Introduction
Gastric cancer is one of the most common cancers
worldwide and has the second highest mortality (1–3). In
2012 there were 951,600 new gastric cancer diagnoses and 723,100
deaths (4,5). The mass screening for gastric cancer
that has developed in recent years has resulted in the diagnosis of
several patients with an early stage of gastric cancer, and these
patients were subsequently treated in a timely fashion, with
surgery or drug treatment (6).
However, the majority of patients with gastric cancer were
diagnosed at an advanced stage, and their mean survival period was
<1 year (1,7). Recurrence, distant metastasis and
resistance to drug treatment are the main barrier to survival of
patients with advanced stage gastric cancer. Therefore, further
study of the molecular mechanisms of gastric cancer may improve the
therapeutic options for gastric cancer.
c-Maf was discovered as an oncogene transduced in
the avian AS42 retrovirus, which was discovered to be overexpressed
in multiple myeloma and angioimmunoblastic T-cell lymphoma
(8,9). c-Maf inducing protein (CMIP) is an
adaptor protein with two isoforms, which is involved in the c-Maf
signaling pathway. The two isoforms of CMIP were observed to be
expressed in the human brain (10,11).
One of the isoforms is a short protein that participates in several
cell-signaling pathways and was reported to be associated with
minimal change nephrotic syndrome (MCNS) (10–13).
The second isoform of CMIP is a longer protein, for which there is
limited functional information at present (10–13).
In kidney-associated diseases CMIP regulates the behavior of
podocytes (14,15). Newbury et al (11) and Scerri et al (16) demonstrated that CMIP is associated
with reading and language related traits. Audard et al
(13) determined that CMIP
contributes to classical Hodgkin lymphoma (13). However, the relationship between
CMIP and human gastric cancer has not yet been reported.
The present study demonstrated that the expression
of CMIP protein was significantly higher in gastric cancer tissues
compared with normal gastric tissues. CMIP was positively
associated with tumor size, lymph node metastasis, histological
grade and clinical stage in gastric cancer. Patients exhibiting
CMIP expression had poorer post-operative relapse-free survival
(RFS) and post-operative overall survival (OS). Furthermore, CMIP
promoted proliferation and metastasis of gastric cancer cells in
vitro. Notably, the present study demonstrated that miR-101-3p
suppresses the expression of CMIP, and CMIP increases the
expression of MDM2 and mitogen activated protein kinase (MAPK).
CMIP was demonstrated to serve an oncogenic role in human gastric
cancer cells, and may be useful as a biomarker or for further
investigation as a therapeutic target in the clinical management of
gastric cancer.
Materials and methods
Clinical samples
A total of 100 paraffin-embedded surgical gastric
cancer tissue specimens and 100 paraffin-embedded surgical paired
normal gastric tissue specimens (n=100 patients; male, n=54;
female, n=46; age, 60.03±8.77 years; Table I) were collected at the First
Affiliated Hospital of Anhui Medical University (Hefei, China)
between January 2009 and December 2015. Patients who had undergone
chemotherapy or radiation therapy prior to surgery were excluded,
as were patients with rheumatic disease, acute infection, human
immunodeficiency virus or other types of cancer. The pathological
tumor stage was defined according to the sixth edition of the
tumor-node-metastasis classification as defined according to the
2008 World Health Organization classification of tumors (17). Primary study endpoints were
post-operative RFS and post-operative OS. RFS and OS were defined
as the time from the date of surgery to the date of mortality from
gastric cancer, or to the date of local recurrence or detection of
distant metastasis, respectively. All tissue diagnoses were
confirmed by permanent histology. The Institutional Review Boards
of the First Affiliated Hospital of Anhui Medical University
approved the protocol for the use of tissue samples from patients
and the follow-up study. Every patient signed a written informed
consent form.
 | Table I.Correlation of CMIP expression with
clinicopathological parameters from gastric cancer patients. |
Table I.
Correlation of CMIP expression with
clinicopathological parameters from gastric cancer patients.
|
|
| CMIP
expression |
|---|
|
|
|
|
|---|
| Parameter | n | Positive, n
(%) | P-value |
|---|
| Age (years) |
|
|
|
|
≤60 | 53 | 39 (73.6) | 0.708 |
|
>60 | 47 | 33 (70.2) |
|
| Sex |
|
|
|
|
Male | 54 | 35 (64.8) | 0.083 |
|
Female | 46 | 37 (80.4) |
|
| Tumor size
(cm) |
|
|
|
| ≤5 | 67 | 43 (64.2) | 0.013 |
|
>5 | 33 | 29 (87.9) |
|
| Lymph node
metastasis |
|
|
|
| No | 37 | 20 (54.1) | 0.002 |
|
Yes | 63 | 52 (82.5) |
|
| Grade |
|
|
|
| I | 7 | 3 (42.9) | 0.010 |
| II | 62 | 41 (66.1) |
|
|
III | 31 | 28 (90.3) |
|
| Stage |
|
|
|
|
I–II | 54 | 34 (63.0) | 0.029 |
|
III–IV | 46 | 38 (82.6) |
|
Immunohistochemistry analysis
Immunohistochemistry (IHC) analyses of CMIP protein
expression was performed using a Two-Step Histostaining kit (Fuzhou
Maixin Biotech Co., Ltd., Fuzhou, China) with a polyclonal primary
antibody against CMIP (1:100; cat. no. 12851-1-AP; Proteintech
Group, Inc., Chicago, USA). The sections (4 µm) were deparaffinized
in xylene and rehydrated in a graded series of ethanol solutions.
For antigen retrieval, slides were heated in a microwave oven in
0.01 M sodium citrate buffer (pH 6.0) for 20 min. The slides were
allowed to cool in the same buffer and were subsequently immersed
in 3% hydrogen peroxide in methanol for 10 min to block endogenous
peroxidase activity. Following 3 washes for 2 min each with
phosphate-buffered saline (PBS), slides were incubated with primary
antibody at 4°C overnight. The slides were rinsed in PBS as
aforementioned, incubated at room temperature for 20 min with
universal horseradish peroxidase (HRP)-conjugated detection reagent
MaxVision™ HRP-Polymer goat anti-mouse/rabbit IHC kit (undiluted;
cat. no. KIT-5030; Fuzhou Maixin Biotech Co., Ltd.) and rinsed
again in PBS as aforementioned. Immunoreactive regions were
visualized using 3,3′-diaminobenzidine tetrahydrochloride (Fuzhou
Maixin Biotech Co., Ltd.). All IHC slides were counterstained with
hematoxylin. Known positive samples were used as positive controls.
An isotype matched negative control was performed using a Negative
Control for Rabbit IgG at room temperature for 20 min (undiluted;
cat. no. RAB-0102; Maixin Biotech Co., Ltd.).
The stained sections were visualized using a light
Olympus microscope (Olympus Corporation, Tokyo, Japan) and were
reviewed and scored for the expression of CMIP protein
independently by two experienced pathologists who had no knowledge
of the patients' identities and clinical status. Staining intensity
and percentage of tissue staining were recorded: ≥10% tumor cells
stained were considered positive for CMIP expression; <10% tumor
cells stained with any intensity was considered as negative
expression.
