Introduction
With the development of pediatric surgery,
increasing numbers of children are exposed to various types of
narcotic (1). If children are only
given local anesthesia in pediatric surgery, the surgery may be
more difficult or even fail because of the child moving therefore,
general anesthesia has become commonly used in pediatric anesthesia
(1). Previous studies suggested
that the removal of a narcotic via the metabolism of the body
causes patients to reawaken (2).
However, the central nervous system is sensitive to anesthesia, and
is readily damaged; long-term anesthesia may damage the central
nervous system, particularly in children, causing neuronal
degeneration, a reduction in the number of nerve cells or brain
atrophy (3). It has been reported
that general anesthetics (including nitrous oxide, ketamine,
midazolam, pentobarbital sodium and isoflurane), used alone or in
combination over an extended period of time, may lead to widespread
neuronal degeneration during brain development, ultimately causing
irreversible damage to the brain which has effects into adulthood
(4). Sevoflurane is a commonly
used anesthetic in clinical practice, although it is not clear
whether sevoflurane is suitable for long term general anesthesia in
children.
The cerebellum is an important motor center; once
the external information is precision computed and synthesized, a
variety of output instructions are issued by Purkinje cells (PCs)
to facilitate the completion of numerous physiological activities
(5,6). PCs, the only output neuron in the
cerebellar cortex, are highly sensitive to narcotics (6). Repeated anesthesia may lead to PC
degeneration, dendritic cell reduction and cerebellar degeneration
in children (7). An in
vitro study indicated that narcotics may affect central nervous
system function primarily through the regulation of receptors and
neurotransmitter delivery, including activation of the
γ-aminobutyric acidA receptor (GABAA-R) and
glutamate release (8). Anesthetic
drugs may increase the synthesis and release of the presynaptic
GABA transmitter in PCs, and subsequently increase the
concentration of glutamate, resulting in a decrease in the
excitability of PCs (9). A
previous study reported that GABAA-R antagonists may
inhibit glutamate-induced neuronal injury in the early stages of
central nervous system development, and that the regulatory effect
of the GABA-R on glutamate might be associated with the level of
permeability to Cl− (10). Therefore, GABAA-Rs serve
an important role in neuronal injury.
It has been demonstrated that
GABAA-R-mediated inhibition serves a decisive role in
the process of sensory information transmission in the cerebellar
cortex; sensory stimulation may evoke PC-induced inhibitory
postsynaptic potential (IPSP), and a subsequent pause in electrical
discharge (11). However, the
effects of anesthesia on neuronal activity, synaptic transmission
and neural network activity in the mammalian cerebellar cortex are
not clear, and the mechanism of sensory information transmission
inhibition induced by anesthesia in the cerebellar cortex remains
to be elucidated.
Since sevoflurane has the advantages of rapid
induction, stability and rapid recovery, it has become a commonly
used narcotic drug in pediatric anesthesia (12). However, it is unclear whether
intermittent or multiple sevoflurane inhalations have an effect on
the cerebellum and motor function in children. In the present
study, field-clamp-clamp-field potential recording and
pharmacological methods were used to observe the effects of
sevoflurane at a sub-anesthetic concentration on sensory
stimulation-induced cerebellar PC-layer potential activity in
vivo. The expression levels of the GABAA-R ε subunit
were detected, and GABAA-R ε subunit knockout mice were
used to investigate the effects and mechanisms of repeated
sevoflurane administration on neuronal activity, synaptic
transmission, neural network, information transmission, the field
potential response of PCs and motor function. Therefore, the
present study sought to elucidate the role of sevoflurane in the
regulation of the cerebellar cortical neural network and motor
function, and to provide a theoretical basis for the future
clinical application of sevoflurane at different doses and lengths
of time in pediatric anesthesia.
Materials and methods
Animal experiment
GABAA-R ε−/− mice (female;
10–12 weeks old; weight 30±4 g) were purchased from Cyagen
Biosciences (Santa Clara, CA, USA) and normal C57 wild-type
(GABAA-R ε+/+) mice (male; 10–12 weeks old;
weight 30±5 g) were provided by the Experimental Animal Center of
Changzhi Medical College (Changzhi, China). As the reproductive
performance of GABAA-R ε−/− mice is weak, the
GABAA-R ε+/+ mice were mated to
GABAA-R ε+/− mice, and the GABAA-R
ε+/− mice were mated to each other to generate the
GABAA-R ε−/− mice and the GABAA-R
ε+/+ mice; GABAA-R ε−/− mice and
GABAA-R ε+/+ mouse pups were used in the
present study. The mice were housed at a temperature of 20–24°C and
a relative humidity of 50–60%, and were maintained on a 12-h
light/dark cycle, the mice had access to food that was sterilized
by irradiation, and the water contained neomycin (4 g/l) and
bacitracin (4 g/l). In the wild-type (GABAA-R
ε+/+) mouse experiments, 220 neonatal mice were used;
200 neonatal mice were divided into 10 groups (n=20 mice/group) and
the control group included 20 neonatal mice. In order to avoid
confusion, each mouse was marked with an ear tag. For the
experimental neonatal mice, the mice were given repeated inhalation
of 1.5% sevoflurane for 1 h daily via a Julian anesthesia machine
(Draeger, Inc., Telford, PA, USA), sustained for 3, 6, 9, 12, 15,
18, 21, 24, 27 and 30 days, respectively. The control group
received 1 l/min O2 and 1 l/min air for 1 h daily,
sustained for 30 days.
In the GABAA-R ε−/− mouse
experiments, 20 neonatal GABAA-R ε−/− mice
were used as the control group, which received 1 l/min
O2 and 1 l/min air; 40 neonatal GABAA-R
ε−/− mice were used to investigate the role of
GABAA-R ε, and all the mice were divided into 2 groups
(n=20). The mice were given repeated inhalation of 1.5% sevoflurane
for 1 h daily, sustained for 12 or 24 days, respectively. A total
of 40 neonatal GABAA-R ε+/+ mice were used as
the control for the GABAA-R ε−/− mice, and
the mice were divided into 2 groups (n=20) and given repeated
inhalation of 1.5% sevoflurane for 1 h daily, sustained for 12 and
24 days, respectively. Mice were placed into self-made inhalation
anesthesia boxes, with a homemade water bath box at a temperature
of 30–34°C. The concentration of sevoflurane in the inlet and
outlet was continuously monitored using a gas monitor (Draeger,
Lübeck, Germany) to control the experimental conditions. Following
repeated anesthesia for 1 month, at 2 months following the birth of
the mice, cerebellar electrophysiological examination was
performed. The present study was approved by the Animal
Experimental Ethics Committee of Changzhi Medical College.
