Introduction
The mechanism that triggers childbirth is complex
and remains to be fully elucidated. Previous studies have
demonstrated that parturition involves interactions between various
processes, factors, regulatory pathways and molecules, which
comprise multi-stage changes (1,2).
Various theories regarding the mechanisms of the triggering of
childbirth, including neurotransmission, mechanical changes,
endocrine control, cervical maturity and lower uterine segment
formation, in addition to immunological theories, have been
proposed. It is widely accepted that normal childbirth relies on
the rhythmic contraction of the uterine smooth muscle and
progressive expansion of the cervix. Additionally, various tissues,
organs and factors are involved in the process of parturition; this
includes endocrine and paracrine signaling by the mother and the
fetus, involving hormones secreted from the placenta, fetal
membranes, uterus, decidua and other adjacent tissues, cytokines
produced in the uterus, and smooth muscle cell membrane receptors.
Alterations in these signaling mechanisms ultimately induce
contraction of the uterine smooth muscle, leading to parturition
(3–6). This evidence suggests that uterine
smooth muscle contraction is a crucial trigger of the childbirth
process.
An intracellular calcium signaling mechanism has
been proposed that induces periodic uterine contractions; this
mechanism involved binding between hormone and receptor, receptor
and G protein activation, and activation of adenylyl cyclase and
phospholipase C (PLC). Adenylyl cyclase catalyzes the production of
cyclic adenosine monophosphate (cAMP) from adenosine triphosphate
(ATP), resulting in the activation of cAMP-dependent protein
kinases. PLC on the membrane surface catalyzes the formation of
diacylglycerol (DAG) and inositol triphosphate (IP3)
from phosphatidylinositol-4,5-bisphosphate (PIP2). DAG
activates protein kinase C (PKC), and PKC subsequently activates
voltage-dependent calcium channels, leading to extracellular
calcium influx. IP3 binds with the IP3
receptor on the endoplasmic reticulum (ER) surface, which induces
the release of calcium from the ER to increase cytoplasmic calcium
levels. Calcium binds with calmodulin, which causes activation of
myosin light chain (MLC) kinase, and phosphorylation of the head of
the MLC filament; this facilitates the contraction of smooth
muscle. Increasing intracellular calcium often depends on the
transmembrane transfer of extracellular calcium into cells, which
can trigger further release of calcium from intracellular stores.
Ca2+ uptake and release by calcium channels in the
plasma membrane and intracellular sarcoplasmic reticulum (SR)
mediate intracellular calcium homeostasis. Thus, calcium channels
serve a crucial role in the regulation of smooth muscle activity.
Under normal circumstances, extracellular Ca2+
predominantly enters cells through calcium channels in the cell
membrane, which triggers further Ca2+ release from the
SR, leading to uterine contractions.
Classical transient receptor potential (TRP)
channels, as six-transmembrane non-selective cation channels,
participate in numerous physiological and pathological processes,
including cell proliferation, angiogenesis, neuronal morphogenesis
and synaptogenesis, and tumor formation. Sodium, calcium and
magnesium ions can enter cells through canonical TRP (TRPC)
channels, which respond to diverse external stimuli. Human cells
express six TRPC isoforms: TRPC1, 3, 4, 5, 6 and 7 (TRPC2 is a
pseudogene in humans). The TRPC family can be classified into four
subgroups according to structural homology and functional
predisposition as follows: TRPC1; TRPC2; TRPC4 and 5; and TRPC3, 6
and 7 (7–10). TRPC channels are expressed during
embryonic development and adulthood. Previous studies have detected
the expression of TRPC channels in the mammalian myometrium, and
mechanical contraction of human uterine smooth muscle has been
demonstrated to cause upregulation of TRPC3 expression.
There are two mechanisms that activate TRP channels
resulting in calcium influx: Store-operated calcium entry (SOCE)
and receptor-operated calcium entry (ROCE). TRP channels can be
activated by G protein-coupled receptors and tyrosine kinase
receptors, or PLCβ and PLCγ. Following activation of a
phosphatidylinositol-specific or PLC-specific G protein-coupled
receptor, or a PLCγ-specific receptor tyrosine kinase, PLC is
induced to hydrolyze PIP2 into IP3 and DAG
(11,12). IP3 acts on the ER or SR,
leading to the release of Ca2+. In SOCE (formally known
as capacitative calcium entry), when calcium stores are depleted,
TRPC channels on the cell membrane open to allow an influx of
extracellular Ca2+. By contrast, ROCE is mediated by
DAG, which directly activates TRPC channel opening.
PKC is a family of Ca2+−activated
phospholipid-dependent serine/threonine protein kinases that are
widely distributed in various organs. PKCs regulate processes in
various cell types, including gene expression, cell proliferation,
apoptosis and cell migration (13). The PKC/PKC-potentiated phosphatase
inhibitor protein of 17 kDa (CPI-17) signaling pathway serves a
critical role in the calcium sensitization of smooth muscle.
CPI-17, a key substrate of PKC, is a PKC-dependent phosphatase
inhibitor that inhibits the enzymatic activity of myosin light
chain phosphatase (MLCP) (14,15).
Su et al (16) silenced
CPI-17 gene expression in bronchial smooth muscle using RNA
interference technology, which decreased bronchial smooth muscle
calcium sensitization, contraction frequency and contractility,
suggesting that CPI-17 affects the contraction of smooth muscle.
Jiang et al (17) reported
that PKC agonists significantly increased the phosphorylation of
myosin light chain (MLC) and the contraction of vascular smooth
muscle. Due to specific MLCP inhibition, exogenous phosphorylated
CPI-17 increases vascular smooth muscle contraction in a
dose-dependent manner. These results suggest that PKC activation
inhibits MLCP activity by inducing CPI-17 phosphorylation, thereby
increasing MLC20 phosphorylation and the calcium sensitivity of
contractile proteins, leading to vascular and bronchial smooth
muscle contraction (13,18).
TRPC channels can mediate extracellular calcium
influx and induce uterine contraction, and additionally exert
extensive physiological effects in the nervous system, vascular
smooth muscle, myocardium and skin. L-type and T-type calcium
channels can enhance the function of TRPC3 channels and can promote
the contraction of uterine spiral arteries in pregnant mice during
parturition. PLCγ can activate TRPC channels and PKC, and the
PKC/CPI-17 pathway serves an important role in vascular smooth
muscle contraction and calcium sensitization. Therefore, it was
hypothesized that TRPC3 triggers parturition via L-type and/or
T-type calcium channels, and that the PLCγ/PKC/CPI-17 pathway is
the major regulator involved in inducing parturition.
In the present study, uterine samples were collected
from patients that experienced preterm delivery with or without
labor onset, full-term delivery with labor onset, full-term
delivery without labor onset, and from a non-pregnant control
group. The effects of TRPC3 on labor onset and on calcium channel
expression were detected by immunohistochemistry and western
blotting. Additionally, pregnant mice were treated with either a
TRPC3 inhibitor or agonist, and uterine samples were harvested at
different time-points in an attempt to investigate the underlying
role of TRPC3 in parturition and its influence on PLCγ/PKC/CPI-17
signaling. Uterine smooth muscle cells from pregnant mice were
isolated, cultured and treated with TRPC3 inhibitors, then the
expression of factors upstream and downstream of TRPC3 were
detected to determine the mechanisms of PLCγ/PKC/CPI-17 in the
regulation of TRPC3 functions in uterine smooth muscle cells from
pregnant mice. The results may provide evidence that could be
useful for the prevention and treatment of premature
childbirth.
Materials and methods
Ethics statement
The present study was approved by the Clinical
Research Ethics Committee of Shenyang Hospital of China Medical
University (Shenyang, China). Written informed consent was obtained
from all patients.
