Introduction
Approximately ten million patients receive general
anesthesia for surgery in China every year. While intravenous
anesthetics cause unconsciousness, the mechanism and neural basis
of unconsciousness are poorly understood (1). At the molecular level, there are
dozens of molecules known to be general anesthetic targets,
including a number of ion channels (2), gap-junction channels (3), and G protein-coupled receptors
(4). It's remarkable that there is
no single molecular target shared by all general anesthetics
(1). Therefore, effects of general
anesthetics must be comprehended in the context of network
connectivity.
There are similarities between general anesthesia
and natural sleep. Imaging studies have shown some parallels
between the anesthetized brain and the brain during deep
non-rapid-eye-movement (NREM) sleep (5,6).
Electroencephalogram (EEG) studies have suggested that loss of
consciousness caused by general anesthetics resembles the rapid
transition from normal wakefulness to sleep (7). Sleep-related EEG waves that resembled
gamma, delta and spindle waves have been observed during general
anesthesia (8). These findings led
an increasingly popular theory that anesthetics may induce
unconsciousness by acting on endogenous sleep-arousal neural
circuitry. But it remains unclear to what extent sleep-arousal
pathway, such as the ventrolateral preoptic nucleus (VLPO)-locus
coeruleus (LC) are involved in generating the hypnotic state.
Commonly used general anesthetic propofol exerts
sedative effects by targeting GABAA receptors. And GABA
is the primary inhibitory neurotransmitter released by
sleep-promoting neurons in the VLPO, which plays a critical role in
inducing and maintaining sleep (9). The VLPO sends GABAergic inhibitory
projections to several wake-promoting nuclei throughout the
neuroaxis (10), including the LC,
tuberomammillary nucleus (TMN) and orexinergic neurons in the
lateral hypothalamus (11).
Previous research has demonstrated that propofol and various
barbiturates activate sleep-promoting VLPO neurons through
different receptors (12–14). Moreover, the inhalational
anesthetic isoflurane directly depolarizes VLPO neurons (15). Nevertheless, lesion of VLPO neurons
could be expected to produce resistance to anesthesia because of
the accrual of sleep debt (16).
Thus, it remains controversial whether VLPO activation contributes
to anesthetic-induced unconsciousness.
In the present study, we hypothesized that propofol
may act on VLPO neurons to stimulate the release of GABA, thereby
inhibiting wakefulness-promoting neurons in the LC. To validate the
hypothesis, we examined the LoRR (loss of righting reflex) and RoRR
(recovery of righting reflex) time following GABAA
receptor agonist or antagonist microinjection. We also recorded the
firing activities of LC neurons under different concentrations of
propofol. The results demonstrated that spontaneous firing of LC
noradrenergic neurons was inhibited by propofol. Furthermore, the
results also showed that firing activities were not significantly
different between sham-lesion and VLPO lesion rats. Suggesting that
VLPO neurons were not likely involved in propofol-mediated
inhibition on LC neurons.
Materials and methods
Animals
Male Sprague-Dawley rats weighing 260–300 g (n=72)
were housed in an isolated chamber at 20–22°C under a 12 h
light/dark cycle (lights on at 8:30 AM). Food and water were
available ad libitum. Twenty-four rats were used for
righting reflex behavioral assays, and the remaining rats were used
for in vivo electrophysiological recordings. Forty-eight
neurons were recorded from forty-eight rats. All animals were
supplied by the laboratory animal center of the Third Military
Medical University (Chongqing, China). Animal experiments were
approved by the Zunyi Medical College Animal Care and Use Committee
(approval number: 2016126). All efforts were made to minimize the
number of rats and their suffering.
Surgical procedures and cannulas
implantation
For behavioral experiments, rats were anesthetized
with sodium pentobarbital [50 mg/kg, intraperitoneally (i.p.)] and
atropine sulfate (0.2 mg, i.p.). Anesthetized animals were mounted
onto a stereotaxic device (68505; RWD Life Science, China) in a
flat-skull position. The core body temperature was maintained at
37–38°C using a heat-controlled pad equipped with a rectal probe
(Stoelting Co., Wood Dale, IL, USA). Sterilized guide cannulas were
implanted as described previously (17,18).
A pair of sterilized guide cannulas made of 23G stainless steel
tubes and plugged internally with 30G stylets were stereotaxically
implanted 2.0 mm above the VLPO (anteroposterior, −0.35 mm;
left-right, ± 1.2 mm; dorsoventral, −7.0 mm from bregma).
Stereotaxic positioning was defined according to the brain atlas of
Paxinos and Watson (2007). Guide cannulas were fixed to the skull
using dental cement.
Lesion formation by ibotenic acid and
saline injection
To induce lesions in the VLPO, rats were
anesthetized and kept in place as mentioned above. The skull of
each rat was exposed, and a glass micropipette (10–12-µm tip
diameter) was lowered into the VLPO region stereotaxically. Fifteen
nanoliters of ibotenic acid solution (10 nmol; Sigma, St. Louis,
MO, USA) (n=24) or saline (n=24) were injected into the VLPO
bilaterally using a microinjection injector (Nanoliter 2000, World
Precision Instruments, Sarasota, FL, USA) as described previously
(16). The glass micropipette was
slowly withdrawn after 5 min. The two holes above VLPO were filled
with bone wax. The wound was stitched with sutures and closed with
wound clips. A prophylactic dose of penicillin (50 ku/kg,
intramuscular injection) was injected into each rat following
surgery. After seven days of recovery, in vivo
electrophysiology experiments were conducted.
Righting reflex behavioral assays
In rodents, loss of consciousness can be measured by
the LoRR and resumption of consciousness can be analyzed by the
RoRR (19,20). Fig.