Cerebellar electrophysiological
examination
Following repeated anesthesia for 1 month, the mice
were placed in a special brain stereotaxic device, with bilateral
ear and maxillary fixing of the head. Craniotomy was performed
using a precision dental drill with a diameter of 1–1.5 mm at the
crus II of the cerebellum; the dura was carefully removed to expose
the recording site. When the control electrode had been mounted,
the mouse and brain stereotaxic apparatus were fixed onto the
microscope (Nikon FN1; Nikon Corporation, Tokyo, Japan), to locate
the operation site at the skull opening, avoiding blood vessels and
perform the electrophysiological recording. The surgical site was
perfused with artificial cerebrospinal fluid (124 mM Nacl, 26 mM
NaHCO3, 2.5 mM KCl, 2 mM CaCl2, 1 mM
MgCl2, 1.25 mM NaH2PO4, 10 mM
D-glucose, and the solution was aerated with 95% O2 and
5% CO2) using a peristaltic pump (MiniPuls 3; Gilson,
Inc., Middleton, WI, USA). Body temperature was maintained at
37±0.2°C during the surgical procedure and was recorded through a
body temperature maintainer and heating pad.
PC layer electrophysiological
recording
A thin glass tube (outer diameter, 1.5 mm; Narishige
Group, Tokyo, Japan) was drawn into a recording electrode using an
automatic drawing apparatus (PB-10; Narishige Group). The electrode
was filled with 20–30 µl artificial cerebrospinal fluid; following
filling, the electrode impedance was 4–6 MΩ. The recording
electrode was mounted on an electric micromanipulator (Sutter
Medical Technologies USA, Atlanta, GA, USA) and the electrode
movement was detected with a microscope (Nikon Eclipse E600FN;
Nikon Corporation). PC discharge was recorded using axopatch-1D
patch-clamp amplifiers (Molecular Devices, LLC, Sunnyvale, CA, USA)
and Clampex 8.1 data acquisition and analysis software (Molecular
Devices, LLC) was used. Following confirmation of the recording
site under a microscope, the electrodes were placed on the brain
surface of the recording area and slowly punctured into the pia
mater from 200 to 400 µm to reach the PC layer of the cerebellar
cortex, adjusting the amplitude of the discharge to a maximum of
0.5 mV. When the discharge frequency and amplitude were stable
following 100 sec, the electrode was recorded. With the
extracellular recording conditions, the confirmation of PC
discharge was based on simple discharge (simple spike) and complex
discharge (complex spike).
Immunofluorescence and
immunohistochemistry
Upon completion of the electrode recording, the
brain was rapidly extracted and placed on ice, and was briefly
washed in ice-cold PBS (Beyotime Institute of Biotechnology,
Haimen, China; 0.2 mol/l; pH 7.4; diluted in double distilled
water). The cerebellum was subsequently isolated and fixed in 4%
paraformaldehyde (Beyotime Institute of Biotechnology) diluted in
PBS at 4°C for 24–48 h, followed by 30% sucrose (Sangon Biotech
Co., Ltd., Shanghai, China; diluted in PBS) for an additional 24
hat 4°C. When the brain tissue had been fully immersed, it was
embedded in Optimum Cutting Temperature embedding agent (Sangon
Biotech Co., Ltd.) and frozen in a liquid nitrogen tank, removed
and frozen at −20°C, and cut on a cryostat (Leica CM3050S; Leica
Microsystems GmbH, Wetzlar, Germany) to a thickness of 15-µm. Brain
slices were blocked with 10% goat serum (OriGene Technologies,
Inc., Beijing, China) at room temperature for 30 min. The goat
serum was diluted in a solution containing 0.3% Triton X-100 and
0.1 mol/l PBS; the Triton X-100 is used to perforate the cell
membrane in order to promote antibody penetration into cells.
Following blocking, the blocking solution was aspirated and the
primary antibody, mouse immunoglobulin (Ig)G
anti-GABAA-R ε (cat. no. sc-271668; 1:300; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) or mouse IgG FluoroPan
Neuronal Marker-Alexa488-conjugated (cat. no. MAB2300X; 1:100; EMD
Millipore, Billerica, MA, USA), was added and incubated at 4°C
overnight. PBS was used as a negative control. The secondary
antibody, goat anti-mouse IgG fluorescein isothiocyanate (1:500;
cat. no. F-2761; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) or goat anti-Mouse IgG phycoerythrin (1:500; cat.
no. P-852; Invitrogen; Thermo Fisher Scientific, Inc.) was added,
and incubated for 2 h at room temperature. DAPI (Sangon Biotech,
Co., Ltd.) was added 10 min prior to the end of incubation.
Following each step, the samples were washed in PBS for 10 min;
antibody incubation and rinsing steps were performed on a shaker.
Fluorescent encapsulated tablets (SouthernBiotech, Birmingham, AL,
USA) were used for 10 min in the dark. Immunohistochemical staining
was performed using an immunohistochemistry detection kit,
according to the manufacturer's protocol (PowerVision™
Two-Step; OriGene Technologies, Inc.). All the samples were
observed by fluorescence microscope (magnification, ×400) (Zeiss
AG, Oberkochen, Germany).
Western blotting
Total proteins of the cerebellum were extracted
using radioimmunoprecipitation assay lysis solution (Beyotime
Institute of Biotechnology), proteins were quantified via a
bicinchoninic acid assay (Beyotime Institute of Biotechnology), and
western blot analysis samples were prepared, 30 µg protein/lane was
separated by 15% SDS-PAGE. Subsequently, the sandwich structure
composed of sponge, filter paper and polyvinylidene fluoride (PVDF)
membrane was placed in the transfer machine at 200 mA for 90 min.
The PVDF membrane was incubated with 5% bovine serum albumin
(Beyotime Institute of Biotechnology) [diluted with PBS-Tween-20
(PBST)] at room temperature for 1 h. The primary antibodies
anti-GABAA-R ε (cat. no. sc-271668; 1:300; Santa Cruz
Biotechnology, Inc.) and anti-GAPDH (cat. no. sc-293335; 1:1,000;
Santa Cruz Biotechnology, Inc.) were incubated at 4°C overnight,
and then washed three times with PBST for 10 min each. The
secondary antibody mouse immunoglobulin (Ig)G conjugated to
CruzFluor™ 488 (cat. no. sc-516176; 1:3,000; Santa Cruz
Biotechnology, Inc.) was incubated at 37°C for 1 h. The PVDF
membrane was washed in PBST three times for 10 min each, and images
were captured using an enhanced chemiluminescence solution
(Beyotime Institute of Biotechnology), and the gray scales were
scanned by Chemidoc XRS+ Imager (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) and the gray scale values were quantified
analysis using Quantity One Analysis Software (version 4.6.9;
Bio-Rad Laboratories, Inc.).
Gait kinematics detection and
analysis
Subsequent to the repeated anesthesia and electrode
recording for 1 month, 2 months following the birth of the mice,
the footprints of mice in each group were recorded using a
transparent table, and the CatWalk Automatic Gait Analysis system
(Noldus Information Technology bv, Wageningen, The Netherlands) was
used to automatically identify and analyze the footprints of the
mice. A total of 97 indices, including walking cycle, step width,
footprint area, speed, standard deviation of body angle, support
duration, swing time, braking duration and propulsion time, were
recorded and analyzed. Each mouse was subjected to at least three
tests. For each test, the mice walked continuously for the length
of the transparent glass plate. The experiment was performed in a
darkroom environment. The gait analysis system was used to
automatically extract and analyze gait video data, the gait
kinematics parameters were obtained and the data were analyzed.
