Introduction
Erythropoietin (Epo), a glycoprotein hormone
(1), triggers erythroid progenitor
cell proliferation and differentiation by binding to its specific
membrane receptor (EpoR), which belongs to the type I family of
single transmembrane cytokine receptors. Epo- and EpoR-null mice
die in utero because of the lack of definitive
erythropoiesis and mature erythrocytes (2). Both Epo and EpoR are expressed in the
nervous system during development and adulthood (3), while Epo-null and EpoR-null mice
exhibit thinning of the neuroepithelium and smaller brain size
(4). Furthermore, Epo signaling
supports neural cell survival during development (5), enhances neuronal recovery after
injury in both the developing and mature brain (6), and enhances neurogenesis following
brain injury (5,7,8). The
neuroprotective and neuroregenerative effects of Epo have been
demonstrated in various central nervous system (CNS) diseases
including ischemic stroke, schizophrenia, multiple sclerosis, brain
injury, and spinal cord injury (SCI) (7). As a result, Epo has been used in
clinical trials in patients with brain injury (7) and SCI (9). Under conditions of brain damage, the
neuroprotective effects of Epo are mediated through EpoR-related
signaling to stimulate neurogenesis (10).
Although the exact mechanisms of the neuroprotective
effects of Epo are not fully understood, several processes have
been implicated, including apoptosis inhibition, reduction of
inflammation, restoration of vascular integrity, reduction of
calcium influx, neurotoxic glutamate release, and reduction of
lipid peroxidation by-products (9). Despite the well-documented
Epo-induced recovery following SCI (7,9,11),
the effects of Epo on adult spinal cord neurogenesis have not been
fully characterized. This may be due to the low rate of NSCs
neuronal differentiation. In the adult spinal cord, endogenous NSCs
proliferate following SCI (12),
resulting strictly in a glial cell fate (13). Thus, the functional recovery
remains less than satisfactory. Therefore, replacing injured or
dead neurons and restoring signal transfer functionality are
important processes in the NSC-related recovery following SCI.
Newly born neurons following SCI have been observed only in cases
of mild lesions, and this process is deemed abortive (14,15).
Thus, we speculated that Epo signaling may stimulate neurogenesis
in endogenous NSCs following SCI. Here, we studied the effects of
Epo treatment on the proliferation and differentiation of NSCs both
in vitro and in vivo in the adult spinal cord under
normal conditions and following SCI.
Materials and methods
Animals and SCI model
Specific pathogen-free adult female Sprague-Dawley
rats (weight, 220–250 g; age, 6–8 weeks) were purchased from the
Animal Center of Anhui, China. All experimental procedures were
performed in accordance with the National Institutes of Health
(NIH) Guide for the Care and Use of Laboratory Animals and were
approved by the Institutional Animal Care and Use Committee of
Anhui Medical University, (Hefei, China). The rats were housed
under temperature-controlled conditions (18–26°C) with a 12-h
light/dark cycle (lights ON at 6 AM) and ad libitum access
to water and food. We adopted a modified version of Allen's method
(16) to develop the rat contusion
model of the T10 spinal cord. Briefly, after rats were anesthetized
with pentobarbital sodium (30 mg/kg), a midline incision was made
to uncover the spinal column at the T8-T11 level, followed by a T10
laminectomy to expose the spinal cord. To induce SCI, a 10-g weight
was dropped from a 2.5-cm height onto the exposed and intact dura
over the dorsal aspect of the spinal cord. Subsequently, the
muscle, fascia, and skin were closed. Following the surgery, rats
were administered subcutaneous saline and kept warm.
In vivo experimental procedures
The rats were randomly divided into four
experimental groups (n=18 each), consisting of two conditions
(Normal or SCI) and two treatments over 7 consecutive days at the
same time of the day (Saline or recombinant human Epo [rhEpo, 5,000
U/kg; Four Rings Biopharmaceutical Co. Ltd, Beijing, China)]. The
Normal+Epo group consisted of normal rats treated with
intraperitoneal (i.p.) injections of rhEpo over 7 consecutive days.
The Normal+Saline group consisted of normal rats treated with i.p.
saline at the same volume. The SCI+Epo group consisted of SCI rats
treated with the i.p. rhEpo (5,000 U/kg) immediately after SCI
onset over 7 consecutive days. The SCI+Saline group consisted of
SCI rats treated with i.p. saline at the same volume.
For mitotic labeling, starting immediately after SCI
onset, all rats received i.p. injections of bromodeoxyuridine
(BrdU, 100 mg/kg; Thermo Fisher Scientific, Inc., MA, USA) once per
day for 7 consecutive days. Rats were sacrificed 2, 8, or 14 days
after SCI.
Cell isolation and culture
NSCs were isolated from the spinal cord of SCI rats
(n=3) as previously described (17) with minor modifications. Whole
spinal cords were mechanically dissociated and cultured in
Dulbecco's modified Eagle's/F12 medium (DMEM/F12; Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
2% B-27 (Invitrogen; Thermo Fisher Scientific, Inc.), 20 ng/ml
human epidermal growth factor (PeproTech, Inc., Rocky Hill, NJ,
USA), and 20 ng/ml human basic fibroblast growth factor-2
(PeproTech, Inc.). The cells were cultured at 37°C under 5%
CO2, 95% air, and 90% humidity. The medium was replaced
every third day, while primary-generated neurospheres were passaged
every fifth day. Tertiary neurospheres were used in all in
vitro experiments.
In vitro experimental procedures
Single-cell suspensions from tertiary neurospheres
were seeded at 2,000 cells/well in 96-well plates. The treatment
group (NSC+Epo) received rhEpo (10 U/ml) in the culture
medium for 7 days. The control group (NSC+Vehicle) consisted
of cells cultured in culture medium without rhEpo. Subsequently,
cell proliferation was assessed by flow cytometry.
Neurospheres were dissociated and plated directly
onto laminin-coated glass coverslips in DMEM/F12 medium containing
2% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 2%
B-27, and 10 U/ml rhEpo at a density of 40,000 cells/coverslip.
After 7 days, immunofluorescence staining for cell differentiation
analysis was performed. The cells in the control group were plated
without rhEpo. The specificity of the Epo effect was investigated
by applying neutralizing monoclonal antibody against Epo (mab287;
0.4 ng/ml; R&D Systems, Inc., Minneapolis, MN, USA) to the cell
culture medium.
Immunofluorescence staining
We performed double immunofluorescence to determine
the fate of newly differentiated cells following SCI. Briefly, 4-µm
thick spinal cord paraffin-embedded sections were incubated at 4°C
overnight with a solution containing rabbit anti-BrdU antibody
(1:1,000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and
one of the following cell-specific mouse antibodies: β-tubulin
(β-tubulin III, 1:10,000; Santa Cruz Biotechnology, Inc.) for
neurons, glial fibrillary acidic protein (GFAP, 1:10,000; Abcam,
Cambridge, UK) for astrocytes, and O4 (1:10,000; R&D Systems,
Inc.) for oligodendrocytes. Subsequently, sections were incubated
with tetramethyl rhodamine isothiocyanate (TRITC)-conjugated goat
anti-rabbit secondary antibody (1:500; Santa Cruz Biotechnology,
Inc.) and fluorescein isothiocyanate (FITC)-conjugated goat
anti-mouse (1:500; Santa Cruz Biotechnology, Inc.) for 2 h at room
temperature. The nuclei were counterstained with
4′,6′-diamidino-2-phenylindole (DAPI) (1 g/ml; Sigma-Aldrich) for
10 min at room temperature.
Cell immunostaining was performed as described
above. The coverslips were incubated with primary antibodies
(β-tubulin, GFAP, or O4) and secondary antibody (TRITC-conjugated
goat anti-rabbit or FITC-conjugated goat anti-mouse). The cell
nuclei were counterstained with DAPI. The primary antibodies were
replaced with normal rabbit serum in phosphate-buffered saline
(PBS, 0.01 M) for the negative control. Both slides and coverslips
were mounted, and images were acquired on a confocal microscope
(Leica SP5; Leica Microsystems, Heidelberg, Germany).