1A illustrates the experimental design used to quantify the
LoRR and RoRR after the VLPO was microinjected with artificial
cerebrospinal fluid (ACSF) (126 mM Na+, 3 mM
K+, 2 mM Ca2+, 1.2 mM Mg2+, 150 mM
Cl−, pH=7.4) and a bolus injection of propofol (11
mg/kg, i.v.). After seven days of recovery from cannulas
implantation surgery, rats (n=24) were put into a Plexiglas
inhalation anesthesia induction chamber for 10-min acclimation
(Fig. 1A; t=−40 min). Then, oxygen
containing 2% isoflurane was introduced into the chamber at 2 l/min
(Fig. 1A; t=−30 min). The 30G
stylet from the guide cannula was removed from the
isoflurane-anesthetized rats, and a 31G injection cannula was
inserted. An injection cannula was attached to a dual-channel
microinjection syringe with PE tubing under the control of an
injection pump (302; RWD Life Science), and then a 24G intravenous
catheter was implanted and fixed to the rat's tail vein. Isoflurane
administration was subsequently discontinued. The ACSF (t=0 min;
0.2 µl/side; n=24) (Fig. 1A) was
microinjected into the bilateral VLPO at an infusion speed of 0.2
µl/min for 1 min (t=1 to t=2) (Fig.
1A). Five min later (t=7 min) (Fig. 1A), propofol (11 mg/kg, n=24) was
bolus injected into the tail vein. The induction time of propofol
was quantified as the time (sec) to LoRR; the resumption time was
quantified as the time to RoRR (sec). Three days later, the
twenty-four rats were subdivided into GABAA agonist group (n=12)
and GABAA antagonist group (n=12). Rats in the agonist group were
microinjected with the GABAA agonist 4,5,6,7-tetrahydroisoxazolo
(5,4-c) pyridin-3-ol (THIP), also called Gaboxadol (t=0; n=12)
(Fig. 1B). Rats in the GABAA
antagonist group received the same dose of gabazine (t=0; n=12)
(Fig. 1C), and the timeline and
procedures were the same as in the ACSF group.
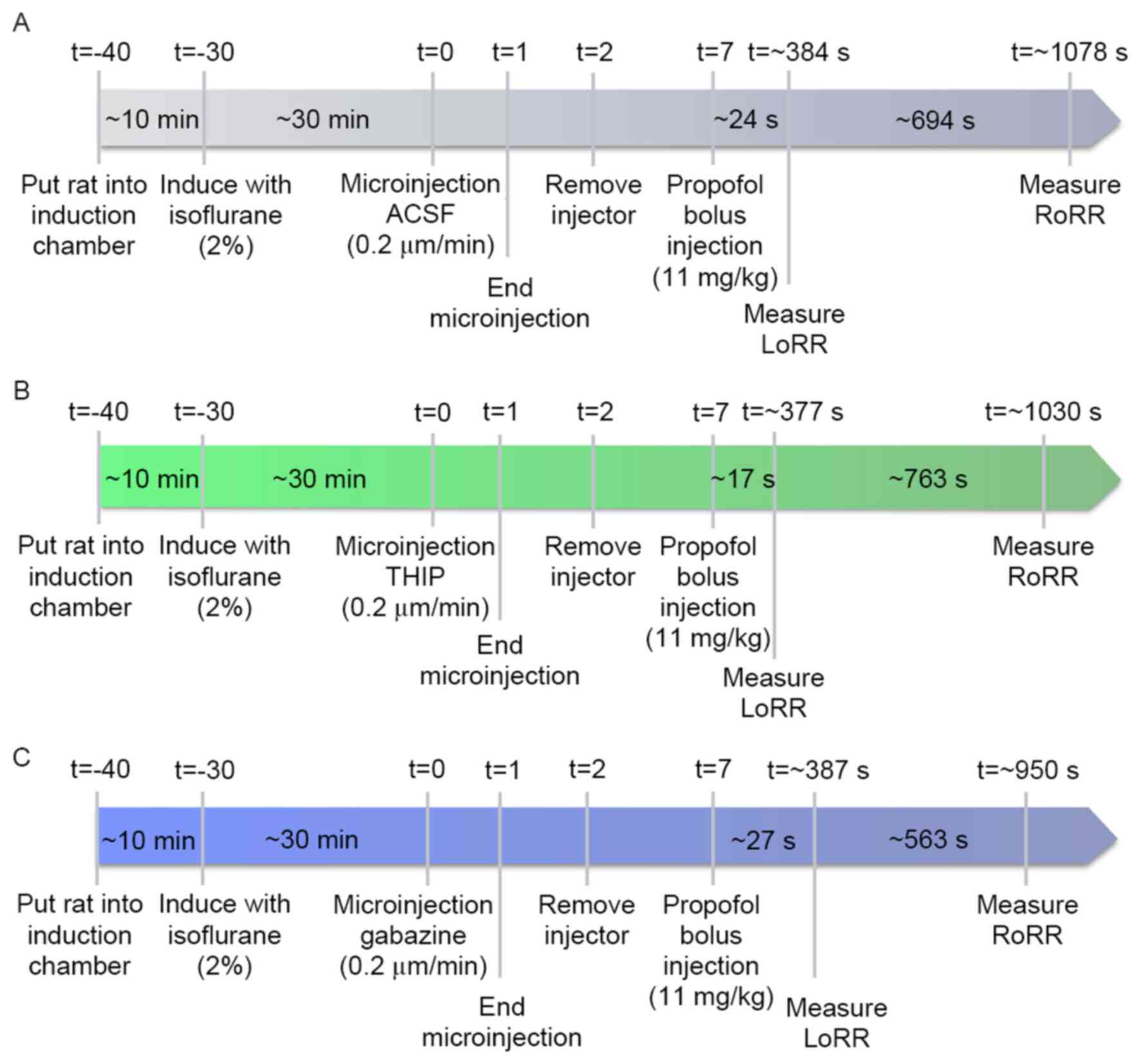 | Figure 1.(A) Timelines of LoRR and RoRR
measurement. Rats were left for 30 min after isoflurane
administration for the righting reflex to recover before
microinjection of ACSF (vehicle control), THIP (B), and gabazine
(C) into the VLPO and before propofol bolus injection. ACSF (A),
THIP (B), and gabazine (C) were microinjected while the rats were
awake. Five min after injection, propofol was bolus injected (11
mg/kg) into the tail vein. The LoRR recorded and quantified
immediately. Then, rats were placed in a supine position and the
RoRR was measured. All experimental procedures were the same in the
agonist (B), antagonist (C), and control (A) groups except for the
microinjection of drugs. LoRR, loss of righting reflex; RoRR,
resumption of righting reflex; ACSF, artificial cerebrospinal
fluid. |
Extracellular single-unit
recordings
After a recovery period of seven days, the
extracellular single-unit recording was performed from
noradrenergic neurons of the LC (LC-NA) with VLPO lesions (n=24)
and sham lesions (n=24) rats. Each group was subdivided into a
consciousness group (n=6, without propofol administration), a low
propofol concentration group (n=6; 20 mg/kg/h, i.v.), medium
concentration group (n=6; 40 mg/kg/h, i.v.) and a high
concentration propofol group (n=6; 60 mg/kg/h, i.v.).