Statistical analysis
All electrophysiological data were analyzed using
Clampfit 8.1 software (Molecular Devices, LLC). All data are
expressed as the mean ± standard deviation. SPSS 19.0 software (IBM
Corp., Armonk, NY, USA) was used for statistical analysis.
Independent t-tests were used for comparisons between groups,
two-way analysis of variance was used for multi-group comparisons,
and further pairwise comparisons were analyzed by the least
significant difference test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Characteristics of field potential
response of PCs induced by sensory stimulation in the mouse
cerebellar cortex
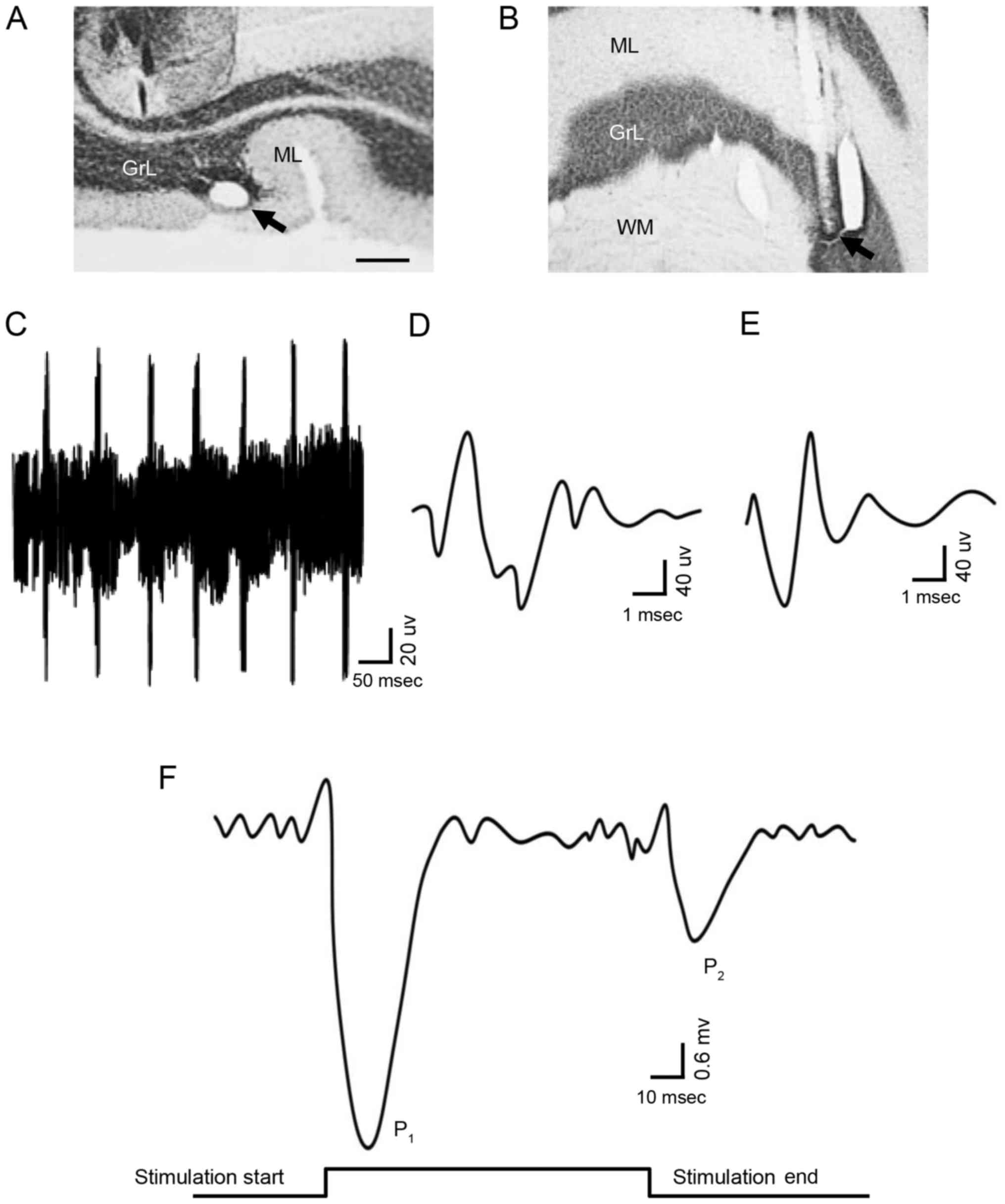
In order to record the potential responses of PCs,
the recording electrodes were located at the specific histological
region where the PCs are existence (Fig. 1A and B). Single-unit spike signals
were excluded and the low noise signals, contributed to only by the
PCs, were detected to record the time axle of electrophysiological
behavior in the experiment. The PCs was identified depending on the
complex spikes and simple spikes, via the waveform analysis
algorithm. The criteria for spike identification of PCs (Fig. 1C) were as follows: i)
Signal-to-noise ratio of individual action potentials >5:1; ii)
existence of the complex spike (Fig.
1D); and iii) presence of the simple spike (Fig. 1E) followed by the complex spike. In
the resting state, PCs did not exhibit synchronous excitatory
activity; therefore, the field potential of the PCs was not
altered. However, stimulation of the ipsilateral whisker pad (60
msec; 50–60 psi) was able to induce PCs to generate a pair of
excitatory field potential responses (Fig. 1F); the presence of the first peak
response (P1) followed by the commencement of
stimulation, and the presence of the second peak (P2)
followed by the end of the stimulation,=indicated that
P1 was induced by the stimulation, while P2
was caused by the end of stimulation. The results of the present
study indicated that sensory stimulation information may be
integrally transmitted by cerebellar PCs, and that the information
contains the onset and end of the stimulus. The above results
suggested that the cerebellar PCs are able to transmit sensory
information with high fidelity.