Cell quantitation
For in vivo analysis, 10 sections within the
SCI region were selected in each rat, and photomicrographs were
analyzed using ImageJ software (National Institutes of Health,
Bethesda, MD, USA). To assess NSCs proliferation,
BrdU+/DAPI+ cells were counted, and the
numbers were normalized to the measured area (512×512 µm); this
value was then presented as number/mm2. NSCs
differentiation was estimated by quantifying BrdU/β-tubulin,
BrdU/GFAP, and BrdU/O4 double-positive cells. Data were presented
as a percentage relative to the total number of BrdU+
cells. All data were averaged to obtain a mean number from 10
sections per rat. DAPI staining was used to determine the total
number of cells.
In vitro, 10 random fields were selected in
each well with three wells per group, and the numbers of DAPI-,
β-tubulin-, GFAP-, and O4-positive cells were estimated. Data were
presented as a percentage of the DAPI-positive cells (total cells).
All cell counts were performed by observers blinded to the
treatment status.
Western blot analysis
Estimation of the Epo and EpoR protein expression
was performed by western blot analysis. Total lysis of the cells
and spinal cords was conducted using the Radio-Immunoprecipitation
Assay buffer (Thermo Fisher Scientific, Inc.), and protein
concentrations were determined using the bicinchoninic acid protein
assay (Thermo Fisher Scientific, Inc.). Equal amounts (20 µg) of
the proteins were separated by SDS-PAGE gel, which were
subsequently electroblotted onto nitrocellulose membranes. The
membranes were blocked with 5% skim milk and were probed overnight
at 4°C with the primary antibodies anti-Epo (1:800; Santa Cruz
Biotechnology, Inc.) and anti-EpoR (1:800; Santa Cruz
Biotechnology, Inc.). The membrane was then incubated with
secondary horseradish peroxidase-conjugated antibody (1:2,000;
Santa Cruz Biotechnology, Inc.) at room temperature for 2 h. The
membranes were then prepared using an electrochemiluminescence
western blotting kit (Pierce, Rockford, IL, USA) according to the
manufacturer's instructions. The optical density value of each band
was quantified using ImageJ and was normalized to the corresponding
β-actin level. Values were expressed as the fold change relative to
the control value.
In vitro cell proliferation assay
NSCs proliferation was determined using flow
cytometry and the number of neurospheres. Briefly, NSCs were fixed
with ice-cold 70% ethanol and washed with PBS. Subsequently, NSCs
were centrifuged and resuspended in 300 µl propidium iodide
solution (Sigma-Aldrich) and 100 g/ml RNase. Cells were then
incubated at 37°C for 1 h followed by fluorescence-activated cell
sorting (FACS) Calibur flow cytometer (GE, Sunnyvale, USA). Data
were collected using the FACS Diva software (Becton Dickinson) and
were analyzed using ModFit LT (Verity Software House, Toshan, ME,
USA). The number and diameter of the neurospheres of each well were
measured using an inverted phase-contrast microscope (Olympus,
Hamburg, Germany) and were analyzed using ImageJ.
Evaluation of the motor neurological
function
Two examiners blinded to the treatment groups
evaluated the rats hindlimb motor function using the Basso,
Beattie, and Bresnahan (BBB) scale score (18). Assessments were performed at 2, 8,
and 14 days after SCI.
Statistical analysis
Quantitative data were first evaluated for normality
with the Kolmogorov-Smirnov test. Data were presented as the mean ±
standard error of the mean (SEM), independent-samples t-tests and
one-way analysis of variance (ANOVA) were used to assess
significance in two-group and three-group comparisons if data were
normal, otherwise nonparametric test was considered and data were
presented as the median and range (P25, P75). BBB score data were
evaluated with repeated measures ANOVA following rank
transformation. All data analyses were performed using SPSS version
21.0 software (IBM Corp., Armonk, NY, USA). A P<0.05 was
considered statistically significant.
Results
Epo increases NSC neurogenesis and
oligodendrogenesis following SCI in vivo
Compared with the SCI+Saline group, Epo
administration for 7 days significantly increased the percentage of
newly generated neurons at all three time points (all P<0.01;
Fig. 1A and D), while
oligodendrocytes increased on days 8 and 14 (both P<0.05;
Fig. 1B and E). Furthermore, Epo
treatment resulted in fewer GFAP+/BrdU+
immunoreactive astrocytes at all three time points compared with
saline treatment (all P<0.05 Fig.
1C and F).
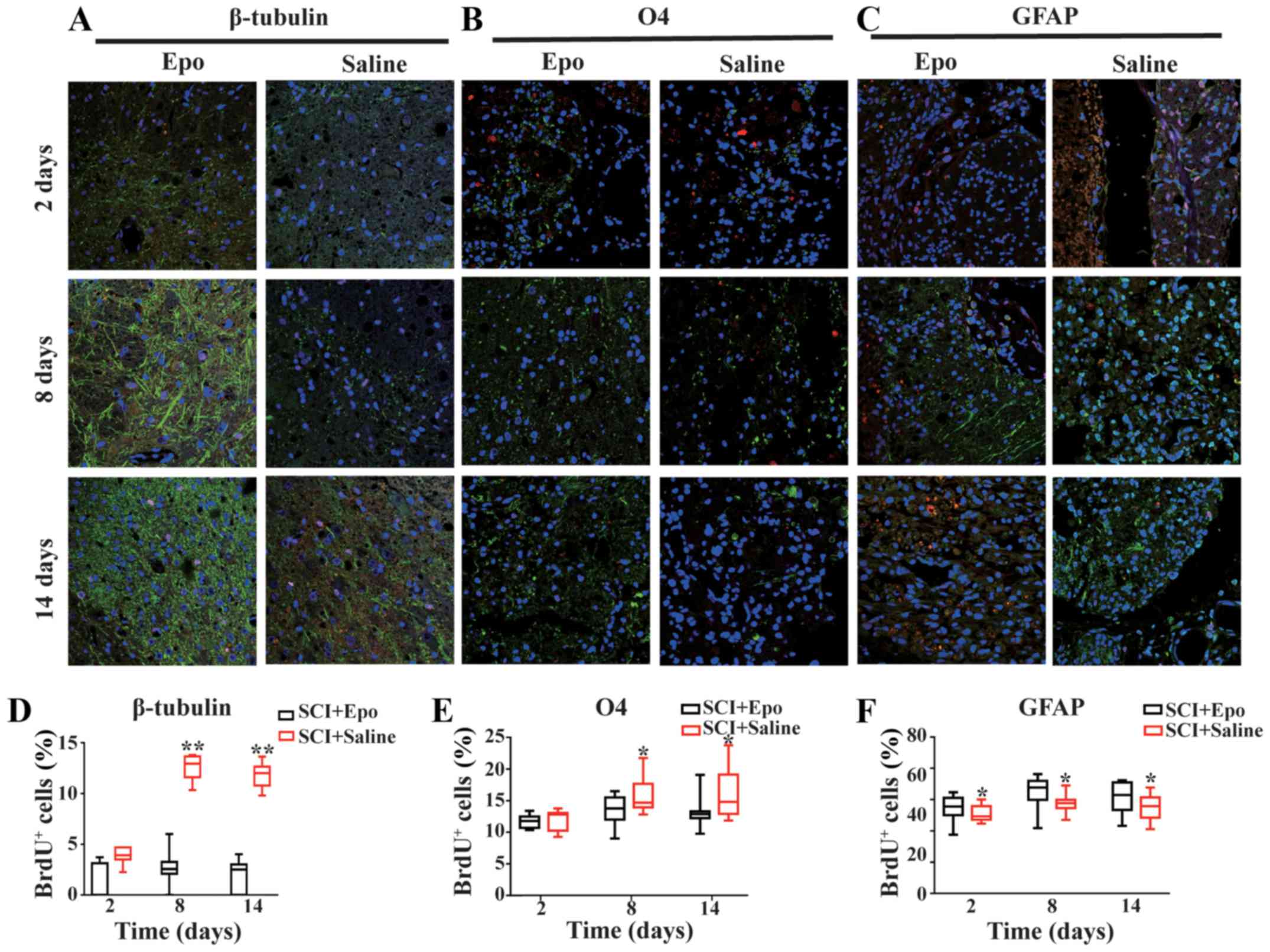 | Figure 1.Effects of Epo signaling on
endogenous NSC differentiation after SCI fluorescence
immunostaining images (merged) showing the co-localization of (A)
β-tubulin (green), (B) O4 (green), and (C) GFAP (green) with BrdU
(red) at 2, 8, and 14 days in the Epo- and Saline-treated groups
after SCI. DAPI (blue) was used as a nuclear counterstain.