Tissue and serum collection
A total of 80 tissue samples (4 groups, 20 samples
per group: Preterm delivery with or without labor onset, full-term
delivery with labor onset, full-term delivery without labor onset
and non-pregnant controls) were obtained from patients who
underwent surgery [caesarean section (pregnant groups) or treatment
for cervical intraepithelial neoplasia III (non-pregnant control
group)] at Shenyang Hospital of China Medical University between
October 2015 and June 2016. Full-term delivery was defined as
parturition at 38–40 weeks of gestation, and labor onset was
defined as the presence of regular contractions and/or cervical
dilation of 2–3 cm. All collected tissue samples were immediately
snap-frozen in liquid nitrogen and stored at −80°C. After placentas
were collected from the pregnant women, whole blood samples were
taken from the placenta and subjected to serum separation by
centrifugation, then stored at a low temperature.
Mouse strains
A total of 80 adult female Kunming mice were
subjected to timed matings. The morning that vaginal plugs were
detected was designated as day 0 of pregnancy. Mice were housed in
a 25°C temperature room with alternating 12-hour light and dark
cycles and had free access to food and water.
Murine preterm labor model
Mice were randomly divided into an unfertilized
group (group A), an ‘infected’ [lipopolysaccharides (LPS)-treated]
preterm group (group B), a 15-day gestation (preterm cesarean
section group (group C), and a full-term cesarean section group
(group D).
At day 15, the pregnant mice in group B received
four injections of LPS in salt solution (350 µg/kg) into the celiac
artery, at intervals of 3 h. In all the groups, mice were injected
with a TRPC3 channel inhibitor SKF96365 (ab120280; Abcam,
Cambridge, UK) or saline (1 h after the injection of group B with
LPS) and the litter delivery was observed and recorded. The mice
were closely observed 24 h/day to record premature deliveries
(i.e., parturition earlier than at day 19). When parturition
occurred in the pregnant mice (full delivery of first offspring),
the uterine tissue was immediately obtained; 10% chloral hydrate
(dose 0.03 ml/kg) was used for anesthetization and the mice were
then sacrificed by cervical spinal dislocation. The obtained
uterine and adipose tissues were irrigated with saline solution and
stored at −80°C.
Plasmid and small interfering RNA
(siRNA) transfection
A TRPC3 construct was generated by subcloning the
TRPC3 coding sequence into a pcDNA3.0 vector using standard
subcloning protocols. In addition, synthetic siRNAs of 17- or
19-nucleotide duplex RNA and 2-nucleotide 3′-dTdT overhangs were
designed and obtained from Guangzhou RiboBio Co., Ltd. (Guangzhou,
China). The green fluorescence-tagged sequences of siRNAs used in
the present study were as follows: Control,
5′-UUCUCCGAACGUGUCACGU-3′; mouse TRPC3-1,
5′-AGCCUUCUGUGCUGAGAAC-3′; and mouse TRPC3-2,
5′-GGACCGCAAAGUGUUUGUG-3′. In order to silence the expression of
TRPC3, cells at 60–70% confluence were seeded overnight and
transfected with either 5 µl of 20 µM siRNA (final concentration 50
nM) against TRPC3 or control siRNA (non-target scramble siRNA) with
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) in OptiMEM (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Transfection was
confirmed as following 3 days, green fluorescence was observable
under the fluorescence microscope. Cells were used in subsequent
assays 48 h following transfection.
Confocal microscopy
Uterine smooth muscle cells were fixed in confocal
chambers using 6% formaldehyde for 15 min at room temperature,
followed by permeabilization with 0.1–0.2% Triton X-100 for 20 min.
After washing, the cells were incubated with the appropriate
primary antibody [TRPC3 (rabbit, 25 kDa; 1:1,000; cat. no. ab51560;
Abcam, Cambridge, UK), PLCγ (rabbit, 1:1,000; Sigma-Aldrich; Merck
KGaA; cat. no. 05-163), PKC (rabbit, 1:1,000; Abcam; cat. no.
ab31), CPI-17 (rabbit, 17 kDa; 1:1,000; Abcam; cat. no. ab131451),
P-CPI-17 (rabbit, 17 kDa, 1:1,000; Abcam; cat. no. ab52174), Cav1.2
(rabbit, 239 kDa, 1:1,000; Abcam; cat. no. ab58552), Cav3.1
(rabbit, 262 kDa, 1:1,000; Abcam; cat. no. ab203577), Cav3.2
(rabbit, 262 kDa; 1:1,000; Abcam; cat. no. ab128251)] at 4°C
overnight followed by incubation at room temperature for 1 h with a
secondary antibody (1:1,000; Abcam; cat. no. ab6721). Confocal
images were obtained using a Zeiss LSM510 laser scanning microscope
(Zeiss, Oberkochen, Germany) with single (488 nm) or multitrack
sequential excitation (488 and 633 nm), and emission [515-540 nm,
boron dipyrromethene fluorescein; 650 nm, Alexa Fluor 633] filter
sets (Chroma Technology Corporation, Bellow Falls, VT, USA). The
color of 488 nm excitation was set as pseudo green, and the color
for 633 nm excitation line was set as pseudo red. The merge of
these two colors (pseudo green and pseudo red) yielded yellow when
colocalization of two proteins occurred.
Western blotting
Samples from the four patient groups were ground
using a pestle and mortar and then resuspended in PBS (pH 7.4)
containing a protease inhibitor cocktail (Roche Diagnostics, Castle
Hill, Australia). The proteins were centrifuged (3,000 × g, 4°C, 5
min) and the supernatant was separated from the pellet then placed
on ice. The pellet was snap frozen and reprocessed as stated above,
and supernatants from each group were pooled and centrifuged
(20,000 × g, 4°C, 1 h). Protein extracts were aliquoted, rapidly
frozen in liquid nitrogen, and stored at −80°C. A Bradford protein
assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used
to determine the protein concentration in samples.
Protein extracts were dissolved in lysis buffer with
a protease inhibitor cocktail, and then separated by 10% SDS-PAGE
and electroblotted onto PVDF membranes for 2 h (80 V) at 4°C. PVDF
membranes were then washed with PBS, blocked and probed with
anti-TRPC3 (rabbit, 25 kDa; cat. no. ab51560; 1:1,000; Abcam)
anti-p38 MAPK antibody (cat. no. ab197348; 1:1,000; Abcam) and
anti-ERK 1/2 (phospho-Thr202/Tyr204; cat. no. ab214362; 1:1,000;
Abcam) antibody overnight at 4°C. Specific binding was visualized
using alkaline phosphate-conjugated secondary antibodies (Abgent;
1:1,000; cat. no. ARS3337) and chemiluminescence, according to
manufacturer's protocol (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). MagicMark XP Western Protein Standard
(Invitrogen; Thermo Fisher Scientific, Inc.) was used to estimate
the migrated protein band sizes. The intensity of bands
corresponding to protein expression was determined using Photoshop
CS3 Extended software (Adobe Systems, Inc., San Jose, CA, USA).
Relative levels of protein expression were determined by
normalizing the intensity of TRPC3 staining on the blots to avoid
potential changes in SMC protein expression in pregnancy, with a
GAPDH primary antibody (mouse, 37 kDa, 1:5,000; cat. no. sc-25778;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) used as the
loading control. Notably, extensive attempts to undertake western
blot protocols using antibodies against L-type and T-type calcium
channel subunits Cav1.2 (rabbit, 239 kDa, 1:1,000;
Abcam; cat. no. ab58552), Cav3.1 (rabbit; 262 kDa;
1:1,000; Abcam; cat. no. ab203577), Cav3.2 (rabbit, 262
kDa; 1:1,000; Abcam; cat. no. ab128251) antibodies to detect
proteins at the respective molecular weights.