Repeated sevoflurane inhalation
inhibits the field potential response of PCs induced by sensory
stimulation in neonatal mice
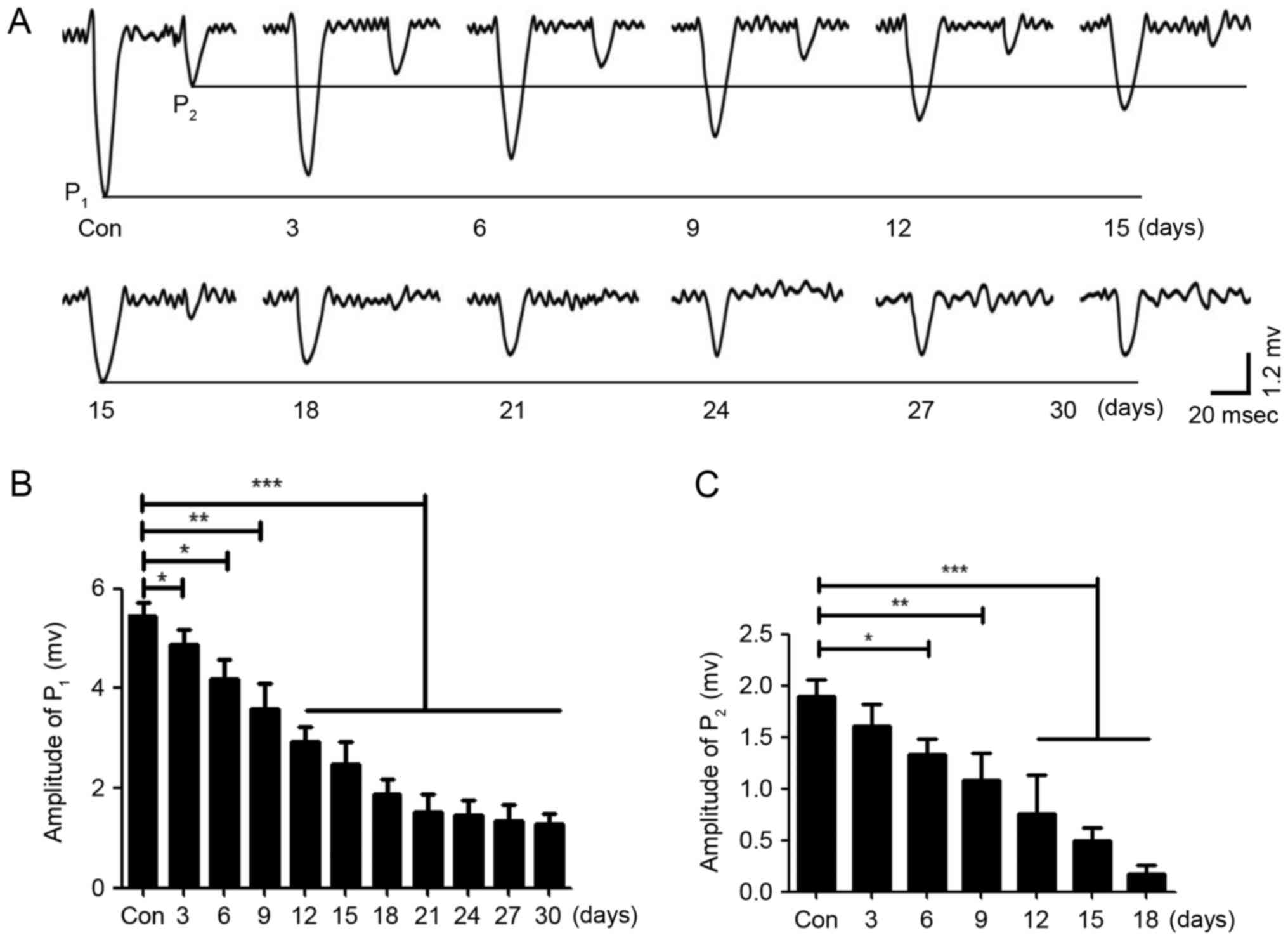
Repeated anesthesia was given for different lengths
of time in neonatal mice, which were housed for 1 month. In the
current clamp mode (I=0), the bimodal excitatory field potential
responses of PCs were induced by the stimulation to the ipsilateral
whisker pad and recorded (Fig.
2A). The results demonstrated that the average peak values of
P1 and P2 decreased with an increased length
of repeated anesthesia; the peak value of P1 no longer
continued to decline when the neonatal mice received repeated
anesthesia for 21 days (Fig. 2A and
B), and the peak value of P2 almost disappeared when
the neonatal mice received repeated anesthesia for 21 days
(Fig. 2A) and statistical analysis
was also performed (Fig. 2C).
These results indicated that the central nervous system may be
damaged by the repeated inhalation of sevoflurane in neonatal mice,
and that the amount of sevoflurane may be negatively associated
with neuronal activity and synaptic transmission, potentially
leading to neuronal dysfunction. Therefore, sevoflurane may damage
the central nervous system in a dose-dependent manner, although
high concentrations of sevoflurane did not completely inhibit the
response of PCs to stimulation; thus, sevoflurane may only
partially inhibit the sensory information transmission of PCs.
Effect of repeated inhalation of
sevoflurane on the movement behavior of neonatal mice
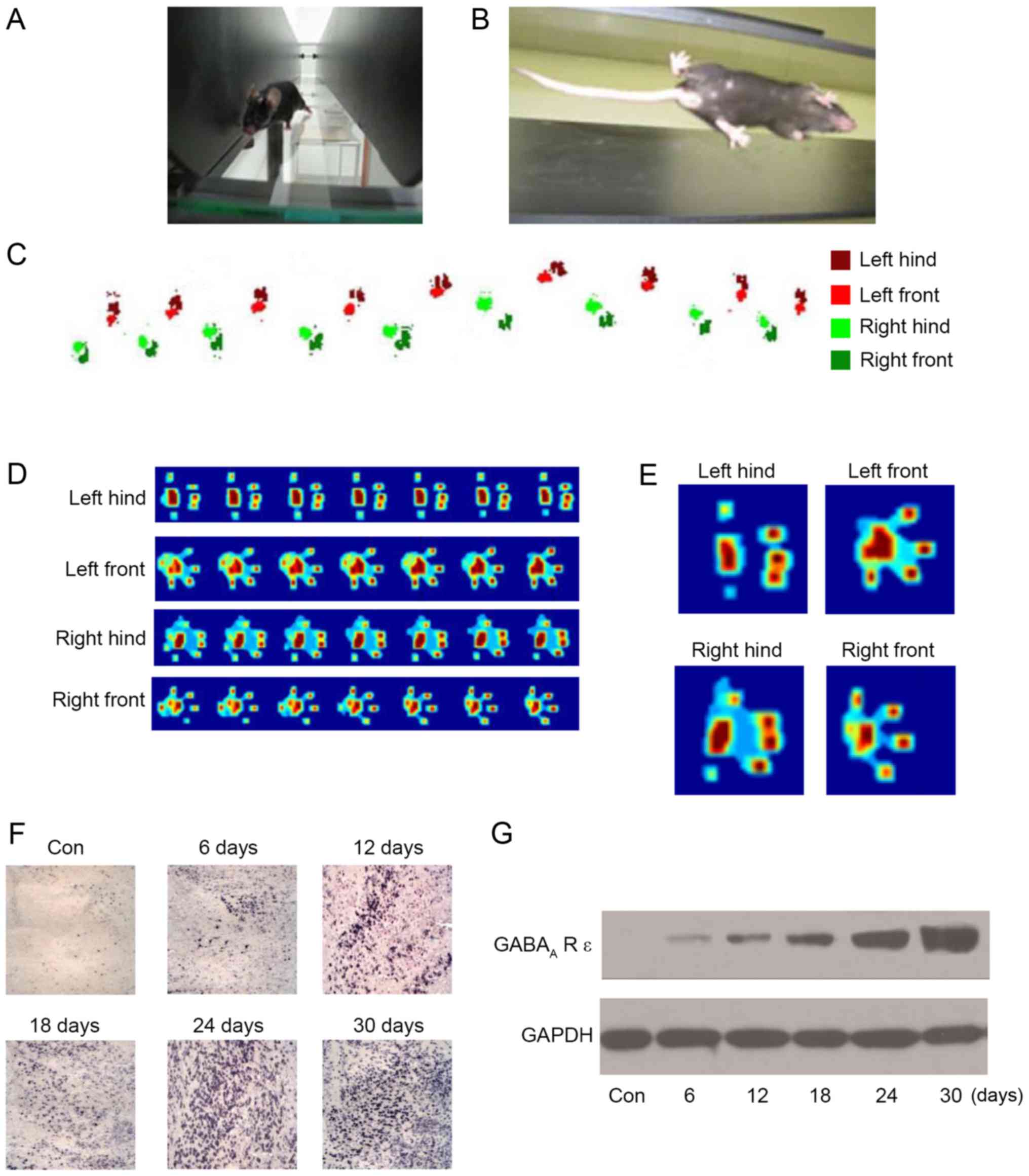
In order to evaluate the effect of sevoflurane on
motor function in neonatal mice, the CatWalk Automatic Gait
Analyzer (Fig. 3A and B) was used
to quantitatively analyze the gait of mice following repeated
inhalation of sevoflurane. The four feet were labeled with
different colors (Fig. 3C), and
the footprints were recorded. Additionally, the footprints were
converted to a heat map, in order for the pressure at different
sites to be determined (Fig. 3D and
E). The results (Table I)
demonstrated that the gait of the mice was abnormal following 6
days of repeated anesthesia and that the abnormality was more
prominent with the increase in repeated anesthesia time, which
primarily manifested in the support phase, swing phase,
coordination, footprint average area, braking index and propulsion
index. In addition, following 24 days of repeated anesthesia, the
gait movement function of the mice was reduced to its worst state;
continuing to increase the days of repeated anesthesia did not lead
to a significant alteration in gait movement function. The results
of the present study demonstrated that repeated inhalation of
sevoflurane may lead to movement behavior disorder in neonatal
mice. Gait indexes were negatively associated with repeated
anesthesia time, which may explain the long-term alterations in
motor function in neonatal mice.
