Introduction
Among cancers, lung cancer has the highest incidence
and cancer-associated mortality worldwide (1–3). In
2015, nearly 733,300 new lung cancer cases and 610,200 lung cancer
deaths occurred in China (4).
Despite improvements in early detection and therapeutic strategies,
the prognosis with standard treatments for patients remains poor,
with a 5-year relapse rate of 80% (5). Many investigations have sought to
identify novel drug targets and effective ways to combat lung
cancer, however, significant research is still required (6–8).
Therefore, there is an urgency to develop novel effective agents to
treat lung cancer.
The Akt signalling pathway is involved in cellular
proliferation, survival and apoptosis in various types of cancer
and has been recognized as a potential molecular target for cancer
therapy (9–11). Activated Akt has been shown to
promote tumour progression in various cancers through cell cycle
arrest and the promotion of apoptosis (11). Abundant evidences indicated that
anti-cancer agents regulate biological behaviour via Akt in various
cancer cell lines. The activity of Akt is also regulated by casein
kinase 2 (CK2) inhibitor (12).
Mitogen-activated protein kinase (MAPK) signalling pathways relay
and integrate signals from a wide range of stimuli and control
cellular proliferation, cell cycle and apoptosis (13–17).
Therefore, the MAPK in these signalling pathways are considered
‘stress-activated protein kinases’ in cellular signalling,
inflammation, apoptosis and the pathogenesis of various diseases.
The signal transducer and activator of transcription-3 (STAT3)
signalling pathway has been shown to regulate the transcriptional
activation of gene products that participate in cell proliferation
and anti-apoptosis, such as cell cycle (cyclins and
cyclin-dependent kinases, Cdks) and apoptotic regulators (Bcl-2 and
Bad) (18). These findings
indicate that the targeting of Akt, MAPK and STAT3 signalling
pathways may be an important therapeutic target in novel cancer
therapy.
Reactive oxygen species (ROS) are one of the major
causes of tumours and have significant roles in the processes of
tumour progression, metastasis and apoptosis (19–21).
Intracellular levels of ROS play key roles in survival and various
physiological functions and also proven to be able to promote cell
proliferation and apoptosis through threshold levels (22). Many studies have shown that ROS
accumulation directly increases mitochondrial dysfunction and the
subsequent initiation of apoptosis (22–24).
Furthermore, ROS or oxidative stress-responses may alter various
cancer developments by activating canonical ROS-responsive
signalling pathways such as MAPK and STAT3 pathways (24). Thus, increasing intracellular ROS
generation and modulating the MAPK and STAT3 signalling pathways
may be an effective approach in the treatment and prevention of
cancers.
Casein kinase 2 (CK2) has been shown to be involved
in malignant proliferation, apoptosis resistance and signal
transduction in different cancer cells (25–27).
Many studies have confirmed the underlying targeting of protein
kinase CK2 inhibitors for various cancer therapies (25–28).
Quinalizarin is a protein kinase CK2 and ATP-competitive inhibitor
that has previously been shown to be a potent, highly selective and
cell-permeable inhibitor (28).
Quinalizarin has been reported to promote apoptosis in human
nasopharyngeal carcinoma CNE-1 and CNE-2 cells (29). However, the underlying molecular
mechanisms of quinalizarin-induced apoptosis in lung cancer cells
remain unknown.
In the present study, we examined the effect of
quinalizarin on anti-proliferation, inducing cell cycle arrest,
apoptosis and ROS generation in lung cancer cell lines. The roles
of the Akt, MAPK, STAT3 and p53 signalling pathways in
quinalizarin-induced cell apoptosis and intracellular ROS
generation were examined.
Materials and methods
Cell culture
Lung cancer A549, NCI-H460, NCI-H23 cell lines and
normal liver QSG-7701 cell lines were obtained from American Type
Culture Collection (Manassas, VA, USA). Cells were cultured in DMEM
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
supplemented with 10% foetal bovine serum (FBS), penicillin (100
U/ml) and streptomycin (100 µg/ml; Gibco) in a 5%
CO2 humidified atmosphere at 37°C. Cells were monitored
daily, and medium was replaced every 2 days at 70% cell
density.
Cell cytotoxicity assays
A549, NCI-H460, NCI-H23 and QSG-7701 cells were
harvested at logarithmic phase and dispensed into 96-well plates at
a density of 5,000 cells per well. After 24 h of incubation, the
A549, NCI-H460, NCI-H23 and QSG-7701 cells were treated with
various concentrations (1, 3, 10, 30 and 100 µmol/l) of
5-fluorouracil (5-FU) and quinalizarin (Sigma-Aldrich, St. Louis,
MO, USA) for 24 h. Controls included untreated and dimethyl
sulfoxide (DMSO; Sigma-Aldrich) treated cells. MTT solution (20
µl; 5 mg/ml) was added to the wells, and 2 h later, the
intracellular formazan crystals that had formed were solubilized
with 100 µl DMSO, and the cells were incubated for an
additional 15 min at 37°C. The absorbance values of the solution
were measured at 540 nm with a microplate luminometer. Percentage
viability was calculated as (OD of drug-treated sample/OD of
control sample) ×100.
Cell cycle analysis
A549 cells in logarithmic phase were seeded onto
6-well culture plates (1×105 cells/well) and cultured 24
h. Then, the cells were treated with 12.1 µmol/l
quinalizarin for 0, 3, 6, 12 and 24 h. Cells were trypsinized and
fixed in 70% ethanol at 4°C for 12 h and washed with PBS 2–3 times.
Cells were resuspended in 195 µl binding buffer and
incubated with RNase and propidium iodide (PI; Beyotime
Biotechnology, Shanghai, China) for 30 min without bright light at
37°C. The cellular DNA content of the treated cells was analysed by
flow cytometry (BD Biosciences, San Jose, CA, USA) with a 488 nm
argon laser.
Cell apoptosis analysis
Early and late apoptosis was analysed by Annexin
V-FITC/PI double staining and flow cytometry. Cells in the
logarithmic growth phase were plated onto 6-well plates at a
density of 1×105 cells/well and incubated overnight.
After being treated with quinalizarin (12.1 µmol/l) for 0,
3, 6, 12 and 24 h, cells were centrifuged at 5,000 × g for 7 min at
4°C and washed with PBS 2–3 times. Each sample was stained with 5
µl Annexin V-FITC and 3 µl PI at room temperature for
20 min in the absence of bright light, and cells were then detected
by flow cytometry. The results are reported as the mean values from
three independent experiments.
Western blotting analysis
Cells were washed in PBS and lysed in cell
extraction buffer (1 M HEPES, pH 7.0; 5 M NaCl; 0.5% Triton X-100;
10% glycerol; 20 mM β-mercaptoethanol; 20 mg/ml AEBSF; 0.5 mg/ml
pepstatin; 0.5 mg/ml leupeptin; and 2 mg/ml aprotinin). Cell
lysates were centrifuged for 30 min at 12,000 × g and 4°C. Then,
the supernatants were dissolved with 5× sample loading buffer and
boiled for 5 min. The resulting supernatants (30 µg) were
separated on 10% SDS-PAGE and transferred to NC membranes. The
membranes were blocked with 5% skim milk for 2 h at room
temperature and incubated for 12 h (overnight) at 4°C with the
following primary antibodies (all obtained from Santa Cruz
Biotechnology, Inc., Dallas, TX, USA): Mouse monoclonal antibodies
against β-actin (1:2,500; cat. no. sc-47778), α-tubulin (1:2,500;
cat. no. sc-8035), Bad (1:1,500; cat. no. sc-8044), Bcl-2 (1:1,500;
cat. no. sc-7382), PARP-1 (1:1,500; cat. no. sc-8007), cleaved
caspase-3 (cle-caspase-3; 1:1,500; cat. no. sc-373730), p-ERK
(1:1,500; cat. no. sc-7383), p-JNK (1:1,500; cat. no. sc-6254), JNK
(1:1,500; cat. no. sc-7345), p-p38 (1:1,500; cat. no. sc-7973),
p-STAT3 (1:1,500; cat. no. sc-8059), STAT3 (1:1,500; cat. no.
sc-8019); and rabbit polyclonal antibodies against CDK2 (1:2,500;
cat. no. sc-163), CDK4 (1:2,500; cat. no. sc-260), CDK6 (1:2,500;
cat. no. sc-177), cyclin D1 (1:2,500; cat. no. sc-753), cyclin E
(1:2,500; cat. no. sc-481), p21 (1:1,500; cat. no. sc-397), p27
(1:2,500; cat. no. sc-528), pro caspase-3 (1:2,500; cat. no.
sc-7148), ERK2 (1:1,500; cat. no. sc-154), p38α/β (1:1,500; cat.
no. sc-7194), p-p53 (1:2,500; cat. no. sc-101762), p53 (1:1,500;
cat. no. sc-6243), Akt1/2/3 (1:1,500; cat. no. sc-8312), and
p-Akt1/2/3 (1:2,500; cat. no. sc-7985-R). Peroxidase-Conjugated
Affini Pure Goat Anti-Mouse IgG (1:5,000; cat. no. ZB-2305) and
Goat Anti-Rabbit IgG (1:5,000; cat. no. ZB-2301) were used as the
secondary antibodies. Membranes were incubated with
chemiluminescence reagent (Millipore, Billerica, MA, USA), detected
by a chemiluminescence instrument and analysed by using ImageJ
1.42q software.