Cell isolation and primary culture of
mouse myometrial smooth muscle cells (mSMCs)
Strips of longitudinal muscle (10–20 mm long) were
cut from the serosal surface of the uterine horn of mouse uteri,
carefully excluding the endometrium and circular muscle, and finely
cut into 2–3 mm-long pieces in ice-cold HBSS and then incubated
with gentle shaking for 40 min at 37°C in 5 ml HBSS containing 1.5
mM CaCl2, collagenase type IA (2 mg/ml; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany), trypsin inhibitor (2 mg/ml;
soybean, type II-S; Sigma-Aldrich; Merck KGaA), and bovine serum
albumin (6 mg/ml; Sigma-Aldrich; Merck KGaA). The cell suspension
was then triturated 15 to 20 times, filtered through a 40-mm
sterile filter, and washed twice in minimum essential medium (MEM;
Gibco; Thermo Fisher Scientific, Inc.) containing 10% fetal calf
serum (FCS; Gibco; Thermo Fisher Scientific, Inc.) with
centrifugation (450 × g, 5 min, 20°C). The cell pellet was
suspended in MEM supplemented with 5% FCS, penicillin (25 U/ml) and
streptomycin (25 U/ml); (Gibco; Thermo Fisher Scientific, Inc.).
For imaging protocols, a cell solution (400 ml) was placed on glass
coverslips and cells were allowed to adhere for ~1 h, after which
the coverslips were flooded with an additional 1.6 ml of culture
medium. For western blot analysis, myocytes were seeded into
25-cm3 culture flasks. Myocytes were incubated at 37°C
in a humidified atmosphere of 95% air/5% CO2 and
maintained as a primary culture until confluent (4 days on
average). The culture medium was changed every 2–3 days. To assess
the purity of myocyte cultures, immunocytochemistry was performed
using α-actin (Sigma-Aldrich; Merck KGaA) and calponin (Abcam)
monoclonal antibodies.
Uterine smooth muscle tissue samples from the
preterm delivery, full-term delivery with labor onset, full-term
delivery without labor onset, and non-pregnant control groups were
placed in sterile petri dishes, and rinsed 2–3 times with saline
solution. Fibrous connective tissue was then removed: The samples
were cut into small pieces and placed in a small 50-ml beaker, then
subjected to trypsin and collagenase I digestion to purify the
fibroblasts and smooth muscle cells according to differences in
their adherence times; purification was performed 2–3 times.
Immunohistochemistry
The distributions of TRPC3, Cav1.2,
Cav3.1, and Cav3.2 were examined in the
smooth muscle tissues from the four groups of human uterine samples
using conventional whole-mount immunohistochemistry and confocal
microscopy. Fixed whole-mount tissues were incubated in blocking
buffer (PBS containing 1% bovine serum albumin and 0.2% Tween-20;
Sigma-Aldrich; Merck KGaA) for 2 h at room temperature, rinsed (3–5
min) in PBS, and incubated with primary antibodies against TRPC3,
Cav1.2, Cav3.1 or Cav3.2 (1:100;
Abcam) in blocking buffer for 18 h at 4°C. Tissue was then rinsed
(3–5 min) in PBS and incubated in secondary antibody (Alexa Fluor
633; Invitrogen; Thermo Fisher Scientific, Inc.) diluted in PBS
containing 0.01% Tween-20 for 2 h, then rinsed again (3–5 min) in
PBS, and mounted in buffered antifade glycerol (AppliChem, GmbH,
Darmstadt, Germany). Tissues were examined with a confocal
microscope (FV1000; Olympus Corporation, Tokyo, Japan) and images
were acquired with uniform settings.
Immunofluorescent staining of
myometrial tissue
Uterine tissue was obtained from pregnant and
unfertilized mice, placed in optimal cutting temperature (Beijing
Solarbio Science & Technology Co., Ltd., Beijing) medium and
frozen. Frozen tissue sections were rehydrated and treated for 30
min with 1X PBS/0.25% Triton X-100 solution, and subsequently
rinsed in PBS twice for 5 min. Tissue sections were then incubated
for 40 min in undiluted Sea Block Blocking Solution (Thermo Fisher
Scientific, Inc.), rinsed in PBS and incubated in undiluted Fc
Receptor Blocker solution (Innovex Biosciences, Inc., Richmond, CA,
USA) for 30 min. After rinsing in PBS, tissue sections were
incubated overnight at 4°C with a primary antibody against TRPC3
(1:300; no. ab51560; Abcam). Sections were then washed in PBS three
times for 10 min and then incubated with the suitable
fluorochrome-labeled secondary antibodies (goat anti-rabbit Alexa
Fluor 488; cat. no. ab150077; Invitrogen/Molecular Probes; Thermo
Fisher Scientific, Inc.). Chromatin was counterstained with Hoechst
(10 mg/ml; Sigma-Aldrich; Merck KGaA) diluted in H2O
(1:10,000). Specimens were examined by confocal microscopy with an
Olympus FV-1000 laser scanning confocal microscope (HeNe/Ar) with a
60× water immersion objective, and images were processed with the
FluoView software (Olympus Corporation). Selected representative
fields are presented.
Statistical analysis
Data are expressed as the means ± standard error,
with n defined in the figure legend or text as the number of
animals used to obtain the data; however, as detailed in the figure
legends, tissues were pooled for western blot analysis. Statistical
significance was determined by one-way analysis of variance
followed by Dunnett's post hoc test for multiple comparisons.
Comparison of two variables was performed with unpaired Student's
t-tests where appropriate, with P<0.05 considered to indicate a
statistically significant difference (GraphPad Prism; GraphPad
Software, Inc., La Jolla, CA, USA).
Results
Expression levels of TRPC3,
Cav1.2, Cav3.1 and Cav3.2 increase
during pregnancy
TRPC3 can be activated by stretch, swelling, heat or
pressure, which are physiological stimuli that are likely to be
involved in uterine contractility. Thus, to determine whether TRPC3
channels modulate myometrial tone during pregnancy, western
blotting was used to measure TRPC3 gene expression in primary mSMCs
obtained from human samples (preterm delivery, full-term delivery
without labor onset, full-term delivery with labor onset, and
non-pregnant control groups; Fig.
1). Western blotting indicated that TRPC3 protein expression
was markedly higher in mSMCs from the full-term without labor onset
group and preterm group compared with samples from the non-pregnant
group (Fig. 1B; P<0.05);
however TRPC3 expression was not significantly different in the
full-term with labor onset group compared with the non-pregnant
control group (P>0.05). During pregnancy, compared with the
non-pregnant controls, Cav1.2, Cav3.1 and
Cav3.2 protein expression levels were markedly increased
(P<0.05) in mSMCs from the preterm delivery group and the
full-term with labor onset group (Fig.
1C-E), however were non-significantly increased in the
full-term without labor onset group. The level of TRPC3 expression
was highest in the preterm group, whereas the levels of
Cav1.2, Cav3.1 and Cav3.2 were
highest in the full-term with labor onset group, and western
blotting results were consistent with each other (Fig. 1).
