;
Fig. 2).
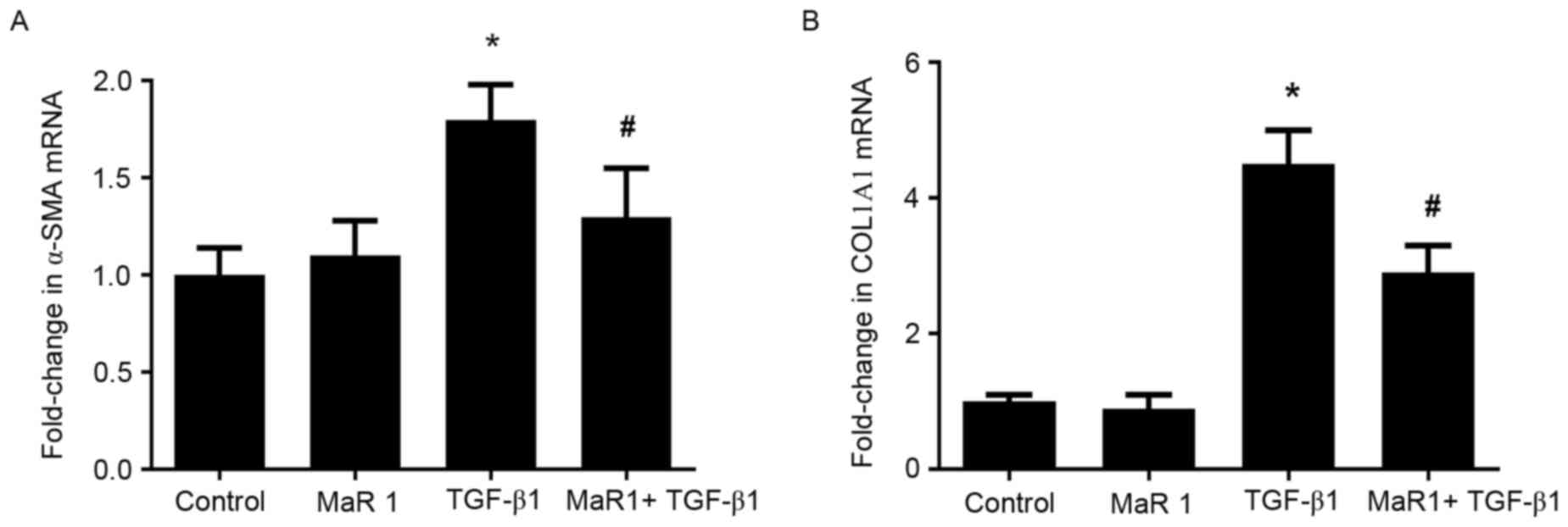
MaR 1 inhibits MRC5 cell
differentiation into myofibroblasts, induced by TGF-β1 in
vitro
In order to determine whether MaR 1 exerts an
inhibitory effect on the TGF-β1-induced differentiation of
fibroblasts into myofibroblasts, the expression of α-SMA and
collagen type I was examined in MRC5 cells treated with TGF-β1, in
the presence or absence of MaR 1. RT-qPCR analysis demonstrated
that administration of TGF-β1 resulted in a significant increase in
α-SMA and COL1A1 (the gene that encodes the pro-α1 chains of
collagen type I) mRNA levels compared with the control and MaR