Detection of ROS
The generation of intracellular ROS in response to
quinalizarin treatment in A549 cells was measured by using
2′,7′-dichlorofluorescein diacetate (DCFH-DA; MERCK, Shanghai,
China) permeates. Cells were plated onto 6-well culture plates
(1×105 cells/well) and incubated for 24 h. Then, the
cells were treated with quinalizarin (12.1 µmol/l) for 0, 3,
6, 12 and 24 h, harvested at 5,000 × g for 5 min and incubated with
DCFH-DA for 30 min in the dark. The results were analysed by flow
cytometry (Beckman Coulter, Inc., Brea, CA, USA).
Statistical analysis
Data were presented as the mean ± standard deviation
and all of the experiments were replicated 3 times. Statistical
analyses were performed using Excel. One-way analysis of variance
or Student's t-test was used to evaluate the statistical
significance between controls with treated groups. P<0.05
indicated statistically significant differences.
Results
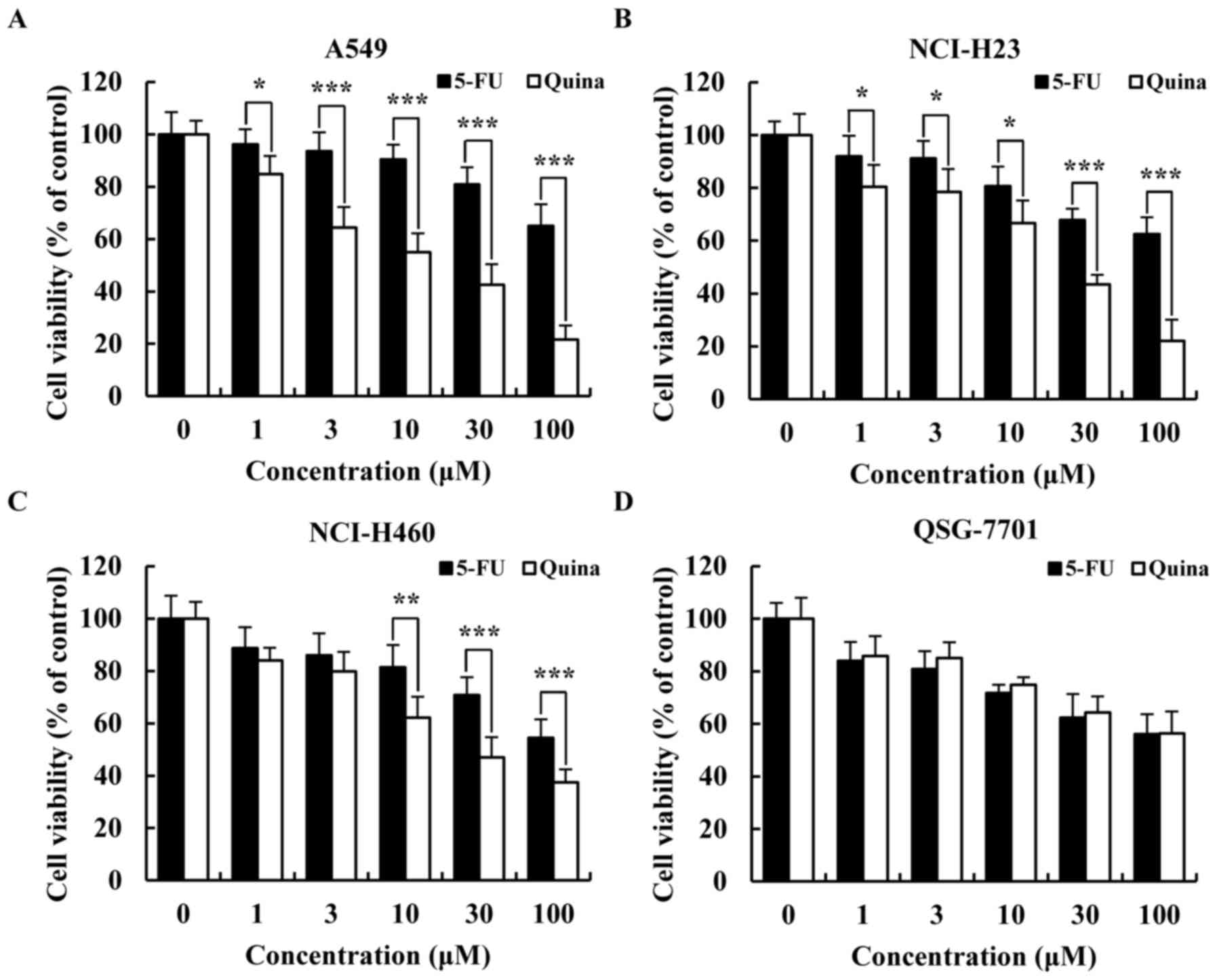
Quinalizarin has cytotoxic effects on
lung cancer cells
To determine whether quinalizarin had cytotoxic
effects in lung cancer cells (A549, NCI-H23 and NCI-H460), cell
viabilities were detected with MTT assays. As shown in Fig. 1A-C, quinalizarin inhibited the cell
viability of the three lung cancer cell types (A549, NCI-H23 and
NCI-H460) in a concentration-dependent manner. Furthermore, there
were significant differences after quinalizarin treatment compared
with 5-FU treatment. The inhibition rate of quinalizarin on A549
cells (IC50, 12.1 µmol/l) was higher than those
on NCI-H23 (IC50, 20.24 µmol/l) and NCI-H460
(IC50, 27.94 µmol/l) cells at 24 h. Furthermore,
there were no significant cytotoxic effects of quinalizarin
compared with 5-FU treatments in normal liver QSG-7701 cells
(Fig. 1D). These results indicated
that quinalizarin has significant dose-dependent cell toxicity
effects in lung cancer cells. Because A549 cells were more
sensitive to quinalizarin, we used A549 cells as a representative
for subsequent experiments.
Quinalizarin induces cell cycle arrest
and regulates expression of cell cycle-related proteins in A549
cells
To investigate whether quinalizarin induced growth
inhibition and cell cycle arrest, cells exposed to quinalizarin
were analysed with flow cytometry and western blotting. As shown in
Fig. 2A and B, the percentage of
cells in G0/G1 phase was significantly
increased compared with the control groups, and cell numbers in
G2/M phase decreased. These results suggested that
quinalizarin induced cell cycle arrest at
G0/G1 phase in a time-dependent manner. To
further investigate whether the CDK/cyclin signalling pathway was
involved in quinalizarin-induced cell cycle arrest,
G0/G1 phase regulatory proteins were
examined. A549 cellular protein expression levels of CDK2/4/6 and
cyclin D1/E were repressed, and p21 and p27 were increased in a
time-dependent manner (Fig. 2C and
D). These results showed that quinalizarin induces A549 cell
cycle arrest at G0/G1 phase in a
time-dependent manner, with decreased expression levels of CDK2/4/6
and cyclin D1/E.