 | Table I.Effect of repeated inhalation of
sevoflurane on the movement behavior of wild-type neonatal
mice. |
Table I.
Effect of repeated inhalation of
sevoflurane on the movement behavior of wild-type neonatal
mice.
| Group | Coordination of
right rear to left front | Ipsilateral
coordination of right front to left front | Swing phase-left
front, sec | Swing phase-right
front, sec | Support phase-left
front, sec | Support phase-right
front, sec | Propulsion index of
left rear | Propulsion index of
left front | Braking index of
left rear | Average area of
right rear footprint, mm2 |
|---|
| Control, n=20 |
0.521±0.074 |
0.573±0.062 |
0.641±0.072 |
0.627±0.059 |
0.315±0.083 |
0.326±0.083 |
0.817±0.115 |
0.571±0.084 |
0.219±0.025 |
2.741±0.413 |
| 6 days, n=20 |
0.463±0.042a |
0.491±0.059a |
0.583±0.058a |
0.542±0.074a |
0.391±0.091a |
0.408±0.062a |
0.742±0.104a |
0.469±0.075a |
0.294±0.053a |
3.915±0.619a |
| 12 days, n=20 |
0.375±0.035a |
0.411±0.058a |
0.491±0.049a |
0.451±0.084a |
0.458±0.071a |
0.471±0.085a |
0.638±0.084a |
0.421±0.058a |
0.357±0.061a |
5.882±0.713a |
| 18 days, n=20 |
0.321±0.047a |
0.324±0.032a |
0.418±0.047a |
0.374±0.037a |
0.527±0.063a |
0.541±0.097a |
0.519±0.078a |
0.349±0.038a |
0.411±0.075a |
8.621±0.914a |
| 24 days, n=20 |
0.262±0.026a |
0.247±0.027a |
0.337±0.046a |
0.261±0.039a |
0.584±0.093a |
0.604±0.071a |
0.457±0.062a |
0.271±0.049a |
0.472±0.085a |
9.884±0.717a |
| 30 days, n=20 |
0.237±0.018a |
0.223±0.016a |
0.329±0.048a |
0.253±0.041a |
0.592±0.101a |
0.613±0.064a |
0.441±0.072a |
0.269±0.062a |
0.483±0.092a |
10.027±0.913a |
Effect of repeated inhalation of
sevoflurane on the GABAA-R ε subunit
There have been reports that the GABAA-R
ε subunit may be regulated by sevoflurane in cortical neurons
(13,14). In order to investigate the role of
the GABAA-R ε subunit in neonatal mice undergoing
repeated inhalation of sevoflurane, the expression of the
GABAA-R ε subunit in cerebellar tissue was detected
using an anti-GABAA-R ε subunit antibody by
immunohistochemistry. The results indicated that the level of
expression of the GABAA-R ε subunit was increased by
repeated inhalation of sevoflurane in the PC layer, and the number
of GABAA-R ε subunit-positive PCs was increased with the
increase in the number of days of repeated anesthesia; similarly to
the effect of repeated inhalation of sevoflurane on movement
behavior and the field potential response of PCs, the increase was
stable at 24 days (Fig. 3F). In
addition, the results of the western blot analysis demonstrated
that the amount of GABAA-R ε subunit protein was
increased by the repeated inhalation of sevoflurane, and was
positively associated with the number of days of repeated
anesthesia (Fig. 3G). The results
of the present study indicated that the GABAA-R ε
subunit was upregulated by the repeated inhalation of sevoflurane,
and that the GABAA-R ε subunit may serve an important
role in neonatal mice receiving repeated inhalation of
sevoflurane.