1-only treated groups. By contrast, treatment with MaR 1 decreased
the mRNA level of α-SMA induced by TGF-β1. It was observed that MaR
1 decreased the TGF-β1-induced expression of COL1A1 mRNA, an
additional marker of myofibroblast differentiation (P<0.05;
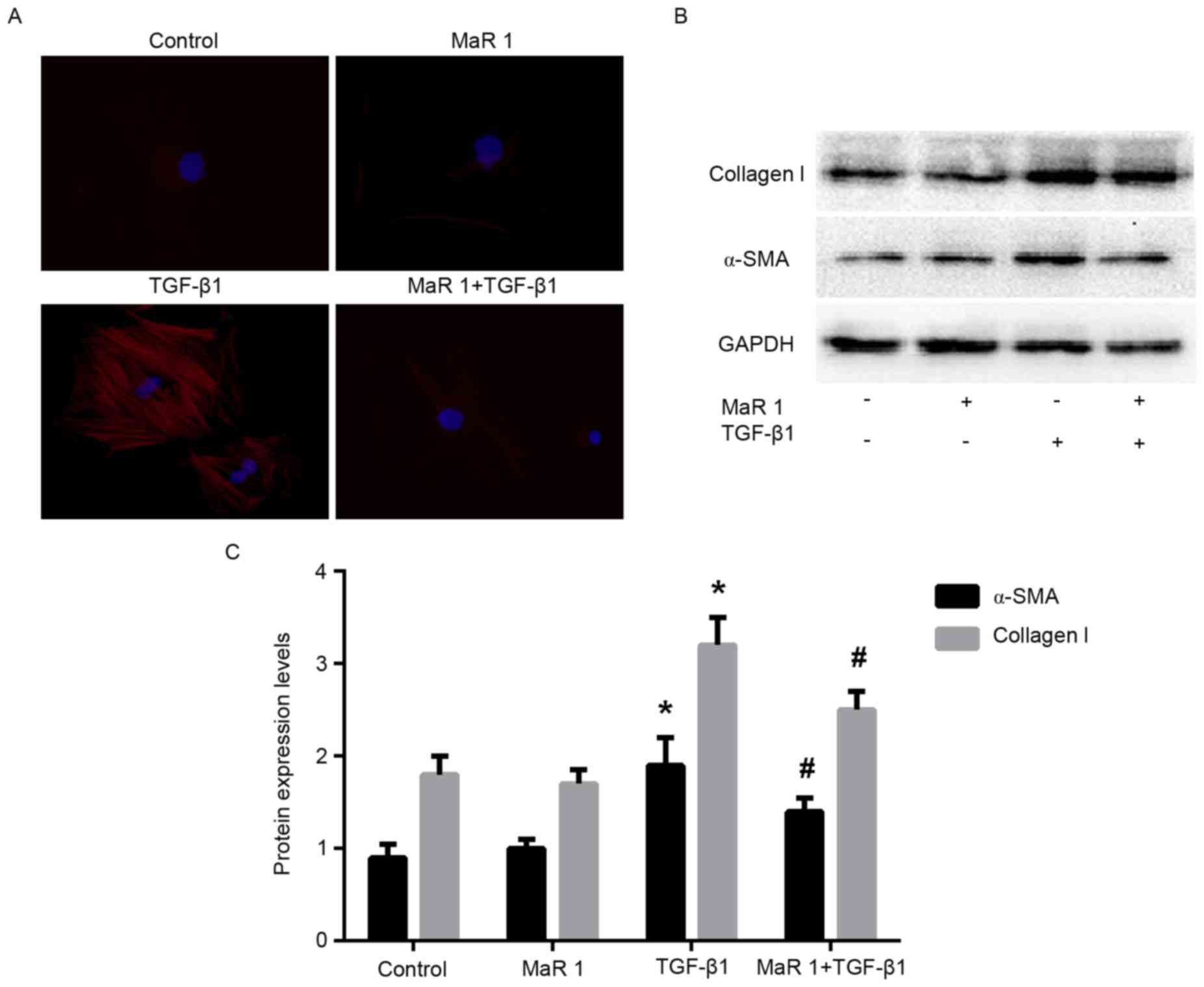
Fig. 3). Immunofluorescence
staining analysis demonstrated that treatment with TGF-β1 increased
the intensity of α-SMA staining in MRC5 cells; whereas, treatment
of MRC5 cells with TGF-β1 following MaR 1 significantly inhibited
this enhancement (P<0.05). In addition, MaR 1 alone exerted no
effect on α-SMA expression (Fig.