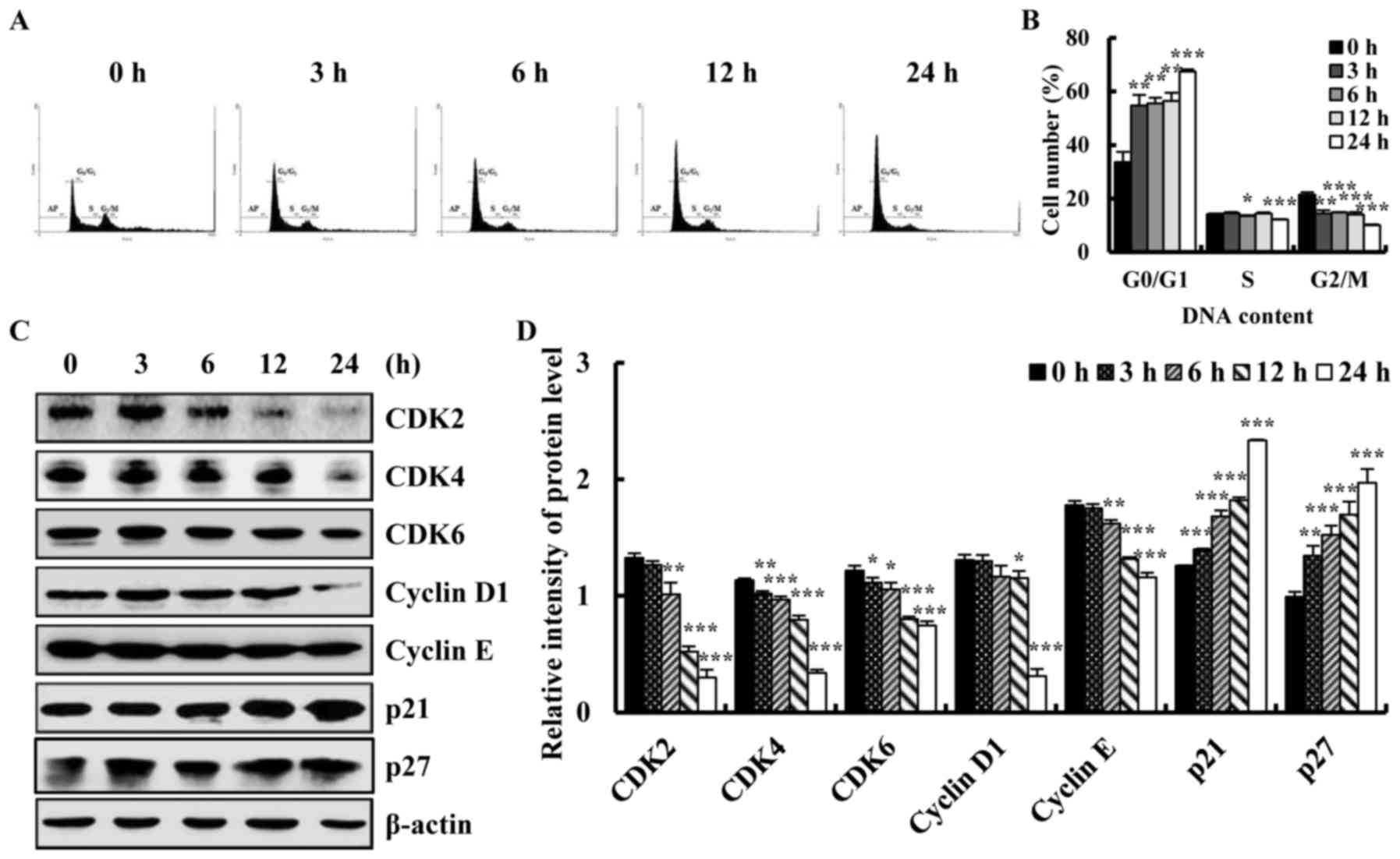 | Figure 2.Quinalizarin induced A549 cell cycle
arrest at G0/G1 phase and affected the cell
cycle regulatory proteins in A549 lung cancer cells. (A) Cells were
treated with 12.1 µmol/l quinalizarin at 0, 3, 6, 12 and 24 h, and
the cell cycle distribution was analysed by flow cytometry, and (B)
quantified. (C) Cells were treated with 12.1 µmol/l quinalizarin
for the indicated times (0 to 24 h), and were then subjected to
western blotting to detect the expression of the cell cycle
associated proteins CDK2/4/6, cyclin D1/E, p21 and p27. (D)
Quantification of western blot analysis. Data are presented as the
mean ± standard deviation. *P<0.05, **P<0.01 and
***P<0.001 vs. control group (0 h). CDK, cyclin dependent
kinase. |
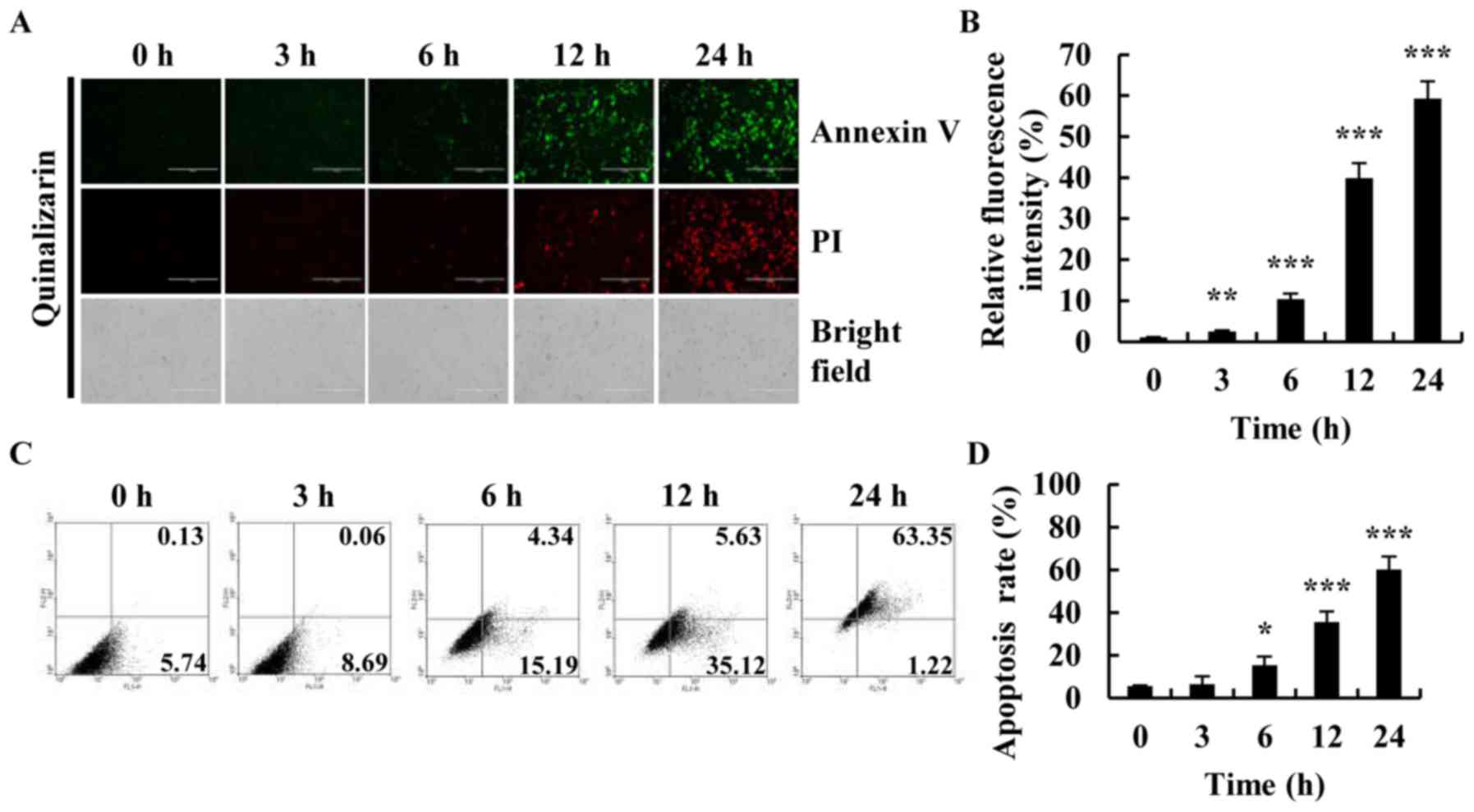
Quinalizarin induces apoptosis in A549
cells
To investigate whether quinalizarin induced
apoptosis in lung cancer cells, A549 cells were treated with
quinalizarin for different time (0, 3, 6, 12 and 24 h), and
apoptosis was observed by Annexin V-FITC/PI double staining.