Quantitative analysis showed an increased density of (D)
β-tubulin+/BrdU+ cells at all time points and
(E) O4+/BrdU+ cells at day 8 and day 14 with
a decreased density of (F) GFAP+/BrdU+
co-labeled cells in the SCI+Epo vs. SCI+Saline groups. All data are
shown as the median (range) from six animals per group in
vivo. BrdU, bromodeoxyuridine; DAPI,
4′,6′-diamidino-2-phenylindole; Epo, erythropoietin; GFAP, glial
fibrillary acidic protein; NSC, neural stem cells; SCI, spinal cord
injury; O4, oligodendrocyte marker O4. *P<0.05 and **P<0.01
vs. SCI+Epo. |
Epo treatment increases NSC
neurogenesis and oligodendrogenesis in vitro
We observed significantly more cells expressing
β-tubulin (P<0.01; Fig. 2A and
B) and O4 (P<0.01; Fig. 2C and
D) in the NSC+Epo cells compared with NSC+Vehicle
cells. In contrast, the number of newly developed astrocytes
(GFAP+/BrdU+ cells) decreased significantly
in NSC+Epo group (P<0.05; Fig. 2E and F). These changes were not
observed when we used a neutralizing monoclonal antibody against
Epo, suggesting that these effects were specifically induced by Epo
(Fig. 2).
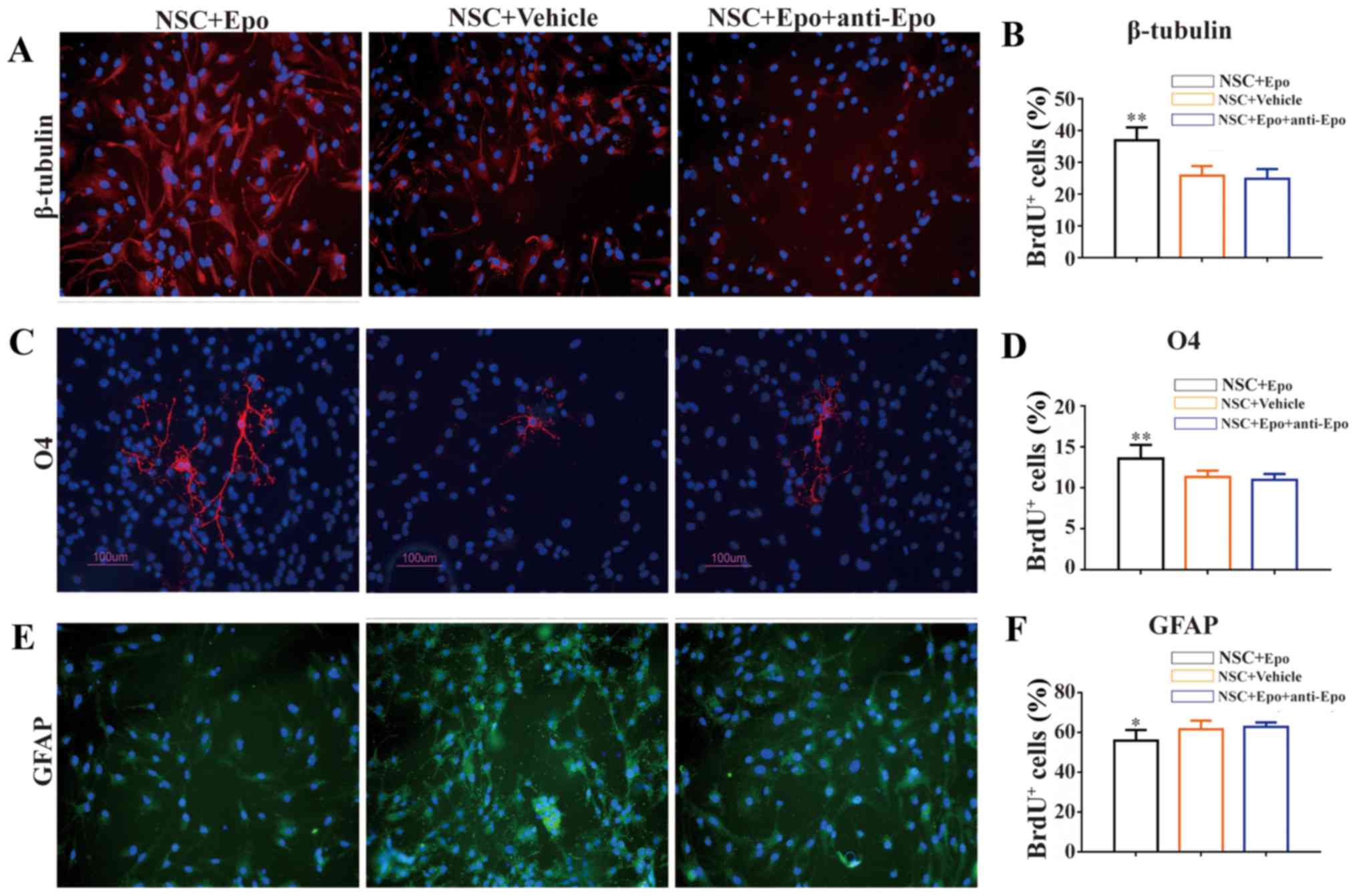 | Figure 2.Effects of Epo signaling on
endogenous NSC differentiation in vitro. (A) Fluorescence
immunostaining images (merged) and (B) quantitative analysis
showing the co-localization of β-tubulin (red). (C) Representative
image (green) and (D) quantitative analysis of O4. (E)
Representative image and (F) quantitative analysis of GFAP (red; E)
with DAPI (blue) at day 8 following exposure to Epo, Vehicle, and
Epo+anti-Epo antibody for 7 days in vitro. DAPI was used as
a nuclear counterstain. Quantitative analysis showed an increased
density of β-tubulin+/BrdU+ cells and
O4+/BrdU+ cells with a decreased density of
GFAP+/BrdU+ cells in the Epo vs. Vehicle
groups. Antibody against Epo neutralized these effects. Scale bars,
100 µm. All data are shown as the mean ± standard error of the mean
from three experiments. BrdU, bromodeoxyuridine; DAPI,
4′,6′-diamidino-2-phenylindole; Epo, erythropoietin; GFAP, glial
fibrillary acidic protein; NSC, neural stem cells; SCI, spinal cord
injury; O4, oligodendrocyte marker O4. *P<0.05 and **P<0.01
vs. control. |
Epo treatment does not affect NSC
proliferation both in vivo and in vitro
We did not observe significant changes in
BrdU+/DAPI+ cells, reflecting NSCs
proliferation, at any time point following SCI between the Epo- and
saline-treated rats (all P>0.05; Fig. 3A and B). Similar results were also
observed in the normal rats treated with Epo or saline (all
P>0.05; Fig. 3C and D).