Cell culture
The mixed human gastric tubular adenocarcinoma cell
line MKN-28 was obtained from the American Type Culture Collection
(Rockville, MD). MKN-28 was cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and
incubated at 37°C in a humidified atmosphere containing 5%
CO2 (18). MKN-28 cells
have previously been reported to be cross-contaminated with MKN-74
cells (19), however, this had no
impact on the outcome of the present study.
Small interfering (si)RNA and micro
(mi)RNA transfection
Anti-CMIP siRNA/control siRNA and
miR-101-3p/negative control (NC) mimics were purchased from
Shanghai GenePharma Co., Ltd. (Shanghai, China). Transient
transfections of siRNAs and miRNAs into 30–50% confluent cells were
performed in 6-well plates using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. The concentration of siRNAs or miRNAs used was 75
pmol/transfection. Cells were harvested 48 h post-transfection. The
sequences were as follows: siCMIP#1, 5′-CAAAGAAGCUCUCGCACAUTT-3′;
siCMIP#2, 5′-CUCACCUCGAAAUUCCUGATT-3′; siNC,
5′-UUCUCCGAACGUGUCACGUTT-3′; miR-101-3p mimic,
5′-UACAGUACUGUGAUAACUGAA-3′ and NC mimics,
5′-UUCUCCGAACGUGUCACGUTT-3′ (all Shanghai GenePharma Co.,
Ltd.).
Cell proliferation assay
MKN-28 cells were plated in 6-well plates and
transfected with anti-CMIP siRNA or control siRNA. Cell
proliferation was assessed by cell counting assay and Cell Counting
Kit (CCK)-8 reagent. Cells were plated at a density of 10,000/well
and the counting assay was performed on days 2, 3, 4 and 5.
Briefly, cells were stained with 0.4% trypan blue for 30 sec at
room temperature and were counted using a blood cell counting board
(XB.K.25; Shanghai Qiujing Biochemical Reagents and Instrument Co.,
Ltd., Shanghai, China). The CCK-8 assay was performed according to
the manufacturer's instructions, and the absorbance was measured at
570 nm.
Colony formation assay
A colony formation assay was performed as described
previously (20,21). Briefly, 60 mm dishes were coated
with 0.5% agar (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), and
a layer of treated MKN-28 cells (transfected with siCMIP#1,
siCMIP#2 or siNC, and incubated at 37°C in a humidified atmosphere
containing 5% CO2 for 48 h; 1×103 cells/dish)
mixed with 0.3% soft agar was added on top. The plates were
incubated for up to 2 weeks, and the assays were stopped when the
colonies were clearly visible by eye.
Flow cytometry analysis
Flow cytometry was performed to examine cell
apoptosis. MKN-28 cells transfected with CMIP-siRNA or NC-siRNA
were double-stained with Annexin V-fluorescein isothiocyanate and
propidium iodide, and then analyzed by flow cytometry according to
the manufacturer's instructions.
Wound healing assay
Cells (5×105) were cultured in 6-well
plates (Sigma-Aldrich; Merck KGaA) in serum-free RPMI-1640 medium
until 100% confluency was reached. The cell layers were scratched
with a sterile 10 µl tip (Sigma-Aldrich; Merck KGaA), washed 3
times with PBS and then incubated in fresh serum-free RPMI-1640
medium for 24 h. An inverted microscope (Olympus Corporation) was
used to examine and image random fields.
Cell migration and invasion
assays
Cell migration and invasion assays were performed in
24-well Matrigel-coated 8-µm pore Transwell chambers (Corning
Incorporated, Corning, NY, USA) according to the manufacturer's
instructions. For the invasion assay, the upper Transwell chambers
were coated with Matrigel (BD Biosciences, Franklin Lakes, NJ,
USA). The Transwell inserts were left uncoated for the migration
assay. The Transwell chambers were seeded with 2×104
cells that had previously been transfected with siRNAs for 24 h, in
media containing 0.1% FBS. The lower chamber contained media with
5% FBS. The chambers were collected 24 h later and cells in the
lower chamber (i.e., migratory or invasive cells) were stained with
Giemsa and imaged under an inverted microscope. The number of cells
on each membrane was determined by counting the number of cells in
ten high-power (×400) fields and determining the mean number of
cells per field.
Target Scan analysis
To identify the potential miRNAs for CMIP-targeting,
TargetScan release 7.1 was used (http://www.targetscan.org/vert_71/) (18).
Luciferase reporter assay
To perform the luciferase reporter assay, luciferase
reporter plasmids (Promega Corporation, Madison, WI, USA)
containing the CMIP wild type 3′untranslated region (UTR;
CMIP-3′UTR-WT; seed region, AGUACUGU) or the CMIP mutated 3′UTR
(CMIP-3′UTR-MUT; seed region, AACACUAC) were constructed.
Co-transfection of miR-101-3p mimic and luciferase
reporter plasmid (CMIP-3′UTR-WT or CMIP-3′UTR-MUT) was performed in
1×106 MKN-28 cells in a 6-well plate using
Lipofectamine® 2000, according to the manufacturer's
instructions. The cells were washed with PBS twice and lysed using
reporter lysis buffer (Promega Corporation). For the luciferase
assay, 20 µl cell extract and 100 µl luciferase assay reagent were
mixed at room temperature. The firefly luciferase activity of this
mixture was then quantified using a Dual-Luciferase Reporter Assay
System (Promega Corporation).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The RNA isolation and RT-qPCR assay were performed
as described previously (20).
Total RNA from the tissues and cells was extracted using an RNeasy
Mini kit (Qiagen, Inc., Valencia, CA, USA) according to the
manufacturer's instructions. Total RNA was treated with DNase I and
purified using phenol-chloroform. RT-qPCR ofmiR-101-3p was
performed using a mirVana qRT-PCR miRNA Detection kit (Invitrogen;
Thermo Fisher Scientific, Inc.). The RT temperature protocol was as
follows: 42°C for 60 min and 70°C for 10 min. The qPCR
thermocycling conditions were as follows: 95°C for 10 min, followed
by 40 cycles of 95°C for 5 sec and 60°C for 30 sec, and then
dissolution curve analysis at 95°C for 15 sec, 60°C for 30 sec and
95°C for 15 sec. U6 was also detected as the endogenous control.
The primers used were as follows: miR-101-3p, forward
5′-ATGCAAGUCAAUAGUGUCAUG-3′ and reverse 5′-GTGCAGGGTCCGAGGT-3′; U6,
forward 5′-TGGAACGATACAGAGAAGATTAGCA-3′ and reverse
5′-AACGCTTCACGAATTTGCGT-3′. The RT-qPCR for MDM2 and MAPK detection
was performed using RT-qPCR kits [RT kit, PrimeScript™ RT reagent
kit; cat. no. DRR037A; qPCR kit, SYBR PremixEx Taq™ II (Perfect
Real Time); cat. no. DRR081A; both Takara Biotechnology, Co., Ltd.,
Dalian, China]. The RT conditions were as follows: 37°C for 15 min
and 85°C for 5 sec. The qPCR thermo cycling conditions were as
follows: 95°C for 10 min, followed by 40 cycles of 95°C for 5 sec
and 60°C for 30 sec, and then dissolution curve analysis at 95°C
for 15 sec, 60°C for 30 sec and 95°C for 15 sec. β-actin was also
detected as the endogenous control. The primers used were as
follows: MDM2, forward 5′-ACCTCACAGATTCCAGCTTCG-3′ and reverse
5′-TTTCATAGTATAAGTGTCTTTTT-3′; MAPK, forward
5′-CAATGGCGGTGTGGTGTTC-3′ and reverse 5′-AGCTCCCTTATGATCTGGTTCC-3′;
β-actin, forward 5′-TTCCTGGGCATGGAGTC-3′ and reverse
5′-CAGGTCTTTGCGGATGTC-3′. The results were quantified using the
2−ΔΔCq method (22).