Fluorescence intensity and morphology changes were observed by
fluorescence microscope. The fluorescence intensity of Annexin
V-FITC and PI of quinalizarin-treatment cells was increased in a
time-dependent manner (Fig. 3A and
B). Especially after 24 h of treatment with quinalizarin,
apoptosis was most apparent as compared with that in the bright
field groups. Cells had decreased in size and were rounded and
floating, thus further demonstrating the ability of quinalizarin to
induce apoptosis in lung cancer cells. The percentage of early and
late apoptosis was quantified by flow cytometry (Fig. 3C and D). The percentages of
apoptotic cells increased from 5.87 to 64.57% for the A549 cells,
thus suggesting that quinalizarin is a potent inducer of apoptosis
in A549 cells.
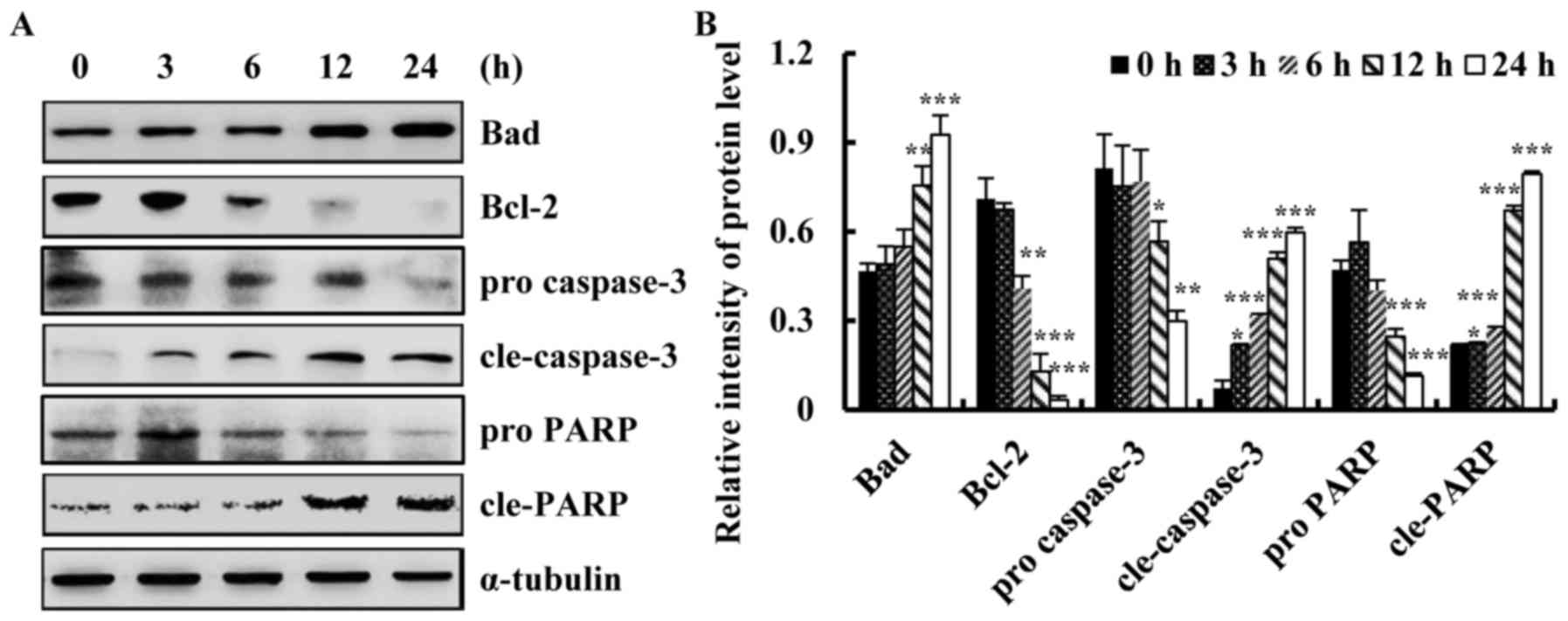
Quinalizarin induces apoptosis in A549
cells via alterations in Bcl-2 family proteins and caspase
activation
To investigate whether quinalizarin induced
apoptosis via the mitochondrial pathway, we detected apoptotic
protein expression levels by western blotting. As shown in Fig. 4A and B, quinalizarin significantly
increased the protein expression level of Bad and decreased that of
Bcl-2. Furthermore, quinalizarin increased caspase-3 and PARP
activity in a time-dependent manner. These results indicated that
the induction of apoptosis was associated with the down-regulation
of Bcl-2 and the up-regulation of Bad, cle-caspase-3 and cleaved
PARP in lung cancer cells. These results showed that quinalizarin
induces apoptosis via the activation of common apoptotic
regulators.
Quinalizarin activates Akt, MAPKs,
STAT3 and p53 signalling pathways
To investigate the mechanism underlying
quinalizarin-induced apoptosis in A549 cells, Akt, MAPK, STAT3 and
p53 signalling pathway-related proteins were analysed by western
blotting. As shown in Fig. 5A-F,
the phosphorylation of Akt, ERK and STAT3 was significantly
decreased and the phosphorylation of JNK, p38 and p53 was
significantly increased after quinalizarin treatment. These results
suggested that quinalizarin-induced activation of the Akt, MAPK,
STAT3 and p53 signalling pathways promote A549 cell apoptosis.
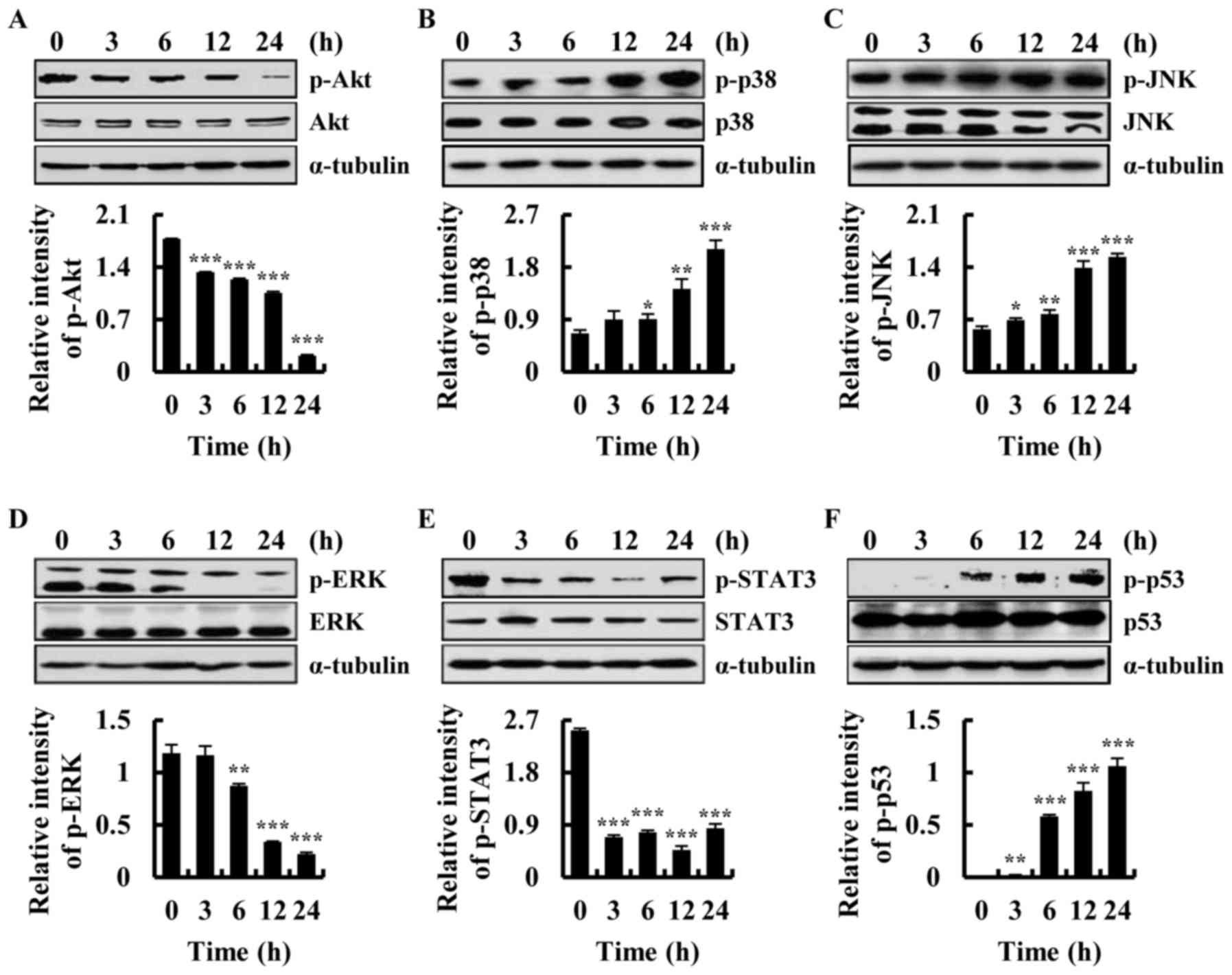 | Figure 5.Quinalizarin induces apoptosis via
Akt, mitogen-activated protein kinases, STAT3 and p53 signalling
pathways. The expression of (A) Akt, (B) p38, (C) JNK, (D) ERK, (E)
STAT3 and (F) p53 proteins was detected by western blot analysis.
