Introduction
Glioma, which is the most commonly occurring highly
malignant primary brain tumor, comprises approximately one-third of
intrinsic tumors of the central nervous system in both children and
adults (1,2). Although major advancements have been
made by integrating neurosurgery, radiotherapy and chemotherapy to
control malignant gliomas, poor prognosis and survival rate remains
characteristic of such tumors (3,4).
Therefore, it is important to further understand the molecular
mechanisms in glioma progression, and develop novel
therapeutics.
MicroRNAs (miRNAs/miRs) are 21–23 nucleotide, small
non-protein coding single-stranded RNA molecules, which are the
important post-transcriptional regulators of gene expression in
animals and plants (5). In
animals, the regulatory effects of miRNAs are achieved by binding
the complimentary sequences within the 3′-untranslated regions
(UTRs) of their cognate messenger RNA (mRNA) targets (6,7).
miRNAs are involved in a variety of tumorigenic activities,
including angiogenesis, proliferation, invasion and metastasis
(8). Previous studies have
suggested that the expression levels of many miRNAs are involved in
glioma (9,10). miR-613 has been identified to serve
a key role in tumorigenesis and cancer progression, such as
inhibition of ovarian cancer proliferation and invasion (11) and induction of lung cancer cell
cycle arrest (12). However, the
effect of miR-613 in glioma remains unknown.
The present study demonstrated that the mRNA
expression level of miR-613 was decreased in glioma cell lines, and
overexpression of miR-613 in both U87 and U251 cells suppressed
invasion, proliferation and angiogenesis. In addition, vascular
endothelial growth factor A (VEGFA) was identified to be a target
gene of miR-613. The effects of miR-613 on tumor angiogenesis and
proliferation were dependent on downregulation of VEGFA. These
findings demonstrated the underlying mechanism and a novel role for
miR-613 as a tumor inhibitor in glioma.
Materials and methods
Cell lines and patient samples
The NHA, H4, U87, U251 and SWO-38 human glioma cell
lines and the HUVEC human endothelial cell line were purchased from
the Institute of Biochemistry and Cell Biology of the Chinese
Academy of Sciences (Shanghai, China). All cells were cultured in
RPMI-1640 medium supplemented with penicillin and streptomycin (100
U/ml and 100 mg/ml, respectively) and 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 37°C in
a humidified atmosphere of 5% CO2.
In the present study, 29 tissue samples were
isolated from patients with stage I–II glioma and 24 samples from
patients with stage III–IV glioma. In addition, 29 samples from
normal adjacent tissue were also collected. The tissues were
collected in the First Affiliated Hospital of Kunming Medical
University (Kunming, China) between May 2014 and March 2016.
Informed consent was obtained from each patient. All tissue samples
had an affirmative pathological diagnosis and were classified on
the basis of the World Health Organization criteria. None of the
patients had received preoperative treatment, including
chemotherapy or radiotherapy. A total of 32 male and 21 female
patients were enrolled in the present study and the mean age of the
patients was 69 years.
Lentivirus (LV) production and
transfection
The LV-miR-613 mimics and mimic controls were
purchased from Shanghai GeneChem Co., Ltd. (Shanghai, China). Cells
were plated in 6-well plates at a density of 1×105
cells/well and were transfected with 200 µl (100 nM) miR-613 mimics
or mimic controls using Lipofectamine 2000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The transfected cells were cultured in RPMI-1640 medium
supplemented with 10% FBS for 48 h at 37°C in a humidified
atmosphere of 5% CO2. The sequences of the
oligonucleotides that were used were as follows: miR-613 mimic
sense, 5′-AGGAAUGUUCCUUCUUUGCC-3′ and antisense,
5′-UGGCAAAGAAGGAACAUUCCUUG-3′; negative control (NC) sequence
sense, 5′-ACUACUGAGUGACAGUAGA-3′ and antisense,
5′-AUCUACUGUCACUCAGUAGUGG-3′.
Plasmid construction and dual
luciferase activity assay
TargetScan (http://targetscan.org/) and MiRanda (http://www.microrna.org/) software were used to
predict the potential target genes of miR-613, and VEGFA was
identified as a possible target. The eukaryotic expression vector
pcDNA3.1 (+) was subcloned with full-length VEGFA cDNA (Invitrogen;
Thermo Fisher Scientific, Inc.). The VEGFA 3′-UTR target site for
miR-613 was amplified by polymerase chain reaction (PCR) and cloned
into the XbaI site of a pGL3 control vector (Promega Corp.,
Madison, WI, USA). This vector was termed wild-type (WT) VEGFA
3′-UTR. The Quick-Change Mutagenesis kit (Agilent Technologies,
Inc., Santa Clara, CA, USA) was used to carry out the site-directed
mutagenesis of the miR-613 target-site in the VEGFA 3′UTR, termed
mutant (Mut) VEGFA 3′-UTR. For the luciferase activity assay, U87
cells were seeded at density of 5×104 cells/well in
24-well plates and co-transfected with WT or Mut VEGFA 3′UTR
vectors and the control vector pRL-(cytomegalovirus) CMV coding for
Renilla luciferase, (Promega Corp.) using Lipofectamine 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. A Dual-Luciferase Reporter Assay
system (Promega Corp.) was used to detect the luciferase activity
after transfection for 36 h.