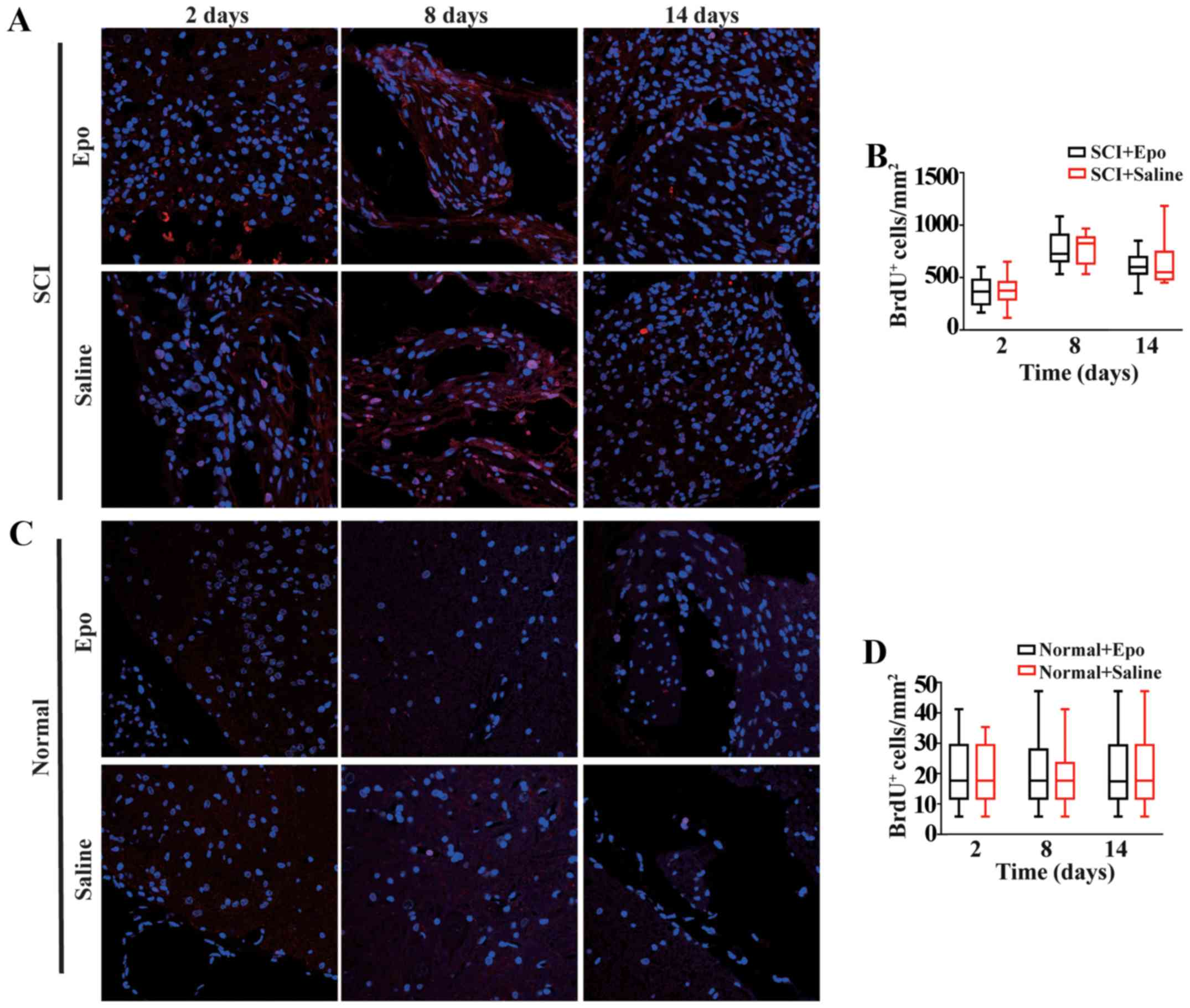 | Figure 3.Effects of Epo signaling on the
proliferation of endogenous NSCs derived from the adult spinal cord
in vivo. (A) Fluorescence immunostaining images (merged) of
BrdU (red) and DAPI (blue) at 2, 8, and 14 days in the Epo- and
Saline-treated groups after SCI. (B) Quantitative analysis showed
no significant difference (P>0.05) in the density of newly
generated cells in either the Epo or Saline groups. (C)
Fluorescence immunostaining images (merged) of BrdU (red) and DAPI
(blue) at 2, 8, and 14 days in the Epo- and Saline-treated groups
in normal rats and (D) quantitative analysis. All data are shown as
the median (range) from six animals per group in vivo. BrdU,
bromodeoxyuridine; DAPI, 4′,6′-diamidino-2-phenylindole; Epo,
erythropoietin; NSC, neural stem cells; SCI, spinal cord
injury. |
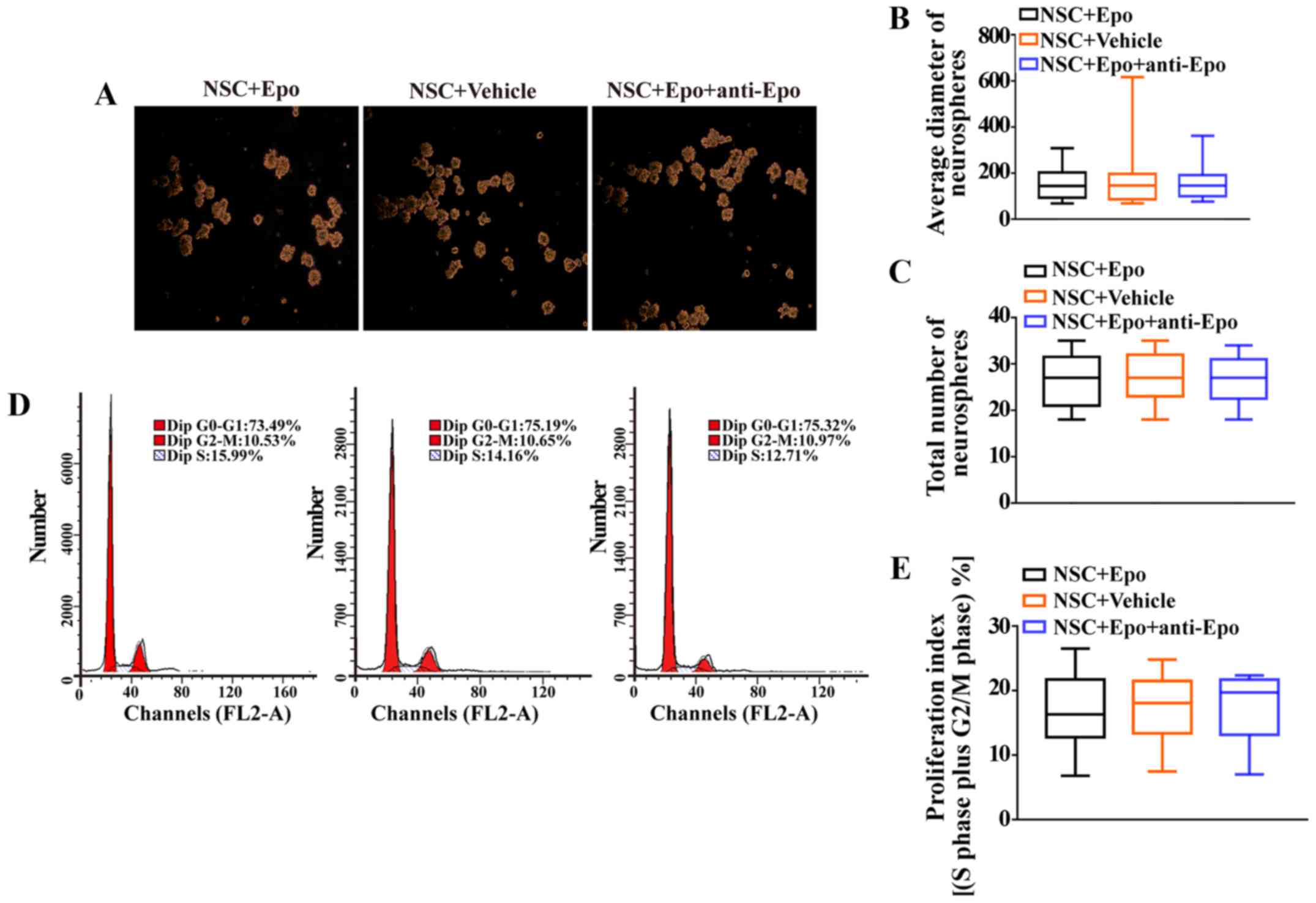
Furthermore, Epo treatment did not alter the average
diameter of neurospheres (P>0.05; Fig. 4A and B) or the total number of
neurospheres compared with NSC+Vehicle cells (P>0.05; Fig. 4A and C). Furthermore, the cell
cycle analysis revealed that the proliferation index (S plus G2/M
phases) was comparable between the Epo-treated cells and controls
(P>0.05; Fig. 4D and E).