*P<0.05, **P<0.01 and ***P<0.001 vs. control group (0 h).
Akt, protein kinase B; JNK, c-Jun N-terminal kinase; ERK,
extracellular signal-regulated kinases; STAT3, signal transducer
and activator of transcription-3; p-, phosphorylated. |
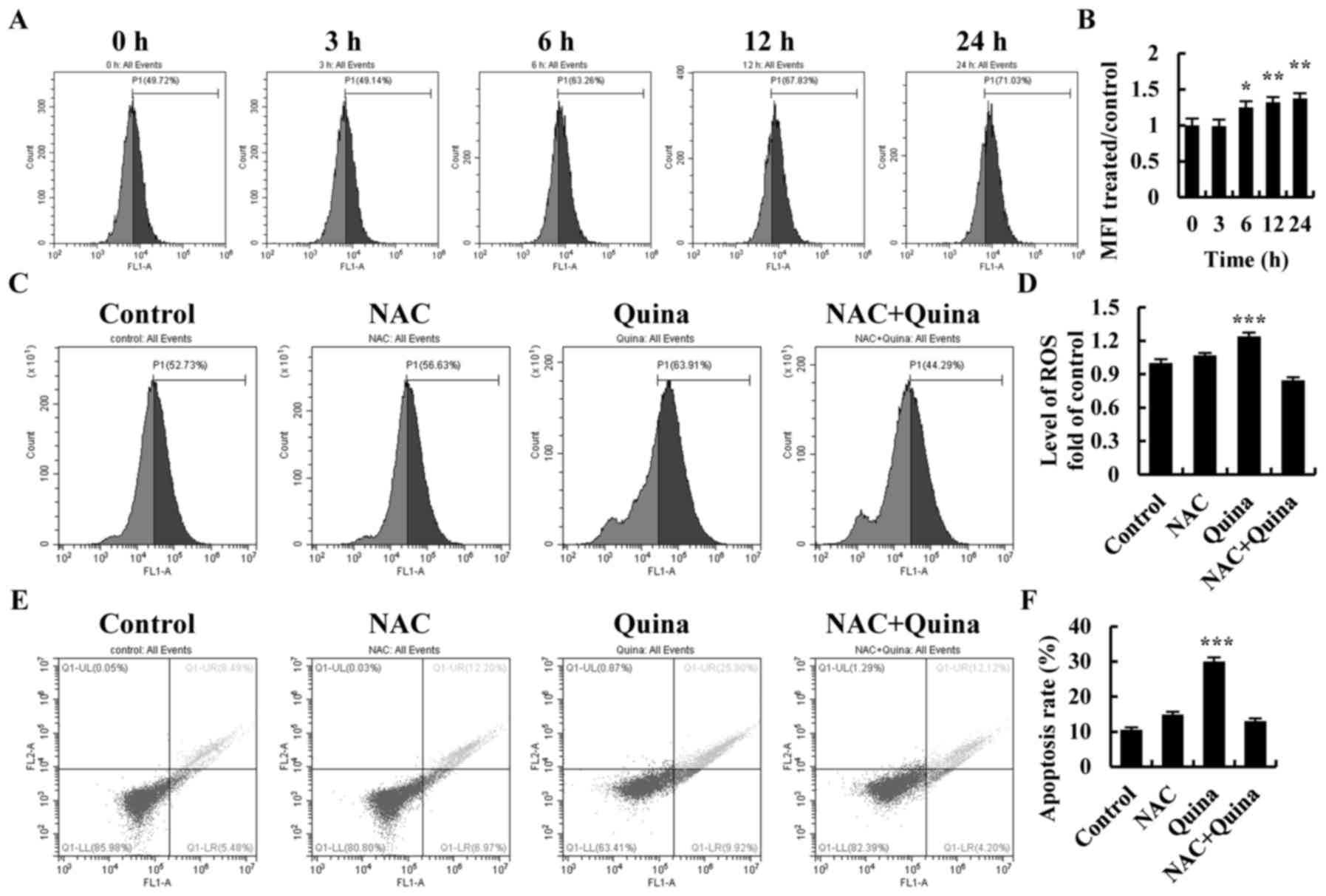
Quinalizarin induces intracellular ROS
generation and ROS scavenger NAC suppression of cell apoptosis
To investigate the relationship between ROS
generation and cell apoptosis, quinalizarin-treated A549 cells were
used to study the mechanism of ROS generation during apoptosis. As
shown in Fig. 6A and B, ROS levels
increased under quinalizarin treatment in a time-dependent manner.
After incubation with quinalizarin and N-acetyl-L-cysteine (NAC),
ROS levels were significantly decreased as compared with the levels
in the quinalizarin group (Fig. 6C and
D). Scavenging of ROS also significantly decreased
quinalizarin-induced cell apoptosis (Fig. 6E and F). Next, we used Western
blotting to confirm the anti-apoptotic mechanism of ROS in
quinalizarin-induced cell apoptosis. After incubation with
quinalizarin and NAC, the phosphorylation of ERK and STAT3
increased and p38, JNK, cle-caspase-3 and cle-PARP were decreased
(Fig. 7A and B). These data
indicated that quinalizarin induces cell apoptosis through the
generation of ROS via the MAPK and STAT3 signalling pathways.
SB203580 (p38 inhibitor), SP600125 (JNK inhibitor) and FR180204
(ERK inhibitor) were applied to confirm the role of MAPK and STAT3
signalling pathways in the quinalizarin-induced apoptosis on lung
cancer cells. A549 cells were pre-treated with 12.1 µmol/l
of SB203580, SP600125 or FR180204 for 30 min followed by treatment
with quinalizarin for 24 h. The decreased protein expression levels
of p-STAT3 induced by quinalizarin were suppressed by adding the
p38MAPK inhibitor and JNK inhibitor, enhanced by adding the ERK
inhibitor (Fig. 8A-F). These
results showed that MAPK was involved in regulating the STAT3
signalling pathway and induced apoptosis in lung cancer A549
cell.