The protocol of extracellular single-unit recordings
in non-anesthetized rats was refereed to previously report
(21). Briefly, the rats were
progressively habituated to the head-restrained position (7–14
days) by placing them inside a plastic chamber, painlessly
restraining the head with a head holder and preventing large body
movements with a cotton-coated plastic cover. The eyes were covered
to reduce visual stimulation. After 7–14 days of habituation, the
rats could be kept in a sphinx position for 3–6 consecutive h
without showing any signs of discomfort. If any signs of discomfort
were seen, the rats were freed from the restrained position.
Then A 24G intravenous catheter was implanted and
fixed as described above. Rats were removed from the induction
chamber and mounted onto a stereotaxic device with blunt ear bars.
Propofol was continuously delivered via a syringe pump (WZS-50F6;
Smith Medical, Plymouth, MN, US) connected to the intravenous
catheter in propofol administration groups. LC-NA extracellular
recordings were obtained as previously described (21). A single barrel glass micropipette
was filled with 0.5% sodium acetate and lowered into the LC (3.5 mm
caudal to lambda, 0.85–1.0 mm lateral to the midline, 5–6.5-mm
below the cortical surface) by using a micromanipulator (Mini 25;
Luigs & Neumann, Ratingen, Germany) according to coordinates
defined in the brain atlas of Paxinos and Watson (2007). The
impedance of the micropipette was 10–20 MΩ, measured in
physiological saline. Recording was performed during a period of at
least 5 min, with only one cell measured from each rat. The
recorded extracellular signals were amplified using a patch clamp
amplifier (EPC10; HEKA Elektronik, Lambrecht, Germany), bandpass
filtered (100–10,000 Hz) and saved to a computer installed with
PATCHMASTER (Heka) and Mini Analysis System (Synaptosoft, Inc.,
Fort Lee, NJ, USA) for offline analysis. The body temperature of
rats was maintained at 37–38°C during the recording period.
Noxious cutaneous stimulation
The discharge patterns of NA-LC neurons were
identified using standard criteria (22,23).
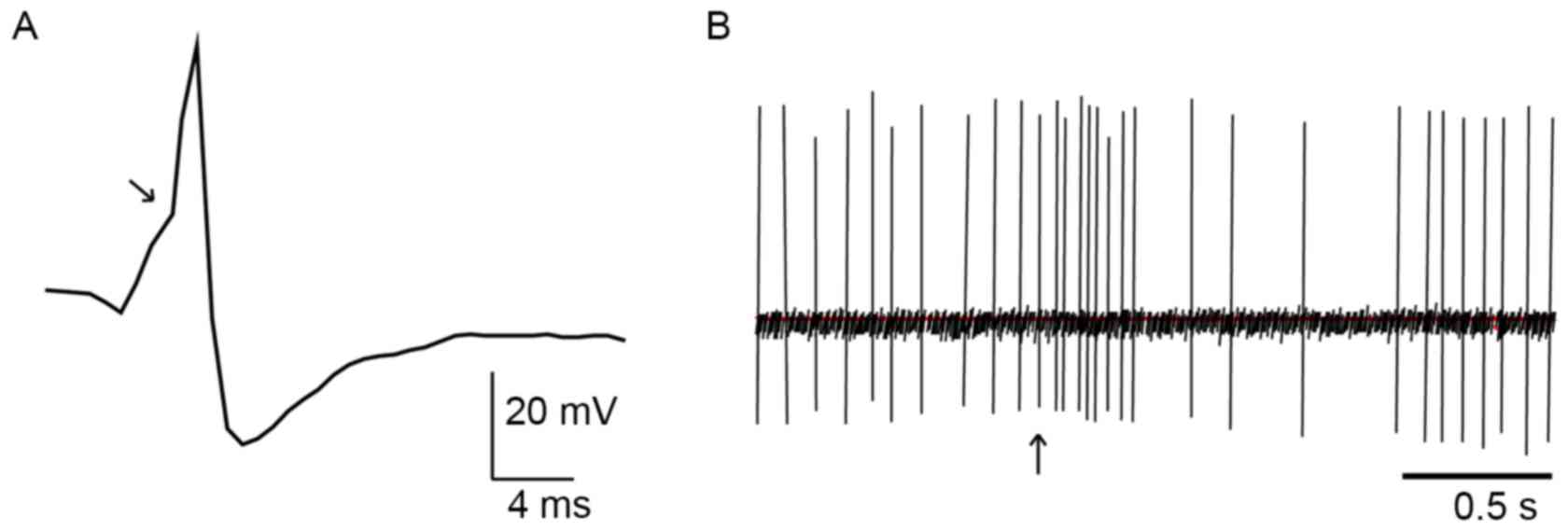
They display a positive-negative action potential shape with a
notch on the ascending limb (Fig.
2A) and a typical, biphasic excitation-inhibition response to
noxious stimulation of the contralateral hind paw (Fig. 2B). We pinched the skin of the rat's
hind paw using toothed forceps for cutaneous nociceptive
stimulation to induce this characteristic firing response. The
noxious stimulation procedure was only operated in anesthetized
rats.