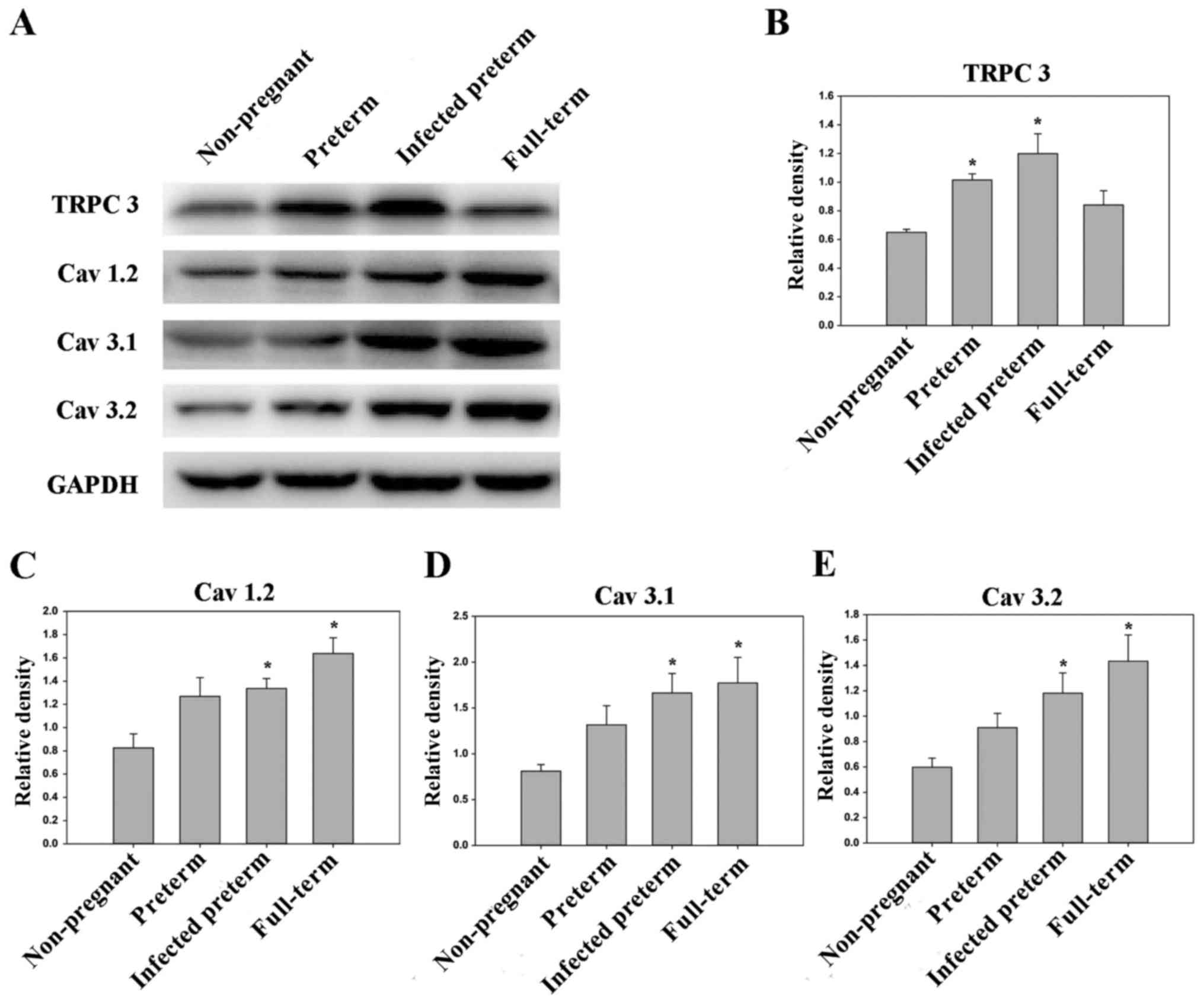
Proteins were subsequently extracted from mouse
uteri and analyzed by western blotting. The levels of TRPC3,
mitogen-activated protein kinase (MAPK), phosphorylated (P-)
extracellular signal-related kinase (ERK), Cav3.2,
Cav3.1, Cav1.2 were all significantly higher
in the LPS-treated preterm group and full-term group compared with
the unfertilized group (all P<0.05). Additionally, the levels of
MAPK and P-ERK were significantly higher in the preterm group than
in the unfertilized group (P<0.05). Furthermore, the levels of
TRPC3, MAPK, P-ERK, Cav3.2, Cav3.1,
Cav1.2 were all higher in the LPS-treated preterm group
than in the preterm group, to varying extents (Fig. 2).
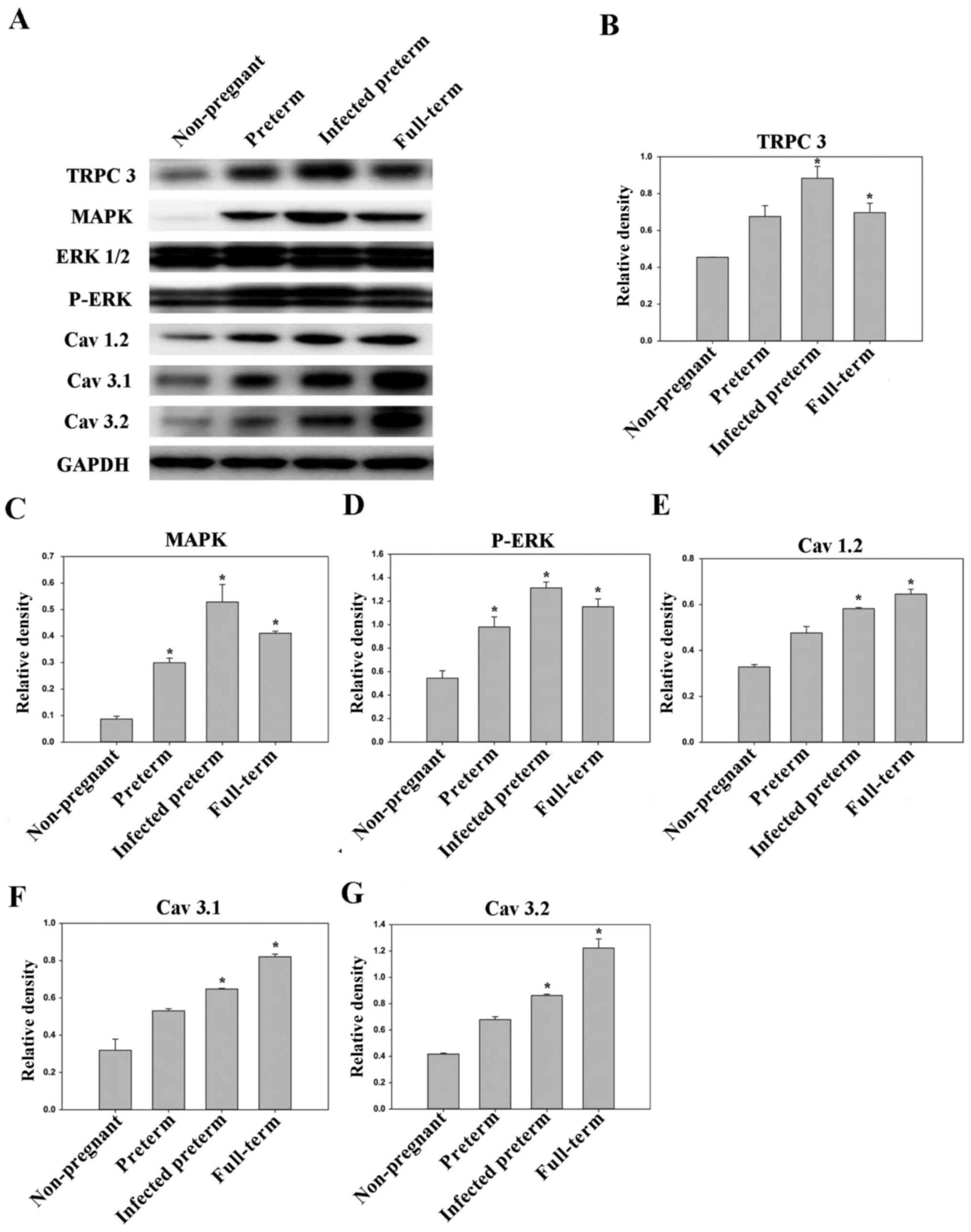 | Figure 2.Expression levels of TRPC3, MAPK,
P-ERK, Cav1.2, Cav.3.1 and Cav3.2
in mouse myometrial smooth muscle cells derived from the
non-pregnant, preterm, infected preterm and full-term groups
(n=10/group). (A) Western blot analysis of protein expression
levels in the different groups. GAPDH was used as the loading