RNA extraction and reverse
transcription-quantitative (RTq)-PCR
Total RNA was extracted with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The mRNA expression level of miR-613 was
detected using an ABI PRISM 7500 Sequence Detection system (ABI)
using TaqMan MicroRNA assay kits (Applied Biosystems, Foster City,
CA, USA) according to the manufacturer's protocol. The
thermocycling conditions that were used were as follows: Initial
denaturation at 95°C for 10 min, followed by 40 cycles at 95°C for
15 sec and at 60°C for 60 sec. U6 served as the control for
normalization.
The gene expression of VEGFA was also detected.
Total RNA was reverse transcribed into cDNA using Revert Aid First
Strand cDNA Synthesis kit (Thermo Fisher Scientific Inc.) according
to the manufacturer's protocol. The temperature conditions used
were as follows: At 25°C for 5 min, at 42°C for 60 min and at 70°C
for 10 min. qPCR was performed on cDNA using SYBR-Green (Takara
Bio, Inc., Otsu, Japan) and gene expression was normalized to
β-actin. Thermocycling conditions were as follows: At 95°C for 30
sec, followed by 40 cycles at 95°C for 5 sec and at 60°C for 34
sec. The primers that were used in the present study were as
follows: VEGFA forward, 5′-ATCCAATCGAGACCCTGGTG-3′ and reverse,
5′-ATCTCTCCTATGTGCTGGCC-3′; β-actin forward,
5′-TGAGAGGGAAATCGTGCGTGAC-3′ and reverse,
5′-GCTCGTTGCCAATAGTGATGACC-3′; miR-613 forward,
5′-AGGAATGTTCCTTCT-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′; and U6
forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. The specificity of primer sequences
were detected by its dissociation curve, and the 2−ΔΔCq
(quantitation threshold) method was used to calculate the relative
gene expression levels (13).
Western blot analysis
Western blot assay was conducted as previously
described (14). Briefly, cells
were lysed with ice-cold lysis buffer (Cell Signaling Technology,
Inc., Danvers, MA, USA). The concentrations of proteins were
measured using a bicinchoninic acid protein assay kit (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Total proteins (30 µg) were separated by 10% SDS-PAGE, transferred
onto polyvinylidene difluoride membranes (EMD Millipore, Billerica,
MA, USA) and blocked in 5% skim milk (BD Biosciences, Franklin
Lakes, NJ, USA) for 2 h at room temperature. Membranes were then
incubated with the following primary antibodies at 4°C overnight:
Anti-VEGFA (cat no. ab1316; 1:1,000; Abcam, Cambridge, UK),
anti-angiopoietin-2 (Ang-2; cat no. sc-20718; 1:50), anti-CD31
(cat. no. sc-71872; 1:200) (both from Santa Cruz Biotechnology,
Inc., Dallas, TX, USA) and anti-GAPDH (cat no. ab9484; 1:5,000;
Abcam). Subsequently, they were incubated with the following
horseradish peroxidase (HRP)-conjugated secondary antibodies for 1
h at room temperature: Anti-mouse immunoglobulin (Ig) G (cat no.
sc-2005; 1:2,000; Santa Cruz Biotechnology, Inc.) and anti-rabbit
IgG (cat no. sc-2004; 1:2,000; Santa Cruz Biotechnology, Inc.).
Enhanced chemiluminescence (GE Healthcare, Chicago, IL, USA) was
used to detect the expression levels of the target proteins. Blots
were semi-quantified using Image Lab software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Immunohistochemistry
According to the manufacturer's protocol,
immunohistochemistry for VEGFA and CD31 was performed on 4-µm
formalin fixed, paraffin-embedded tissue sections. The sections
were incubated with anti-VEGFA (1:200) and anti-CD31 (1:50) primary
antibodies overnight at 4°C. The sections were then incubated for 1
h at room temperature with HRP-conjugated anti-mouse secondary
antibody (cat. no. sc-2005; 1:200; Santa Cruz Biotechnology, Inc.).
Immunohistochemical stains were completed using an automated
immunostainer (Ventana Medical Systems Inc., Tucson, AZ, USA) and
stained sections were observed under a light microscope.
Cell proliferation and tube formation
assay
For the colony formation test, U87 and U251 cells
(1,000 cells/well) were treated with miRNA control, miR-613 mimc,
or miR-613 mimic and pcDNA3.1-VEGFA, seeded into 6-well plates and
cultured for 14 days. The colonies (>50 cells; diameter, 60–100
µm) were stained with 0.1% methylene blue for 5 min at room
temperature and imaged using an optical microscope. The tube
formation assay was conducted as previously described (14), in 24-well plates using growth
factor reduced Matrigel (BD Biosciences). Cells (2×104
cells/well) were resuspended in serum-free medium and plated on the
Matrigel-coated plates. Following incubation at 37°C overnight,
each well was observed under a light microscope. Tubules in each
field were imaged and an average of tubules from 3–5 random fields
of view/well was counted.