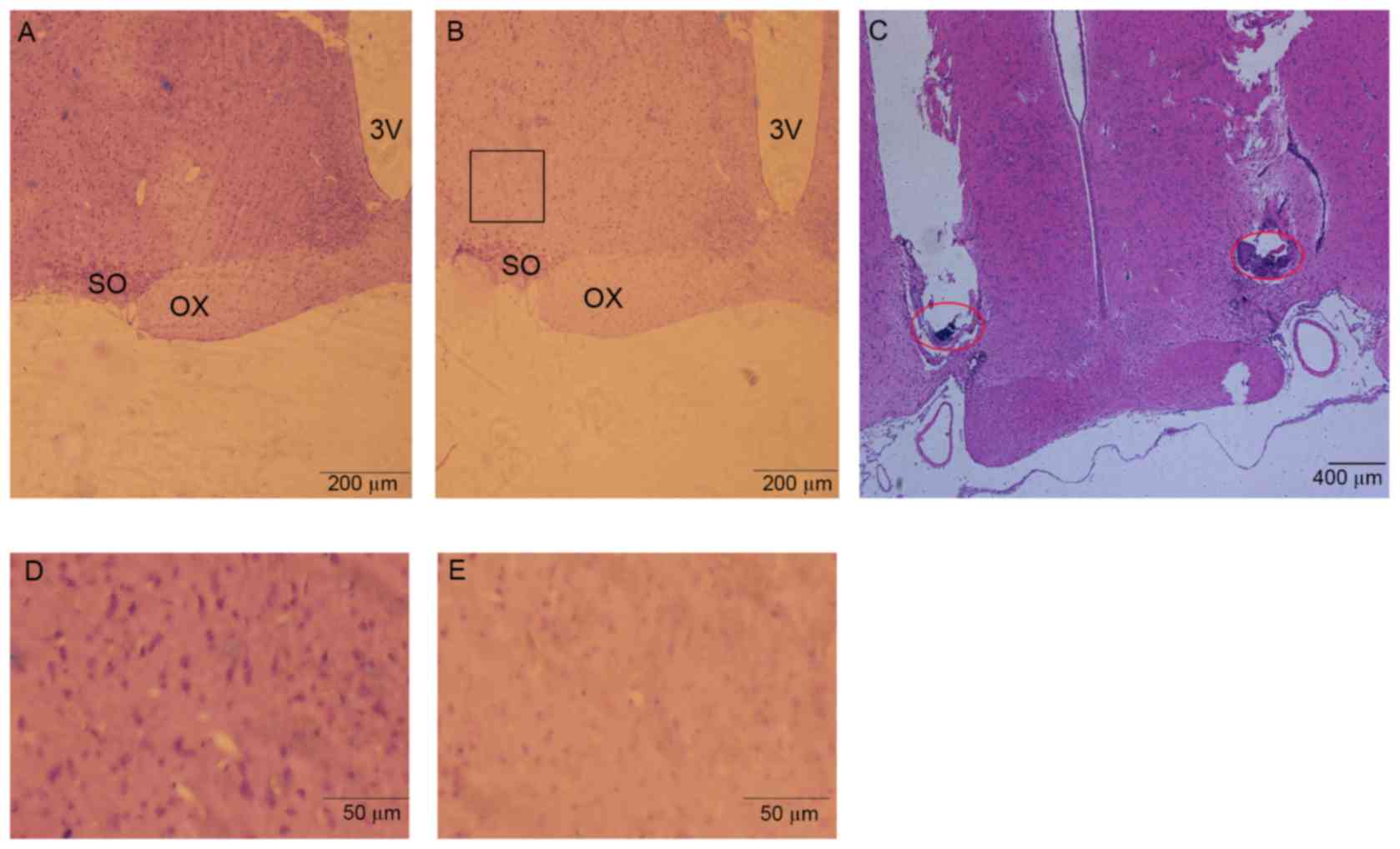
Histological localization of
microinjection and lesion areas
After behavioral and electrophysiology experiments,
rats were deeply anesthetized with sodium pentobarbital. Cold 100
ml physiological saline was perfused through the ascending aorta,
followed by 200 ml 4% paraformaldehyde. The brains were then
removed, post-fixed for 24 h at 4°C in paraformaldehyde, and
transferred to glass containers filled with 30% sucrose for 48 h at
4°C. 20-µm coronal sections containing microinjection or lesion
sites were cut and stained with hematoxylin-eosin for histological
verification (Fig. 3). Stereotaxic
coordinates were identified according to the brain atlas of Paxinos
and Watson (2007).
Statistical analyses
Statistical analyses were performed using SPSS 17.0
for Windows. All data were expressed as means ± standard deviation.
A P-value of <0.05 was considered statistically significant
using paired-samples t-test in behavioral experiments. N refers to
the number of rats studied. The firing frequencies of noradrenergic
neurons in different groups were compared using independent-samples
t-test. The firing frequencies of noradrenergic neurons in the same
group were compared using one-way ANOVA, and n refers to the number
of neurons analyzed.
Results
The effects of GABAA
receptors in the VLPO on LoRR and RoRR
To determine whether GABAergic neurons are involved
in unconsciousness induced by propofol. We microinjected GABAA
agonist (THIP) or GABAA antagonist (gabazine) into VLPO and
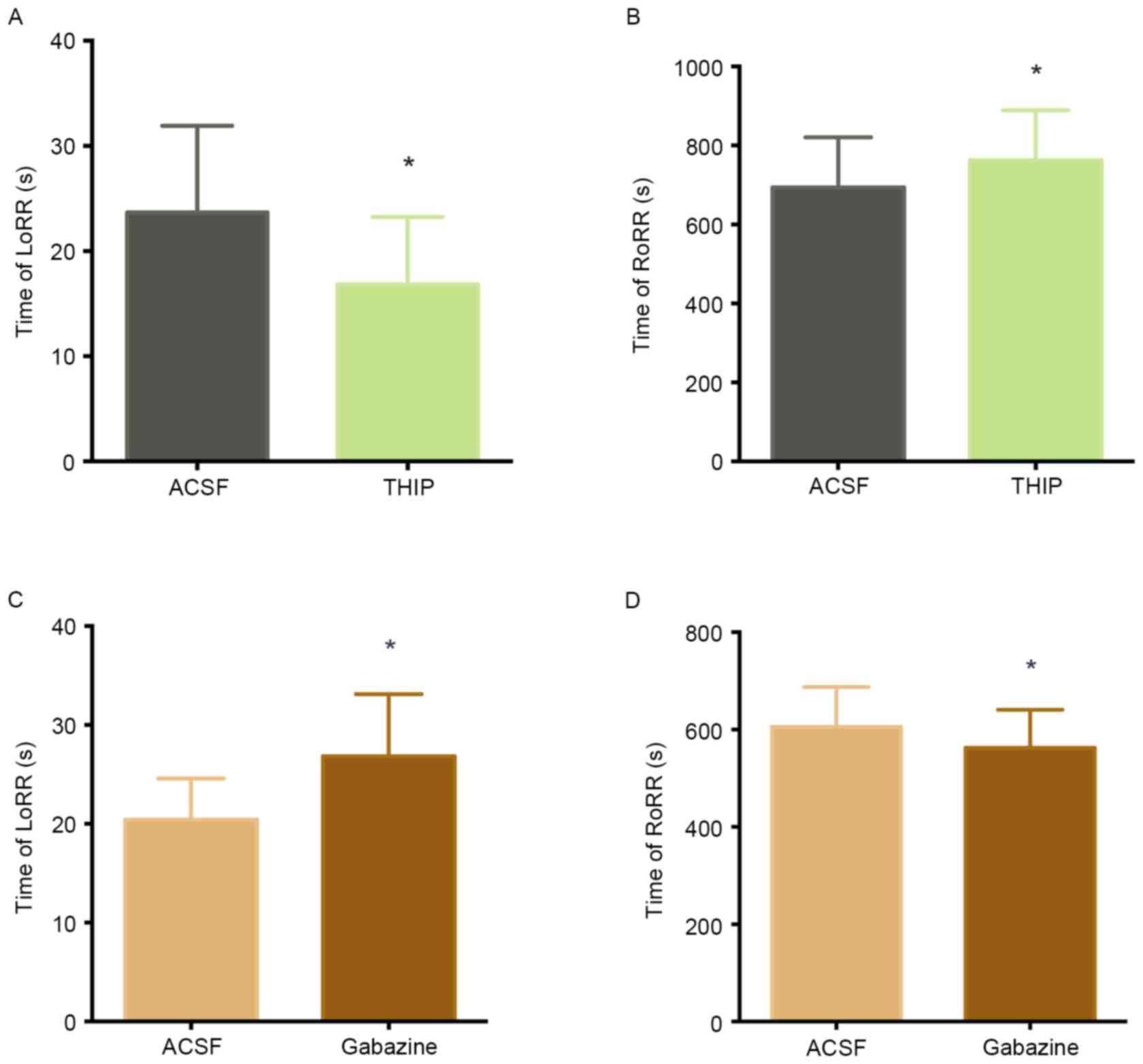
observed the LoRR and RoRR. Fig. 4A
and B illustrates how ACSF and THIP microinjection into the
VLPO of the same rats (n=12) affected propofol-mediated anesthesia
and subsequent recovery. Microinjection of THIP into the VLPO
decreased LoRR (23.67±8.24 sec to 16.83±6.41 sec, P<0.05)
(Fig. 4A) and increased RoRR
(694.00±126.73 sec to 762.83±126.28 sec, P<0.05) (Fig. 4B) before propofol administration.
(Fig. 4C and D) illustrates the
effects of ACSF and the GABAA antagonist gabazine on
propofol-mediated anesthesia and recovery in the same rats (n=12).
Microinjection of gabazine into the VLPO increased LoRR (20.42±4.14
sec to 26.83±6.28 sec, P<0.05) (Fig. 4C) and decreased RoRR (605.67±81.79
sec to 562.58±78.05 sec, P<0.05, Fig. 4D) before propofol
administration.
The effect of propofol on the
spontaneous firing activity of LC neurons in rats with VLPO
lesion
To explore the effect of propofol on LC firing
activity, we anesthetized the VLPO sham-lesion and VLPO lesion rats
with different concentrations of propofol. Fig. 5 shows that propofol inhibited the
spontaneous firing activity of LC neurons in a dose-dependent