control. Quantified protein expression levels of (B) TRPC3, (C)
MAPK, (D) P-ERK, (E) Cav1.2, (F) Cav.3.1 and
(G) Cav3.2. Data are presented as the mean ± standard
error. *P<0.05 vs. non-pregnant group. TRPC3, canonical
transient receptor potential 3; MAPK, mitogen-activated protein
kinase; ERK, extracellular signal-related kinase; P-,
phosphorylated. |
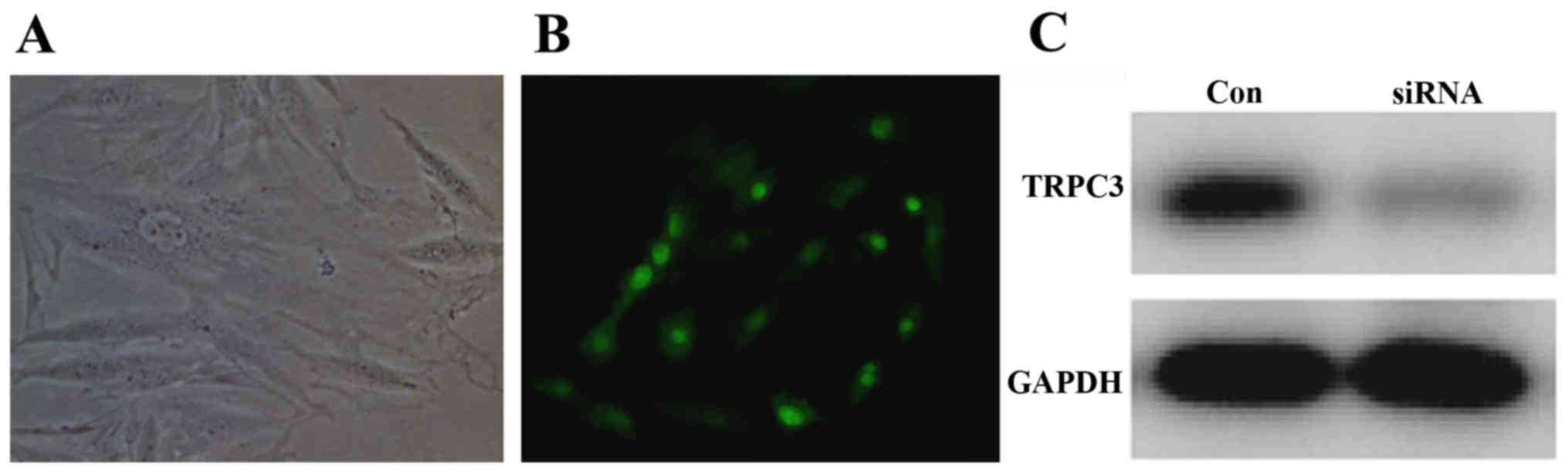
Mouse uterine SMCs were transfected with the TRPC3
plasmid. After 3 days, green fluorescence was observable under the
fluorescence microscope, and cell growth was normal (Fig. 3). Additionally, mouse uterine SMCs
were transfected with TRPC3-1 and TRPC3-2 siRNA for 3 days, and
western blot analysis was subsequently used to detect TRPC3
expression. It was observed that TRPC3 expression was decreased by
transfection with either of the TRPC3 siRNA compared with the
control siRNA (Fig. 3).
Effect of siRNA-mediated TRPC3
downregulation
To assess the role of TRPC3 in mSMCs, TRPC3 was
downregulated by siRNA silencing. Transfection of the mSMCs with
