Introduction
Hepatocellular carcinoma (HCC) ranked as the 5th
most frequent cancer and was also one of the lethal cancers,
particularly in People's Republic of China where liver cancer was
the most commonly diagnosed cancer and the most prevalent cause of
cancer-related deaths followed by lung, stomach, and esophageal
cancers (based on the statistics in 2015) (1,2).
Because HCC was often diagnosed at an advanced stage, the prognosis
of HCC patients was not optimistic (3). Therefore, a better understanding of
the pathogenesis of HCC and a novel target for the early screening
of HCC might improve the survival of HCC patients (4).
MicroRNAs (miRNAs) are small non-coding RNAs (18–25
nucleotides in length) that regulate the expression of multiple
mRNAs at the post-transcriptional level by suppressing the
stability and the translation of mRNAs (5,6). The
aberrant expressions of miRNAs was observed in various human
cancers, and extensive studies suggested that these deregulated
miRNAs had the capacity to distinguish malignant tumors of liver,
breast, lung, pancreas and leukemia from adjacent non-tumorous
tissue (7–12). miR-338-3p and miR-338-5p originate
from an intron of the gene encoding apoptosis-associated tyrosine
kinase (AATK). Both miR-338-3p and miR-338-5p are co-expressed
because they enjoyed the same promoter together (13). miR-338 was the prvious ID of
miR-338-3p, which had been reported in variety of diseases
(13–16). miR-338* is one of the members of
miR-338 family and usually represented as miR-338-5p (17). As a member of the miRNA family,
miR-338-5p was reported to be correlated with the carcinogenesis
and progression of several human cancers including gastric cancer
(18), colorectal cancer (19), and glioblastoma (20). However, there were limited studies
on the clinical significance of miR-338-5p in HCC. Chen et
al reported the overexpression of miR-338-5p in tumor tissues
of the liver and preoperative plasma by miRNA array in Asian
patients (21). Whether miR-338-5p
is indeed a qualified diagnostic biomarker for HCC and the
underlying molecular mechanism of miR-338-5p in HCC remained
unclarified.
Therefore, this study aimed to investigate the
diagnostic significance of miR-338-5p in HCC tissues and the
molecular mechanism of miR-338-5p in HCC with a combination of