Cell invasion assay
The cell invasion assay was conducted as previously
described (5) using
Matrigel-coated Transwell inserts (Costar; Corning Incorporated,
Corning, NY, USA). Briefly, 50 µl (2.0 mg/ml) Matrigel (BD
Biosciences) was added in the upper chambers of the Transwell
inserts for 30 min at 37°C. Subsequently, 3×103 cells in
RPMI-1640 medium without serum were seeded in the upper chambers of
the inserts. In the lower chambers, 600 µl culture medium
supplemented with 10% FBS was added as a chemoattractant. Following
24 h incubation, non-invaded cells were removed using a cotton
swab. Cells that had invaded the lower membrane were fixed with 4%
paraformaldehyde for 30 min at room temperature, stained with 0.1%
crystal violet for 20 min at room temperature, and photographed
under a phase-contrast microscope (Olympus Corp., Tokyo, Japan). A
total of 5 random fields of view were selected for cell
counting.
Animal tumor studies
Animal experiments were approved by the Ethics
Committee of Health and Planning Commission of Qujing City (Qujing,
China). A total of 20 male BALB/c nude mice (age, 5–6 weeks;
weight, 18–22 g) were purchased from Shanghai SLAC Laboratory
Animal Co., Ltd. (Shanghai, China). Mice were housed in a
specific-pathogen-free animal center and fed with autoclaved water
and laboratory rodent chow ad libitum. Mice were maintained
in a controlled temperature (21–23°C) under a 12-h light/dark
cycle, in standard vinyl cages with air filter tops. Cages, bedding
and water containers were autoclaved prior to use. At the beginning
of the experiment, mice were subcutaneously injected in the front
right leg with 0.2 ml U87 cells (2.5×106), which had
been previously transfected with miRNA control or miR-613 mimics
using Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Tumor
sizes were recorded at the indicated time points, and tumor volume
(mm3) was calculated as follows: Tumor volume = 1/2
(tumor length) × (tumor width)2. On day 42
post-injection, mice were sacrificed and the tumor nodules were
dissected and weighed.
Statistical analysis
All data are presented as the mean ± standard
deviation of 3 independent experiments. One-way analysis of
variance or two-tailed Student's t-test were used to analyze the
significant differences between groups. SPSS 18.0 (SPSS, Inc.,
Chicago, IL, USA) and GraphPad Prism 6.0 (National Institutes of
Health, Bethesda, MD, USA) software were used for data analysis.
P<0.05 was considered to indicate a statistically significant
difference.
Results
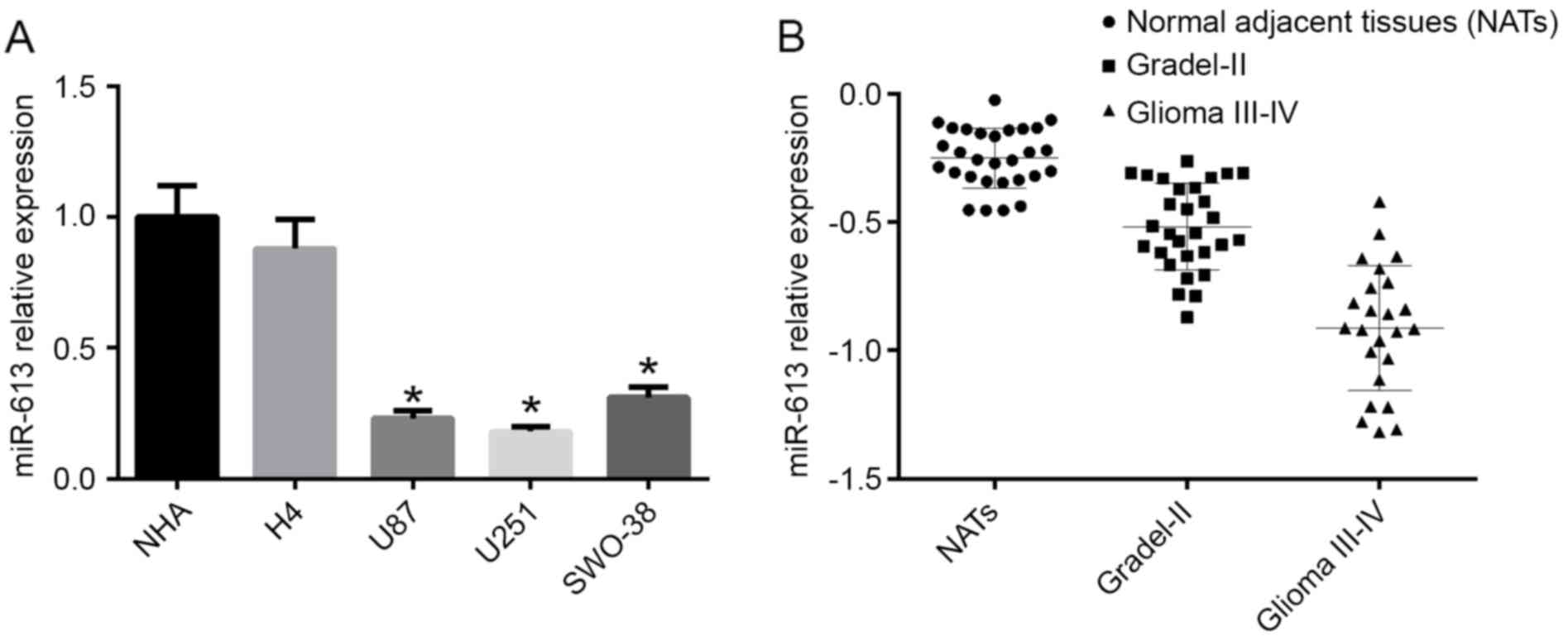
miR-613 expression is downregulated in
glioblastoma multiforme (GBM) cell lines and is associated with the
glioma grade
The expression level of miR-613 was detected in NHA,
H4, U87, U251 and SWO-38 cell lines by RT-qPCR. The results
demonstrated that the mRNA expression level of miR-613 was
decreased in high-grade glioma cell lines (U87, U251 and SWO-38)
compared with NHA cells and the low-grade glioma cell line H4
(P<0.05; Fig. 1A). As U87 and
U251 cells had the lowest expression of miR-613, they were selected
for subsequent experiments. To further confirm the different
expression levels of miR-613 in different grade gliomas, the mRNA
expression level of miR-613 in different grade glioma tissues was
examined by RT-qPCR. As a result, miR-613 levels in glioma I–II
were significantly reduced compared with normal adjacent tissues
(NATS), and in glioma III–IV it was the lowest (Fig. 1B).