4A). Consistent with the results of the RT-qPCR and
immunofluorescence staining assays, western blot analysis
demonstrated that MaR 1 decreased the TGF-β1-induced expression of
α-SMA and collagen type I (P<0.05). However, MaR 1 alone
exhibited no effect on protein expression (Fig. 4B and C).
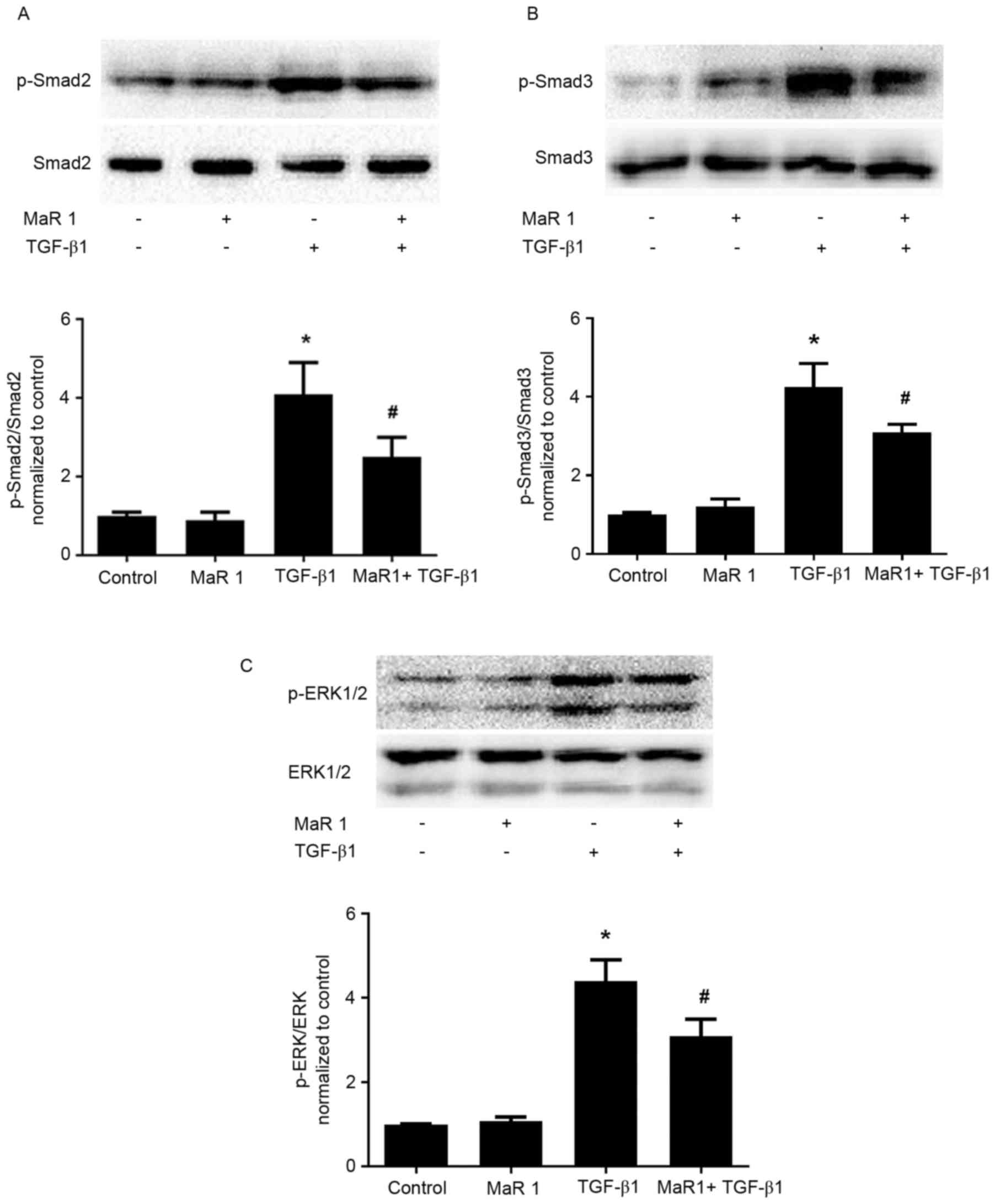
MaR 1 decreases TGF-β1-induced Smad2/3
and ERK1/2 phosphorylation in MRC5 cells
TGF-β1 signals were followed by the activation of
cytoplasmic effectors, including Smad and non-Smad mediator
proteins. In order to elucidate the possible mechanism involved in
the effect of MaR 1 on TGF-β1-stimulated profibrotic activity, the
expression of p-Smad2, p-Smad3, and p-ERK1/2 was evaluated. As
presented in Fig. 5, treatment
with TGF-β1 resulted in significantly increased phosphorylation of
Smad2, Smad3 and ERK1/2 compared with the control group.