Epo treatment in vivo does not affect
the proliferation or differentiation of endogenous NSCs in the
normal spinal cord
At all three time points, no changes in the density
of endogenous NSCs in the normal spinal cord were observed after a
7-day treatment with Epo compared with saline-treated normal rats
(all P>0.05; Fig. 3C and D).
Moreover, we did not observe any BrdU/β-tubulin double-positive
newly generated cells in both normal spinal cord groups.
Furthermore, the numbers of BrdU+/GFAP+ and
BrdU+/O4+ cells remained unchanged in both
groups (all P>0.05; Fig.
5).
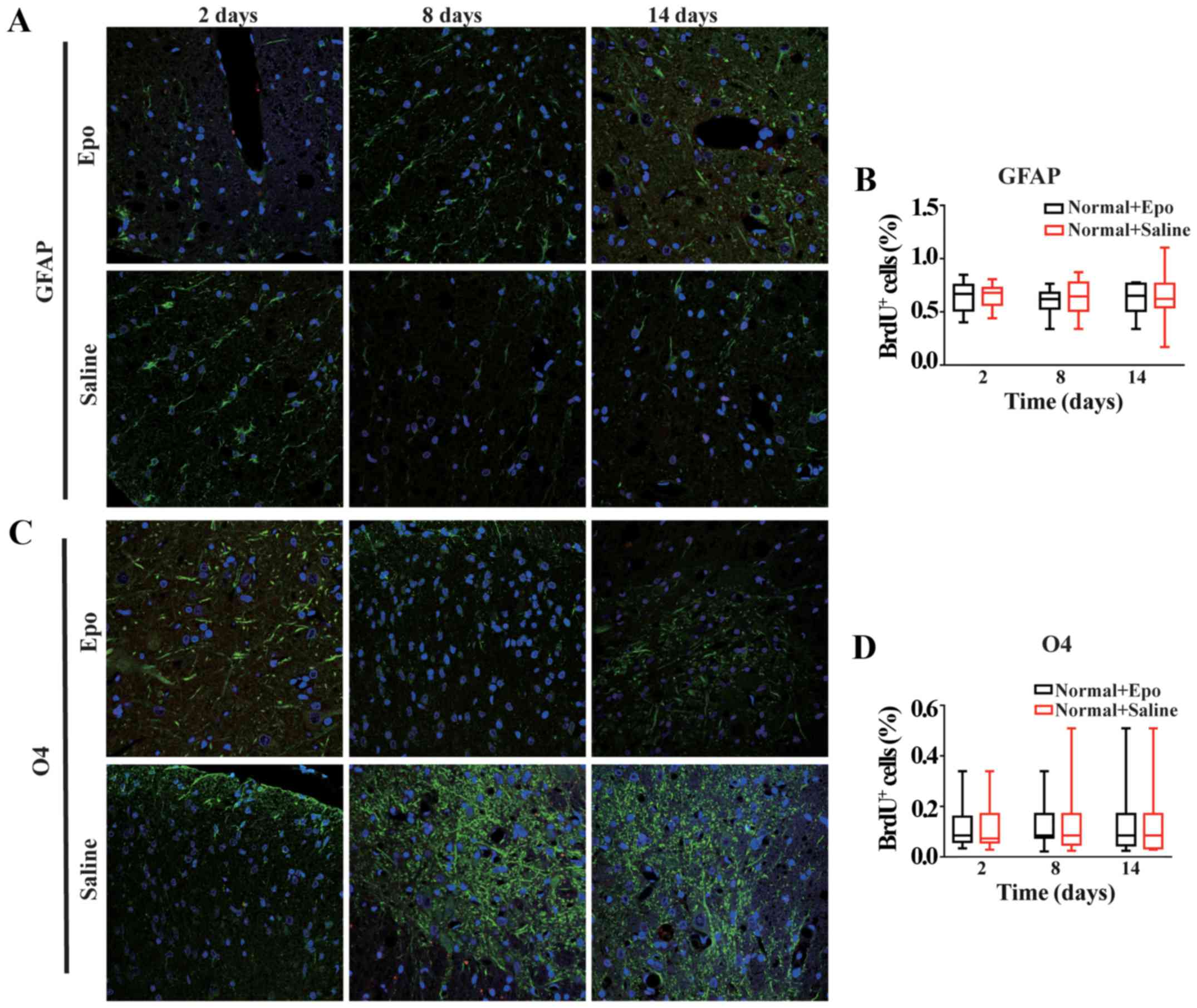 | Figure 5.Effects of Epo signaling on
endogenous NSC differentiation in the normal spinal cord. (A)
Fluorescence immunostaining images (merged) showing the
co-localization of GFAP (green), (B) Quantitative analysis showed
no significant difference in GFAP+/BrdU+ (all
P>0.05). (C) Fluorescence immunostaining images (merged) showing
the co-localization of O4 (green) with BrdU (red) at 2, 8, and 14
days in the Epo- and Saline-treated groups in the normal spinal
cord. DAPI (blue) was used as a nuclear counterstain. (D)
Quantitative analysis showed no significant difference in
O4+/BrdU+ (all P>0.05, D) co-labeled cells
at all three time points in the Normal+Epo vs. Normal+Saline
groups. All data are shown as the median (range) from six animals
per group in vivo. BrdU, bromodeoxyuridine; DAPI,
4′,6′-diamidino-2-phenylindole; Epo, erythropoietin; GFAP, glial
fibrillary acidic protein; NSC, neural stem cells; O4,
oligodendrocyte marker O4. |
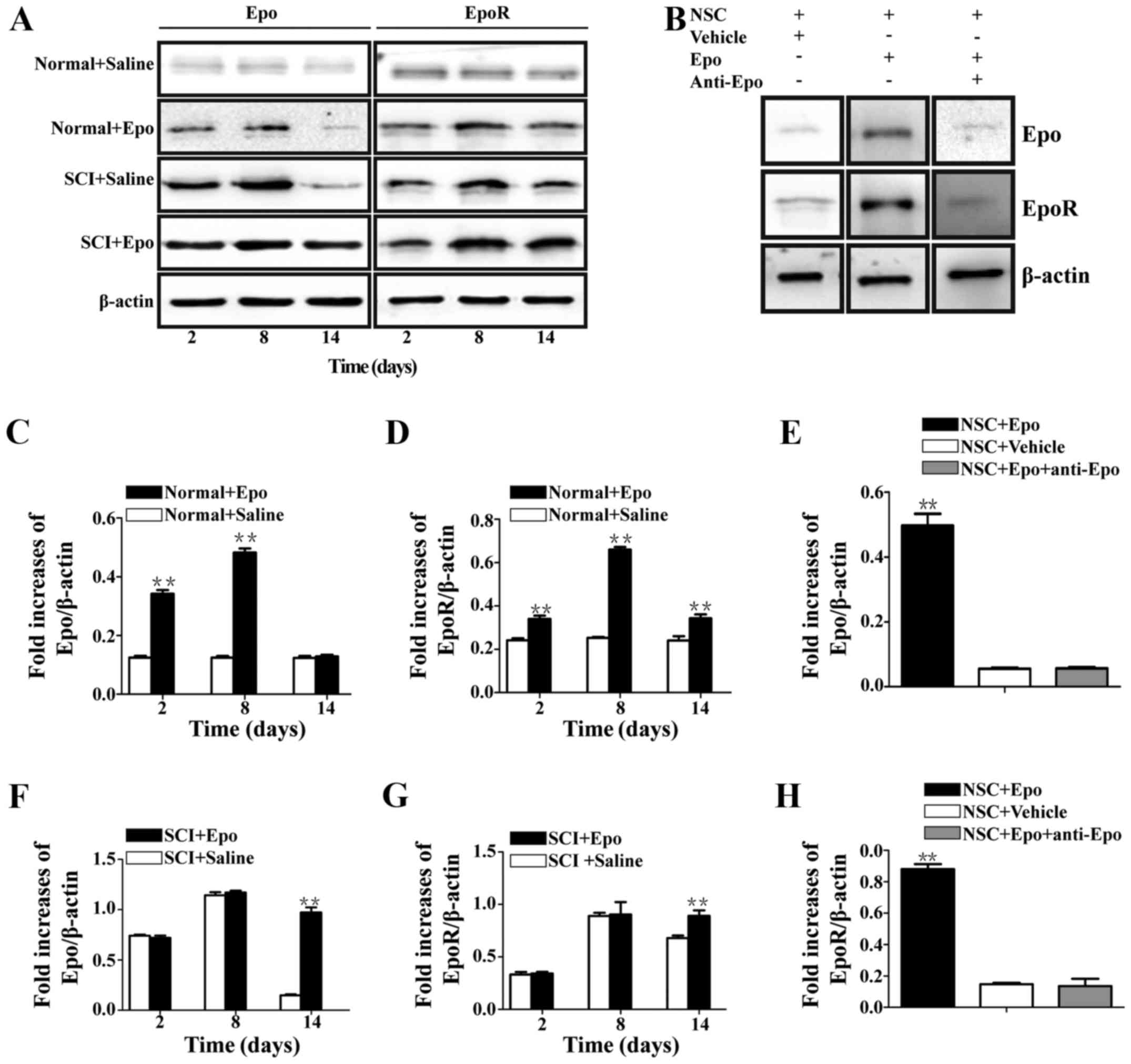
Epo treatment upregulates Epo and EpoR
expression both in vivo and in vitro
Compared with the low Epo and EpoR immunoreactivity
in the Normal+Saline rats (Fig.
6A), these levels significantly increased in the
Normal+Epo group at day 2 (P<0.01), with a maximum Epo
increase of 4-fold (P<0.01; Fig.
6C) and a maximum EpoR increase of 2.7-fold (P<0.01;
Fig. 6D) at day 8. In contrast,
Epo expression returned to normal control levels at day 14
(P>0.05; Fig. 6C), while the
EpoR expression, which also significantly decreased at day 14,
remained significantly higher than the Normal+Saline rats
(P>0.05; Fig. 6D).
Following SCI, both Epo and EpoR immunoreactivities
increased (Fig. 6A); however,
there were no significant differences at days 2 or 8 (P>0.05;
Fig. 6F and G) between the
SCI+Saline and SCI+Epo rats. Furthermore, Epo and EpoR expression
decreased in both groups after day 8, but remained significantly
higher in the SCI+Epo rats at day 14 (P<0.01; Fig. 6F and G) compared with the
SCI+Saline rats.
Similar results were obtained in vitro
(Fig. 6B), where a 9-fold increase
in Epo (P<0.01; Fig. 6E) and a
5.9-fold increase in EpoR (P<0.01; Fig. 6H) were observed in NSC+Epo cells
compared with NSC+Vehicle cells. The specificity of these effects
was confirmed by the neutralizing monoclonal antibody against Epo,
which resulted in no differences (P>0.05; Fig. 6E and H).
Epo treatment promotes motor function
recovery following SCI
The initial trauma following SCI resulted in flaccid
paralysis of the hindlimbs accompanied by urinary retention that
was evacuated by bladder palpation twice per day. The results of
BBB scores are summarized in Table
I. At 2 days post-SCI, no differences were observed between the
SCI+Epo and SCI+Saline rats (P>0.05). However, by
days 8 and 14, the treated rats scored significantly higher than
the controls (P<0.05).
 | Table I.Basso, Beattie and Bresnahan scores