meta-analysis and bioinformatics analysis. Our study confirmed the
significance of miR-338-5p for the diagnosis of HCC and might
promote the understanding of the molecular mechanism underlying it.
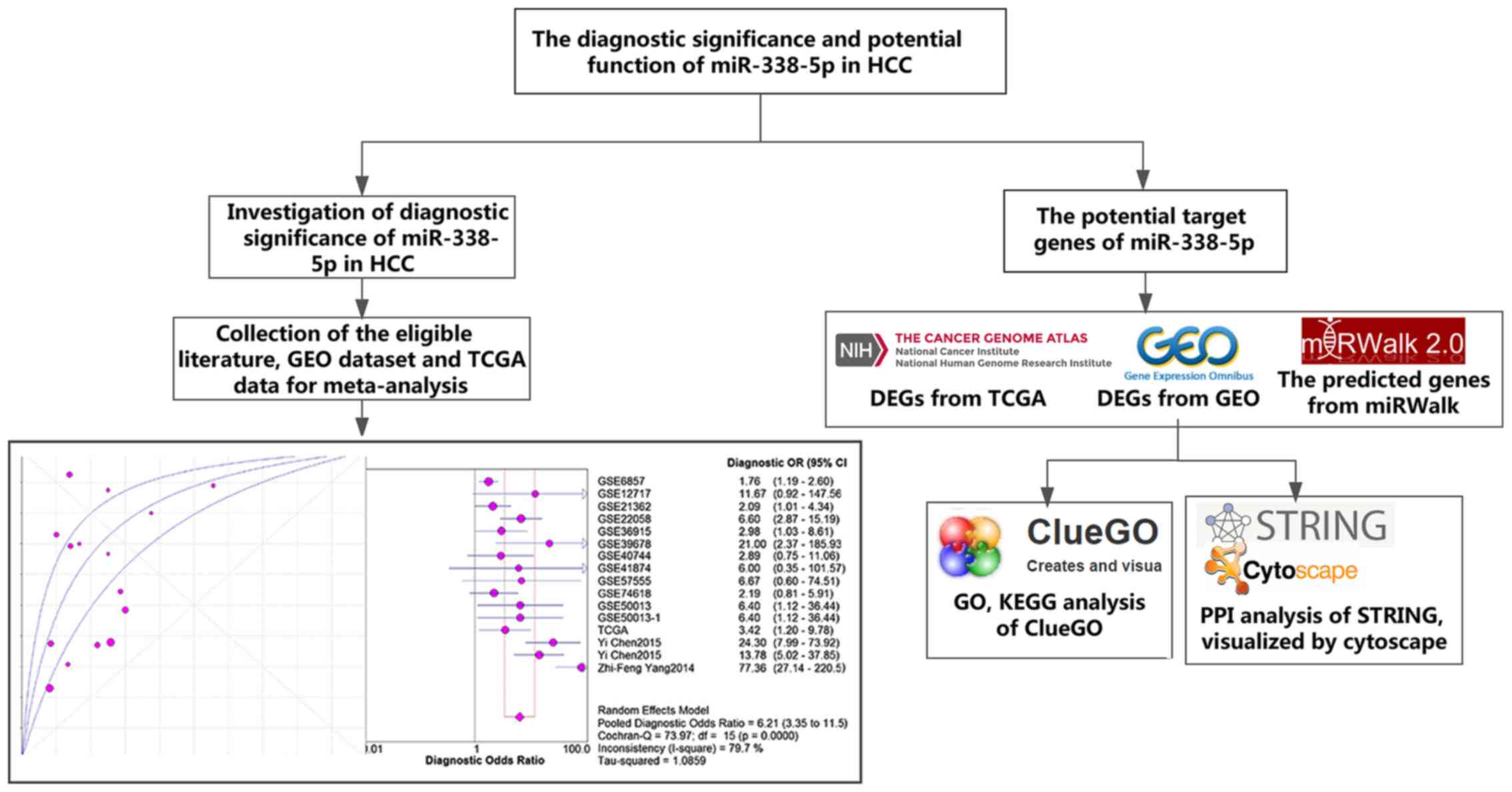
The framework of this article was displayed in Fig. 1.
Materials and methods
The process of study selection
In order to obtain the comprehensive data of the
diagnostic value of miR-338-5p in HCC, a thorough search for the
related studies was conducted in Gene Expression Omnibus (GEO)
dataset and other database including PubMed, Embase, Cochrane, Web
of Science, Sinomed, Chinese VIP, Wanfang database and China
National Knowledge Infrastructure (CNKI) until December 15, 2016
with the searching strategies: (miR-338 or miRNA-338 or
microRNA-338 or miR338 or miRNA338 or microRNA338 or ‘miR 338’ or
‘miRNA 338’ or ‘microRNA 338’) and (malignan* or cancer or tumor or
tumour or neoplas* OR carcinoma) AND (hepatocellular or liver or
hepatic or HCC). Studies that meet the following criteria were
eligible for the meta-analysis: i) Studies evaluated the expression
of miR-338-5p for the diagnosis of HCC; ii) the disease of the
patients were validated with golden standard; iii) the number of
cases were reported in the study; and iv) the sensitivity and
specificity of the diagnostic test were available directly or
indirectly from the study. The exclusion criteria of the studies
were as follows: i) The content of the studies were irrelevant with
HCC; ii) the subjects of experiment were not human beings; iii)
there was no sufficient data for researchers to directly acquire or
calculate sensitivity or specificity of the diagnostic test; and
iv) studies were classified as review, meta-analysis, case study or
conference note. Moreover, data of the diagnostic value of
miR-338-5p in HCC was also downloaded from the cancer genome atlas
(TCGA) (https://cancergenome.nih.gov/).
Data extraction and statistical
analysis
The following information and data were extracted
from the included studies: The ID no. of each GSE chip, first
author, year of publication, country, experiment type, platform of
each GSE chip, sample number for the experiment group and control
group, tissue types, true positivity (TP), false positivity (FP),
false negativity (FN) and true negativity (TN).