Application of MaR 1 decreased the TGF-β1-induced expression of
p-Smad2, p-Smad3 and p-ERK1/2 in MRC5 cells, compared with
treatment with TGF-β1 alone. Treatment with MaR 1 alone exhibited
no effect on Smad and ERK mediator proteins.
Discussion
In the present study, it was demonstrated that
pretreatment with MaR 1 was able to attenuate TGF-β1-dependent lung
fibroblast proliferation and migration, TGF-β1-dependent increases
in lung fibroblast α-SMA and collagen type I expression and
TGF-β1-induced Smad2/3 and ERK1/2 phosphorylation in lung
fibroblasts. It was additionally observed that administration of
MaR 1 post-TGF-β1 stimulation was able to inhibit human lung
fibroblast proliferation, migration and differentiation in
vitro (data not presented).
The fibroblast/myofibroblast has been identified to
be an important pathogenic cell in all fibrotic diseases through
the secretion of excess ECM, cytokines and growth factors, and
tissue contraction. Myofibroblasts have three biological origins:
Tissue resident fibroblasts; epithelial to mesenchymal transition;
and bone marrow-derived mesenchymal precursors (19). Previous evidence has suggested that
myofibroblasts may be a target for anti-fibrotic therapy (6–8).
Lung fibroblast migration, proliferation and differentiation into a
myofibroblast-like cell type are regarded important steps in lung
fibrogenesis (5). It is well-known
that TGF-β1 is the most potent stimulator of the activation and the
differentiation of fibroblasts into myofibroblasts (20). Preventing myofibroblast
differentiation in migratory and proliferative fibroblasts has been
demonstrated to attenuate lung fibrosis (21).
Recently discovered pro-resolving lipid mediators,
including lipoxins, protectins, resolvins and maresins, have been
reported to exhibit potent anti-inflammatory and pro-resolving
effects in multiple disease models, by acting on neutrophils,
macrophages and other inflammatory cells (9,10).
Previous research demonstrated that pro-resolving lipid mediators
exhibit anti-fibrotic and anti-proliferative abilities. Lipoxin A4
was demonstrated to inhibit the proliferation of human lung
fibroblasts evoked by connective tissue growth factor, and
suppressed the tumor necrosis factor-α-induced proliferation of rat
mesangial cells (22,23). Aspirin-triggered-15R-lipoxin A4 and
lipoxin A4 inhibited endothelial cell proliferation stimulated with
vascular endothelial growth factor or leukotriene D4 (24). Systemic administration of resolvin
2 and MaR 1 attenuated neointimal hyperplasia and vessel remodeling
in a mouse model of arterial injury, by reducing early
proliferation in the vessel wall and inhibiting vascular smooth
muscle cell migration mediated by platelet-derived growth factor
(16). As demonstrated by the MTT
and scratch wound healing assays, the present study demonstrated
that MaR 1 is able to inhibit fibroblast proliferation and
migration stimulated by TGF-β1.