Effects of repeated inhalation of
sevoflurane on neonatal mice regulated by the GABAA-R ε
subunit
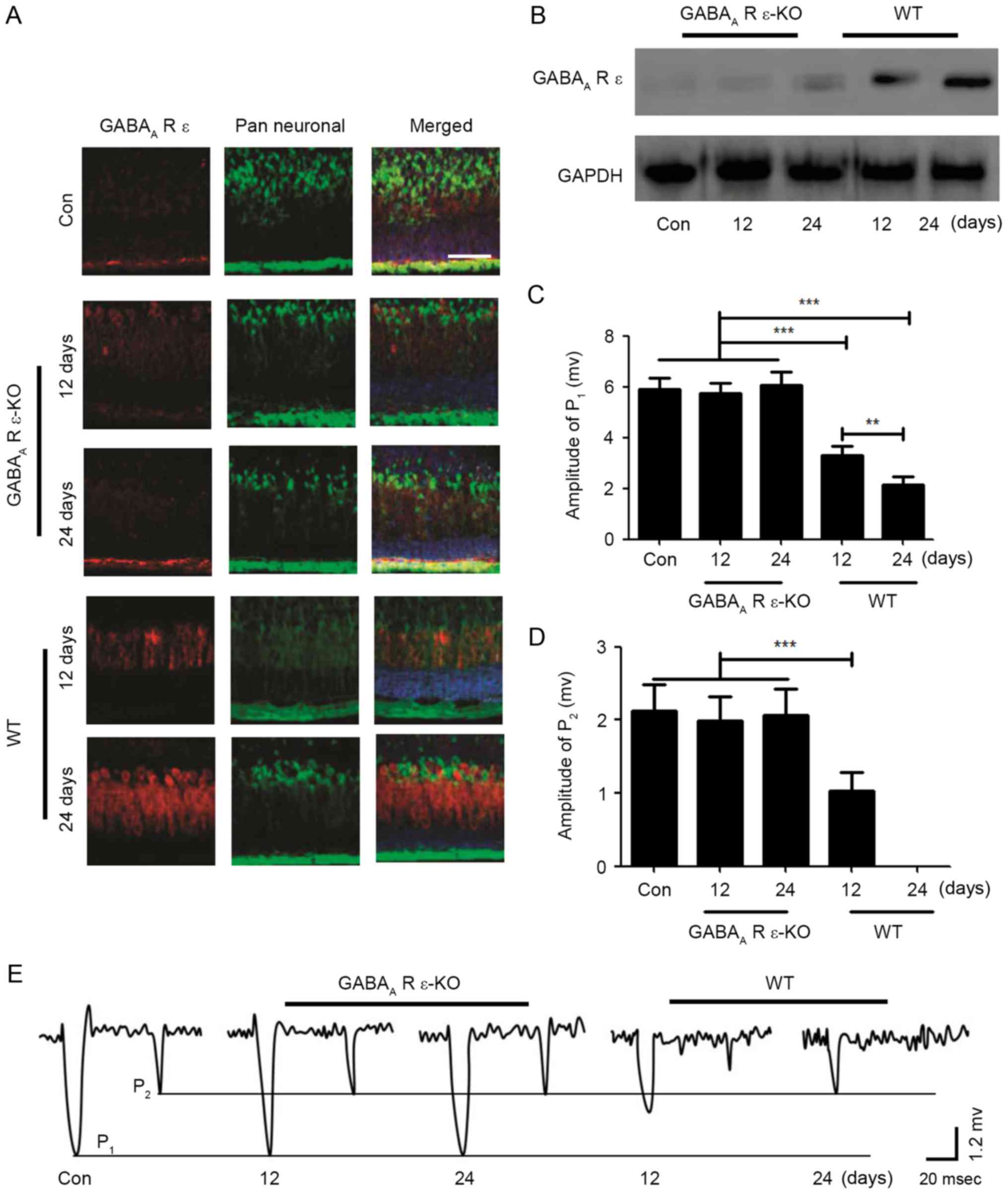
In order to determine whether the GABAA-R
ε subunit served a role in the effects of repeated inhalation of
sevoflurane in neonatal mice on the field potential response of PCs
and movement behavior, GABAA-R ε subunit knockout mice
were used in the present study. In the GABAA-R ε subunit
knockout mice, following repeated inhalation of sevoflurane for 12
or 24 days, compared with the control group, the expression level
of the GABAA-R ε subunit was not altered, whereas the
expression level of the GABAA-R ε subunit was markedly
increased in a time-dependent manner in the wild-type mice
(Fig. 4A); the results of the
western blot analysis confirmed this result (Fig. 4B). For the GABAA-R ε
subunit knockout neonatal mice, the mice were subjected to repeated
inhalation of sevoflurane for different lengths of time; when the
mice were 2 months old, the excitatory field potential responses of
PCs were induced by stimulation of the ipsilateral whisker pad in
the current clamp mode (I=0). The results indicated that the values
of P1 and P2 in GABAA-R ε subunit
knockout mice were consistent with the control group, although in
the wild-type mice, the values of P1 and P2
were significantly altered in a time-dependent manner (Fig. 4C-E). Additionally, the effect of
repeated inhalation of sevoflurane on motor function in the
GABAA-R ε subunit knockout mice was examined using the
CatWalk Automatic Gait Analyzer. The results demonstrated that
repeated inhalation of sevoflurane did not affect motor function in
the GABAA-R ε subunit knockout mice, although motor
function in wild-type mice was inhibited by repeated inhalation of
sevoflurane (Table II). These
results indicated that the GABAA-R ε subunit may serve
an important role in the field potential response of PCs and the
development of movement behaviors.
| Table II.Effect of repeated inhalation of
sevoflurane on the movement behavior of GABAA R ε
knockout and wild-type neonatal mice. |
Table II.
Effect of repeated inhalation of
sevoflurane on the movement behavior of GABAA R ε
knockout and wild-type neonatal mice.
| Group | Coordination of
right rear to left front | Ipsilateral
coordination of right front to left front | Swing phase-left
front, sec | Swing phase-right
front, sec | Support phase-left
front, sec | Support phase-right
front, sec | Propulsion index of
left rear | Propulsion index of
left front | Braking index of
left rear | Average area of
right rear footprint, mm2 |
|---|
| Control, n=20 |
0.547±0.092 |
0.552±0.093 |
0.633±0.114 |
0.636±0.113 |
0.323±0.077 |
0.337±0.064 |
0.804±0.121 |
0.563±0.076 |
0.226±0.032 |
2.512±0.375 |
| GABAA-R
ε knockout mice |
|
|
|
|
|
|
|
|
|
|
| 12 days, n=20 |
0.536±0.093 |
0.549±0.073 |
0.634±0.079 |
0.632±0.095 |
0.331±0.064 |
0.341±0.059 |
0.796±0.114 |
0.559±0.067 |
0.224±0.074 |
2.449±0.483 |
| 24 days, n=20 |
0.521±0.115 |
0.533±0.069 |
0.618±0.093 |
0.619±0.075 |
0.329±0.084 |
0.339±0.067 |
0.785±0.097 |
0.562±0.089 |
0.231±0.043 |
2.593±0.618 |
| Wild-type mice |
|
|
|
|
|
|
|
|
|
|
| 12 days, n=20 |
0.378±0.063a |
0.395±0.104a |
0.479±0.062a |
0.442±0.095a |
0.463±0.085a |
0.483±0.067a |
0.615±0.078a |
0.416±0.074a |
0.368±0.074a |
6.114±0.817a |
| 24 days, n=20 |
0.283±0.062a |
0.257±0.053a |
0.341±0.057a |
0.273±0.053a |
0.569±0.107a |
0.668±0.093a |
0.463±0.057a |
0.268±0.028a |
0.485±0.075a |
9.634±0.829a |
Discussion
The cerebellum is an important motor center;
external information is precision computed and synthesized by the
cerebellar cortex, and e PCs issue a variety of output instructions
to complete physiological activities (15). PCs are the principal neurons in the
cerebellar cortex, and their axons form the only efferent pathway
of the cerebellar cortex (16).
GABA is released via the axon terminals of PCs, which contribute to
the inhibitory effect of PCs on the dominant nucleated neurons of
the cerebellum (17). In the
afferent fibers of the cerebellum, climbing fibers and mossy fibers
may combine with PC dendrites to form excitatory synaptic
connections (18). Excitatory
afferents of climbing fibers may cause PC excitability,
characterized by complex spikes and, subsequent to the formation of
synaptic connection between the mossy fibers and granular cells, PC
excitability is indirectly excited through the axons of granulosa
cells and parallel fibers, and manifests as simple spikes (6,18).