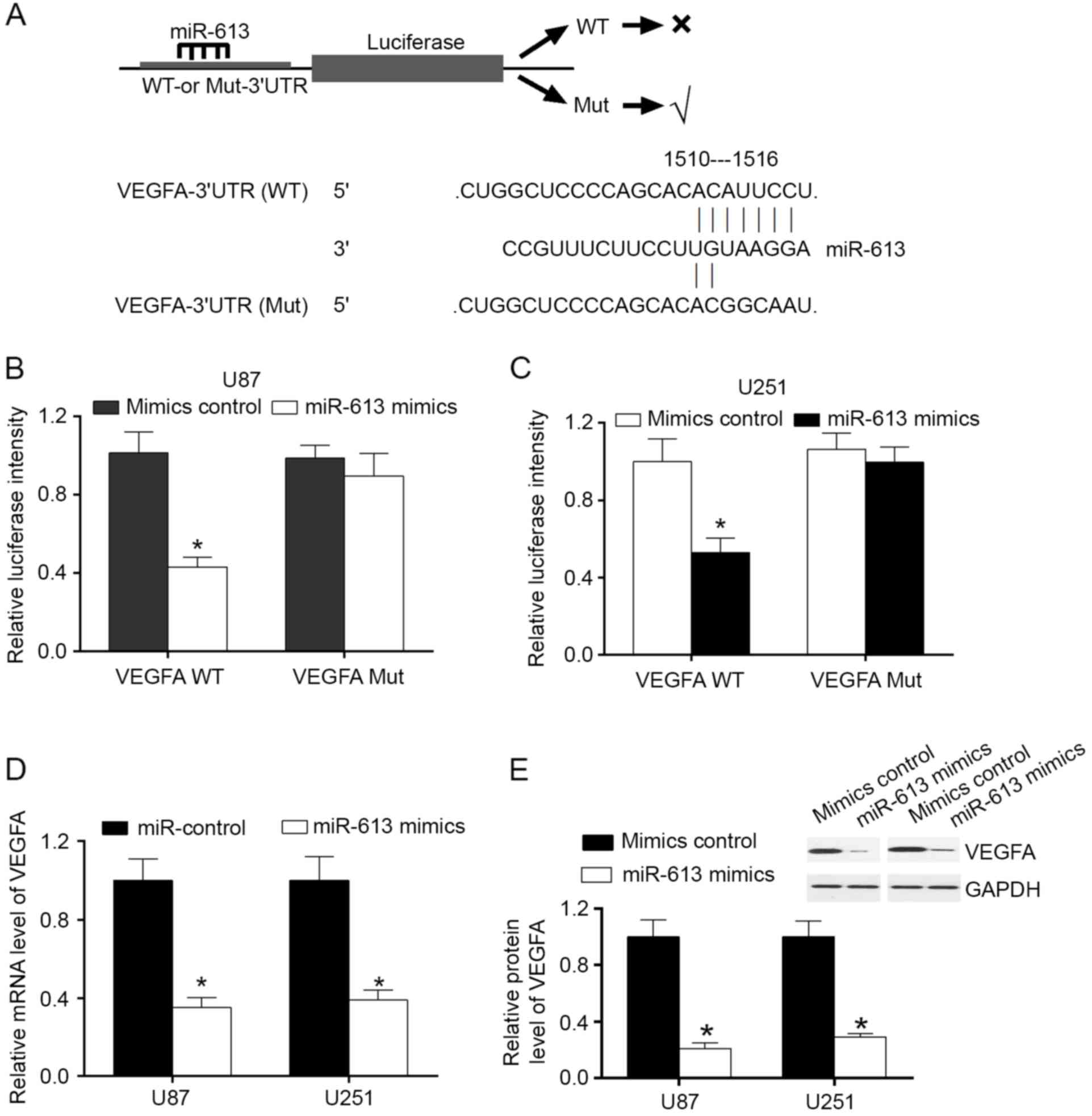
miR-613 serves as a regulator of VEGFA
by attaching to VEGFA 3′-UTR
MiRanda algorithms and TargetScan were used to
search for target genes of miR-613. Among the mRNAs involving
miR-613 recognition sites in their 3′-UTRs, VEGFA was focused on.