MetaDiSc1.4 and STATA12.0 were applied for all the
statistical analysis. To explore expression of miR-338-5p, the
continuous outcomes of GEO and TCGA datasets were calculated with
standard mean difference (SMD). The sensitivity (SEN), specificity
(SPE), positive likelihood ratio (PLR), negative likelihood ratio
(NLR) and diagnostic odds ratio (DOR) of the included studies were
pooled with the bivariate meta-analysis model (22,23).
The summary receiver operator characteristic (SROC) curve was
plotted according to the sensitivity and specificity from each
study. The area under the SROC curve (AUC) calculated from the SROC
reflected the capacity of miR-338-5p to differentiate HCC patients
from non-cancer patients accurately. An AUC value of 0.5 or 1.0
represents a poor or perfect diagnostic value, respectively
(24). Additionally, Q test and
I2 statistics were employed to assess the heterogeneity
between studies. The random-effects model would be used to pool the
results if an I2 value was more than 50% with a P-value
<0.10; otherwise, a fixed-effects model would be applied
(25,26). To identify the source of
heterogeneity, the subgroup analysis was conducted based on the
number and features of the included studies. With regard to the
publication bias, the Deeks' funnel plot asymmetry test was carried
out to detect the publication bias, and P-value <0.05 was
indicative of significance.
The target genes of miR-338-5p
The potential target genes of miR-338-5p came from
three sources: The differentially expressed genes from GEO, TCGA
and the predicted genes from 12 online software (miRWalk, MicroT4,
miRanda, mirBridge, miRDB, miRMap, miRNAMap, PicTar2, PITA, RNA22,
RNAhybrid and TargetScan). We firstly searched Gene Expression
Omnibus (GEO) datasets for deregulated target genes of miR-338-5p
from the mRNA profiling data of HCC samples on December 15, 2016.
All the GSE chips shared the same platform: GPL570 (Affymetrix
Human Genome U133 Plus 2.0 Array). After preliminary screening, 54
studies remained for further selection. Among the 54 studies,
Homo sapiens tissue samples instead of cell lines samples
were included for further analysis. Finally, 10 HCC datasets
(GSE29721, GSE45436, GSE55092, GSE62232, GSE9843, GSE41804,
GSE6764, GSE33006, GSE6222 and GSE19665) comprising 431 HCC samples
and 198 control samples were chosen for further analysis.
Differentially expressed genes (DEGs) between cancer and normal
samples of 10 datasets were acquired via GCBI online tool
(https://www.gcbi.com.cn/gclib/html/index). Fold-change
>1.5, and a P-value <0.05 was set as the threshold for the
DEGs. Another database containing high-throughput data: TCGA was
also searched. Publicly available miRNA-seq and RNA-seq data of
liver HCC was downloaded from the TCGA data portal (December 2016,
https://gdc-portal.nci.nih.gov/). Since
the TCGA data were a community resource project, additional
approval by an ethics committee of our hospital was not mandatory.
And the present study adhered to the TCGA publication guidelines
and data access policies. From the downloaded data of 377 HCC
samples and 50 normal liver samples. R language package DESeq was
subsequently used for the calculation of DEGs (Padj <0.05 and
the absolute log2 fold-change >1). As for the predicted genes,
selection was based on the condition that they were recorded in
more than 4 of the 12 prediction software. The selected qualified
target genes from the online software and the validated target
genes from miRWalk were considered as the potential target genes of
miR-338-5p.
The protein-protein-interaction (PPI)
network and validation of target genes
To illustrate the interaction between the targets of
miR-338-5p, a PPI network was drawn by Cytoskape v.5.3.0. The nodes
and edges represented target genes and the interactions between
target genes, respectively. Hub genes were identified according to
the value of degrees of each node. Protein expression of hub gene
was validated by The Human Protein Atlas (HPA), an
immunohistochemisty (IHC) database (27). Each antibody in the database has
been used for IHC staining of both normal and HCC tissues.
The Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analysis of the target genes
The GO and KEGG pathway analysis were performed by
the BiNGO and ClueGO plug-in unit in Cytoscape v.3.5.0 for the
functional annotation of the target genes. Three GO terms including
biological process (BP), cellular component (CC) and molecular
function (MF) were utilized to identify the enrichment of target
genes. P-value <0.05 was significant.
Results
Eligible studies for the
meta-analysis
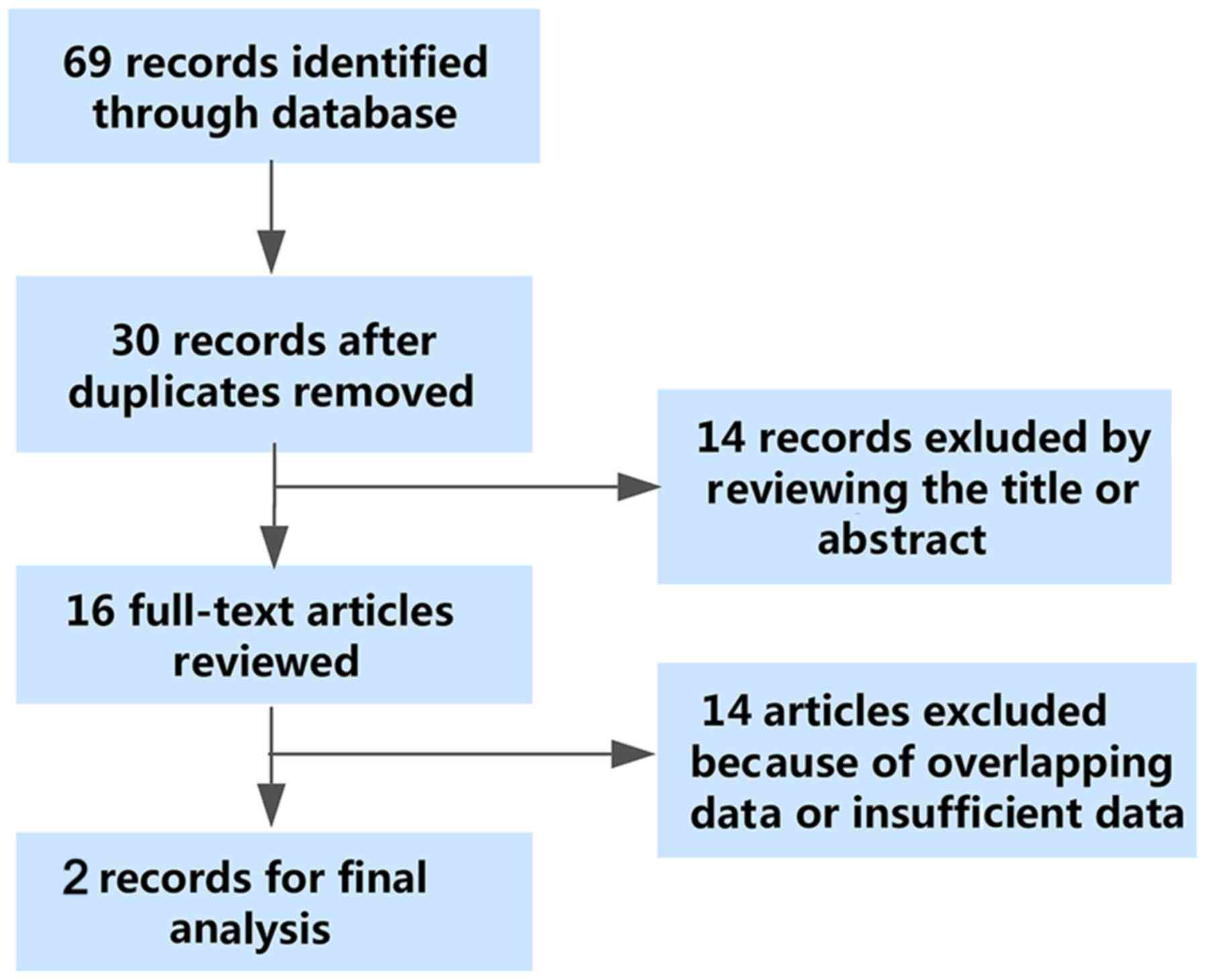
As shown in Fig. 2,
the flowchart exhibited the selection and retrieval process of the
qualified studies. A total of 69 studies were identified as the
initial records, and 30 studies remained after the removal of
duplicate records. Then, 14 records were excluded in the
preliminary screening of the titles and abstracts of the articles.