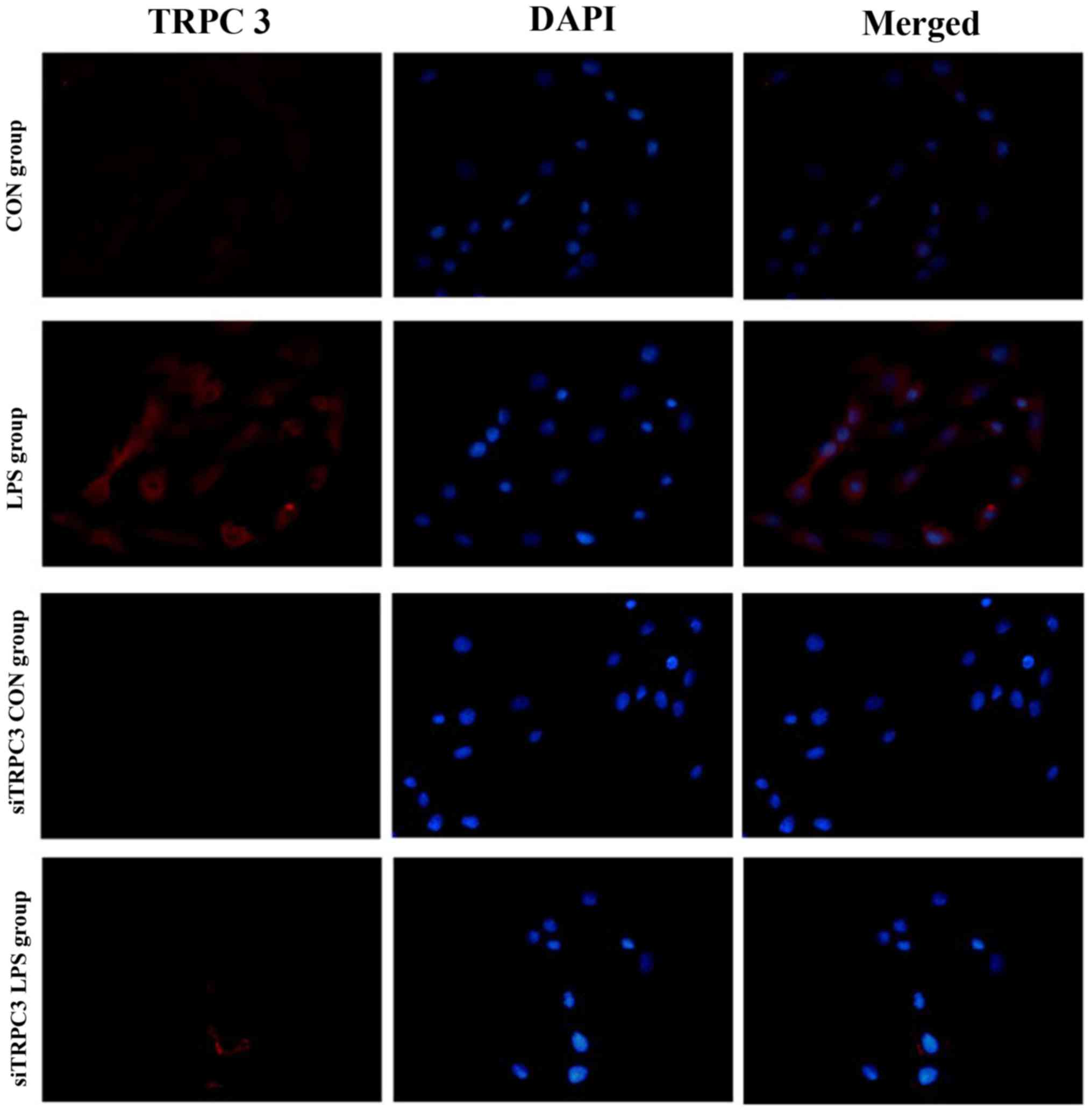
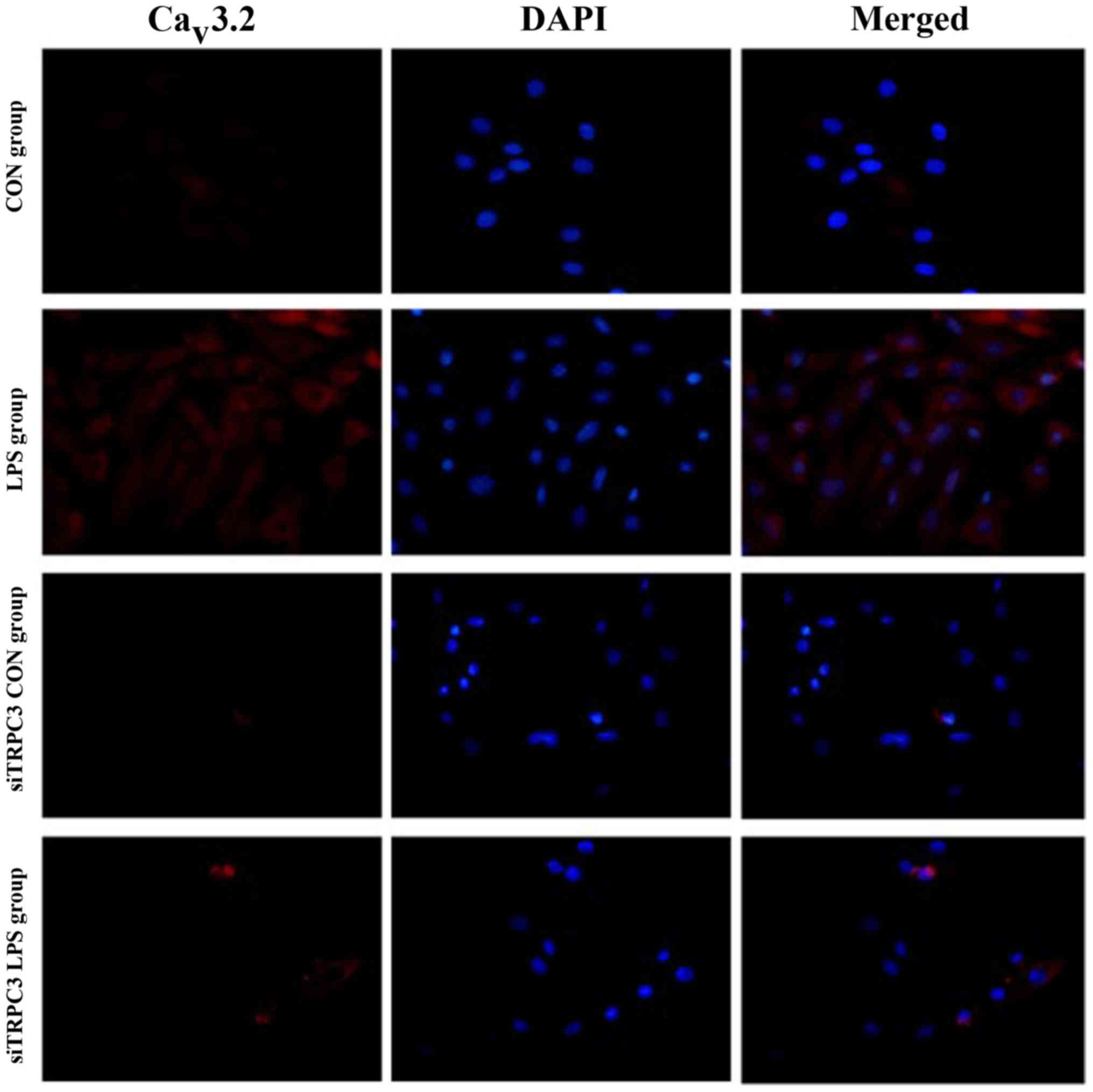
anti-TRPC3 siRNA effectively depleted TRPC3 protein (Fig. 3). Immunofluorescence analysis
indicated that knockdown of TRPC3 expression reduced the expression
levels of PLCγ, PKC, CPI-17, P-CPI-17, Cav1.2,
Cav3.1 and Cav3.2 in mSMCs (Figs. 4–11). In mSMCs obtained from the
LPS-treated group, higher expression levels of TRPC3, PLCγ, PKC,
CPI-17, P-CPI-17, Cav1.2, Cav3.1 and
Cav3.2 were detected in the control siRNA-transfected
group compared with the TRPC3-siRNA-transfected group (Figs. 4–11). Additionally, quantified western
blot analysis confirmed that TRPC3 knockdown reduced the expression
levels of TRPC3, PLCγ, PKC, CPI-17, P-CPI-17, Cav1.2,
Cav3.1 and Cav3.2 compared with the control
siRNA-transfected group in non-LPS-treated cells (Fig. 12).
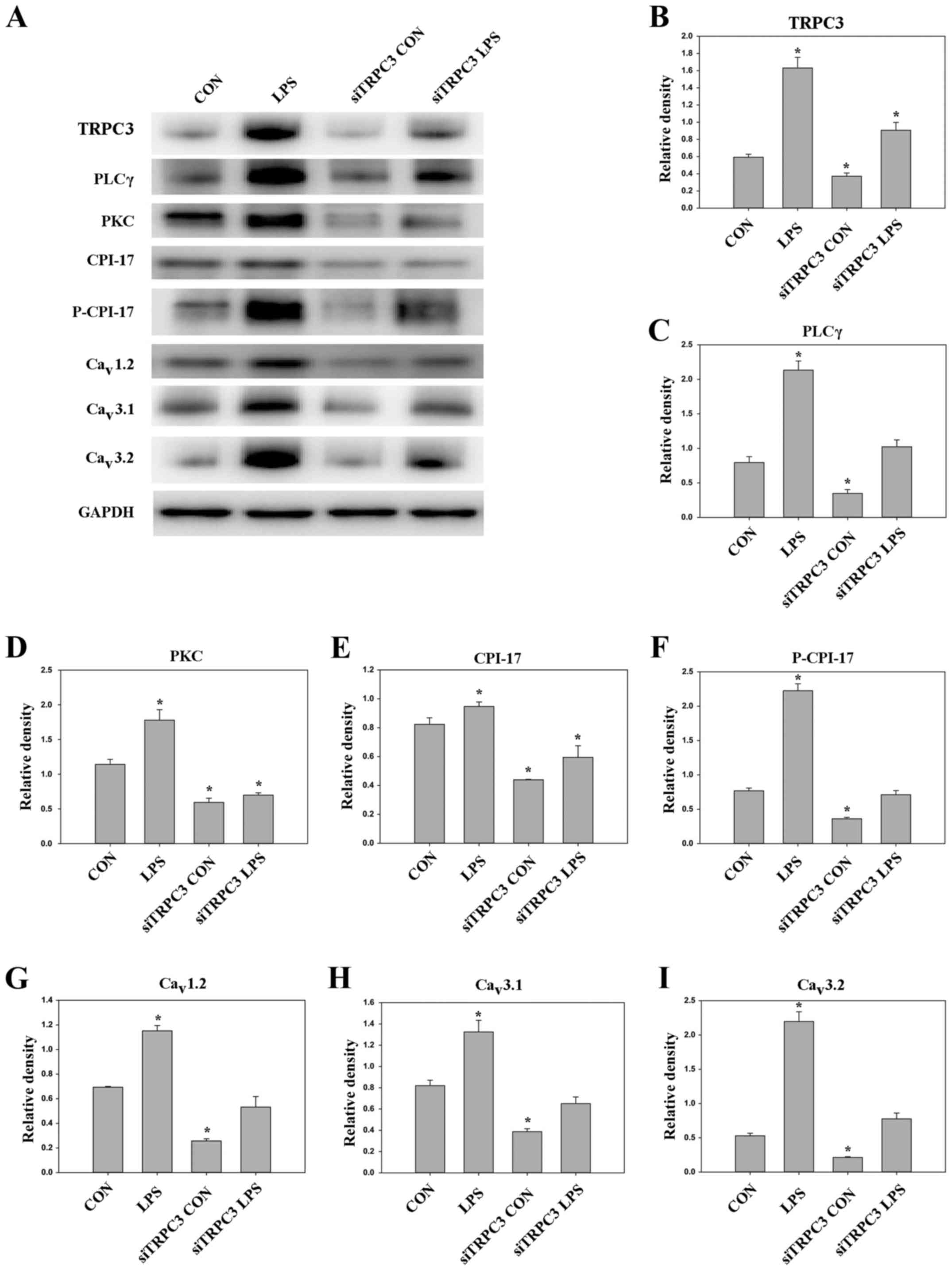 | Figure 12.(A) Western blot analysis of proteins
in mouse myometrial smooth muscle cells with and without
TRPC3-siRNA transfection. GAPDH was used as the loading control.