The mRNA levels of checkpoint kinase 2 (CHEK2), RB transcriptional
corepressor 1 (RB1), B-cell lymphoma 2 (Bcl-2)-associated X (BAX),
Bcl-2-associated agonist of cell death (BAD), caspase 8 and
FADD-like apoptosis regulator (CFLAR), Fos proto-oncogene, AP-1
transcription factor subunit (FOS) and nuclear factor-kB1 (NF-kB1)
were also determined using the same method, however, their
expression levels were not associated with CMIP (data not shown).
The primers used for these were as follows: CHEK2, forward
5′-AGTGGTGGGGAATAAACGCC-3′ and reverse
5′-TCTGGCTTTAAGTCACGGTGTA-3′; RB1, forward
5′-GAACATCGAATCATGGAATCCCT-3′ and reverse
5′-AGAGGACAAGCAGATTCAAGGTGAT-3′; BAX, forward
5′-GGGTGGTTGGGTGAGACTC-3′ and reverse
5′-AGACACGTAAGGAAAACGCATTA-3′; BAD, forward
5′-CCCAGAGTTTGAGCCGAGTG-3′ and reverse 5′-CCCATCCCTTCGTCGTCCT-3′;
CFLAR, forward 5′-GTGGAGACCCACCTGCTCA-3′ and reverse
5′-GGACACATCAGATTTATCCAAATCC-3′; FOS, forward
5′-TGCCTCTCCTCAATGACCCTGA-3′ and reverse
5′-ATAGGTCCATGTCTGGCACGGA-3′; NF-kB1, forward
5′-TGCCAACAGATGGCCCATAC-3′ and reverse
5′-TGTTCTTTTCACTAGAGGCACCA-3′.
Western blot analysis
Total protein was extracted from MKN-28 cells using
the Nuclear and Cytoplasmic Protein Extraction kit (Beyotime
Institute of Biotechnology, Haimen, China). Proteins (25 µg) were
resolved by 12% SDS-PAGE and the proteins were subsequently
electrotransferred to polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). Membrane blocking was performed at
room temperature for 2 h with 5% (w/v) nonfat milk powder.
Following blocking, the membranes were incubated with polyclonal
primary antibodies against CMIP (cat. no. 12851-1-AP) and β-actin
(cat. no. 20536-1-AP; both diluted at 1:1,000; ProteinTech Group,
Inc., Chicago, IL, USA) overnight at 4°C. The membranes were
subsequently incubated with a secondary antibody [goat anti-rabbit
IgG (H+L) HRP-conjugated secondary antibody; 1:50,000; cat. no.
31460; Invitrogen; Thermo Fisher Scientific, Inc.) for 2 h at room
temperature. Protein bands were identified using an enhanced
chemiluminescence system (EMD Millipore) and the substrates, Femto
(cat. no. 34095; Thermo Fisher Scientific, Inc.) and Pico (cat. no.
34077; Thermo Fisher Scientific, Inc.).
Statistical analysis
All statistical analyses were performed with using
SPSS for Windows (version 18.0; SPSS, Inc., Chicago, IL, USA). All
data were presented as the mean ± standard error of the mean of at
least 3independent experiments. For RT-qPCR, cell counting assay,
CCK-8 assay, cell colony formation assay, cell migration assay,
cell invasion assay, wound healing assay, luciferase reporter assay
and flow cytometry analysis, a one-way analysis of variance
followed by Bonferroni or Tamhane post hoc tests were used.
Pearson's chi-square test was used to analyze the results of the
immunohistochemistry assay and the clinicopathological parameters.
P<0.05 was considered to indicate a statistically significant
difference. Variables associated with OS and RFS rates were tested
using Kaplan-Meier estimates and compared by log-rank test.
Results
Expression of CMIP in gastric cancer
tissues and normal gastric tissues
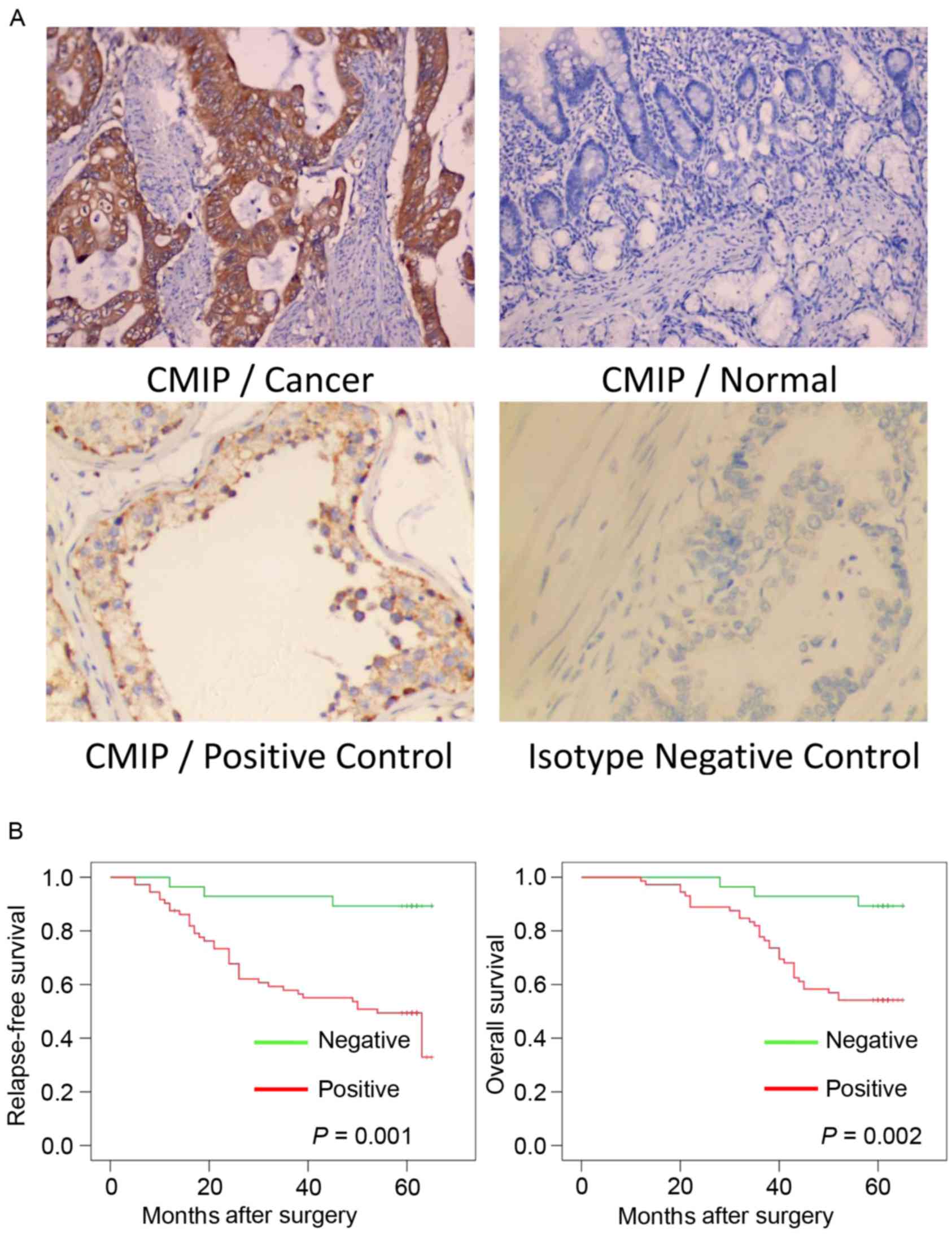
The positive CMIP protein expression signals were
predominantly located in the cytoplasm of the gastric cancer cells
(Fig. 1A). IHC analyses were
performed on 100 gastric cancer tissue specimens and 100 normal
gastric tissue specimens. As presented in Table II, the positive rate of CMIP
protein expression was 72 (72%) in the gastric cancer tissues,
whereas 48 (48%) of the normal gastric tissues positively expressed
CMIP (P<0.001).