In order to confirm that VEGFA was a direct target of miR-613,
VEGFA wild-type (WT) or mutant 3′-UTR was subcloned into a
luciferase reporter vector and co-transfected with miR-613 mimics
or a negative control into U87 and U251 cells (Fig. 2A). The results demonstrated that in
U87 and U251 cell lines, miR-613 significantly inhibited the
luciferase activity of the VEGFA WT 3′-UTR, but had no influence on
the mutant (Fig. 2B and C). To
evaluate the effect of miR-613 on VEGFA expression, U87 and U251
cells were transfected with miR-613 mimics and control. The results
demonstrated that overexpression of miR-613 downregulated the mRNA
and protein expression levels of VEGFA (Fig. 2D and E). These findings illustrated
that VEGFA was a direct downstream target of miR-613 in GBM
cells.
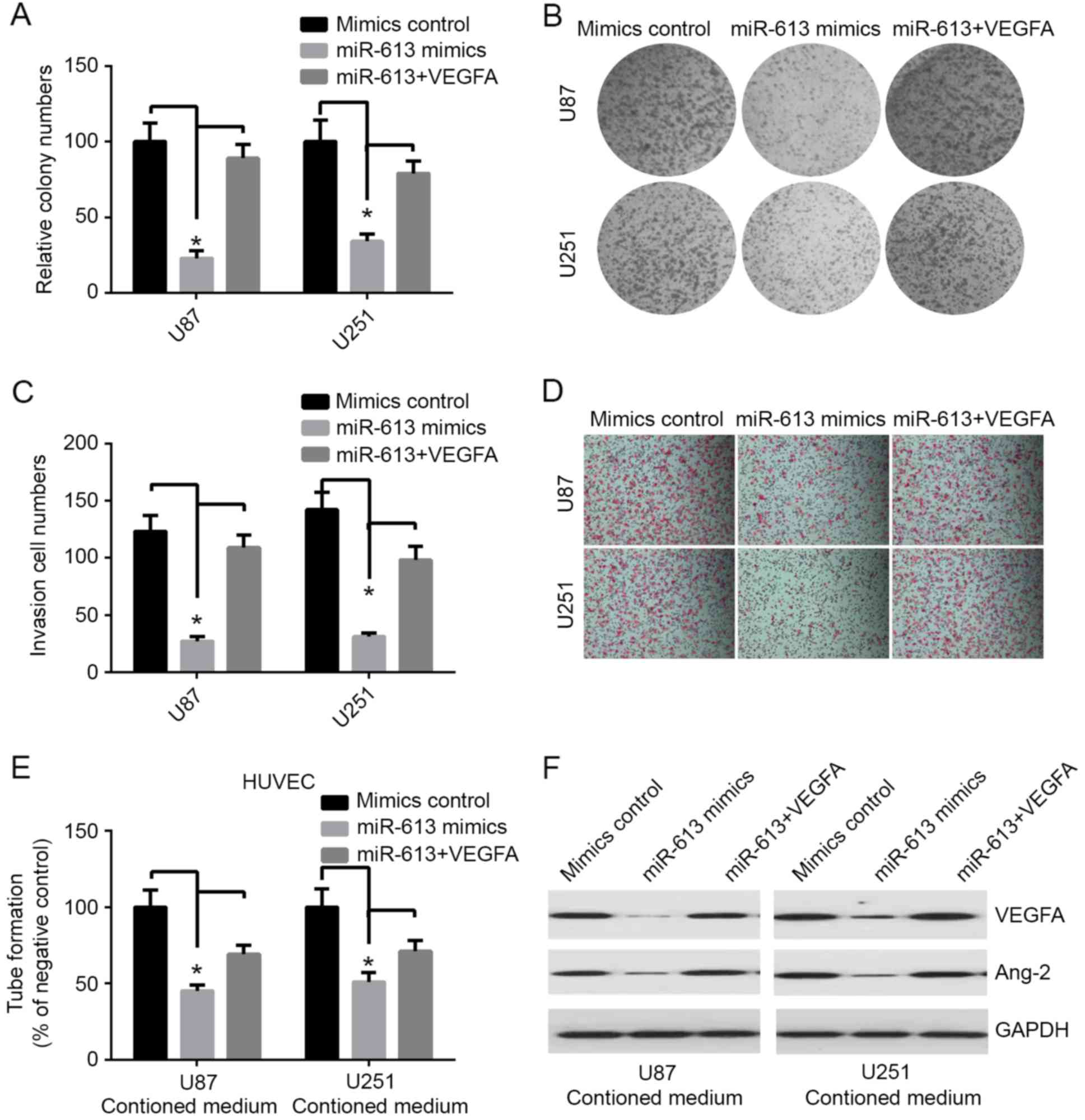
miR-613 suppresses the proliferation,
invasion and angiogenesis of GBM cells via targeting VEGFA in
vitro
VEGFA has been identified to be potentially the most
important mediator in glioma. Previous studies demonstrated that
VEGFA serves a key role in regulating tumor-induced angiogenesis in
glioma (15,16). Studies have indicated that
overexpression of VEGFA was associated with poor prognosis and the
risk for glioma recurrence (17,18).