Differentiated myofibroblasts are characterized by
increased expression of collagen and α-SMA, the most commonly used
molecular markers (6). The results
of the present study demonstrated that MaR 1 prevents
TGF-β1-induced myofibroblast differentiation in cultured lung
fibroblasts, as exhibited by the underexpression of collagen type I
and α-SMA at the mRNA and protein levels. It was hypothesized that,
in addition to inhibiting epithelial-mesenchymal transition in lung
alveolar epithelial cells (17),
MaR 1 may inhibit the effects of TGF-β1 on fibroblasts,
demonstrating a second, synergistic mechanism whereby MaR 1 exerts
its anti-fibrotic effects. To the best of our knowledge, the
present study is the first to directly demonstrate that MaR 1
inhibits TGF-β1-induced myofibroblast differentiation from lung
fibroblasts.
TGF-β1 exhibits numerous biological functions
associated with airway remodeling, fibrosis and myofibroblast
differentiation, including enhanced cellular migration,
proliferation and ECM synthesis (25,26).
TGF-β1 elicits its biological activities by signaling through a
receptor complex of serine/threonine kinase type I and type II
receptors, which induces signal transduction through
receptor-mediated Smad signaling pathway in addition to non-Smad
signaling pathways (27). In order
to investigate the possible signaling cascade by which MaR 1 may
suppress human lung fibroblast cell activation, the effects of MaR
1 on the Smad2/3 and ERK1/2 signaling pathways in TGF-β1-stimulated
fibroblasts were investigated. It has been demonstrated that the
TGF-β1/Smad pathway is activated in myofibroblast differentiation,
and inhibiting its phosphorylation may attenuate myofibroblast
differentiation and the fibrogenic response. Dong et al
(28) demonstrated that
interleukin-27 inhibited the proliferation, differentiation and
collagen synthesis of lung fibroblasts by reducing the activation
of the TGF-β1/Smad signaling pathways. In cystic fibrosis lungs,
TGF-β1 signaling was markedly increased, as observed by p-Smad2
expression, increased myofibroblast differentiation and tissue
fibrosis (29). Tumelty et
al (30) reported that aortic
carboxypeptidase-like protein increased α-SMA and collagen protein
expression by inducing Smad2/3 phosphorylation in primary lung
fibroblasts and promoted myofibroblast differentiation. As
expected, the results of the present study demonstrated that
p-Smad2 and p-Smad3 were highly expressed in TGF-β1 treated cells
compared with controls, and that this effect was inhibited by
treatment with MaR 1. The results of the present study suggested
that MaR 1 inhibited myofibroblast differentiation, in part through
the inhibition of Smad2 and Smad3 phosphorylation.
In addition to Smad signaling pathways, previous
studies have demonstrated that non-Smad pathways, including ERK1/2,
may serve a role in TGF-β1-mediated biological activities. The
ratio of p-ERK to ERK indicates the degree of activation of the ERK
pathway, which serves a role in a number of
fibroblast/myofibroblast functions, including proliferation,
migration and differentiation. A previous study demonstrated that
MaR 1 may protect against liver injury induced by carbon
tetrachloride by inactivating mitogen-activated protein
kinase/ERK1/2 signaling pathways (15). An additional previous study
demonstrated that the downregulation of ERK1/2 inhibited high
mobility group protein B1-induced myofibroblast differentiation and
migration, using the human lung fibroblast cell line WI-38
(31). In addition, early reports
indicated that IM-412 inhibited TGF-β1 induced expression of the
fibrotic markers α-SMA and fibronectin, and collagen accumulation
in human lung fibroblasts by decreasing Smad2 and Smad3
phosphorylation, in addition to ERK activity (32). Chung et al (33) reported that resistin like α induced
myofibroblast differentiation by increasing the rapid
phosphorylation of the ERK signaling pathway. Consistent with this
report, the results of the present study demonstrated that the
effects of MaR 1 were partly mediated by decreasing ERK1/2
phosphorylation in TGF-β1-treated human lung fibroblasts. A number
of growth factors stimulate ERK1/2, and it has been reported that
activated ERK phosphorylates Smad proteins within their linker
regions, resulting in the maintenance of Smad-mediated biological
activities (34). However, the
precise mechanisms through which MaR 1 attenuates the
TGF-β1-induced proliferation, migration and differentiation of
human lung fibroblasts requires further clarification.