| Table II.Expression of CMIP in gastric cancer
and normal tissues. |
Table II.
Expression of CMIP in gastric cancer
and normal tissues.
|
|
| CMIP
expression |
|---|
|
|
|
|
|---|
| Group | n | Negative, n
(%) | Positive, n
(%) |
|---|
| Cancer | 100 | 28 (28.0) | 72
(72.0)a |
| Normal | 100 | 52 (52.0) | 48 (48.0) |
The relationship between CMIP
expression and clinicopathological parameters within gastric cancer
patients
The association of CMIP expression with gastric
cancer prognosis was evaluated. The clinicopathological
characteristics and CMIP expression of the gastric cancer patients
involved in the present study are presented in Table I. Positive expression of CMIP was
associated with tumor size (P=0.013), lymph node metastasis
(P=0.002), histological grade (P=0.010) and clinical stage
(P=0.029) in gastric cancer.
Correlation between CMIP expression
and patient survival
Kaplan-Meier estimates and log-rank tests were
performed to assess whether CMIP expression was associated with
post-operative RFS or OS of gastric cancer patients with complete
follow-up data. The results indicated that patients with primary
tumors expressing CMIP protein had a significantly poorer RFS and
OS compared with those without CMIP protein expression (P=0.001 and
P=0.002, respectively; Fig.
1B).
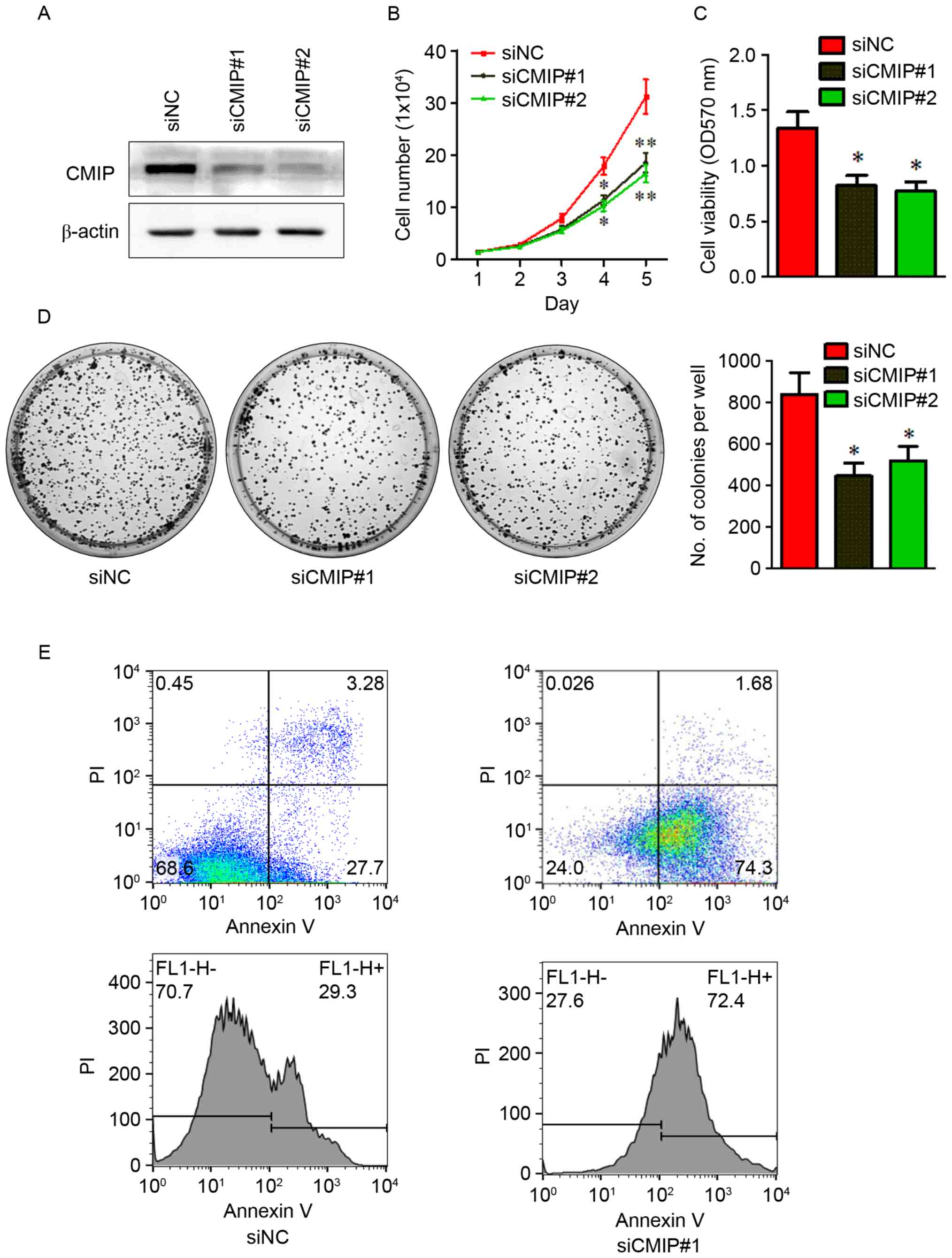
Effects of CMIP inhibition on cell
proliferation and apoptosis of MKN-28 cells
To investigate the role of CMIP in MKN
tumorigenesis, CMIP knockdown was performed in MKN-28 cells by
transfection with CMIP-siRNA. Following transfection, CMIP-siRNA
treated cells had significantly lower CMIP protein levels compared
with cells transfected with control-siRNA (Fig. 2A), indicating that the CMIP
knockdown was successful. The impact of CMIP knockdown on MKN-28
proliferation and growth were assessed using cell counting, CCK-8
and colony formation assays, following treatment with CMIP-siRNA
(75 pmol). As presented in Fig.
2B-D, cells transfected with CMIP-siRNA exhibited a marked
decrease in proliferation, viability and growth, compared with
control-siRNA treated cells. Furthermore, cell apoptosis was
examined using flow cytometry (Fig.
2E). Cells transfected with CMIP-siRNA had greater levels of
apoptosis compared with control-siRNA treated cells (30.98% vs.
75.98%).