The expression level of miR-613 was decreased in GBM, and its
inhibitory action on VEGFA was involved with miR-613, which may
have a role in GBM carcinogenesis by downregulating the expression
of VEGFA. Therefore, the effects of miR-613 overexpression and
VEGFA restoration in GBM cells was investigated. In the colony
formation assay, overexpression of miR-613 markedly decreased
colony formation in both U87 and U251 cell lines (Fig. 3A and B). The invasion experiment
results demonstrated that upregulation of miR-613 significantly
inhibited invasion in both GBM cell lines, compared with the mimic
controls (Fig. 3C and D). The tube
formation assay of endothelial HUVEC cells was suppressed by
treatment with medium preconditioned by miR-613-overexpressing U87
and U251 cells (Fig. 3E). In
addition, western blot analysis indicated that restoration of VEGFA
ameliorated the miR-613-induced downregulation of VEGFA (Fig. 3F). The growth and invasion of tumor
depends on tumor angiogenesis, there are various angiogenic factors
regulating this process. Ang-2 is one of the most important
proteins for this process (19).
In this study, the results demonstrated that overexpression of
miR-613 markedly decreased the level of Ang-2 compared with
control, and it was markedly improved by transfection of VEGFA
(Fig. 3F). However, the decrease
in colony formation, invasion and tube formation produced by stable
upregulation of miR-613 was restored by transfecting VEGFA into
both U87 and U251 cell lines (Fig.
3A-E). These results indicated that miR-613 markedly inhibits
invasion, proliferation and angiogenesis, at least partially, via
targeting VEGFA.
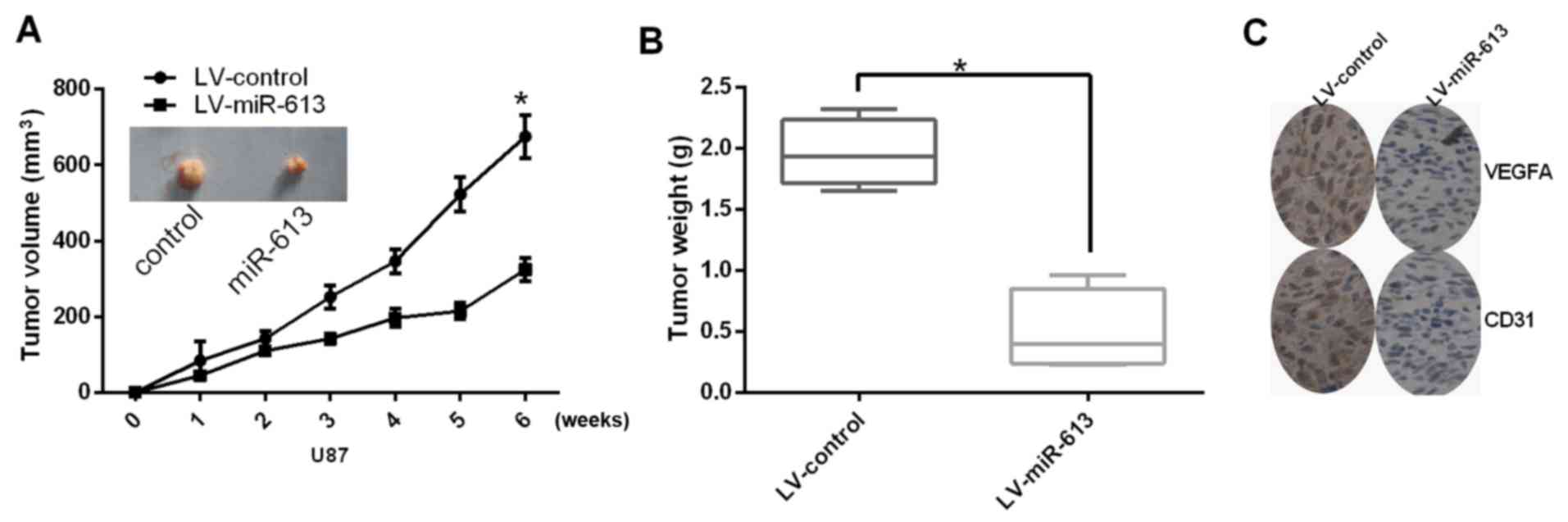
Upregulation of miR-613 inhibits
angiogenesis and growth of GBM cells in vivo
Next, the role of miR-613 on tumor angiogenesis and
growth was studied in vivo. The tumor volume and tumor
weight of subcutaneous xenograft tumors apparently decreased in
nude mice with U87 cells transfected with miR-617, compared with
those stably transfected with LV-control (Fig. 4A and B). Furthermore, the
expression level of VEGFA was also reduced by stable transfection
of miR-613. In addition, miR-613 decreased the protein expression
levels of VEGFA and CD31 (Fig.
4C). In conclusion, these results indicated that miR-613 could
suppress angiogenesis and growth of glioma cells in
vivo.
Discussion
miRNAs serve key roles in many biological processes
including proliferation, differentiation, development and apoptosis
(20). Previous studies have
demonstrated that miRNAs may represent potential therapeutic
targets for a variety of diseases (21,22).