of hindlimb movements following spinal cord injury. |
Table I.
Basso, Beattie and Bresnahan scores
of hindlimb movements following spinal cord injury.
|
| BBB score |
|---|
|
|
|
|---|
| Groups | 2 days | 8 days | 14 days |
|---|
| Epo+SCI | 0 (0,1.0) | 6.0
(4.5,7.0)a | 11.5
(9.8,12.3)a |
| Saline+SCI | 0 (0,0.2) | 2.5 (1.7,3.2) | 6.5 (5.0,8.0) |
Discussion
In the present study, we investigated the effect of
rhEpo treatment on neuronal recovery following SCI. Our findings
provide evidence that 7 consecutive days of systemic delivery of
rhEpo can activate Epo signaling and enhance not only neuronal but
also oligodendrocyte differentiation following SCI both in
vivo and in vitro. However, no effects on proliferation
or in the normal adult spinal cord were observed.
Recent studies reported a protective role of Epo and
EpoR in ischemic and non-ischemic CNS injuries (1,7). Epo
administration leads to substantial neuroprotection and provides a
marked recovery of motor function following SCI; thus, the
Epo-induced prevention of secondary injury has been proposed
(10–12).
We chose the SCI contusion model because it
generates highly reproducible trauma and best mimics common SCI in
humans (19). The Epo dose (5,000
U/kg) was selected based on the best biochemical results obtained
in preliminary studies (20). The
7-day Epo treatment regimen, which allows safe hematocrit levels,
was used based on a previous study (21).
We first examined the effect of Epo signaling in the
non-injured spinal cord. Ransome and Turnley (22) reported that a 7-day systemic
delivery of rhEpo transiently increased the number of
BrdU+ cells by 30%, while neurons were increased by 40%
in the normal sub-granular zone of the dentate gyrus. Furthermore,
a 3-week systemic administration of Epo induced BrdU+
cells and CA1/CA3 neurons, in addition to an approximate 20%
increase in oligodendrocytes in the healthy young mouse hippocampus
(23). In contrast, our data
suggest no effect of 7-day Epo treatment on the number and cell
fate of endogenous NSCs of healthy spinal cords despite marked
increases in Epo and EpoR expressions. Although SCI shares many
pathophysiological processes with brain injury, the adult spinal
cord remains a highly non-neurogenic region that is nearly
completely devoid of neurogenesis under normal conditions (12,13).
Furthermore, adult NSCs transplanted into the neurogenic
hippocampus differentiate into neurons, whereas the same cells
transplanted into the spinal cord produce astrocytic progeny
(24). However, based on our
results, both Epo and EpoR expression were remarkably low in the
Normal+Saline group, which is in agreement with previous studies
(4,20). Notably, under physiological
conditions, Epo has a limited capacity to pass an intact
blood-brain barrier (BBB) via a saturable transport mechanism due
to its relatively high molecular weight and high degree of
glycosylation structures (25). In
line with this, neonates treated weekly with rhEpo either
subcutaneously (1,200 U/kg) or intravenously (1,400 U/kg) to
prevent transfusions did not display higher Epo levels in the
cerebrospinal fluid compared with controls (26). It has been demonstrated that Epo
does not cross the intact BBB of mouse (27), rat (28), and rhesus monkey (29). Furthermore, in a clinical trial,
the intravenous Epo delivery within the first 6 h of stroke failed
to exert any neuroprotective effects (30), which may be due to the fact that
the BBB remains intact in the early hours following stroke
(31).