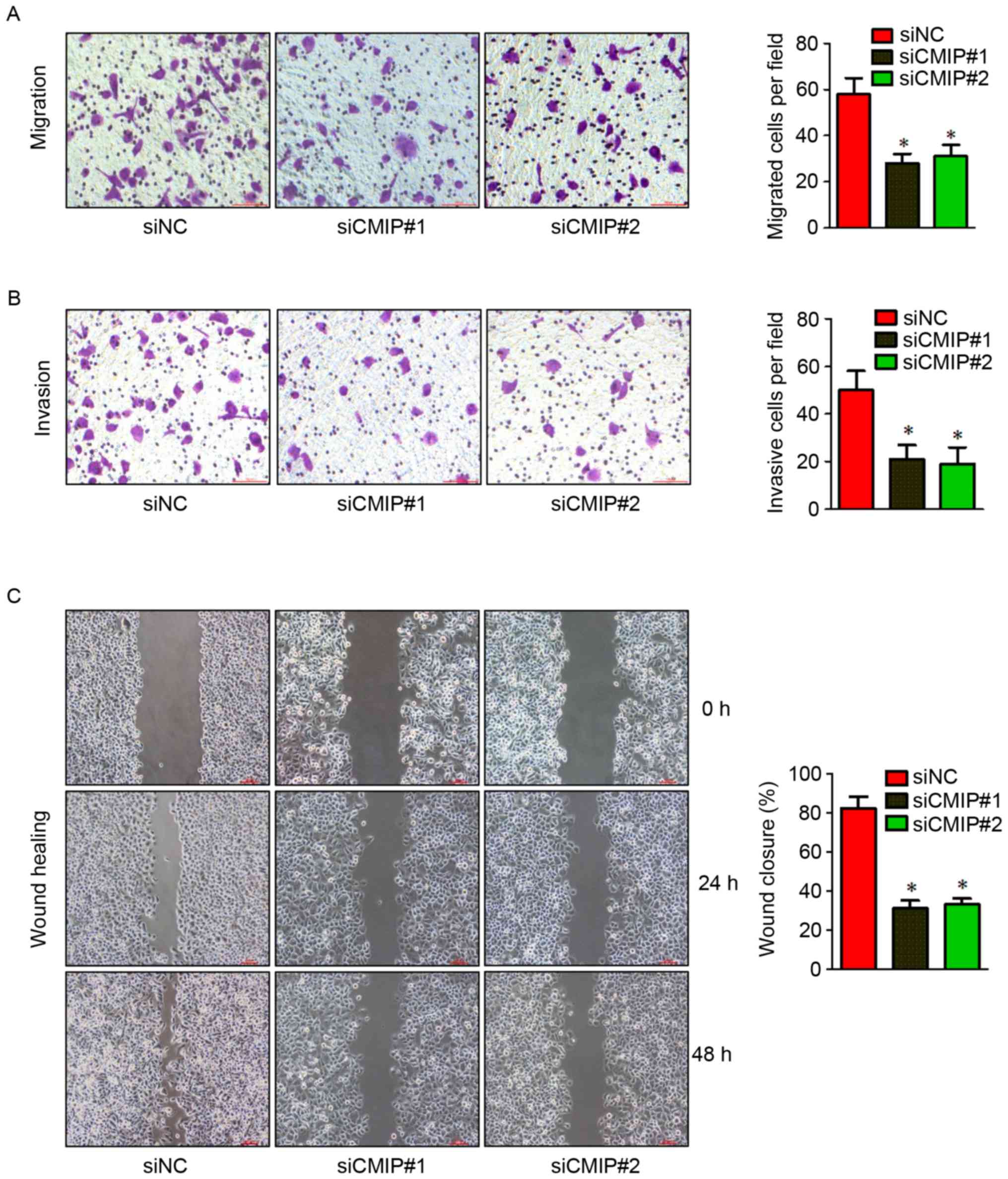
Effects of CMIP inhibition on the
migration and invasion of MKN-28 cells
To explore the role of CMIP in gastric cancer cell
migration and invasion, the effects of CMIP knockdown on MKN-28
cells were assessed using Transwell and wound healing assays.
MKN-28 cells transfected with CMIP-siRNA displayed reduced
migration and invasion in Transwell and wound healing assays
(Fig. 3), compared with
control-siRNA treated cells.
CMIP is a direct target of miR-101-3p
in MKN-28 cells
To investigate the regulation of CMIP in gastric
cancer, TargetScan was used to search for potential CMIP-targeting
miRNAs. The TargetScan analysis indicated that CMIP may be a target
gene of miR-101-3p. The miR-101-3p-binding site was found in the
wild-type 3′-UTR of CMIP, and a luciferase reporter plasmid was
constructed containing the wild type binding site (CMIP-3′UTR-WT).
A reporter plasmid containing a mutant 3′-UTR of CMIP
(CMIP-3′UTR-MUT) was also constructed (Fig. 4A). RT-qPCR was performed to detect
the miR-101-3p expression levels in gastric cancer tissues and
normal gastric tissues. The results indicated that miR-101-3p was
significantly downregulated in gastric cancer tissues (Fig. 4B). To verify whether CMIP was a
direct target of miR-101-3p, a dual luciferase reporter assay was
performed. MiR-101-3p suppressed the luciferase activity of the
wild type CMIP 3′-UTR upon co-transfection of the luciferase vector
in MKN-28 cells (Fig. 4C). This
inhibition was reversed when the seed sequences of the miR-101-3p
target sequences were mutated in the CMIP-3′UTR-MUTvector (Fig. 4C). To further investigate the
effects of miR-101-3p, MKN-28 cells were treated with the
miR-101-3p mimic. The results indicated that overexpression of
miR-101-3p reduced the protein level of CMIP (Fig. 4D).