On the whole, there are two ways for developing miRNA-based
therapeutics: miRNA mimics and miRNA antagonists. miRNA antagonists
restrain endogenous miRNAs that exhibit gain-of-function in
diseased tissues (23). miRNA
mimics are used to recover loss-of-function. This approach, also
termed ‘miRNA replacement therapy’, aims to transfect miRNAs into
diseased cells that are regularly expressed in healthy cells
(24). These miRNA-based treatment
methods may be effective on tumor therapy. Although thousands of
miRNAs have been detected in humans (25), only a few of them have been
functionally characterized.
miR-613 was firstly reported to participate in lipid
metabolism in macrophages (26)
and HepG2 cells (27). Previous
studies demonstrated a link between miR-613 and tumorigenesis
(11,28,29).
The present study detected the expression level of miR-613 by
RT-qPCR, and the results demonstrated that the mRNA expression
level of miR-613 was decreased in high grade glioma cell lines
(U87, U251 and SWO-38) compared with NHA cells and the H4 low-grade
glioma cell line. Furthermore, the level of miR-613 was markedly
associated with the grade of glioma, and the mRNA expression level
of miR-613 was decreased in the poor development of glioma. In
order to further investigate the potential therapeutic effect of
overexpression of miR-613 in GBM cells, the role of miR-613 in the
growth of subcutaneous xenograft tumors in nude mice was examined.
Consistent with the result in vitro, restoration of miR-613
inhibited the growth of xenograft tumors. Studies also identified
that upregulation of miR-613 reduced tumor angiogenesis and
invasion in vitro (28,30–32).
These results emphasized the therapeutic value of recovery of
miR-613 expression level in the therapy of tumors.
Angiogenesis serves an important role in tumor
growth and tumor progression. A large number of studies have
demonstrated VEGFA as the most potential adjuster of tumor-induced
angiogenesis in glioma (15,16).
Increased the expression of VEGFA is associated with aggravation of
risk for glioma recurrence, poor prognosis and increased tumor
microvessel density (17,18). In the present study,
miR-613-overexpressing cells were transfected with VEGFA, and
examined the colony forming ability, the invasion ability and tube
formation recovery. Certain direct targets of miR-613 have been
confirmed (26,29). The present study identified a novel
direct target of miR-613 and VEGFA. The results revealed that
miR-613 had the ability to negatively modulate the expression of
VEGFA, which serves important roles by connecting with a site in
the VEGFA 3′-UTR. VEGFA increased the expression level of VEGFA in
miR-613-overexpressing U87 and U251 cells. Ang-2 is another
important protein to regulate tumor angiogenesis (19). The present study demonstrated that
Ang-2 and VEGFA serve synergistic roles in tumor angiogenesis
(33). Therefore, it was concluded
that VEGFA may be a direct target for miR-613, implicating miR-613
as a novel target for glioma therapy.
In the present study, there were many lines of
evidence to confirm the interaction between miR-613 and VEGFA.
Firstly, the 3′-UTR of VEGFA had an assumed binding site of
miR-613. Secondly, miR-613 inhibited the activity of a luciferase
reporter merged with the 3′-UTR of VEGFA mRNA. Furthermore, miR-613
suppressed the protein and mRNA expression levels of VEGFA. These
findings facilitate understanding of the role of VEGFA in
regulating glioma cells.
In conclusion, the results of the present study
suggested that miR-613 is involved in glioma. In addition, it was
demonstrated that miR-613 serves an important role in the
malignancy of glioma cells via inhibition of VEGFA expression. To
the best of our knowledge, this study was the first to assess the
association between miR-613 and VEGFA in glioma. The results
implicate miR-613 as a potential target to treat gliomas in the
future.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Shanxi, China (grant no. 2013011052-2).
References
|
1
|
Sun J, Zheng G, Gu Z and Guo Z: MiR-137
inhibits proliferation and angiogenesis of human glioblastoma cells
by targeting EZH2. J Neurooncol. 122:481–489. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Y, Zhao S and Xu Z: Network and
pathway analysis of microRNAs, transcription factors, target genes
and host genes in human glioma. Oncol Lett. 11:3534–3542.
2016.PubMed/NCBI
|
|
3
|
Lefranc F, Rynkowski M, DeWitte O and Kiss
R: Present and potential future adjuvant issues in high-grade
astrocytic glioma treatment. Adv Tech Stand Neurosurg. 34:3–35.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yue X, Wang P, Xu J, Zhu Y, Sun G, Pang Q
and Tao R: MicroRNA-205 functions as a tumor suppressor in human
glioblastoma cells by targeting VEGF-A. Oncol Rep. 27:1200–1206.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010.PubMed/NCBI
|
|
7
|
Dykxhoorn DM: MicroRNAs and metastasis:
Little RNAs go a long way. Cancer Res. 70:6401–6406. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Di Leva G, Garofalo M and Croce CM:
MicroRNAs in cancer. Annu Rev Pathol. 9:287–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ciafrè SA, Galardi S, Mangiola A, Ferracin
M, Liu CG, Sabatino G, Negrini M, Maira G, Croce CM and Farace MG:
Extensive modulation of a set of microRNAs in primary glioblastoma.