Basket cells (BCs) stellate cells (SCs) are the inhibitory
interneurons of PCs; the long axon terminals of BCs are wrapped in
the soma and axon segments of 3–5 PCs (19). The excitations of BCs may directly
inhibit PCs via the soma, and the axons of SCs selectively combine
with the dendrites of PCs to form inhibitory synaptic connections,
contributing to the inhibition of PC dendrites and shunting the
excitatory afferents of parallel fibers (20,21).
It was previously suggested that one bundle of parallel fibers and
its dominant PCs are excited by sensory information through the
mossy fiber-granulosa-paracellular pathway (22). Subsequent studies demonstrated that
sensory stimulation may stimulate PCs, in addition to leading to PC
IPSP accompanied by discharge pause (23–25).
Sensory stimulation may induce PC excitation following blocking of
GABAA receptor activity, indicating that inhibitory
interneurons serve a role in the process of cerebellar cortex
sensory information transmission (11,26).
In the present study, it was demonstrated that stimulation to the
ipsilateral whisker pad may stimulate the high-fidelity response of
PCs in the crus II region of the cerebellum and as the stimulus
response at the beginning was greater than the stimulus response at
the end, it indicates that the sensory stimulation information was
preserved intact through mossy fiber-PC pathway transmission into
cerebellar cortex. The integrity of the stimulation beginning and
end information coding indicated that cerebellar PCs have a high
fidelity for the transmission of sensory information.
Experimental animal behavior analysis is widely used
in studies of motor function, learning, memory and other higher
central nervous system functions (27). Compared with the analysis of
electrophysiological, biochemical and other parameters,
experimental animal behavior analysis may fully reflect the overall
state of animals, and gait analysis is commonly used in the study
of motor behavior (28). The gait
of mice was detected using the CatWalk Gait Analysis system
following repeated inhalation of sevoflurane, which may effectively
reflect the severity of brain injury and movement functions, and
guarantee objectivity (29).
Therefore, the CatWalk Gait Analysis system may be used as a method
to evaluate animal behavior in the repeated inhalation of
sevoflurane model. The parameters associated with cerebellar injury
in gait analysis are: Ipsilateral coordination, contralateral
coordination, support phase, swing phase, propulsion index, braking
index and average footprint (30).
In the present study, the results indicated that the gait analysis
indexes were significantly associated with cerebellar injury in
mice undergoing repeated inhalation of sevoflurane with different
time. When the anesthesia time increased, the gait analysis indexes
deteriorated, which indicated that the cerebellar injury was
positively associated with the length of time of repeated
inhalation of sevoflurane.
In the present study, the results indicated that
repeated inhalation of sevoflurane exerted inhibitory effects on
cerebellar sensory information transmission and motor function
development, and there was a certain dose-dependent effect;
however, long-time repeated inhalation of sevoflurane was unable to
completely inhibit the sensory stimulation-induced response of PCs
and motor function development. It was additionally demonstrated
that the GABAA-R ε subunit was significantly activated,
suggesting that repeated inhalation of sevoflurane may inhibit the
transmission of sensory information in PCs and motor function
development by enhancing GABAA-R ε subunit activity. In
the GABAA-R ε subunit knockout mice, the inhibitory
effects of repeated inhalation of sevoflurane on the transmission
of sensory information in PCs and motor function development were
completely inhibited. The results of the present study indicated
that the GABAA-R ε subunit served a role in the field
potential response of PCs and the development of movement
behavior.
The results of the present study may be associated
with a number of factors. Repeated inhalation of sevoflurane may
affect central nervous system function by regulating receptor and
neurotransmitter transmittance, contributing to GABAA-R
ε subunit activation (31).
Therefore, the repeated inhalation of sevoflurane may directly
activate the GABAA-R ε subunit in PCs, which may result
in a sustained inhibition of the field potential response in the
PCs, contributing to inhibition of information transmission and the
development of motor function. Increasing the concentration of
sevoflurane in the cerebellum may synergize with the endogenous
GABAA-R ε subunit, thereby enhancing the inhibitory
effect of the endogenous GABAA-R ε subunit on the field
potential response of PCs, delaying or inhibiting the transmission
of movement information and, consequently, the development of motor
function (32). Sevoflurane
increases the excitability of Golgi cells by inhibiting the
activity of Na+/K+ ATPase, thereby enhancing
the paroxysmal inhibition of PCs and increasing the leakage of the
GABAA-R ε subunit around PCs, resulting in an increase
in the GABAA-R ε subunit concentration in the cerebellum
and further increasing the sustained inhibition of information
transmission in PCs, affecting motor function and development
(33–35). Therefore, an increase in
sevoflurane concentration in the cerebellar cortex may excite Golgi
cells, increase the release of the GABAA-R ε subunit,
increase the concentration of GABAA-R ε subunits around
the PCs, and enhance the sustained inhibitory effect on the sensory
stimulus-induced responses of PCs and the transmission of motion
information, delaying motor function development.
In conclusion, repeated inhalation of sevoflurane
exerted an effect on GABAA-R ε subunit activity in
neonatal mice. The field potential response of the PCs was markedly
inhibited by sevoflurane through GABAA-R ε subunit
activation. Sevoflurane was additionally able to inhibit sensory
information transmission in the cerebellar cortex, further delaying
the cerebellar motor function development. These results
demonstrated that repeated inhalation of sevoflurane may damage the
cerebellum, and revealed a partially dose-dependent effect,
suggesting that repeated inhalation of anesthesia may require
careful consideration in children; GABAA-R ε subunit
antagonists may be applied in combination, in order to reduce the
incidence of side-effects associated with repeated inhaled
anesthesia.
References
|
1
|
Brandt ML, Harmon CM, Helmrath MA, Inge
TH, McKay SV and Michalsky MP: Morbid obesity in pediatric diabetes
mellitus: Surgical options and outcomes. Nat Rev Endocrinol.
6:637–645. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mallineni SK and Yiu CK: Dental treatment
under general anesthesia for special-needs patients: Analysis of
the literature. J Investig Clin Dent. 7:325–331. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parekh S, Gardener C, Ashley PF and Walsh
T: Intraoperative local anaesthesia for reduction of postoperative
pain following general anaesthesia for dental treatment in children
and adolescents. Cochrane Database Syst Rev. 12:CD0097422014.
|
|
4
|
Stratmann G: Review article: Neurotoxicity
of anesthetic drugs in the developing brain. Anesth Analg.
113:1170–1179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hirata Y, Katagiri K and Tanaka Y: Direct
causality between single-Purkinje cell activities and motor
learning revealed by a cerebellum-machine interface utilizing VOR
adaptation paradigm. Cerebellum. 11:455–456. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barmack NH and Yakhnitsa V: Climbing
fibers mediate vestibular modulation of both ‘complex’ and ‘simple
spikes’ in Purkinje cells. Cerebellum. 14:597–612. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang JJ, Yen CT, Tsao HW, Tsai ML and
Huang C: Neuronal oscillations in Golgi cells and Purkinje cells
are accompanied by decreases in Shannon information entropy.