Although Epo transport across the intact
blood-spinal cord barrier (BSCB) has not been studied, we assume
that Epo access to the spinal cord may be restricted. However,
because a high Epo concentration is a prerequisite for its
neuroprotective function (32), we
postulate that Epo that is transported across the BSCB into the
spinal cord contributes to an increased Epo and EpoR expression,
but may be insufficient for promoting neuroprotection.
While SCI triggers endogenous NSC proliferation,
most of the newly formed cells differentiate into astrocytes, and
neurons are rarely found (12,13).
Epo signaling has been reported to enhance neuronal differentiation
(4,8,22,33)
and production of new oligodendrocytes (34,35)
under both physiological and pathological conditions in the adult
brain. Furthermore, Gonzalez et al (36) reported that Epo treatment promoted
neurogenesis and oligodendrogliosis in neonatal rats after stroke
and in healthy young mice (23).
This is in line with our findings indicating a significant increase
in newly generated neurons at all post-SCI time points and
oligodendrocytes at days 8 and 14 in the Epo-treated SCI rats along
with marked and prolonged Epo signaling activity. While local
injury initiates Epo signaling, the endogenous Epo levels are
insufficient for neuroprotection and repair (21). Epo has been demonstrated to
increase in the spinal cord due to ruptured BSCB following SCI
(25), which may be sufficient to
induce a neuroprotective effect. According to our results, EpoR
expression peaks on day 8 post-SCI and remains relatively high
until day 14, whereas Epo remained nearly undetectable (37). Thus, we suggest that Epo
administration accommodated the excessive vacant EpoR, resulting in
a prolonged Epo signaling activity up until day 14. Oligodendrocyte
progenitor cells and multipotential ependymal cells account for
approximately 85% of endogenous NSCs in the non-injured spinal
cord. However, the rate of division of NSCs increases following
SCI, resulting in the generation of new cells (19). Therefore, Epo acts most likely
through promoting oligodendrogenesis rather than by protecting
oligodendrocytes (37). The effect
of Epo on NSC proliferation remains controversial. Indeed, previous
studies reported contradictory results on proliferation by showing
that Epo induced an increase (8,22,35),
inhibition (23,38), or no effect (35,39,40).
Our results suggested no increase in the density of newly generated
NSCs, which is similar to the effect of Epo in the hematopoietic
system, i.e., driving differentiation rather than directly
stimulating proliferation.
Our in vivo results were also confirmed in
vitro, where we found that 10 U/ml Epo, which has a maximal
effect on both neuronal production (23) and oligodendrogenesis (35), induced neuronal and oligodendrocyte
differentiation to a higher level than that of the saline group
with a concomitant increase in Epo and EpoR expression. Exogenous
Epo directly promoted the neuronal production of NSCs derived from
the brain with a 2.2-fold increase and decreased the secondary NSC
sphere formation in vitro (38). Exposure to Epo and EpoR
neutralizing antibodies dramatically reduced the extent of neuronal
and oligodendrocyte differentiation to a level comparable to
controls.
However, we observed no effect of Epo treatment on
the number and diameter of neurospheres. Similarly, the percentage
of cells at each cell cycle phase was not influenced by Epo. The
in vitro results were also confirmed in vivo. These
findings are in line with findings reported by Marfia et al
(40) in which NSCs from
postmortem mouse brains yielded primarily neurons (approximately
30–40%) following the activation of Epo/EpoR signaling without
altering proliferation.
Furthermore, our results suggested that Epo
administration over 7 days following SCI promotes motor functional
recovery, which is in line with previous reports (41).
To date, several SCI animal models have been
evaluated including trauma, ischemia, inflammation, and
radiation-induced models. In line with these previous studies, our
results demonstrated the effectiveness of Epo treatment in
promoting motor functional recovery (9,11).
Costa et al (41) recently
reported their primary clinical trial data of Epo treatment in
acute traumatic SCI vs. methylprednisolone (the current gold
standard pharmacological treatment). Their results indicated a 27%
absolute difference favoring Epo as an effective treatment.
Based on our present findings and on previous
reports, we suggest that SCI rapidly activates the Epo signaling
pathway, which could be prolonged and reinforced with exogenous Epo
administration. This subsequently results in stimulating neuronal
and oligodendrocyte differentiation and ultimately promoting the
repair process. Scrutinizing the precise mechanisms underlying the
effect of Epo signaling on neuronal repair is crucial for advancing
therapeutic approaches after SCI. Epo signaling has been shown to
be involved in multiple neuroprotective processes after injury,
including anti-apoptotic and anti-inflammatory functions and edema
reduction (1,7,11).
However, the precise mechanism underlying the Epo signaling pathway
and its neuroprotective effects remains unknown and requires
further research.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81171173) and the
Anhui Provincial Natural Science Foundation (grant no.
11040606Q25).
Glossary
Abbreviations
Abbreviations:
|
BBB
|
Basso, Beattie, and Bresnahan
|
|
BrdU
|
bromodeoxyuridine
|
|
BSCB
|
blood-spinal cord barrier
|
|
CNS
|
central nervous system
|
|
DAPI
|
4′,6′-diamidino-2-phenylindole
|
|
DMEM/F12
|
Dulbecco's modified Eagle's/F12
medium
|
|
Epo
|
erythropoietin
|
|
EpoR
|
erythropoietin receptor
|
|
FACS
|
fluorescence-activated cell
sorting
|
|
FITC
|
fluorescein isothiocyanate
|
|
GFAP
|
glial fibrillary acidic protein
|
|
NSCs
|
neural stem cells
|
|
PBS
|
phosphate-buffered saline
|
|
rhEpo
|
recombinant human erythropoietin
|
|
SCI
|
spinal cord injury
|
|
SEM
|
standard error of the mean
|
|
TRITC
|
tetramethyl rhodamine
isothiocyanate
|
References
|
1
|
Nekoui A and Blaise G: Erythropoietin and
nonhematopoietic effects. Am J Med Sci. 353:76–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu H, Liu X, Jaenisch R and Lodish HF:
Generation of committed erythroid BFU-E and CFU-E progenitors does
not require erythropoietin or the erythropoietin receptor. Cell.
83:59–67. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Digicaylioglu M, Bichet S, Marti HH,
Wenger RH, Rivas LA, Bauer C and Gassmann M: Localization of
specific erythropoietin binding sites in defined areas of the mouse
brain. Proc Natl Acad Sci USA. 92:3717–3720. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu X, Shacka JJ, Eells JB, Suarez-Quian C,
Przygodzki RM, Beleslin-Cokic B, Lin CS, Nikodem VM, Hempstead B,
Flanders KC, et al: Erythropoietin receptor signaling is required
for normal brain development. Development. 129:505–516.
2002.PubMed/NCBI
|
|
5
|
Kaneko N, Kako E and Sawamoto K:
Enhancement of ventricular-subventricular zone-derived neurogenesis
and oligodendrogenesis by erythropoietin and its derivatives. Front
Cell Neurosci. 7:2352013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McPherson RJ and Juul SE: Erythropoietin
for infants with hypoxic-ischemic encephalopathy. Curr Opin
Pediatr. 22:139–145. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Simon FH, Erhart P, Vcelar B, Scheuerle A,
Schelzig H and Oberhuber A: Erythropoietin preconditioning improves
clinical and histologic outcome in an acute spinal cord ischemia
and reperfusion rabbit model. J Vasc Surg. 64:1797–1804. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu D, Mahmood A, Qu C, Goussev A,
Schallert T and Chopp M: Erythropoietin enhances neurogenesis and
restores spatial memory in rats after traumatic brain injury. J
Neurotrauma. 22:1011–1017. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Carelli S, Marfia G, Di Giulio AM,
Ghilardi G and Gorio A: Erythropoietin: Recent developments in the
treatment of spinal cord injury. Neurol Res Int. 2011:4531792011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Castaneda-Arellano R, Beas-Zarate C,
Feria-Velasco AI, Bitar-Alatorre EW and Rivera-Cervantes MC: From
neurogenesis to neuroprotection in the epilepsy: Signalling by
erythropoietin. Front Biosci (Landmark Ed). 19:1445–1455. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Simon F, Scheuerle A, Gröger M, Vcelar B,
McCook O, Möller P, Georgieff M, Calzia E, Radermacher P and
Schelzig H: Comparison of carbamylated erythropoietin-FC fusion
protein and recombinant humanerythropoietin during porcine aortic
balloon occlusion-induced spinal cord ischemia/reperfusion injury.