Quantified protein expression levels of (B) TRPC3, (C) PLCγ, (D)
PKC, (E) CPI-17, (F) P-CPI-17, (G) Cav1.2, (H)
Cav3.1 and (I) Cav3.2. Data are presented as
the mean ± standard error. *P<0.05 vs. CON group. CON,
untransfected non-infected with LPS; LPS, infected with LPS;
siTRPC3 CON, transfected with siTRPC3 and non-infected with LPS;
siTRPC3 LPS, transfected with siTRPC3 and infected with LPS. TRPC3,
canonical transient receptor potential 3; siRNA, small interfering
RNA; PLC, phospholipase C; PKC, protein kinase C; CPI-17,
C-kinase-activated protein phosphatase-1 inhibitor; LPS,
lipopolysaccharides; CON, control. |
Discussion
The effect of uterine canonical TRPC3 expression
affected the parturition-triggering mechanism and PLCγ/PKC/CPI-17
signaling pathways. In the preterm and full-term without labor
onset patient groups, TRPC3 gene expression in the mSMCs was
overexpressed compared with the non-pregnant group; however, TRPC3
expression was not elevated in the full-term with labor onset
group, exhibiting no difference compared with the non-pregnant
group. During pregnancy, compared with the non-pregnant controls,
Cav1.2, Cav3.1 and Cav3.2 gene
expression levels were markedly increased in mSMCs in the preterm
delivery group and the full-term with labor onset group, however
were non-significantly increased in the full-term without labor
onset group. The level of TRPC3 was highest in the preterm group,
while the levels of Cav1.2, Cav3.1 and
Cav3.2 were highest in the full-term with labor onset
group. LPS-treated preterm and full-term groups, TRPC3, MAPK,
ERK1/2, P-ERK, Cav3.2, Cav3.1 and
Cav1.2 were all expressed at higher levels than in the
unfertilized group. In the LPS-treated preterm group, the levels of
TRPC3, MAPK, ERK1/2, P-ERK, Cav3.2, Cav3.1
and Cav1.2 were increased compared with the preterm
group. Furthermore, following transfection of siTRPC3 into cells,
it was demonstrated that the levels of TRPC3, PLCγ, PKC, CPI-17,
P-CPI-17, Cav1.2, Cav3.1 and
Cav3.2 expression were decreased in the LPS siTRPC3
group compared with the LPS-treated untransfected control
group.
CPI-17 is an important signaling molecule involved
in the regulation of calcium sensitivity in smooth muscle
contraction. It is a PKC-activated phosphatase inhibitor with a
relative molecular weight of 17 kDa, and is widely expressed in
vascular and visceral smooth muscle cells (11). Studies have demonstrated that PKC
phosphorylates CPI-17 at Thr-38, then phosphorylated CPI-17 binds
with the 38 kDa MLCP catalytic subunit (the main unit responsible
for enzymatic activity), which causes a 1,000-fold inhibition of
MLCP, thus increasing calcium sensitivity and smooth muscle
contraction (12–14). Additionally, nitric oxide (NO)
binds with soluble guanylate cyclase in smooth muscle cells, which
increases cyclic guanosine monophosphate (cGMP) levels; this
activates a cGMP-dependent kinase, which then dephosphorylates
CPI-17 and inhibits the calcium sensitization mediated by
phosphorylated CPI-17; thus, this pathway reduces the
Ca2+ sensitivity of smooth muscle cells (15).
A previous study identified that there were no
significant differences in CPI-17 mRNA and protein expression
levels between the unfertilized group, full-term delivery without
labor onset group and full-term delivery with labor onset group;
however, phosphorylated CPI-17 protein in uterine smooth muscle
tissue was lower in the unfertilized group compared with the
full-term delivery without labor onset group and full-term delivery
with labor onset group (16,17).
Furthermore phosphorylated CPI-17 protein levels were significantly
higher in the full-term delivery with labor onset group than in the
full-term delivery without labor onset group. These differences may
be explained by the following: During pregnancy, endothelin-1 and
TNF-α (16,17) increase the phosphorylation of
CPI-17, compared with the unfertilized group, whereas the high NO
in uterine smooth muscle tissue inhibits the effect of
phosphorylated CPI-17; during late pregnancy, prostaglandin and
oxytocin levels in uterine smooth muscle gradually increase, and
PKCβ expression is upregulated, which induces CPI-17
phosphorylation at Thr-38 and increases the expression level and
activity of phosphorylated CPI-17; prior to parturition, the NO
level sharply decreases (13), and
the inhibitory effect on phosphorylated CPI-17 is reduced;
therefore, phosphorylated CPI-17 levels increase rapidly, which
inhibits the activity of MLCP, resulting in increased
phosphorylation of MLC20. Phosphorylated MLC20 causes myosin head
swinging and binding with actin, which induces uterine smooth
muscle contraction and triggers parturition.
Although PKC and Rho kinase can act on CPI-17,
CPI-17 activation-induced contraction is only inhibited by
GF109203X, a PKC inhibitor; other inhibitors of
contraction-regulating kinases, such as the Rho kinase inhibitor
Y-27632, exert few inhibitory effects in rat aortic smooth muscle
cells and pulmonary arterial endothelial cells (18–20).
This evidence indicates that PKC is the main regulator of CPI-17
and uterine smooth muscle contraction.
PKC, initially discovered by Nishizuka in 1977, is a
family of phospholipid-dependent Ca2+−activated protein
serine/threonine kinases, and is an important component in cell
signal transduction pathways (21). PKC proteins include two domains:
The catalytic domain located in the C-terminal; and the regulatory
domain located in the N-terminal. In the regulatory domain, there
is an amino acid sequence with similarity to PKC substrates, which
acts as a false substrate sequence. PKC is commonly present in the
cytoplasm in an inactive form, where the substrate cannot enter the
catalytic domain due to the interaction with the false substrate
sequence (22). When extracellular
signals cause PLC activation, IP3 is produced by
hydrolysis of PIP2. IP3 subsequently promotes
Ca2+ release from the ER, which increases the
cytoplasmic Ca2+ concentration and activates PKC. DAG
promotes PKC to translocate from the cytoplasm to the cell
membrane. Under physiological conditions, PKC can only be activated
by binding with the cell membrane (23).
The administration of a nonspecific PKC inhibitor to
full-term pregnant women reduced smooth muscle contraction by
50–87%, indicating that PKC has some effect on smooth muscle
contraction (23). There are 12
subtypes of PKC (23), with varied
distribution among different tissues. PKCα, β, γ, δ, z and ι are
expressed in the uterine smooth muscle (24). However, it remains unclear which
PKC subtype or subtypes contribute to the regulation of uterine
smooth muscle contraction. Sakai et al (25,26)
demonstrated that a PKCβ-specific antagonist inhibits calcium
sensitization mediated by phorbol 12,13-dibutyrate, and causes
uterine smooth muscle relaxation. These studies suggest that PKCβ
subtypes may serve an important role in regulating uterine smooth
muscle contraction.
PKCβ mRNA and protein were expressed in the uterine
smooth muscle of the full term, preterm group, and full term entry
group as detected by RT-qPCR and western blot analyses. PKCβ mRNA
and protein expression levels in the full-term delivery with labor
onset group were significantly increased compared with the
non-pregnant group and full-term delivery without labor onset
group, and the expression level in full-term delivery with labor
onset group was obviously higher than that of without labor onset
group (5,17,27).
This difference may be explained by the observation that
prostaglandins, oxytocin receptors and endothelin can stimulate the
expression of PKCβ (5,17,27).
As the pregnancy continues, the expression of prostaglandins and
oxytocin receptor is increased and reaches a peak prior to the
parturition. These substances activate specific PLC on uterine
smooth muscle cell membrane, which then hydrolyzes PIP2
to IP3 and DAG. IP3 promotes the release of
ER Ca2+ and increases the cytoplasmic Ca2+.
Ca2+ binding to the regulatory domain of PKCβ area can
induce conformational changes, and rapidly increase the catalytic
activity of the protein. DAG stimulates translocation of PKCβ from
the cytoplasm to the cell membrane, which then triggers a
downstream signaling cascade that increases MLC20 phosphorylation
in the uterine smooth muscle cells to enhance uterine smooth muscle
contraction and induce labor.