manner. In the VLPO sham-lesion group, the spontaneous firing
activity of LC neurons in medium propofol concentration (40
mg/kg/h) was reduced to 2.98±0.6 Hz compared with 6.54±1.16 Hz of
the low propofol concentration (20 mg/kg/h) group (P<0.05). The
spontaneous firing activity of LC neurons in high propofol
concentration (60 mg/kg/h) was reduced to 1.36±0.9 Hz compared with
2.98±0.6 Hz of the medium propofol concentration (40 mg/kg/h) group
(P<0.05). The firing activity of LC neurons in medium propofol
concentration was reduced to 2.98±0.6 Hz compared with 7.32±1.33 Hz
of the consciousness group (P<0.05). And the firing activity of
LC neurons in high propofol concentration was reduced to 1.36±0.9
Hz compared with 7.32±1.33 Hz of the consciousness group
(P<0.05). However, there's no statistical significant
differences in the firing activity between consciousness and low
propofol concentration group, (Fig.
5A).
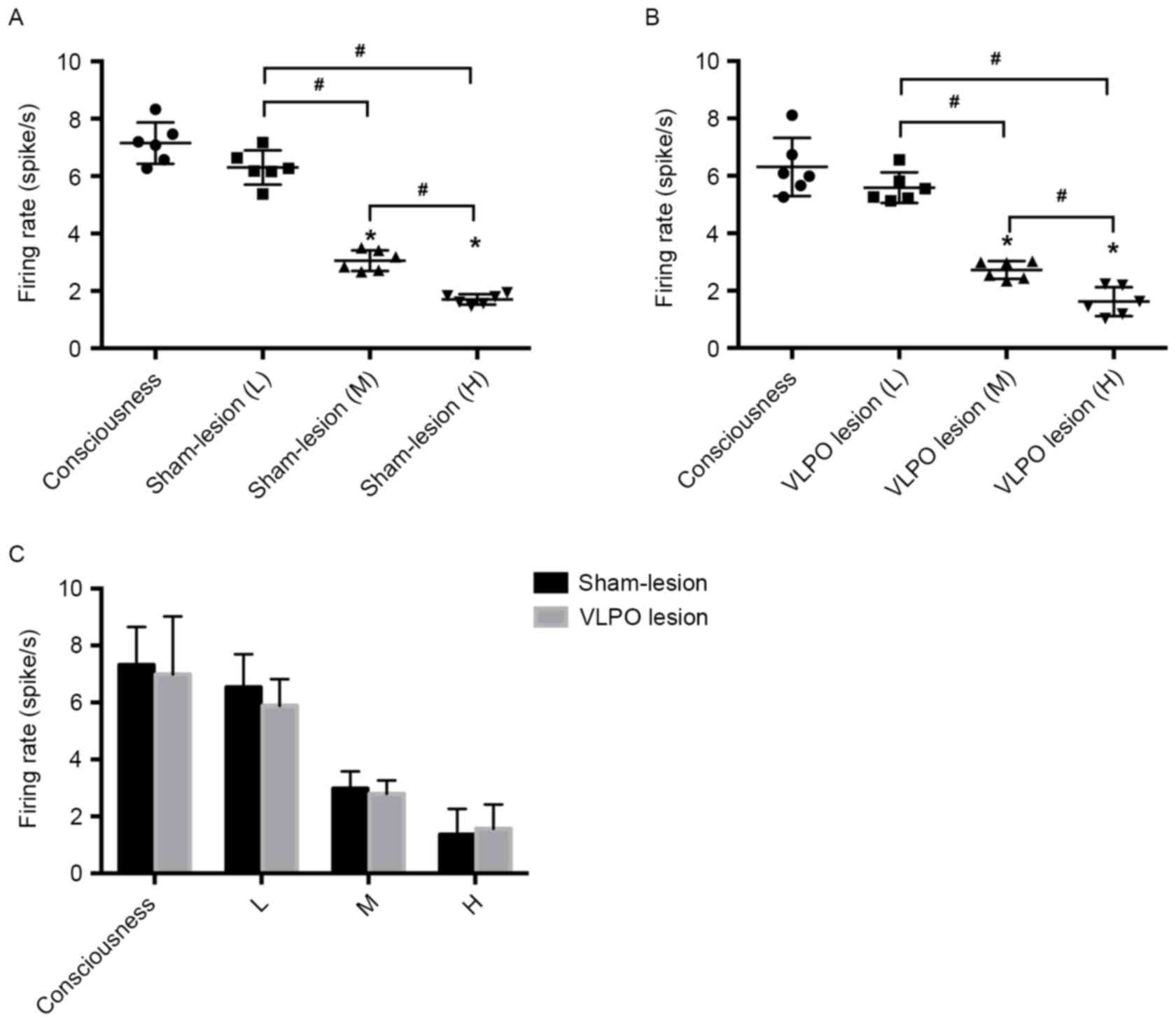 | Figure 5.Effect of different propofol
concentration on firing rate of LC noradrenergic neurons in VLPO
sham-lesion and lesion rats. (A) Shows the low (20 mg/kg/h, i.v.),
medium (40 mg/kg/h, i.v.), high (60 mg/kg/h, i.v.) propofol
concentration administration and without propofol administration on
LC noradrenergic neurons firing activity in VLPO sham-lesion rats.
(B) Shows the low (20 mg/kg/h, i.v.), medium (40 mg/kg/h, i.v.),
high (60 mg/kg/h, i.v.) propofol concentration administration and
without propofol administration on LC noradrenergic neurons firing
activity in VLPO lesion rats. (C) Shows different propofol
concentration administration and without propofol administration on
LC noradrenergic neurons firing activity in VLPO sham-lesion and
lesion rats. #P<0.05 high propofol concentration
group vs. medium propofol concentration group, n=6; medium propofol
concentration group vs. low propofol concentration group, n=6.
*P<0.05 medium propofol concentration vs. consciousness group,
n=6; high propofol concentration vs. consciousness group, n=6. L,
low propofol concentration administration (20 mg/kg/h, i.v.); M,
medium propofol concentration administration (40 mg/kg/h, i.v.); H,
high propofol concentration administration (60 mg/kg/h, i.v.). |
The similar inhibitory effect was observed in the
VLPO lesion group. In this group, the spontaneous firing activity
of LC neurons in medium propofol concentration rats was 2.80±0.46
Hz compared with 5.89±0.93 Hz in the low propofol concentration
group (P<0.05). The spontaneous firing activity of LC neurons in
high propofol concentration rats was 1.57±0.85 Hz compared with
2.80±0.46 Hz in the medium propofol concentration group