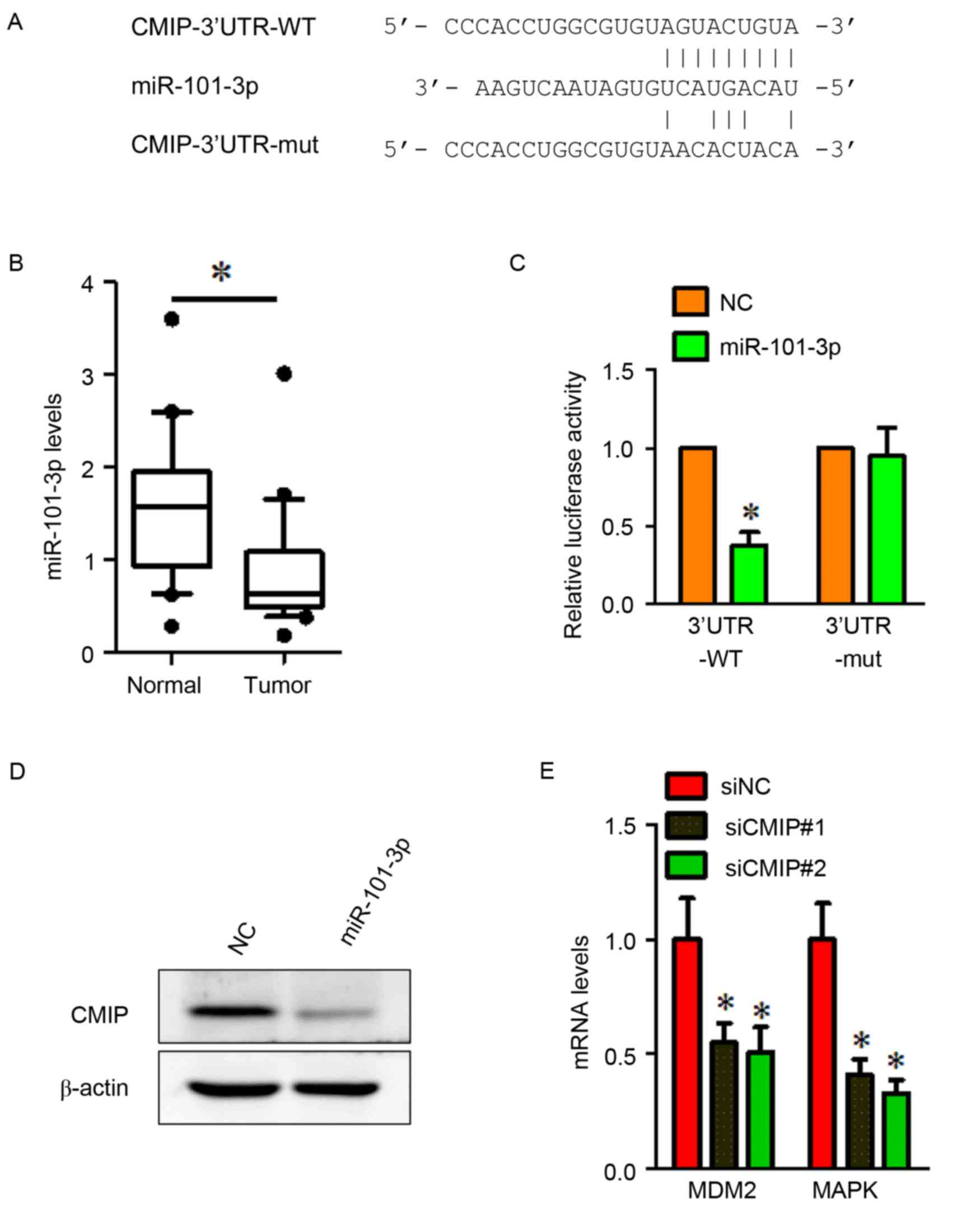 | Figure 4.CMIP is a direct target of miR-101-3p
and CMIP regulates the expression of MDM2 and MAPK. (A) The
TargetScan-predicted binding site between miR-101-3p and the 3′-UTR
of CMIP. The mutant 3′-UTR of CMIP is also presented. (B)
miR-101-3p was down-regulated in gastric cancer tissues compared
with normal gastric tissues. *P<0.05 vs. Normal. (C) Luciferase
assay of MKN-28 cells cotransfected with miR-101-3p mimic or NC,
and a luciferase reporter containing CMIP 3′-UTR wildtype or mutant
constructs. *P<0.05 vs. NC. (D) MKN-28 cells were transfected
with miR-101-3p mimics or NC. miR-101-3p overexpression inhibited
the protein expression of CMIP. β-actin served as a loading
control. (E) Cells transfected with CMIP-siRNA demonstrated a
significant decrease in MDM2 and MAPK mRNA expression compared with
cells transfected with siNC. All data were presented as the mean ±
standard error of the mean of at least 3 independent experiments.
*P<0.05 vs. siNC. CMIP, c-Maf inducing protein; MAPK, mitogen
activated protein kinase; miR, microRNA; NC, negative control; si,
small interfering; 3′UTR, 3-untranslated region; 3′UTR-WT, wild
type 3′UTR construct; 3′UTR-MUT, mutant 3′UTR construct; MAPK,
mitogen activated protein kinase. |
CMIP regulates MDM2 and MAPK
expression in gastric cancer cells
To obtain further insight into the role of CMIP in
MKN-28 cell invasion and metastasis, the expression of several
mRNAs (including cell cycle control and DNA damage repair genes
MDM2, CHEK2 and RB1, apoptosis and cell senescence genes BAX, BAD
and CFLAP and signal transduction molecules and transcription
factors MAPK, FOS and NFKB1) was assessed using RT-qPCR in
siCMIP-transfected MKN-28 cells, compared with si-NC transfected
cells (data not shown). Two downregulated genes (MDM2 and MAPK)
were identified in siCMIP-transfected MKN-28 cells. The results
demonstrated that mRNA levels of MDM2 and MAPK were downregulated
following CMIP knockdown in gastric cancer MKN-28 cells (Fig. 4E).
Discussion
The present study documented for the first time that
CMIP may serve an oncogenic role in human gastric cancer cells. In
200 samples of human gastric tissues (containing 100 cancer and 100
normal tissues), CMIP expression was significantly higher in
gastric cancer tissues compared with normal gastric tissues.
Patients with positive CMIP expression were associated with a
larger tumor size, lymph node metastasis, higher grade, higher
stage, and poorer RFS and OS. Using cell counting, CCK-8 and colony
formation assays, we determined that in MKN-28 gastric cancer
cells, cell proliferation was markedly decreased following
knockdown of CMIP. Furthermore, flow cytometry indicated that cell
apoptosis was significantly increased following CMIP knockdown.
Cell migratory and invasive abilities were also decreased following
knockdown of CMIP in MKN-28 cells, as determined by migration,
invasion and wound healing assays. The oncogene c-Maf was recently
demonstrated to be overexpressed in multiple myeloma and
angioimmunoblastic T-cell lymphoma (8,9).
Overexpression of c-Maf is a frequent oncogenic event in multiple
myeloma that promotes proliferation and pathological interactions
with the bone marrow stroma (23).
CMIP is involved in the c-Maf signaling pathway, which serves an
important role in MCNS and in human reading and language related
behavior (11–13,16).
However, the relationship between CMIP and human carcinoma has not
been well studied. Therefore, the present study examined the role
of CMIP in human gastric cancer.
Upstream of CMIP, a luciferase assay indicated that
CMIP may be a direct target of miR-101-3p in gastric cancer MKN-28
cells. Furthermore, CMIP was significantly reduced by
overexpression with miR-101-3p. miRNAs are non-coding RNAs
consisting of 20–25 nucleotides, which were identified to serve
important roles in the initiation, development, growth,
proliferation and metastasis of human cancer, including gastric
cancer (24–26). miR-25, miR-214 and miR-132, were
demonstrated to promote tumor growth or metastasis of gastric
cancer (27–29), whereas miR-320a, miR-205 and
miR-26b were revealed to suppress tumor proliferation, migration or
invasion of gastric cancer (30–32).
The present study demonstrated that the expression of miR-101-3p
was markedly lower in gastric cancer tissues compared with normal
gastric tissues, indicating that miR-101-3p may serve a tumor
suppressive role in gastric cancer. miR-101-3p suppressed the
expression of CMIP, and miR-101-3p and CMIP exhibited opposing
expression patterns. These results supported the earlier
observation that CMIP expression was elevated in gastric cancer
tissues. As previously reported, miR-101-3p has demonstrated a
tumor-suppressing role in human breast cancer, salivary gland
adenoid cystic carcinoma and hepatocellular carcinoma (33–35),
and these results are consistent with the current findings.