As a consequence, 16 studies were reviewed in the full text. Among
the 16 studies, 14 studies were ineligible due to insufficient data
of the diagnostic parameters or duplicate data. Eventually, two
studies were enrolled for the meta-analysis. Though two included
studies were conducted by the same authors, we failed to validate
that the two studies shared the same patient cohorts. Thus, we
regard the two studies of Chen et al as two different
studies (21,28).
Assessment of the diagnostic value and
the integrated meta-analysis
To comprehensively evaluate the diagnostic value of
miR-338-5p, we supplemented the literature analysis with GEO data
and TCGA data. We searched the GEO dataset with the same searching
strategies in literature meta-analysis. Finally, a total of
eligible 11 GSE chips were included in our meta-analysis (29–38)
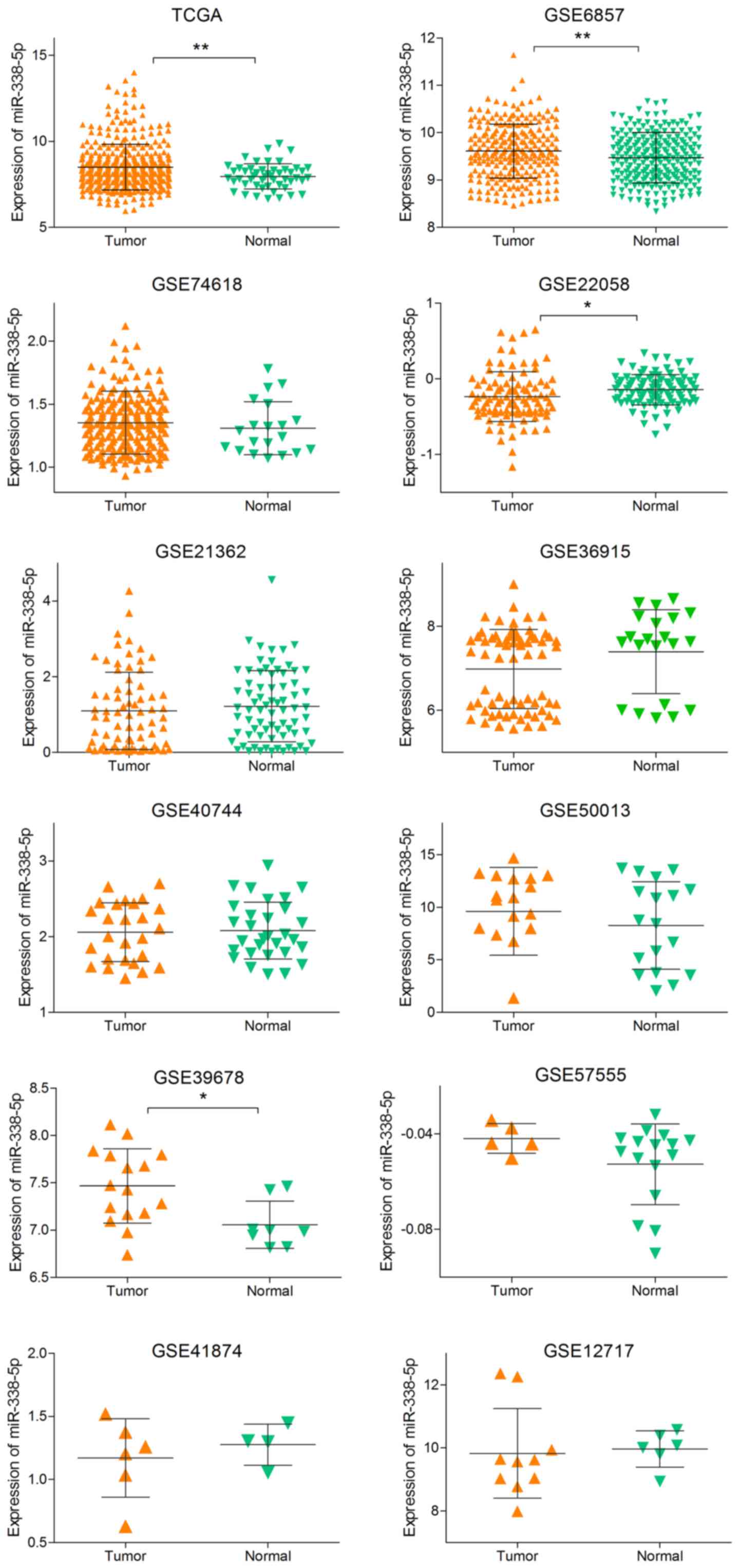
(Table I), and the expression
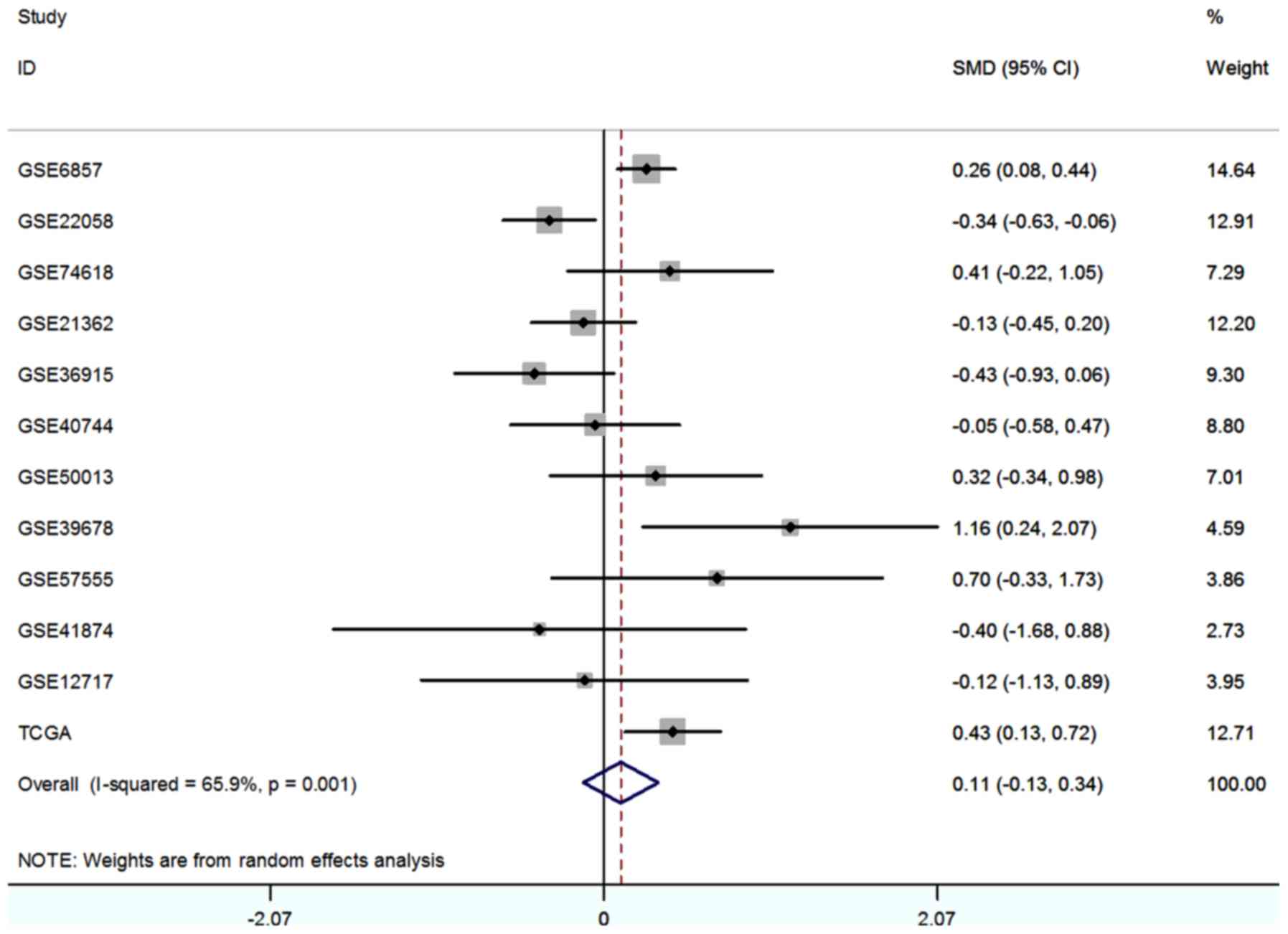
level of each study were showed in Fig. 3. With the random-effects model, the
forest-plot represented that no significant difference expression
was observed between HCC tissue and normal tissue. The pooled SMD
(0.11, 95% CI: −0.13, 0.34) was showed in Fig. 4.