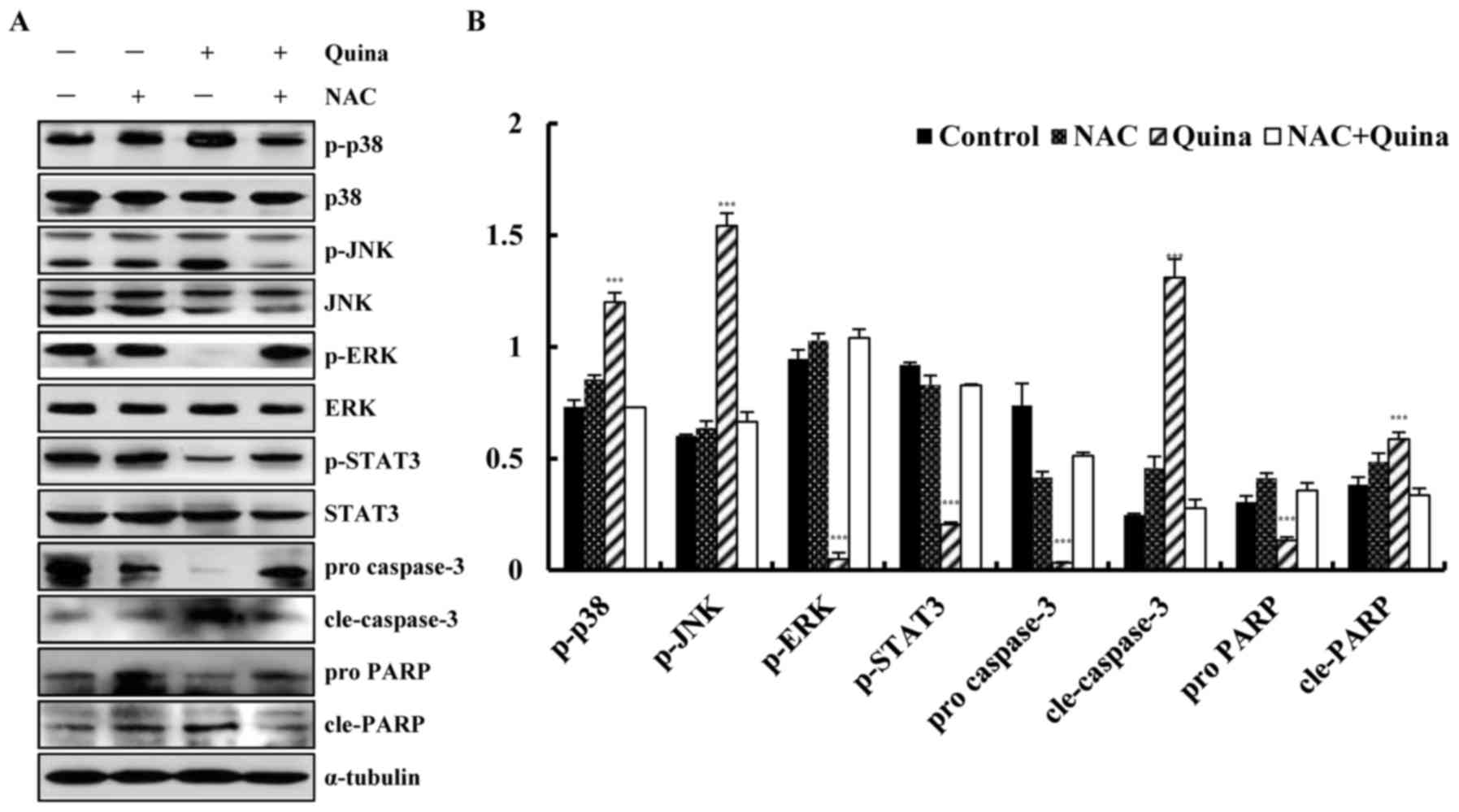 | Figure 7.Quinalizarin induces ROS-mediated
apoptosis via mitogen-activated protein kinase and STAT3 signalling
pathways. (A) The relative protein expression levels of p38, JNK,
ERK, STAT3, cleaved-caspase-3 and cleaved-PARP were detected by
western blotting and (B) quantified. ***P<0.001 vs. NAC + Quina
group. ROS, reactive oxygen species; NAC, N-acetyl-L-cysteine;
Quina, quinalizarin; JNK, c-Jun N-terminal kinase; ERK,
extracellular signal-regulated kinases; STAT3, signal transducer
and activator of transcription-3; p-, phosphorylated; PARP, poly
(adenosine diphosphate-ribose) polymerase; cle, cleaved. |
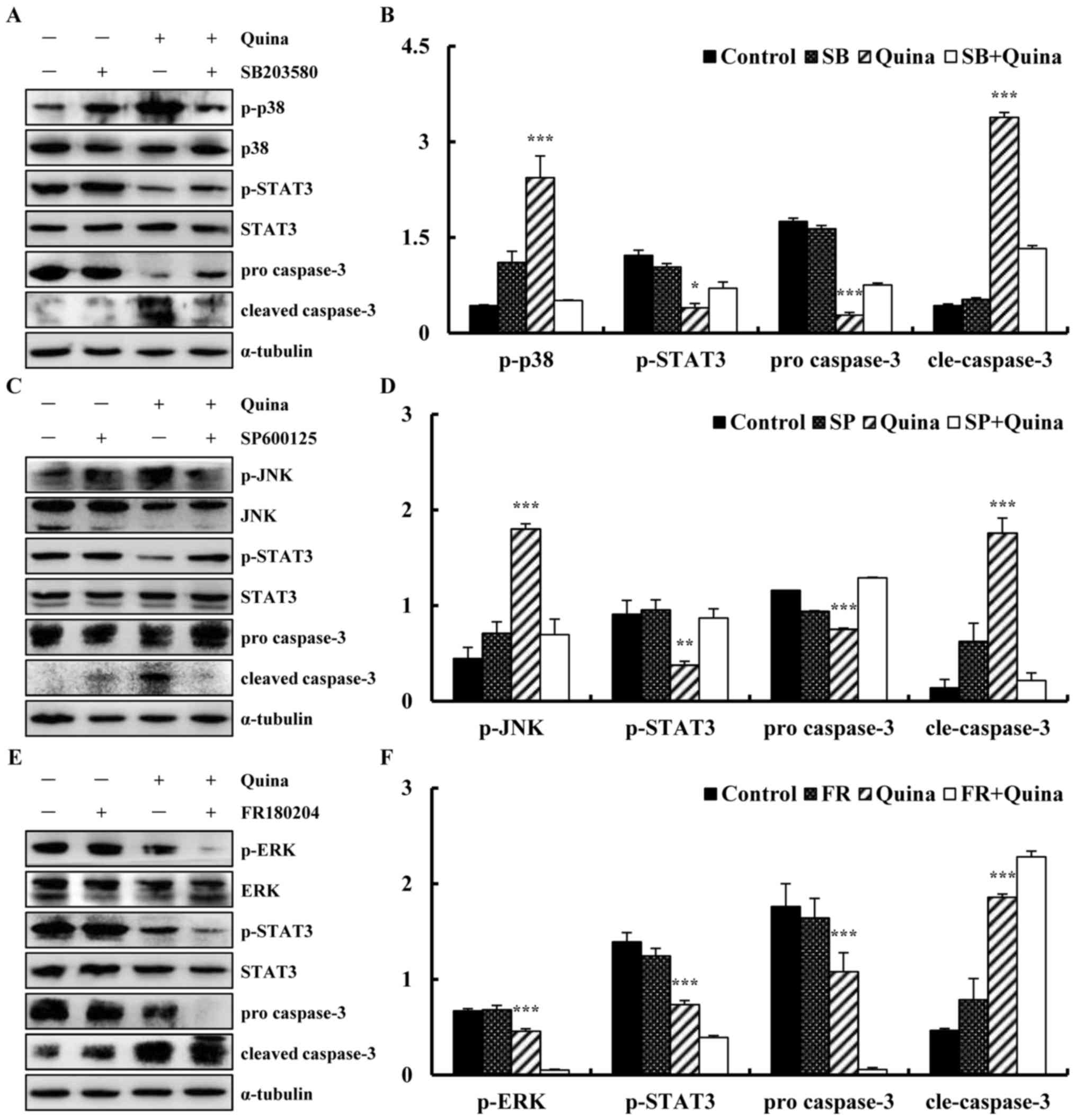 | Figure 8.Effect of quinalizarin on
mitogen-activated protein kinase and STAT3 signalling pathways in
A549 cells. (A) Expression of p-p38, p-STAT3 and cleaved caspase-3,
in quinalizarin and p38 inhibitor SB203580-treated A549 cells. (B)
Relative levels of p-p38, p-STAT3 and cleaved caspase-3 were
calculated using ImageJ. (C) Expressions of p-JNK, p-STAT3 and
cleaved caspase-3, in the quinalizarin and JNK inhibitor
SP600125-treated A549 cells. (D) Relative levels of p-JNK, p-STAT3
and cleaved caspase-3 were calculated using Image J. (E)
Expressions of p-ERK, p-STAT3 and cleaved caspase-3, in the
quinalizarin and ERK inhibitor FR180204-treated A549 cells. (F)
Relative levels of p-ERK, p-STAT3 and cleaved caspase-3 were
calculated using Image J. *P<0.05, **P<0.01 and ***P<0.001
vs. inhibitor + Quina group. SB, SB203580; SP, SP600125; FR,
FR180204; Quina, quinalizarin; STAT3, signal transducer and
activator of transcription-3; JNK, c-Jun N-terminal kinase; ERK,
extracellular signal-regulated kinases; p-, phosphorylated; cle,
cleaved. |
Discussion
Potential growth inhibition and apoptosis-inducing
effects of quinalizarin on lung cancer cell lines were explored.