When investigating the downstream pathway, CMIP
knockdown was revealed to downregulate the expression of MDM2 and
MAPK. MDM2 has previously been identified as an oncogene in gastric
cancer; MDM2 promoted tumor initiation and development by targeting
and reducing tumor suppressor genes, including p53 (36,37).
Furthermore, it was reported that MDM2 serves a promoting role in
the migration and invasion of gastric cancer (38), and inhibition of MDM2 expression
could induce the apoptosis of gastric cancer cells (39). MDM2 has also demonstrated a tumor
promoting ability in breast cancer and non-small cell lung cancer
(40,41). MAPK was reported to serve as an
oncogene in gastric cancer, where it promoted tumor growth and
metastasis (42,43). In breast cancer, colon cancer and
hepatocellular carcinoma, MAPK contributed to tumor development,
migration and invasion (44–46).
These results supported the results of the present study, and
suggest that CMIP may serve an oncogenic role in gastric cancer
cells partly via regulating the expression of MDM2 and MAPK.
Therefore, the results imply that CMIP was regulated by miR-101-3p
and itself regulated MDM2 and MAPK; this signaling pathway appears
to be involved in promoting a cancer phenotype in vitro and
may promote the oncogenicity of gastric cancer.
The present study investigated the role of CMIP in
the human gastric cancer cell line MKN-28. This is the first report
on the role of CMIP in human gastric cancer. The cell line selected
for the present study was MKN-28, which was previously reported to
be cross-contaminated with MKN-74 cells (19). MKN-28 and MKN-74 cells have been
reported to contain different genetic and epigenetic alterations of
p53, p21, cluster of differentiation 44 and adenomatous polyposis
coli, and different expression levels of B-cell lymphoma 2,
epidermal growth factor (EGF), EGF receptor, transforming growth
factor-α, interleukin (IL)-1α, IL-8, vascular endothelial growth
factor, cyclin E and platelet derived growth factor (19,47).
These genes have no direct relation with CMIP, the CMIP-related
upstream miR-101-3p and downstream proteins MDM2 and MAPK.
Therefore, use of the mixed gastric tubular adenocarcinoma cell
line MKN-28 had no impact on the outcome of the study.
In conclusion, the present study demonstrated a
tumor-promoting function of CMIP in the MKN-28 cell line.
miR-101-3p targeted CMIP, and knockdown of CMIP downregulated MDM2
and MAPK. It is possible that this signaling pathway may contribute
to the oncogenic role of CMIP in gastric cancer. In patients with
gastric cancer, expression of CMIP was associated with poorer
clinical parameters, RFS and OS. These results suggest that CMIP
may be a novel therapeutic target for gastric cancer; however
further studies are required to investigate this.
Acknowledgements
This study was supported by a grant from the Natural
Science Foundation of Anhui Province (grant no. 1508085MH173).
References
|
1
|
Lee KB, Jin H, Ye S, Park BH and Kim SM:
Recombinant human bone morphogenetic protein-2 inhibits gastric
cancer cell proliferation by inactivating Wnt signaling pathway via
c-Myc with aurora kinases. Oncotarget. 7:73473–73485.
2016.PubMed/NCBI
|
|
2
|
Yin K, Liu M, Zhang M, Wang F, Fen M, Liu
Z, Yuan Y, Gao S, Yang L, Zhang W, et al: miR-208a-3p suppresses
cell apoptosis by targeting PDCD4 in gastric cancer. Oncotarget.
7:67321–67332. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scott RB, Harrison J, Boulton C, Wilson J,
Gregory R, Parkin S, Bain PG, Joint C, Stein J and Aziz TZ: Global
attentional-executive sequelae following surgical lesions to globus
pallidus interna. Brain. 125:562–574. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Feng T, Sun L, Qi W, Pan F, Lv J, Guo J,
Zhao S, Ding A and Qiu W: Prognostic significance of Tspan9 in
gastric cancer. Mol Clin Oncol. 5:231–236. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zakko L, Lutzke L and Wang KK: Screening
and preventive strategies in esophagogastric cancer. Surg Oncol
Clin N Am. 26:163–178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Van Cutsem E, Moiseyenko VM, Tjulandin S,
Majlis A, Constenla M, Boni C, Rodrigues A, Fodor M, Chao Y, Voznyi
E, et al: Phase III study of docetaxel and cisplatin plus
fluorouracil compared with cisplatin and fluorouracil as first-line
therapy for advanced gastric cancer: A report of the V325 Study
Group. J Clin Oncol. 24:4991–4997. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Murakami Y, Yatabe Y, Sakaguchi T, Sasaki
E, Yamashita Y, Morito N, Yoh K, Fujioka Y, Matsuno F, Hata H, et
al: c-Maf expression in angioimmunoblastic T-cell lymphoma. Am J
Surg Pathol. 31:1695–1702. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Benkhelifa S, Provot S, Nabais E, Eychéne
A, Calothy G and Felder-Schmittbuhl MP: Phosphorylation of MafA Is
essential for its transcriptional and biological properties. Mol
Cell Biol. 21:4441–4452. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Szymańska K, Szczałuba K, Lugowska A,
Obersztyn E, Radkowski M, Nowakowska BA, Kuśmierska K, Tryfon J and
Demkow U: The analysis of genetic aberrations in children with
inherited neurometabolic and neurodevelopmental disorders. Biomed
Res Int. 2014:4247962014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Newbury DF, Winchester L, Addis L,
Paracchini S, Buckingham LL, Clark A, Cohen W, Cowie H, Dworzynski
K, Everitt A, et al: CMIP and ATP2C2 modulate phonological
short-term memory in language impairment. Am J Hum Genet.
85:264–272. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grimbert P, Valanciute A, Audard V, Pawlak
A, Le Gouvelo S, Lang P, Niaudet P, Bensman A, Guellaën G and
Sahali D: Truncation of C-mip (Tc-mip), a new proximal signaling
protein, induces c-maf Th2 transcription factor and cytoskeleton
reorganization. J Exp Med. 198:797–807. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Audard V, Zhang SY, Copie-Bergman C,
Rucker-Martin C, Ory V, Candelier M, Baia M, Lang P, Pawlak A and
Sahali D: Occurrence of minimal change nephrotic syndrome in
classical Hodgkin lymphoma is closely related to the induction of
c-mip in Hodgkin-Reed Sternberg cells and podocytes. Blood.
115:3756–3762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Y, Su L, Lin Q, Han Y, You P and Fan
Q: Induction of C-Mip by IL-17 plays an important role in
Adriamycin-induced Podocyte damage. Cell Physiol Biochem.
36:1274–1290. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ory V, Fan Q, Hamdaoui N, Zhang SY,
Desvaux D, Audard V, Candelier M, Noel LH, Lang P, Guellaën G, et
al: c-mip down-regulates NF-κB activity and promotes apoptosis in
podocytes. Am J Pathol. 180:2284–2292. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scerri TS, Morris AP, Buckingham LL,
Newbury DF, Miller LL, Monaco AP, Bishop DV and Paracchini S:
DCDC2, KIAA0319 and CMIP are associated with reading-related
traits. Biol Psychiatry. 70:237–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fletcher CDM, Unni KK and Meretens F:
Pathology and Genetics of Tumours of Soft Tissue and BoneWorld
Health Organization Classification of Tumors. IARC Press; Lyon:
2002
|
|
18
|
Ding K, Wu Z, Wang N, Wang X, Wang Y, Qian
P, Meng G and Tan S: MiR-26a performs converse roles in
proliferation and metastasis of different gastric cancer cells via
regulating of PTEN expression. Pathol Res Pract. 213:467–475. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, MacLeod RA, Masters JR, Nakamura Y, Reid YA,
Reddel RR, et al: Check your cultures! A list of cross-contaminated
or misidentified cell lines. Int J Cancer. 127:1–8. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang T, Zhao B, Li X and Wan J: ARPP-19
promotes proliferation and metastasis of human glioma. Neuroreport.