 | Table I.Basic information and clinical data
of the included studies. |
Table I.
Basic information and clinical data
of the included studies.
| GEO accession | Author, year | Public year | Country | Experiment
type | Platform | HCC | Control | Sample type | TP | FP | FN | TN | (Refs.) |
|---|
| GSE6857 | Budhu et al,
2008 | 2008 | USA | Non-coding RNA
profiling by array | GPL4700 | 240 | 241 | Tissue | 91 | 62 | 149 | 179 | (29) |
| GSE12717 | Su et al,
2009 | 2008 | USA | Non-coding RNA
profiling by array | GPL7274 | 10 |
6 | Tissue |
7 | 1 |
3 |
5 | (30) |
| GSE22058 | Burchard et
al, 2010 | 2010 | USA | Non-coding RNA
profiling by array | GPL10457 | 96 | 96 | Tissue | 36 | 8 | 60 | 88 | (31) |
| GSE40744 | Diaz et al,
2013 | 2013 | USA | Non-coding RNA
profiling by array | GPL14613 | 26 | 19 | Tissue |
8 | 4 | 18 | 26 | (32) |
| GSE50013 | Shen et al,
2013 | 2013 | USA | Non-coding RNA
profiling by array | GPL15497 | 18 | 18 | Plasma | 16 | 10 |
2 |
8 | (33) |
| GSE41874 | Morita
(unpublished) | 2013 | Japan | Non-coding RNA
profiling by array | GPL7722 |
6 |
4 | Tissue |
4 | 1 |
2 |
3 | – |
| GSE57555 | Murakami et
al, 2015 | 2015 | Japan | Non-coding RNA
profiling by array | GPL16699 |