(P<0.05). The firing activity of LC neurons in medium propofol
concentration was reduced to 2.80±0.46 Hz compared with 6.99±2.03
Hz of the consciousness group (P<0.05). And the firing activity
of LC neurons in high propofol concentration was reduced to
1.57±0.85 Hz compared with 6.99±2.03 Hz of the consciousness group
(P<0.05). In addition, there's no statistical significant
differences in the firing activity between consciousness and low
propofol concentration group, (Fig.
5B). Nevertheless, our experimental results showed that firing
activities were not significantly different between sham-lesion and
VLPO lesion rats after administration of the same concentration of
propofol (P>0.05), (Fig.
5C).
Discussion
When we microinjected the GABAA agonist
THIP into the VLPO before propofol administration, results showed
that LoRR decreased while RoRR increased. In contrast,
microinjection of the GABAA antagonist gabazine into the
VLPO increased LoRR and decreased RoRR. The results indicate that
GABAA receptors in the VLPO are involved in
propofol-induced unconsciousness, which is consistent with previous
reports demonstrating that potentiating GABAergic transmission in
the VLPO promotes sleep (15,24,25).
The VLPO was the first nucleus to be identified as a
sleep-regulating center containing GABAergic neurons (26). Two-thirds of VLPO neurons are
inhibited by the wake-promoting neurotransmitter noradrenaline
(NA). These are putative sleep-promoting neurons and can be
activated by general anesthetics. The remaining one-third of
neurons can be excited by NA and inhibited by general anesthetics
(14). Patch clamp experiments
(27) have shown that propofol
directly activates NA(−) neurons via GABAA receptors.
THIP is a specific GABAA receptor agonist (28) and microinjection of THIP into the
pontine reticular formation promotes wakefulness (29). On the other hand, microinjection of
THIP into the VLPO improves sleep (30).
Our work suggests that early activation of VLPO
GABAergic neurons enhances the anesthetic effect of propofol. This
effect may be the result of more GABA releasing into the arousal
system in response to THIP. Nelson (12) demonstrated that the sedative
effects of GABA-responsive agents were attenuated when gabazine was
microinjected into the TMN nucleus. In our study, the gabazine
microinjection significantly increased LoRR and decreased RoRR.
After the GABAA receptor was blocked, propofol-induced
anesthesia was attenuated. In conclusion, potentiating GABAergic
transmission in the VLPO may represent a mechanism for
propofol-induced anesthesia. In contrast, blocking GABAergic
transmission may attenuate unconsciousness following propofol
administration.