In conclusion, the results of the present study
demonstrated that MaR 1 inhibited TGF-β1-dependent profibrotic
activity in lung fibroblasts when administered prior to or
following TGF-β1 stimulation, and that the protective effect of MaR
1 may be associated with inactivation of the Smad2/3 and ERK1/2
signaling pathways in lung fibroblasts. The present study provided
evidence for the potential role of MaR 1 in fibrotic lung disease.
In addition to lung fibrosis, it is hypothesized that MaR 1 may act
on other types of fibroblasts, representing a potentially useful
therapeutic strategy in other fibrotic diseases.
References
|
1
|
Postma DS and Timens W: Remodeling in
asthma and chronic obstructive pulmonary disease. Proc Am Thorac
Soc. 3:434–439. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ding NH, Li JJ and Sun LQ: Molecular
mechanisms and treatment of radiation-induced lung fibrosis. Curr
Drug Targets. 14:1347–1356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ramirez AM, Shen Z, Ritzenthaler JD and
Roman J: Myofibroblast transdifferentiation in obliterative
bronchiolitis: Tgf-beta signaling through smad3-dependent and
-independent pathways. Am J Transplant. 6:2080–2088. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wilson MS and Wynn TA: Pulmonary fibrosis:
Pathogenesis, etiology and regulation. Mucosal Immunol. 2:103–121.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Noble PW, Barkauskas CE and Jiang D:
Pulmonary fibrosis: Patterns and perpetrators. J Clin Invest.
122:2756–2762. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maharaj S, Shimbori C and Kolb M:
Fibrocytes in pulmonary fibrosis: A brief synopsis. Eur Respir Rev.
22:552–557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sivakumar P, Ntolios P, Jenkins G and
Laurent G: Into the matrix: Targeting fibroblasts in pulmonary
fibrosis. Curr Opin Pulm Med. 18:462–469. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Phan SH: The myofibroblast in pulmonary
fibrosis. Chest. 122 Suppl 6:S286–S289. 2002. View Article : Google Scholar
|
|
9
|
Serhan CN: Pro-resolving lipid mediators
are leads for resolution physiology. Nature. 510:92–101. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Serhan CN and Chiang N: Resolution phase
lipid mediators of inflammation: Agonists of resolution. Curr Opin
Pharmacol. 13:632–640. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dalli J, Zhu M, Vlasenko NA, Deng B,
Haeggström JZ, Petasis NA and Serhan CN: The novel
13S,14S-epoxy-maresin is converted by human macrophages to maresin
1 (MaR1), inhibits leukotriene A4 hydrolase (LTA4H), and shifts
macrophage phenotype. FASEB J. 27:2573–2583. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marcon R, Bento AF, Dutra RC, Bicca MA,
Leite DF and Calixto JB: Maresin 1, a proresolving lipid mediator
derived from omega-3 polyunsaturated fatty acids, exerts protective
actions in murine models of colitis. J Immunol. 191:4288–4298.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nordgren TM, Heires AJ, Wyatt TA, Poole
JA, LeVan TD, Cerutis DR and Romberger DJ: Maresin-1 reduces the
pro-inflammatory response of bronchial epithelial cells to organic
dust. Respir Res. 14:512013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gong J, Wu ZY, Qi H, Chen L, Li HB, Li B,
Yao CY, Wang YX, Wu J, Yuan SY, et al: Maresin 1 mitigates
LPS-induced acute lung injury in mice. Br J Pharmacol.
171:3539–3550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li R, Wang Y, Zhao E, Wu K, Li W, Shi L,
Wang D, Xie G, Yin Y, Deng M, et al: Maresin 1, a proresolving
lipid mediator, mitigates carbon tetrachloride-induced liver injury
in mice. Oxid Med Cell Longev. 2016:92037162016.PubMed/NCBI
|
|
16
|
Akagi D, Chen M, Toy R, Chatterjee A and
Conte MS: Systemic delivery of proresolving lipid mediators
resolvin D2 and maresin 1 attenuates intimal hyperplasia in mice.