Cell proliferation follows the progression of the cell cycle. In
eukaryotes, mitosis is dependent on the completion of DNA synthesis
(30). Checkpoints play important
roles in the regulation of the cell cycle in eukaryotic cells, and
abnormal regulation of these checkpoints frequently occurs in
tumour cells (31). Cells may be
arrested at G0/G1, S and G2/M
phases of the cell cycle (32).
Indeed, quinalizarin has been reported to decrease CDK1 and cdc25C
phosphatase levels and to induce apoptosis in human prostate
cancers (33). In the present
study, quinalizarin was found to induce A549 cells arrest at
G0/G1 phase, thus suggesting that
quinalizarin may induce cell cycle arrest at different phases.
CDK2/4/6 and cyclin D1/E are known to be involved in the regulation
of G0/G1 phase. Because activation of the
CDK/cyclin complex initiates entry into G1 phase, the
G0/G1 transition usually requires a
functional CDK/cyclin complex (34). Our results indicated that
quinalizarin induced A549 cell cycle arrest at
G0/G1 phase by suppressing CDK2/4/6 and
cyclin D1/E and increasing p21 and p27 protein expression
levels.
Apoptosis is a conserved programmed cell death
mechanism that is involved in the elimination of cancer cells.
Intrinsic apoptosis is also known as mitochondrial apoptosis
because it depends on factors released from the mitochondria. The
mitochondrial pathway is regulated by Bcl-2 family members, which
include pro-apoptotic Bad and anti-apoptotic Bcl-2 proteins
(35). Caspase-3 is a critical
enzyme in apoptosis that cleaves several essential cellular
proteins such as PARP (36). Many
anti-cancer agents induce the intrinsic apoptotic pathway, which is
characterized by increasing activation of caspase-3 and cleavage of
PARP. Our present study showed that quinalizarin markedly induced
the apoptosis of A549 cells by up-regulating Bad, down-regulating
the expression of Bcl-2 and activating caspase-3 and PARP. These
findings suggested that quinalizarin may induce
mitochondrial-dependent apoptosis, regulate Bcl-2 and Bad
expressions, and activate the mitochondrial cascade in lung cancer
cells.
Furthermore, the underlying molecular mechanisms of
quinalizarin-induced apoptosis indicated that quinalizarin induces
cell apoptosis via suppression of the Akt, MAPK, STAT3 and p53
signalling pathways. The Akt signalling pathway is closely
associated with CK2 and plays important roles in cell survival and
apoptosis (37). The MAPK
signalling pathways can be divided into three major subgroups,
correlated with apoptosis: the ERK, JNK and p38 pathways (14). ERK is involved in the regulation of
cell proliferation, differentiation and apoptosis in cancer cells
(15). JNK is activated by
environmental and toxic stresses and is important in inflammation
through the control of cell proliferation, differentiation,
survival and migration of various cell types (16). p38 is activated by cell
stress-induced signalling in response to various factors including
toxic chemicals and oxidative stress (17). STAT3 activation also leads to
increased cell proliferation, angiogenesis, multidrug resistance
and decreased cell apoptosis (18). p53 activates target genes involved
in cell cycle arrest, apoptosis, senescence, anti-angiogenesis and
autophagy, thereby suppressing malignant tumour transformation and
preserving genomic integrity (38). Our results revealed that
quinalizarin induces apoptosis by regulating the expression levels
of proteins involved in the Akt, MAPK, STAT3 and p53 signalling
pathways.
ROS are a group of highly reactive molecular oxygens
continuously produced during mitochondrial respiration. ROS are
usually maintained at tolerable levels under physiological
conditions, whereas high levels of ROS are an important mechanism
of cell death (39). Intracellular
ROS levels have been reported to activate the MAPK signalling
pathways (40). Imbalances due to
either increased ROS production or decreased ROS degradation may
cause excessive ROS accumulation, thus damaging to cell structures
and compromising cellular functions (41). Our study showed that NAC completely
restored the apoptotic mechanism of A549 cells induced by
quinalizarin. This finding suggested that ROS mediate cell
apoptosis. The expression levels of proteins involved in the MAPK
and STAT3 signalling pathways were detected and revealed to
increase expression levels of ERK and STAT3 and decrease expression
levels of JNK and p38 with a corresponding decrease in ROS levels.
These results revealed that quinalizarin regulates the MAPK and
STAT3 signalling pathways and is closely linked to ROS generation.
To determine whether STAT3 was inactivated in response to
quinalizarin-induced MAPK activation, the p38 inhibitor, JNK
inhibitor or ERK inhibitor was used to investigation the
interaction between MAPK and STAT3. The results showed that the
reactivation of the p38 inhibitor, JNK inhibitor or ERK inhibitor
by quinalizarin-inhibited the STAT3 signalling pathway, indicating
that STAT3 was regulated by the MAPK signalling pathway.
The present study demonstrated that quinalizarin
inhibits lung cancer A549 cell proliferation,
G0/G1 phase cell cycle arrest and apoptosis
by increasing ROS generation and the activation or inactivation of
the Akt, MAPK, STAT3 and p53 signalling pathways. This study
provides evidence that quinalizarin is a potential therapeutic
agent for the treatment of lung cancer. In future research, the
effects of quinalizarin in vivo should be evaluated.
Acknowledgements
The present study was funded by Program of
Cultivation and Support Projects of Heilongjiang Bayi Agricultural
University (XA2015-04), Nature Science Foundation of Heilongjiang
Province of China (LC2015036) and the Research Project of
Heilongjiang Bayi Agricultural University (XYB2013-24).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ruan Y, Hu K and Chen H: Autophagy
inhibition enhances isorhamnetin-induced mitochondria-dependent
apoptosis in non-small cell lung cancer cells. Mol Med Rep.
12:5796–5806. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao GF, Huang ZA, Du XK, Yang ML, Huang
DD and Zhang S: Molecular docking studies of traditional chinese
medicinal compounds against known protein targets to treat
non-small cell lung carcinomas. Mol Med Rep. 14:1132–1138. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dai GH, Meng GM, Tong YL, Chen X, Ren ZM,
Wang K and Yang F: Growth-inhibiting and apoptosis-inducing
activities of Myricanol from the bark of Myrica rubra in human lung
adenocarcinoma A549 cells. Phytomedicine. 21:1490–1496. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu K, Liu B and Liu Y: Impact of Brachyury
on epithelial-mesenchymal transitions and chemosensitivity in
non-small cell lung cancer. Mol Med Rep. 12:995–1001. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo W, Xie L, Zhao L and Zhao Y: mRNA and
microRNA expression profiles of radioresistant NCI-H520 non-small
cell lung cancer cells. Mol Med Rep. 12:1857–1867. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen X, Yang Z, Sun R, Mo Z, Jin G, Wei F,
Hu J, Guan W and Zhong N: Preparation of lung-targeting,
emodin-loaded polylactic acid microspheres and their properties.
Int J Mol Sci. 15:6241–6251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Y, Guo G, Song J, Cai Z, Yang J, Chen
Z, Wang Y, Huang Y and Gao Q: B7-H3 promotes the migration and
invasion of human bladder cancer cells via the PI3K/Akt/STAT3
signaling pathway. J Cancer. 8:816–824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu S, Huang L, Meng L, Sun H, Zhang W and
Xu Y: Isorhamnetin inhibits cell proliferation and induces
apoptosis in breast cancer via Akt and mitogen-activated protein
kinase kinase signaling pathways. Mol Med Rep. 12:6745–6751. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang G, Wang C, Sun M, Li J, Wang B, Jin
C, Hua P, Song G, Zhang Y, Nguyen LL, et al: Cinobufagin inhibits
tumor growth by inducing intrinsic apoptosis through AKT signaling
pathway in human nonsmall cell lung cancer cells. Oncotarget.
7:28935–28946. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang D, Chen B, Zhou J, Zhou L, Li Q, Liu
F, Chou KY, Tao L and Lu LM: Low concentrations of trichosanthin
induce apoptosis and cell cycle arrest via c-Jun N-terminal protein
kinase/mitogen-activated protein kinase activation. Mol Med Rep.