In the present study, we also found that the firing
rate of LC neurons was dramatically decreased when the
concentration of propofol was higher. It was reported that the
persistent inward calcium current and the cAMP-activated inward
sodium current formed the spontaneous firing activity of LC neurons
(31). Previously in vitro
patch clamp experiments demonstrated that propofol inhibited the LC
neurons by activation of outward chloride current, and the
GABAA (rather than GABAB) receptor
antagonists could block the inhibitory effect (32). Accordingly, in our in vivo
extracellular single-unit recordings experiments, we hypothesize
that outward chloride current activated by propofol can balance the
inward calcium current and the cAMP-activated sodium current. Thus,
the spontaneous firing activity of LC will be inhibited.
Furthermore, higher propofol concentration could increase the
outward chloride current as to hyperpolarize the LC neurons.
The LC neurons fire continually during consciousness
and the firing rate decreases during non-rapid eye movement (NREM)
and rapid eye movement (REM) sleep (33). Halothane is an agent that could
suppress such activity of LC neurons (34). Moreover, α2-adrenergeic receptors
in the LC are a major target for the sedative agent dexmedetomidine
(35), which directly inhibits LC
neurons. Our data showed that the firing rate of LC neurons was not
influenced by VLPO lesions, indicating that VLPO GABAergic neurons
were not involved in the propofol-mediated inhibition on LC
neurons.
VLPO lesions could induce acute sleep loss in rats
(16,36) and confer a short-term resistance to
isoflurane (15). Our previous
work (37) conducted acute lesions
in VLPO culminated in opposite results, which indicated that VLPO
is necessary for the propofol-induced inhibition of LC activity.
Another thing to note is that sleep debt caused by sleep loss can
amplify the sensitivity of propofol and isoflurane (38). In order to minimize the effect of
sleep debt, we performed experiments seven days after the lesions.
VLPO lesions did not affect the firing rate of LC neurons after
propofol was administered. This could also be explained by the
presence of different sleep-promoting neuron populations in the
median preoptic nucleus (MNPO) (39), basal forebrain (BF) (40) and cortex (41). If one region is experimentally
lesioned, other sleep-promoting systems remain intact. Thus, LC
neurons also received inhibitory afferent projections from these
nuclei, which may have compensated for the loss of VLPO GABAergic
neurons in this study.
However, this study is limited in the following
aspects. We aim to investigate the role NA(−) neurons play in
propofol-induced anesthesia. Nevertheless, NA(+) neurons in the
VLPO were also lesioned by ibotenic acid. Specific inactivation of
NA(−) neurons by pharmacogenetics or genetic manipulation would be
a better approach to conducting our study. Additionally,
quantifying the changes of neurotransmitters GABA in the LC area
would make our conclusion more persuasive.
In conclusion, we identified that VLPO GABAergic
neurons play a crucial role in propofol-induced unconsciousness.
Propofol could suppress the spontaneous firing activity of LC
noradrenergic neurons in vivo, but VLPO neurons are not
involved in propofol-mediated inhibition on LC neurons. Present
findings are consistent with the hypothesis that general
anesthetics act on endogenous sleep-wake circuitry to exert
sedative effects.
Acknowledgements
This research was supported by grants from the
National Natural Science Foundation of China (NO. 81460219). All
work was conducted in the Guizhou Key Laboratory of Anesthesia and
Organ Protection, Zunyi, China. JY, YZ and BF contributed to the
conception and design of the work. JY, ZL, and YZ performed the
behavioral assays. SC, BF, and JY performed histological staining
and analysis. JY, LZ, ZW and YZ were responsible for the in
vivo recording. JY analyzed the data and drafted the
manuscript. All authors approved the final version of the
manuscript.
References
|
1
|
Brown EN, Lydic R and Schiff ND: General
anesthesia, sleep, and coma. N Engl J Med. 363:2638–2650. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Franks NP: Molecular targets underlying
general anaesthesia. Br J Pharmacol. 147:(Suppl 1). S72–S81. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yasui Y, Masaki E and Kato F: Sevoflurane
directly excites locus coeruleus neurons of rats. Anesthesiology.
107:992–1002. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ishizawa Y, Pidikiti R, Liebman PA and
Eckenhoff RG: G protein-coupled receptors as direct targets of
inhaled anesthetics. Mol Pharmacol. 61:945–952. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Braun AR, Balkin TJ, Wesenten NJ, Carson
RE, Varga M, Baldwin P, Selbie S, Belenky G and Herscovitch P:
Regional cerebral blood flow throughout the sleep-wake cycle. An
H2(15)O PET study. Brain. 120:1173–1197. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kajimura N, Uchiyama M, Takayama Y, Uchida
S, Uema T, Kato M, Sekimoto M, Watanabe T, Nakajima T, Horikoshi S,
et al: Activity of midbrain reticular formation and neocortex
during the progression of human non-rapid eye movement sleep. J
Neurosci. 19:10065–10073. 1999.PubMed/NCBI
|
|
7
|
Boly M, Moran R, Murphy M, Boveroux P,
Bruno MA, Noirhomme Q, Ledoux D, Bonhomme V, Brichant JF, Tononi G,
et al: Connectivity changes underlying spectral EEG changes during
propofol-induced loss of consciousness. J Neurosci. 32:7082–7090.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Voss L and Sleigh J: Monitoring
consciousness: The current status of EEG-based depth of anaesthesia
monitors. Best Pract Res Clin Anaesthesiol. 21:313–325. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Franks NP: General anaesthesia: From
molecular targets to neuronal pathways of sleep and arousal. Nat
Rev Neurosci. 9:370–386. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saper CB, Scammell TE and Lu J:
Hypothalamic regulation of sleep and circadian rhythms. Nature.
437:1257–1263. 2015. View Article : Google Scholar
|
|
11
|
Saper CB, Fuller PM, Pedersen NP, Lu J and
Scammell TE: Sleep state switching. Neuron. 68:1023–1042. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nelson LE, Guo TZ, Lu J, Saper CB, Franks
NP and Maze M: The sedative component of anesthesia is mediated by
GABA(A) receptors in an endogenous sleep pathway. Nat Neurosci.
5:979–984. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu J, Nelson LE, Franks N, Maze M,
Chamberlin NL and Saper CB: Role of endogenous sleep-wake and
analgesic systems in anesthesia. J Comp Neurol. 508:648–662. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li KY, Guan YZ, Krnjević K and Ye JH:
Propofol facilitates glutamatergic transmission to neurons of the
ventrolateral preoptic nucleus. Anesthesiology. 111:1271–1278.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Moore JT, Chen J, Han B, Meng QC, Veasey
SC, Beck SG and Kelz MB: Direct activation of sleep-promoting VLPO
neurons by volatile anesthetics contributes to anesthetic hypnosis.