FASEB J. 29:2504–2513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Li R, Chen L, Tan W, Sun Z, Xia H,
Li B, Yu Y, Gong J, Tang M, et al: Maresin 1 inhibits
epithelial-to-mesenchymal transition in vitro and attenuates
bleomycin induced lung fibrosis in vivo. Shock. 44:496–502. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hinz B, Phan SH, Thannickal VJ, Galli A,
Bochaton-Piallat ML and Gabbiani G: The myofibroblast: One
function, multiple origins. Am J Pathol. 170:1807–1816. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Santana A, Saxena B, Noble NA, Gold LI and
Marshall BC: Increased expression of transforming growth factor
beta isoforms (beta 1, beta 2, beta 3) in bleomycin-induced
pulmonary fibrosis. Am J Respir Cell Mol Biol. 13:34–44. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang S, Cui H, Xie N, Icyuz M, Banerjee S,
Antony VB, Abraham E, Thannickal VJ and Liu G: miR-145 regulates
myofibroblast differentiation and lung fibrosis. FASEB J.
27:2382–2391. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu SH, Wu XH, Lu C, Dong L and Chen ZQ:
Lipoxin A4 inhibits proliferation of human lung fibroblasts induced
by connective tissue growth factor. Am J Respir Cell Mol Biol.
34:65–72. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu SH, Wu XH, Lu C, Dong L, Zhou GP and
Chen ZQ: Lipoxin A4 inhibits connective tissue growth
factor-induced production of chemokines in rat mesangial cells.
Kidney Int. 69:248–256. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fierro IM, Kutok JL and Serhan CN: Novel
lipid mediator regulators of endothelial cell proliferation and
migration: Aspirin-triggered-15R-lipoxin A(4) and lipoxin A(4). J
Pharmacol Exp Ther. 300:385–392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Blobe GC, Schiemann WP and Lodish HF: Role
of transforming growth factor beta in human disease. N Engl J Med.
342:1350–1358. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Camoretti-Mercado B and Solway J:
Transforming growth factor-beta1 and disorders of the lung. Cell
Biochem Biophys. 43:131–148. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Samarakoon R, Overstreet JM and Higgins
PJ: TGF-β signaling in tissue fibrosis: Redox controls, target
genes and therapeutic opportunities. Cell Signal. 25:264–268. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dong Z, Zhao X, Tai W, Lei W, Wang Y, Li Z
and Zhang T: IL-27 attenuates the TGF-β1-induced proliferation,
differentiation and collagen synthesis in lung fibroblasts. Life
Sci. 146:24–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Harris WT, Kelly DR, Zhou Y, Wang D,
MacEwen M, Hagood JS, Clancy JP, Ambalavanan N and Sorscher EJ:
Myofibroblast differentiation and enhanced TGF-B signaling in
cystic fibrosis lung disease. PLoS One. 8:e701962013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tumelty KE, Smith BD, Nugent MA and Layne
MD: Aortic carboxypeptidase-like protein (ACLP) enhances lung
myofibroblast differentiation through transforming growth factor β
receptor-dependent and -independent pathways. J Biol Chem.
289:2526–2536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee CC, Wang CN, Lee YL, Tsai YR and Liu
JJ: High mobility group box 1 induced human lung myofibroblasts
differentiation and enhanced migration by activation of MMP-9. PLoS
One. 10:e01163932015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park S, Ahn JY, Lim MJ, Kim MH, Lee SL,
Yun YS, Jeong G and Song JY: IM-412 inhibits transforming growth
factor beta-induced fibroblast differentiation in human lung
fibroblast cells. Biochem Biophys Res Commun. 399:268–273. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chung MJ, Liu T, Ullenbruch M and Phan SH:
Antiapoptotic effect of found in inflammatory zone (FIZZ)1 on mouse
lung fibroblasts. J Pathol. 212:180–187. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hough C, Radu M and Doré JJ: Tgf-beta
induced Erk phosphorylation of smad linker region regulates smad
signaling. PLoS One. 7:e425132012. View Article : Google Scholar : PubMed/NCBI
|