11:349–356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zheng Y, McFarland BC, Drygin D, Yu H,
Bellis SL, Kim H, Bredel M and Benveniste EN: Targeting protein
kinase CK2 suppresses prosurvival signaling pathways and growth of
glioblastoma. Clin Cancer Res. 19:6484–6494. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pan B, Zhong W, Deng Z, Lai C, Chu J, Jiao
G, Liu J and Zhou Q: Inhibition of prostate cancer growth by
solanine requires the suppression of cell cycle proteins and the
activation of ROS/P38 signaling pathway. Cancer Med. 5:3214–3222.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhai H, Hu S, Liu T, Wang F, Wang X, Wu G,
Zhang Y, Sui M, Liu H and Jiang L: Nitidine chloride inhibits
proliferation and induces apoptosis in colorectal cancer cells by
suppressing the ERK signaling pathway. Mol Med Rep. 13:2536–2542.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park KR, Yun HM, Quang TH, Oh H, Lee DS,
Auh QS and Kim EC: 4-Methoxydalbergione suppresses growth and
induces apoptosis in human osteosarcoma cells in vitro and in vivo
xenograft model through down-regulation of the JAK2/STAT3 pathway.
Oncotarget. 7:6960–6971. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsai WC, Bai LY, Chen YJ, Chu PC, Hsu YW,
Sargeant AM and Weng JR: OSU-A9 inhibits pancreatic cancer cell
lines by modulating p38-JAK-STAT3 signaling. Oncotarget.
8:29233–29246. 2017.PubMed/NCBI
|
|
18
|
Miao D and Zhang L: Leptin modulates the
expression of catabolic genes in rat nucleus pulposus cells through
the mitogen-activated protein kinase and Janus kinase 2/signal
transducer and activator of transcription 3 pathways. Mol Med Rep.
12:1761–1768. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Principe M, Borgoni S, Cascione M,
Chattaragada MS, Ferri-Borgogno S, Capello M, Bulfamante S,
Chapelle J, Di Modugno F, Defilippi P, et al: Alpha-enolase (ENO1)
controls alpha v/beta 3 integrin expression and regulates
pancreatic cancer adhesion, invasion, and metastasis. J Hematol
Oncol. 10:162017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi XY, Xiong LX, Xiao L, Meng C, Qi GY
and Li WL: Downregulation of caveolin-1 upregulates the expression
of growth factors and regulators in co-culture of fbroblasts with
cancer cells. Mol Med Rep. 13:744–752. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Khan M, Khan M, Al-Marri AH, Al-Warthan A,
Alkhathlan HZ, Siddiqui MR, Nayak VL, Kamal A and Adil SF:
Apoptosis inducing ability of silver decorated highly reduced
graphene oxide nanocomposites in A549 lung cancer. Int J
Nanomedicine. 11:873–883. 2016.PubMed/NCBI
|
|
22
|
Chen Z, Teo AE and McCarty N: ROS induced
CXCR4 signaling regulates mantle cell lymphoma (MCL) cell survival
and drug resistance in the bone marrow microenvironment via
autophagy. Clin Cancer Res. 22:187–199. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang W, Zhang Q, Jiang Y, Li F and Xin H:
Effects of ophiopogonin B on the proliferation and apoptosis of
SGC-7901 human gastric cancer cells. Mol Med Rep. 13:4981–4986.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Y, Liu JM, Xiong XX, Qiu XY, Pan F,
Liu D, Lan SJ, Jin S, Yu SB and Chen XQ: Piperlongumine selectively
kills hepatocellular carcinoma cells and preferentially inhibits
their invasion via ROS-ER-MAPKs-CHOP. Oncotarget. 6:6406–6421.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cozza G, Mazzorana M, Papinutto E, Bain J,
Elliott M, di Maira G, Gianoncelli A, Pagano MA, Sarno S, Ruzzene
M, et al: Quinalizarin as a potent, selective and cell-permeable
inhibitor of protein kinase CK2. Biochem J. 421:387–395. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song C, Gowda C, Pan X, Ding Y, Tong Y,
Tan BH, Wang H, Muthusami S, Ge Z, Sachdev M, et al: Targeting
casein kinase II restores Ikaros tumor suppressor activity and
demonstrates therapeutic efficacy in high-risk leukemia. Blood.
126:1813–1822. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hung MS, Xu Z, Chen Y, Smith E, Mao JH,
Hsieh D, Lin YC, Yang CT, Jablons DM and You L: Hematein, a casein
kinase II inhibitor, inhibits lung cancer tumor growth in a murine
xenograft model. Int J Oncol. 43:1517–1522. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou Y, Li K, Zhang S, Li Q, Li Z, Zhou F,
Dong X, Liu L, Wu G and Meng R: Quinalizarin, a specific CK2
inhibitor, reduces cell viability and suppresses migration and
accelerates apoptosis in different human lung cancer cell lines.
Indian J Cancer. 2 Suppl 52:e119–e124. 2015.
|
|
29
|
Kim J, Choi WJ, Moon SH, Jung J, Park JK,
Kim SH and Lee JO: Micropillar arrays as potential drug screens:
Inhibition of micropillar-mediated activation of the
FAK-Src-paxillin signaling pathway by the CK2 inhibitor CX-4945.
Acta Biomater. 27:13–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang NH, Shin HC, Oh S, Lee KH, Lee YB and
Choi KC: Soy milk digestion extract inhibits progression of
prostate cancer cell growth via regulation of prostate
cancer-specific antigen and cell cycle-regulatory genes in human
LNCaP cancer cells. Mol Med Rep. 14:1809–1816. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu L, Wang D, Li L, Ding X and Ma H:
Dehydroepiandrosterone inhibits cell proliferation and improves
viability by regulating S phase and mitochondrial permeability in
primary rat Leydig cells. Mol Med Rep. 14:705–714. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen L, Tian H, Li M, Ge C, Zhao F, Zhang
L, Li H, Liu J, Wang T, Yao M and Li J: Derivate isocorydine
inhibits cell proliferation in hepatocellular carcinoma cell lines
by inducing G2/M cell cycle arrest and apoptosis. Tumour Biol.
37:5951–5961. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schneider CC, Götz C, Hessenauer A,
Günther J, Kartarius S and Montenarh M: Down-regulation of CK2
activity results in a decrease in the level of cdc25C phosphatase
in different prostate cancer cell lines. Mol Cell Biochem.
356:177–184. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shi W, Deng J, Tong R, Yang Y, He X, Lv J,
Wang H, Deng S, Qi P, Zhang D and Wang Y: Molecular mechanisms
underlying mangiferin-induced apoptosis and cell cycle arrest in
A549 human lung carcinoma cells. Mol Med Rep. 13:3423–3432. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Woo SM, Choi YK, Kim AJ, Cho SG and Ko SG:
p53 causes butein-mediated apoptosis of chronic myeloid leukemia
cells. Mol Med Rep. 13:1091–1096. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu Y, Chen Y, Wu Q, Jia L and Du X:
Minocycline inhibits PARP-1 expression and decreases apoptosis in
diabetic retinopathy. Mol Med Rep. 12:4887–4894. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zeng J, Chen S, Li N, Chen L, Su J, Niu G,
Zhu S and Liang Y: Sasanquasaponin from Camellia oleifera Abel.
induces apoptosis via Bcl-2, Bax and caspase-3 activation in HepG2
cells. Mol Med Rep. 12:1997–2002. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jin J, Lin G, Huang H, Xu D, Yu H, Ma X,
Zhu L, Ma D and Jiang H: Capsaicin mediates cell cycle arrest and
apoptosis in human colon cancer cells via stabilizing and
activating p53. Int J Biol Sci. 10:285–295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Conway GE, Casey A, Milosavljevic V, Liu
Y, Howe O, Cullen PJ and Curtin JF: Non-thermal atmospheric plasma
induces ROS-independent cell death in U373MG glioma cells and
augments the cytotoxicity of temozolomide. Br J Cancer.
114:435–443. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao W, Lu M and Zhang Q: Chloride
intracellular channel 1 regulates migration and invasion in gastric
cancer by triggering the ROS-mediated p38 MAPK signaling pathway.
Mol Med Rep. 12:8041–8047. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Aredia F, Czaplinski S, Fulda S and
Scovassi AI: Molecular features of the cytotoxicity of an NHE
inhibitor: Evidence of mitochondrial alterations, ROS
overproduction and DNA damage. BMC Cancer. 16:8512016. View Article : Google Scholar : PubMed/NCBI
|