In summary, the results of the current study suggest
that PKCβ/CPI-17 signaling may serve a role in triggering labor,
and it is suggested that inhibiting this pathway may suppress
birth; however, the pathway regulatory mechanism and its
interaction with other signaling pathways remains unclear. Further
investigation of PKCβ/CPI-17 signaling will aid in the
clarification of the mechanism and may potentially provide novel
treatments for the prevention of preterm delivery.
Acknowledgements
The present study supported by the National Natural
Science Youth Foundation of China (grant nos. 81501287 and
81501260); Construction of Liaoning Provincial Center for
Translational Medicine Research and Collaborative Network
Construction Project (grant no. 2014225007); and the National
Health and Family Planning Commission of the Public Welfare
Industry Research Projects (grant no. 201402006).
References
|
1
|
Senturk MB, Cakmak Y, Gündoğdu M, Polat M
and Atac H: Does performing cesarean section after onset of labor
has positive effect on neonatal respiratory disorders? J Matern
Fetal Neonatal Med. 29:2457–2460. 2016.PubMed/NCBI
|
|
2
|
Hanley GE, Munro S, Greyson D, Gross MM,
Hundley V, Spiby H and Janssen PA: Diagnosing onset of labor: A
systematic review of definitions in the research literature. BMC
Pregnancy Childbirth. 16:712016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ohno Y, Terauchi M, Tamakoshi K, Shiozaki
A and Saito S: The risk factors for labor onset hypertension.
Hypertens Res. 39:260–265. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Neal JL and Lowe NK: Physiologic
partograph to improve birth safety and outcomes among low-risk,
nulliparous women with spontaneous labor onset. Med Hypotheses.
78:319–326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kiesewetter B and Lehner R: Maternal
outcome monitoring: Induction of labor versus spontaneous onset of
labor-a retrospective data analysis. Arch Gynecol Obstet.
286:37–41. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ijaiya MA, Adesina KT, Raji HO, Aboyeji
AP, Olatinwo AO, Adeniran AS, Adebara IO and Isiaka-Lawal S:
Duration of labor with spontaneous onset at the University of
Ilorin Teaching Hospital, Ilorin, Nigeria. Ann Afr Med. 10:115–119.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kapur NK, Qiao X, Paruchuri V, Mackey EE,
Daly GH, Ughreja K, Morine KJ, Levine J, Aronovitz MJ, Hill NS, et
al: Reducing endoglin activity limits calcineurin and TRPC-6
expression and improves survival in a mouse model of right
ventricular pressure overload. J Am Heart Assoc. 3:pii: e000965.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Myeong J, Kwak M, Hong C, Jeon JH and So
I: Identification of a membrane-targeting domain of the transient
receptor potential canonical (TRPC)4 channel unrelated to its
formation of a tetrameric structure. J Biol Chem. 289:34990–35002.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dhar M, Wayman GA, Zhu M, Lambert TJ,
Davare MA and Appleyard SM: Leptin-induced spine formation requires
TrpC channels and the CaM kinase cascade in the hippocampus. J
Neurosci. 34:10022–10033. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Wang Y, Yang K, Tian L, Fu X,
Wang Y, Sun Y, Jiang Q, Lu W and Wang J: BMP4 increases the
expression of TRPC and basal [Ca2+]i via the p38MAPK and ERK1/2
pathways independent of BMPRII in PASMCs. PLoS One. 9:e1126952014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ilatovskaya DV, Levchenko V, Lowing A,
Shuyskiy LS, Palygin O and Staruschenko A: Podocyte injury in
diabetic nephropathy: Implications of angiotensin II-dependent
activation of TRPC channels. Sci Rep. 5:176372015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamada H, Yoshida M, Ito K, Dezaki K, Yada
T, Ishikawa SE and Kakei M: Potentiation of glucose-stimulated
insulin secretion by the GPR40-PLC-TRPC pathway in pancreatic
β-cells. Sci Rep. 6:259122016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ziemba BP, Burke JE, Masson G, Williams RL
and Falke JJ: Regulation of PI3K by PKC and MARCKS: Single-molecule
analysis of a reconstituted signaling pathway. Biophys J.
110:1811–1825. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ruiz-Loredo AY, López E and López-Colomé
AM: Thrombin stimulates stress fiber assembly in RPE cells by
PKC/CPI-17-mediated MLCP inactivation. Exp Eye Res. 96:13–23. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hagel C, Dornblut C, Schulz A, Wiehl U,
Friedrich RE, Huckhagel T, Mautner VF and Morrison H: The putative
oncogene CPI-17 is up-regulated in schwannoma. Neuropathol Appl
Neurobiol. 42:664–668. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su W, Xie Z, Liu S, Calderon LE, Guo Z and
Gong MC: Smooth muscle-selective CPI-17 expression increases
vascular smooth muscle contraction and blood pressure. Am J Physiol
Heart Circ Physiol. 305:H104–H113. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang W, Li Z, Zhao W, Chen H, Wu Y, Wang
Y, Shen Z, He J, Chen S, Zhang J and Fu G: Breviscapine attenuatted
contrast medium-induced nephropathy via PKC/Akt/MAPK signalling in
diabetic mice. Am J Transl Res. 8:329–341. 2016.PubMed/NCBI
|
|
18
|
Li W, Lv J, Wu J, Zhou X, Jiang L, Zhu X,
Tu Q, Tang J, Liu Y, He A, et al: Maternal high-salt diet altered
PKC/MLC20 pathway and increased ANG II receptor-mediated
vasoconstriction in adult male rat offspring. Mol Nutr Food Res.
60:1684–1694. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Murthy KS, Zhou H, Grider JR, Brautigan
DL, Eto M and Makhlouf GM: Differential signalling by muscarinic
receptors in smooth muscle: m2-mediated inactivation of myosin
light chain kinase via Gi3, Cdc42/Rac1 and p21-activated kinase 1
pathway, and m3-mediated MLC20 (20 kDa regulatory light chain of
myosin II) phosphorylation via Rho-associated kinase/myosin
phosphatase targeting subunit 1 and protein kinase C/CPI-17
pathway. Biochem J. 374:145–155. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kolosova IA, Ma SF, Adyshev DM, Wang P,
Ohba M, Natarajan V, Garcia JG and Verin AD: Role of CPI-17 in the
regulation of endothelial cytoskeleton. Am J Physiol Lung Cell Mol
Physiol. 287:L970–L980. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Eto M, Ohmori T, Suzuki M, Furuya K and
Morita F: A novel protein phosphatase-1 inhibitory protein
potentiated by protein kinase C. Isolation from porcine aorta media
andaracterization. J Biochem. 18:1104–1107. 1995. View Article : Google Scholar
|
|
22
|
MacDonald JA, Eto M, Borman MA, Brautigan
DL and Haystead TA: Dual Ser and Thr phosphorylation of CPI-17, an
inhibitor of myosin phosphatase, by MYPT associated kinase. FEBS
Lett. 493:91–94. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zemlickova E, Johannes FJ, Aitken A and
Dubois T: Association of CPI-17 with protein kinase C and casein
kinase I. Biochem Biophys Res Commun. 316:39–47. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kitazawa T, Semba S, Huh YH, Kitazawa K
and Eto M: Nitric oxide-induced biphasic mechanism of vascular
relaxation via dephosphorylation of CPI-17 and MYPT1. J Physiol.
587:3587–3603. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sakai H, Hirano T, Takeyama H, Chiba Y and
Misawa M: Acetylcholine-induced phosphorylation of CPI-17 in rat
bronchial smooth muscle: The roles of Rho-kinase and protein kinase
C. Can J Physiol Pharmacol. 83:375–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sakai H, Chiba Y and Misawa M:
Augmentation of endothelin-1-induced phosphorylation of CPI-17 and
myosin light chain in bronchial smooth muscle from airway
hyperresponsive rats. Biol Pharm Bull. 29:1897–1899. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sladek SM, Magness RR and Conrad KP:
Nitric oxide and pregnancy. Am J Physiol. 272:R441–R463.
1997.PubMed/NCBI
|