Cerebellum. 13:97–108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rostain JC, Lavoute C, Risso JJ, Vallée N
and Weiss M: A review of recent neurochemical data on inert gas
narcosis. Undersea Hyperb Med. 38:49–59. 2011.PubMed/NCBI
|
|
9
|
Irie T, Kikura-Hanajiri R, Usami M,
Uchiyama N, Goda Y and Sekino Y: MAM-2201, a synthetic cannabinoid
drug of abuse, suppresses the synaptic input to cerebellar Purkinje
cells via activation of presynaptic CB1 receptor.
Neuropharmacology. 95:479–491. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Walls AB, Waagepetersen HS, Bak LK,
Schousboe A and Sonnewald U: The glutamine-glutamate/GABA cycle:
Function, regional differences in glutamate and GABA production and
effects of interference with GABA metabolism. Neurochem Res.
40:402–409. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kueh SL, Dempster J, Head SI and Morley
JW: Reduced postsynaptic GABAA receptor number and enhanced
gaboxadol induced change in holding currents in Purkinje cells of
the dystrophin-deficient mdx mouse. Neurobiol Dis. 43:558–564.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gueli SL and Lerman J: Controversies in
pediatric anesthesia: Sevoflurane and fluid management. Curr Opin
Anaesthesiol. 26:310–317. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ando N, Sugasawa Y, Inoue R, Aosaki T,
Miura M and Nishimura K: Effects of the volatile anesthetic
sevoflurane on tonic GABA currents in the mouse striatum during
postnatal development. Eur J Neurosci. 40:3147–3157. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ishizeki J, Nishikawa K, Kubo K, Saito S
and Goto F: Amnestic concentrations of sevoflurane inhibit synaptic
plasticity of hippocampal CA1 neurons through gamma-aminobutyric
acid-mediated mechanisms. Anesthesiology. 108:447–456. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cerminara NL, Lang EJ, Sillitoe RV and
Apps R: Redefining the cerebellar cortex as an assembly of
non-uniform Purkinje cell microcircuits. Nat Rev Neurosci.
16:79–93. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Marcaggi P: Cerebellar endocannabinoids:
Retrograde signaling from purkinje cells. Cerebellum. 14:341–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Valenzuela CF and Jotty K: Mini-review:
Effects of ethanol on GABAA receptor-mediated neurotransmission in
the cerebellar cortex-recent advances. Cerebellum. 14:438–446.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barmack NH and Yakhnitsa V: Topsy turvy:
Functions of climbing and mossy fibers in the vestibulo-cerebellum.
Neuroscientist. 17:221–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Whitney ER, Kemper TL, Rosene DL, Bauman
ML and Blatt GJ: Density of cerebellar basket and stellate cells in
autism: Evidence for a late developmental loss of Purkinje cells. J
Neurosci Res. 87:2245–2254. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu H, Zhao SN, Zhao GY, Sun L, Chu CP and
Qiu DL: N-methyl-d-aspartate inhibits cerebellar Purkinje cell
activity via the excitation of molecular layer interneurons under
in vivo conditions in mice. Brain Res. 1560:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lennon W, Hecht-Nielsen R and Yamazaki T:
A spiking network model of cerebellar Purkinje cells and molecular
layer interneurons exhibiting irregular firing. Front Comput
Neurosci. 8:1572014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hashimoto K, Yoshida T, Sakimura K,
Mishina M, Watanabe M and Kano M: Influence of parallel
fiber-Purkinje cell synapse formation on postnatal development of
climbing fiber-Purkinje cell synapses in the cerebellum.
Neuroscience. 162:601–611. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng N and Raman IM: Synaptic inhibition,
excitation, and plasticity in neurons of the cerebellar nuclei.
Cerebellum. 9:56–66. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tanaka S, Kawaguchi SY, Shioi G and Hirano
T: Long-term potentiation of inhibitory synaptic transmission onto
cerebellar Purkinje neurons contributes to adaptation of
vestibulo-ocular reflex. J Neurosci. 33:17209–17220. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gmaz JM and McKay BE: Toluene decreases
Purkinje cell output by enhancing inhibitory synaptic transmission
in the cerebellar cortex. Neurosci Lett. 560:1–6. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ono Y, Saitow F and Konishi S:
Differential modulation of GABAA receptors underlies postsynaptic
depolarization- and purinoceptor-mediated enhancement of cerebellar
inhibitory transmission: A non-stationary fluctuation analysis
study. PLoS One. 11:e01506362016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Batchelor PE, Skeers P, Antonic A, Wills
TE, Howells DW, Macleod MR and Sena ES: Systematic review and
meta-analysis of therapeutic hypothermia in animal models of spinal
cord injury. PLoS One. 8:e713172013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cimolin V and Galli M: Summary measures
for clinical gait analysis: A literature review. Gait Posture.
39:1005–1010. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Parvathy SS and Masocha W: Gait analysis
of C57BL/6 mice with complete Freund's adjuvant-induced arthritis
using the CatWalk system. BMC Musculoskelet Disord. 14:142013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bozkurt A, Scheffel J, Brook GA, Joosten
EA, Suschek CV, O'Dey DM, Pallua N and Deumens R: Aspects of static
and dynamic motor function in peripheral nerve regeneration: SSI
and CatWalk gait analysis. Behav Brain Res. 219:55–62. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stojanovic T, Capo I, Aronica E,
Adle-Biassette H, Höger H, Sieghart W, Kovacs GG and Milenkovic I:
The α1, α2, α3, and γ2 subunits of GABAA receptors show
characteristic spatial and temporal expression patterns in
rhombencephalic structures during normal human brain development. J
Comp Neurol. 524:1805–1824. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li ZX, Yu HM and Jiang KW: Tonic GABA
inhibition in hippocampal dentate granule cells: Its regulation and
function in temporal lobe epilepsies. Acta Physiol (Oxf).
209:199–211. 2013.PubMed/NCBI
|
|
33
|
Groundwater PW, Hamid K, Ng I,
Tallapragada VJ, Hibbs DE and Hanrahan J: The differential effects
of resveratrol and trans-ε-viniferin on the GABA-induced current in
GABAA receptor subtypes expressed in xenopus laevis oocytes. J
Pharm Pharm Sci. 18:328–338. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chou WH, Wang D, McMahon T, Qi ZH, Song M,
Zhang C, Shokat KM and Messing RO: GABAA receptor trafficking is
regulated by protein kinase C(epsilon) and the
N-ethylmaleimide-sensitive factor. J Neurosci. 30:13955–13965.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Eisinger BE, Zhao C, Driessen TM, Saul MC
and Gammie SC: Large scale expression changes of genes related to
neuronal signaling and developmental processes found in lateral
septum of postpartum outbred mice. PLoS One. 8:e638242013.
View Article : Google Scholar : PubMed/NCBI
|