Curr Biol. 22:2008–2016. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu J, Greco MA, Shiromani P and Saper CB:
Effect of lesions of the ventrolateral preoptic nucleus on NREM and
REM sleep. J Neurosci. 20:3830–3842. 2000.PubMed/NCBI
|
|
17
|
Zhang J, Yin D, Wu F, Zhang G, Jiang C, Li
Z, Wang L and Wang K: Microinjection of adenosine into the
hypothalamic ventrolateral preoptic area enhances wakefulness via
the A1 receptor in rats. Neurochem Res. 38:1616–1623. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sukhotinsky I, Zalkind V, Lu J, Hopkins
DA, Saper CB and Devor M: Neural pathways associated with loss of
consciousness caused by intracerebral microinjection of GABA
A-active anesthetics. Eur J Neurosci. 25:1417–1436. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tung A, Herrera S, Szafran MJ, Kasza K and
Mendelson WB: Effect of sleep deprivation on righting reflex in the
rat is partially reversed by administration of adenosine A1 and A2
receptor antagonists. Anesthesiology. 102:1158–1164. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vanini G, Watson CJ, Lydic R and Baghdoyan
HA: Gamma-aminobutyric acid-mediated neurotransmission in the
pontine reticular formation modulates hypnosis, immobility, and
breathing during isoflurane anesthesia. Anesthesiology.
109:978–988. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takahashi K, Kayama Y, Lin JS and Sakai K:
Locus coeruleus neuronal activity during the sleep-waking cycle in
mice. Neuroscience. 169:1115–1126. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sugiyama D, Hur SW, Pickering AE, Kase D,
Kim SJ, Kawamata M, Imoto K and Furue H: In vivo patch-clamp
recording from locus coeruleus neurones in the rat brainstem. J
Physiol. 590:2225–2231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang T, Zhang QJ, Liu J, Wu ZH and Wang S:
Firing activity of locus coeruleus noradrenergic neurons increases
in a rodent model of Parkinsonism. Neurosci Bull. 25:15–20. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lydic R and Baghdoyan HA: Sleep,
anesthesiology, and the neurobiology of arousal state control.
Anesthesiology. 103:1268–1295. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiong M, Li J, Wang D, Delphin E and Ye
JH: Intra-ventrolateral preoptic nucleus injection of
γ-aminobutyric acid induces sedation in rats. Int J Physiol
Pathophysiol Pharmacol. 4:94–98. 2012.PubMed/NCBI
|
|
26
|
Sherin JE, Shiromani PJ, McCarley RW and
Saper CB: Activation of ventrolateral preoptic neurons during
sleep. Science. 271:216–219. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu YW, Zuo W and Ye JH: Propofol
stimulates noradrenalin-inhibited neurons in the ventrolateral
preoptic nucleus by reducing GABAergic inhibition. Anesth Analg.
117:358–363. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Boehm SL II, Homanics GE, Blednov YA and
Harris RA: delta-Subunit containing GABAA receptor knockout mice
are less sensitive to the actions of
4,5,6,7-tetrahydroisoxazolo-[5,4-c]pyridin-3-ol. Eur J Pharmacol.
541:158–162. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vanini G and Baghdoyan HA: Extrasynaptic
GABAA receptors in rat pontine reticular formation increase
wakefulness. Sleep. 36:337–343. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu J and Greco MA: Sleep circuitry and the
hypnotic mechanism of GABAA drugs. J Clin Sleep Med. 2:(Suppl).
S19–S26. 2006.PubMed/NCBI
|
|
31
|
Williams JT and North RA: Opiate-receptor
interactions on single locus coeruleus neurones. Mol Pharmacol.
26:489–497. 1984.PubMed/NCBI
|
|
32
|
Chen CL, Yang YR and Chiu TH: Activation
of rat locus coeruleus neuron GABA(A) receptors by propofol and its
potentiation by pentobarbital or alphaxalone. Eur J Pharmacol.
386:201–210. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Berridge CW and Waterhouse BD: The locus
coeruleus-noradrenergic system: Modulation of behavioral state and
state-dependent cognitive processes. Brain Res Brain Res Rev.
42:33–84. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sirois JE, Lei Q, Talley EM, Lynch C III
and Bayliss DA: The TASK-1 two-pore domain K+ channel is a
molecular substrate for neuronal effects of inhalation anesthetics.
J Neurosci. 20:6347–6354. 2000.PubMed/NCBI
|
|
35
|
Correa-Sales C, Rabin BC and Maze M: A
hypnotic response to dexmedetomidine, an alpha 2 agonist, is
mediated in the locus coeruleus in rats. Anesthesiology.
76:948–952. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eikermann M, Vetrivelan R, Grosse-Sundrup
M, Henry ME, Hoffmann U, Yokota S, Saper CB and Chamberlin NL: The
ventrolateral preoptic nucleus is not required for isoflurane
general anesthesia. Brain Res. 1426:30–37. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Yu T, Yuan J and Yu BW: The
ventrolateral preoptic nucleus is required for propofol-induced
inhibition of locus coeruleus neuronal activity. Neurol Sci.
36:2177–2184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tung A, Szafran MJ, Bluhm B and Mendelson
WB: Sleep deprivation potentiates the onset and duration of loss of
righting reflex induced by propofol and isoflurane. Anesthesiology.
97:906–911. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Suntsova N, Szymusiak R, Alam MN,
Guzman-Marin R and McGinty D: Sleep-waking discharge patterns of
median preoptic nucleus neurons in rats. J Physiol. 543:665–677.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Szymusiak R and McGinty D: Sleep
suppression following kainic acid-induced lesions of the basal
forebrain. Exp Neurol. 94:598–614. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gerashchenko D, Wisor JP, Burns D, Reh RK,
Shiromani PJ, Sakurai T, de la Iglesia HO and Kilduff TS:
Identification of a population of sleep-active cerebral cortex
neurons. Proc Natl Acad Sci USA. 105:10227–10232. 2008. View Article : Google Scholar : PubMed/NCBI
|