27:960–966. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding ZY, Jin GN, Wang W, Chen WX, Wu YH,
Ai X, Chen L, Zhang WG, Liang HF, Laurence A, et al: Reduced
expression of transcriptional intermediary factor 1 gamma promotes
metastasis and indicates poor prognosis of hepatocellular
carcinoma. Hepatology. 60:1620–1636. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hurt EM, Wiestner A, Rosenwald A, Shaffer
AL, Campo E, Grogan T, Bergsagel PL, Kuehl WM and Staudt LM:
Overexpression of c-maf is a frequent oncogenic event in multiple
myeloma that promotes proliferation and pathological interactions
with bone marrow stroma. Cancer Cell. 5:191–199. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nip H, Dar AA, Saini S, Colden M, Varahram
S, Chowdhary H, Yamamura S, Mitsui Y, Tanaka Y, Kato T, et al:
Oncogenic microRNA-4534 regulates PTEN pathway in prostate cancer.
Oncotarget. 7:68371–68384. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yoo HI, Kim BK and Yoon SK:
MicroRNA-330-5p negatively regulates ITGA5 expression in human
colorectal cancer. Oncol Rep. 36:3023–3029. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Y, Peng Z, Zhao Y and Chen L:
microRNA-25 inhibits cell apoptosis of human gastric adenocarcinoma
cell line AGS via regulating CCNE1 and MYC. Med Sci Monit.
22:1415–1420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xin R, Bai F, Feng Y, Jiu M, Liu X, Bai F,
Nie Y and Fan D: MicroRNA-214 promotes peritoneal metastasis
through regulating PTEN negatively in gastric cancer. Clin Res
Hepatol Gastroenterol. 40:748–754. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li W, Zhang J, Chen T, Yin P, Yang J and
Cao Y: miR-132 upregulation promotes gastric cancer cell growth
through suppression of FoxO1 translation. Tumour Biol. Aug
23–2015.(Epub ahead of print).
|
|
30
|
Wang Y, Zeng J, Pan J, Geng X, Li L, Wu J,
Song P, Wang Y, Liu J and Wang L: MiR-320a inhibits gastric
carcinoma by targeting activity in the FoxM1-P27KIP1 axis.
Oncotarget. 7:29275–29286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu C, Li M, Zhang L, Bi Y, Wang P, Li J
and Jiang X: MicroRNA-205 suppresses the invasion and
epithelial-mesenchymal transition of human gastric cancer cells.
Mol Med Rep. 13:4767–4773. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tsai MM, Huang HW, Wang CS, Lee KF, Tsai
CY, Lu PH, Chi HC, Lin YH, Kuo LM and Lin KH: MicroRNA-26b inhibits
tumor metastasis by targeting the KPNA2/c-jun pathway in human
gastric cancer. Oncotarget. 7:39511–39526. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu P, Ye F and Xie X, Li X, Tang H, Li S,
Huang X, Song C, Wei W and Xie X: mir-101-3p is a key regulator of
tumor metabolism in triple negative breast cancer targeting AMPK.
Oncotarget. 7:35188–35198. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu XY, Liu ZJ, He H, Zhang C and Wang YL:
MicroRNA-101-3p suppresses cell proliferation, invasion and
enhances chemotherapeutic sensitivity in salivary gland adenoid
cystic carcinoma by targeting Pim-1. Am J Cancer Res. 5:3015–3029.
2015.PubMed/NCBI
|
|
35
|
Sheng Y, Li J, Zou C, Wang S, Cao Y, Zhang
J, Huang A and Tang H: Downregulation of miR-101-3p by hepatitis B
virus promotes proliferation and migration of hepatocellular
carcinoma cells by targeting Rab5a. Arch Virol. 159:2397–2410.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cui W, Wu R, Cao H, Gao J, Wang X and Ren
Q: P53 gene mutation and expression of MDM2, P53, P16 protein and
their relationship in human glioma. J Huazhong Univ Sci Technolog
Med Sci. 25:622–624, 635. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Danilovskyi SV, Minchenko DO, Moliavko OS,
Kovalevska OV, Karbovskyi LL and Minchenko OH: ERN1 knockdown
modifies the hypoxic regulation of TP53, MDM2, USP7 and PERP gene
expressions in U87 glioma cells. Ukr Biochem J. 86:90–102. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shen J, Niu W, Zhou M and Zhang H, Ma J,
Wang L and Zhang H: MicroRNA-410 suppresses migration and invasion
by targeting MDM2 in gastric cancer. PLoS One. 9:e1045102014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang BY, Cao J, Chen JW and Liu QY:
Triptolide induces apoptosis of gastric cancer cells via inhibiting
the overexpression of MDM2. Med Oncol. 31:2702014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Saji S, Nakashima S, Hayashi S, Toi M,
Saji S and Nozawa Y: Overexpression of MDM2 in MCF-7 promotes both
growth advantage and p53 accumulation in response to estradiol. Jpn
J Cancer Res. 90:210–218. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hai J, Sakashita S, Allo G, Ludkovski O,
Ng C, Shepherd FA and Tsao MS: Inhibiting MDM2-p53 interaction
suppresses tumor growth in patient-derived non-small cell lung
cancer Xenograft models. J Thorac Oncol. 10:1172–1180. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wei L, Li Y and Suo Z: TSPAN8 promotes
gastric cancer growth and metastasis via ERK MAPK pathway. Int J
Clin Exp Med. 8:8599–8607. 2015.PubMed/NCBI
|
|
43
|
Yang M, Gu YY, Peng H, Zhao M, Wang J,
Huang SK, Yuan XH, Li J, Sang JL, Luo Q and Huang C: NAIF1 inhibits
gastric cancer cells migration and invasion via the MAPK pathways.
J Cancer Res Clin Oncol. 141:1037–1047. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao M, Howard EW, Parris AB, Guo Z, Zhao
Q and Yang X: Alcohol promotes migration and invasion of
triple-negative breast cancer cells through activation of p38 MAPK
and JNK. Mol Carcinog. 56:849–862. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim GT, Lee SH and Kim YM: Torilis
japonica extract-generated intracellular ROS induces apoptosis by
reducing the mitochondrial membrane potential via regulation of the
AMPK-p38 MAPK signaling pathway in HCT116 colon cancer. Int J
Oncol. 49:1088–1098. 2016.PubMed/NCBI
|
|
46
|
Peng W and Fan H: Long noncoding RNA CCHE1
indicates a poor prognosis of hepatocellular carcinoma and promotes
carcinogenesis via activation of the ERK/MAPK pathway. Biomed
Pharmacother. 83:450–455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yokozaki H: Molecular characteristics of
eight gastric cancer cell lines established in Japan. Pathol Int.
50:767–777. 2000. View Article : Google Scholar : PubMed/NCBI
|