Biochem Biophys Res Commun. 334:1351–1358. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moller HG, Rasmussen AP, Andersen HH,
Johnsen KB, Henriksen M and Duroux M: A systematic review of
microRNA in glioblastoma multiforme: Micro-modulators in the
mesenchymal mode of migration and invasion. Mol Neurobiol.
47:131–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fu X, Cui Y, Yang S, Xu Y and Zhang Z:
MicroRNA-613 inhibited ovarian cancer cell proliferation and
invasion by regulating KRAS. Tumour Biol. 37:6477–6483. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao Q, Li J, Yan J, Liu S, Guo Y, Chen D
and Luo Q: Lycium barbarum polysaccharides ameliorates renal injury
and inflammatory reaction in alloxan-induced diabetic nephropathy
rabbits. Life Sci. 157:82–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Qi M, Li S, Qi T, Mei H, Huang K,
Zheng L and Tong Q: microRNA-9 targets matrix metalloproteinase 14
to inhibit invasion, metastasis, and angiogenesis of neuroblastoma
cells. Mol Cancer Ther. 11:1454–1466. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim KJ, Li B, Winer J, Armanini M, Gillett
N, Phillips HS and Ferrara N: Inhibition of vascular endothelial
growth factor-induced angiogenesis suppresses tumour growth in
vivo. Nature. 362:841–844. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Plate KH, Breier G, Weich HA and Risau W:
Vascular endothelial growth factor is a potential tumour
angiogenesis factor in human gliomas in vivo. Nature. 359:845–848.
1992. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chaudhry IH, O'Donovan DG, Brenchley PE,
Reid H and Roberts IS: Vascular endothelial growth factor
expression correlates with tumour grade and vascularity in gliomas.
Histopathology. 39:409–415. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Varlet P, Guillamo JS, Nataf F, Koziak M,
Beuvon F and Daumas-Duport C: Vascular endothelial growth factor
expression in oligodendrogliomas: A correlative study with
Sainte-Anne malignancy grade, growth fraction and patient survival.
Neuropathol Appl Neurobiol. 26:379–389. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maisonpierre PC, Suri C, Jones PF,
Bartunkova S, Wiegand SJ, Radziejewski C, Compton D, McClain J,
Aldrich TH, Papadopoulos N, et al: Angiopoietin-2, a natural
antagonist for Tie2 that disrupts in vivo angiogenesis. Science.
277:55–60. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang D, Qiu C, Zhang H, Wang J, Cui Q and
Yin Y: Human microRNA oncogenes and tumor suppressors show
significantly different biological patterns: From functions to
targets. PLoS One. 5(pii): e130672010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006;
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kota J, Chivukula RR, O'Donnell KA,
Wentzel EA, Montgomery CL, Hwang HW, Chang TC, Vivekanandan P,
Torbenson M, Clark KR, et al: Therapeutic microRNA delivery
suppresses tumorigenesis in a murine liver cancer model. Cell.
137:1005–1017. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bader AG, Brown D and Winkler M: The
promise of microRNA replacement therapy. Cancer Res. 70:7027–7030.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao XM, Liu KQ, Zhu G, He F, Duval B,
Richer JM, Huang DS, Jiang CJ, Hao JK and Chen L: Identifying
cancer-related microRNAs based on gene expression data.
Bioinformatics. 31:1226–1234. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao R, Feng J and He G: miR-613 regulates
cholesterol efflux by targeting LXRa and ABCA1 in PPARγ activated
THP-1 macrophages. Biochem Biophys Res Commun. 448:329–334. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhong D, Zhang Y, Zeng YJ, Gao M, Wu GZ,
Hu CJ, Huang G and He FT: MicroRNA-613 represses lipogenesis in
HepG2 cells by downregulating LXRa. Lipids Health Dis. 12:322013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ren W, Li C, Duan W, Du S, Yang F, Zhou J
and Xing J: MicroRNA-613 represses prostate cancer cell
proliferation and invasion through targeting Frizzled7. Biochem
Biophys Res Commun. 469:633–638. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li D, Li DQ, Liu D and Tang XJ: MiR-613
induces cell cycle arrest by targeting CDK4 in non-small cell lung
cancer. Cell Oncol (Dordr). 39:139–147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li B, Xie Z, Li Z, Chen S and Li B:
MicroRNA-613 targets FMNL2 and suppresses progression of colorectal
cancer. Am J Transl Res. 8:5475–5484. 2016.PubMed/NCBI
|
|
31
|
Qiu W, Yang Z, Fan Y and Zheng Q:
MicroRNA-613 inhibits cell growth, migration and invasion of
papillary thyroid carcinoma by regulating SphK2. Oncotarget.
7:39907–39915. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu H, Duan P, Zhu H and Rao D: miR-613
inhibits bladder cancer proliferation and migration through
targeting SphK1. Am J Transl Res. 9:1213–1221. 2017.PubMed/NCBI
|
|
33
|
Scheuer W, Thomas M, Hanke P, Sam J, Osl
F, Weininger D, Baehner M, Seeber S, Kettenberger H, Schanzer J, et
al: Anti-tumoral, anti-angiogenic and anti-metastatic efficacy of a
tetravalent bispecific antibody (TAvi6) targeting VEGF-A and
angiopoietin-2. MAbs. 8:562–573. 2016. View Article : Google Scholar : PubMed/NCBI
|