Intensive Care Med. 37:1525–1533. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grégoire CA, Goldenstein BL, Floriddia EM,
Barnabé-Heider F and Fernandes KJ: Endogenous neural stem cell
responses to stroke and spinal cord injury. Glia. 63:1469–1482.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Johansson CB, Momma S, Clarke DL, Risling
M, Lendahl U and Frisén J: Identification of a neural stem cell in
the adult mammalian central nervous system. Cell. 96:25–34. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qin Y, Zhang W and Yang P: Current states
of endogenous stem cells in adult spinal cord. J Neurosci Res.
93:391–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vessal M, Aycock A, Garton MT, Ciferri M
and Darian-Smith C: Adult neurogenesis in primate and rodent spinal
cord: comparing a cervical dorsal rhizotomy with a dorsal column
transection. Eur J Neurosci. 26:2777–2794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin ZS, Zu B, Chang J and Zhang H: Repair
effect of Wnt3a protein on the contused adult rat spinal cord.
Neurol Res. 30:480–486. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yin ZS, Zhang H, Wang W, Hua XY, Hu Y,
Zhang SQ and Li GW: Wnt-3a protein promote neuronal differentiation
of neural stem cells derived from adult mouse spinal cord. Neurol
Res. 29:847–854. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sharif-Alhoseini M, Khormali M, Rezaei M,
Safdarian M, Hajighadery A, Khalatbari MM, Safdarian M, Meknatkhah
S, Rezvan M, Chalangari M, et al: Animal models of spinal cord
injury: A systematic review. Spinal Cord. 55:714–721. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang J, Li Y, Cui Y, Chen J, Lu M, Elias
SB and Chopp M: Erythropoietin treatment improves neurological
functional recovery in EAE mice. Brain Res. 1034:34–39. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaptanoglu E, Solaroglu I, Okutan O,
Surucu HS, Akbiyik F and Beskonakli E: Erythropoietin exerts
neuroprotection after acute spinal cord injury in rats: Effect on
lipid peroxidation and early ultrastructural findings. Neurosurg
Rev. 27:113–120. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ransome MI and Turnley AM: Systemically
delivered Erythropoietin transiently enhances adult hippocampal
neurogenesis. J Neurochem. 102:1953–1965. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hassouna I, Ott C, Wüstefeld L, Offen N,
Neher RA, Mitkovski M, Winkler D, Sperling S, Fries L, Goebbels S,
et al: Revisiting adult neurogenesis and the role of erythropoietin
for neuronal and oligodendroglial differentiation in the
hippocampus. Mol Psychiatry. 21:1752–1767. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shihabuddin LS, Horner PJ, Ray J and Gage
FH: Adult spinal cord stem cells generate neurons after
transplantation in the adult dentate gyrus. J Neurosci.
20:8727–8735. 2000.PubMed/NCBI
|
|
25
|
Brines ML, Ghezzi P, Keenan S, Agnello D,
de Lanerolle NC, Cerami C, Itri LM and Cerami A: Erythropoietin
crosses the blood-brain barrier to protect against experimental
brain injury. Proc Natl Acad Sci USA. 97:10526–10531. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Juul SE, Harcum J, Li Y and Christensen
RD: Erythropoietin is present in the cerebrospinal fluid of
neonates. J Pediatr. 130:428–430. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Banks WA, Jumbe NL, Farrell CL, Niehoff ML
and Heatherington AC: Passage of erythropoietic agents across the
blood-brain barrier: a comparison of human and murine
erythropoietin and the analog darbepoetin alfa. Eur J Pharmacol.
505:93–101. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lieutaud T, Andrews PJ, Rhodes JK and
Williamson R: Characterization of the pharmacokinetics of human
recombinant erythropoietin in blood and brain when administered
immediately after lateral fluid percussion brain injury and its
pharmacodynamic effects on IL-1beta and MIP-2 in rats. J
Neurotrauma. 25:1179–1185. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Boado RJ, Hui EK, Lu JZ and Pardridge WM:
Drug targeting of erythropoietin across the primate blood-brain
barrier with an IgG molecular Trojan horse. J Pharmacol Exp Ther.
333:961–969. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ehrenreich H, Weissenborn K, Prange H,
Schneider D, Weimar C, Wartenberg K, Schellinger PD, Bohn M, Becker
H, Wegrzyn M, et al: Recombinant human erythropoietin in the
treatment of acute ischemic stroke. Stroke. 40:e647–e656. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Latour LL, Kang DW, Ezzeddine MA, Chalela
JA and Warach S: Early blood-brain barrier disruption in human
focal brain ischemia. Ann Neurol. 56:468–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Castañeda-Arellano R, Feria-Velasco AI and
Rivera-Cervantes MC: Activity increase in EpoR and Epo expression
by intranasal recombinant human erythropoietin (rhEpo)
administration in ischemic hippocampi of adult rats. Neurosci Lett.
583:16–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xiong Y, Mahmood A, Zhang Y, Meng Y, Zhang
ZG, Qu C, Sager TN and Chopp M: Effects of posttraumatic
carbamylated erythropoietin therapy on reducing lesion volume and
hippocampal cell loss, enhancing angiogenesis and neurogenesis, and
improving functional outcome in rats following traumatic brain
injury. J Neurosurg. 114:549–559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang L, Chopp M, Zhang RL, Wang L, Zhang
J, Wang Y, Toh Y, Santra M, Lu M and Zhang ZG: Erythropoietin
amplifies stroke induced oligodendrogenesis in the rat. PLoS One.
5:e110162010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jantzie LL, Miller RH and Robinson S:
Erythropoietin signaling promotes oligodendrocyte development
following prenatal systemic hypoxic-ischemic brain injury. Pediatr
Res. 74:658–667. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gonzalez FF, Larpthaveesarp A, McQuillen
P, Derugin N, Wendland M, Spadafora R and Ferriero DM:
Erythropoietin increases neurogenesis and oligodendrogliosis of
subventricular zone precursor cells after neonatal stroke. Stroke.
44:753–758. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Grasso G, Sfacteria A, Passalacqua M,
Morabito A, Buemi M, Macrì B, Brines ML and Tomasello F:
Erythropoietin and erythropoietin receptor expression after
experimental spinal cord injury encourages therapy by exogenous
erythropoietin. Neurosurgery. 56:821–827. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shingo T, Sorokan ST, Shimazaki T and
Weiss S: Erythropoietin regulates the in vitro and in vivo
production of neuronal progenitors by mammalian forebrain neural
stem cells. J Neurosci. 21:9733–9743. 2001.PubMed/NCBI
|
|
39
|
McAdams RM, McPherson RJ, Mayock DE and
Juul SE: Outcomes of extremely low birth weight infants given early
high-dose erythropoietin. J Perinatol. 33:226–230. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Marfia G, Madaschi L, Marra F, Menarini M,
Bottai D, Formenti A, Bellardita C, Di Giulio AM, Carelli S and
Gorio A: Adult neural precursors isolated from post mortem brain
yield mostly neurons: An erythropoietin -dependent process.
Neurobiol Dis. 43:86–98. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Costa DD, Beghi E, Carignano P, Pagliacci
C, Faccioli F, Pupillo E, Messina P, Gorio A and Redaelli T:
Tolerability and efficacy of erythropoietin (EPO) treatment in
traumatic spinal cord injury: A preliminary randomized comparative
trial vs. methylprednisolone (MP). Neurol Sci. 36:1567–1574. 2015.
View Article : Google Scholar : PubMed/NCBI
|