5 | 16 | Tissue |
4 | 6 |
1 | 10 | (34) |
| GSE74618 | Martinez-Quetglas
et al, 2016 | 2016 | Spain | Non-coding RNA
profiling by array | GPL14613 | 223 | 20 | Tissue | 131 | 10 | 92 | 10 | (35) |
| GSE36915 | Shih et al,
2012 | 2012 | Taiwan | Non-coding RNA
profiling by array | GPL8179 | 68 | 21 | Tissue | 37 | 6 | 31 | 15 | (36) |
| GSE39678 | Noh et al,
2013 | 2013 | South Korea | Non-coding RNA
profiling by array | GPL15852 | 16 |
8 | Tissue | 14 | 2 |
2 |
6 | (37) |
| GSE21362 | Sato et al,
2011 | 2011 | Japan | Non-coding RNA
profiling by array | GPL10312 | 73 | 73 | Tissue | 27 | 16 | 46 | 57 | (38) |
| TCGA |
|
|
| miRNA-Seq | Illumina | 375 | 50 | Tissue | 156 | 8 | 218 | 42 |
|
| Literaure | Chen et al,
2015 | 2015 | China | qRT-PCR |
| 37 | 31 | Plasma | 27 | 0 | 10 | 31 | (21) |
| Literaure | Chen et al,
2015 | 2015 | China | qRT-PCR |
| 39 | 25 | Plasma | 35 | 4 |
4 | 21 | (28) |
From the chi-square test and I2 test,
significant heterogeneity existed in all the pooled effects (SE,
SP, PLR, NLR and DOR) between studies (All I2>50%;
P<0.05). Therefore, random effects model were employed to
estimate the overall SE, SP, PLR, NLR and DOR of all the data. As
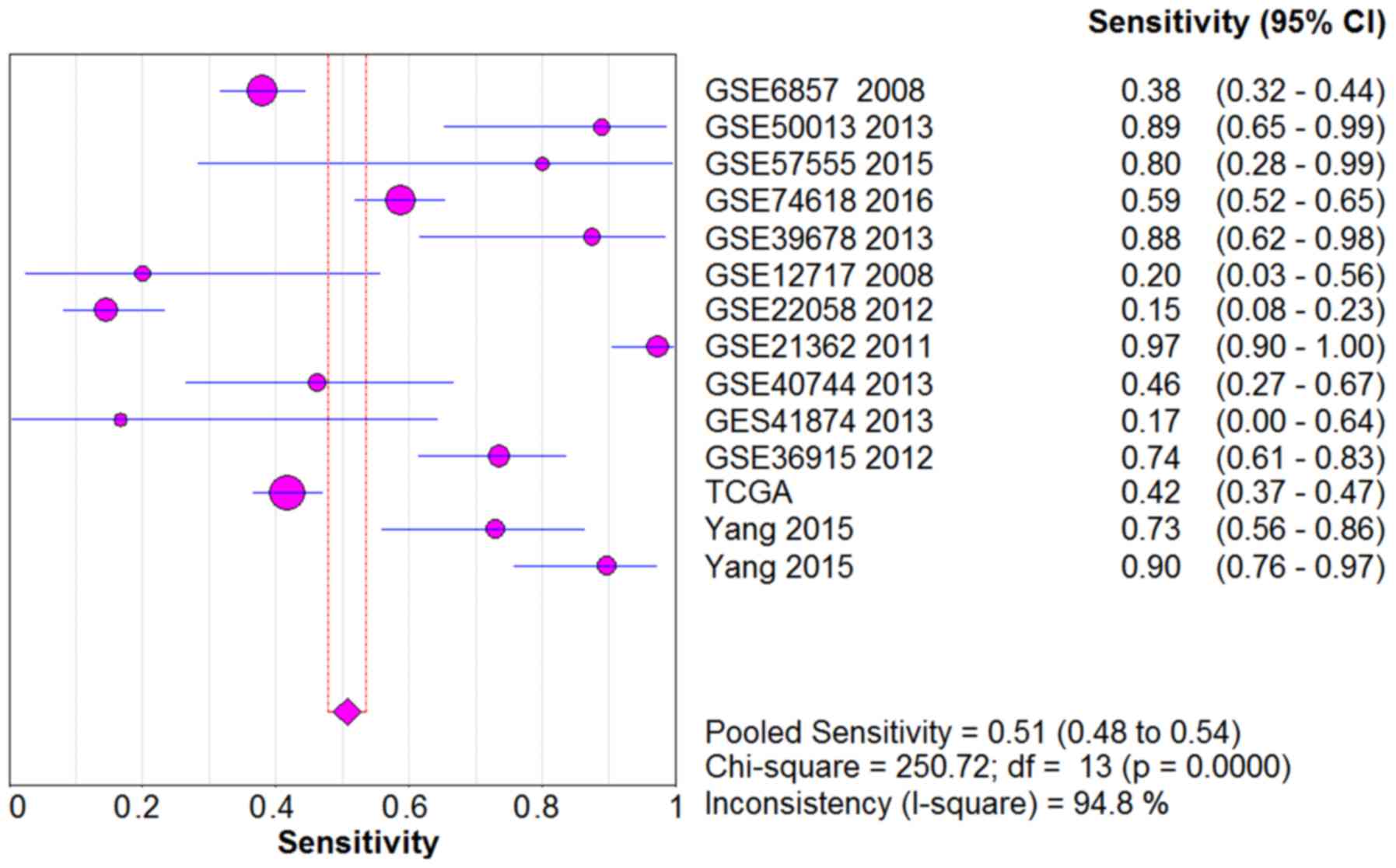
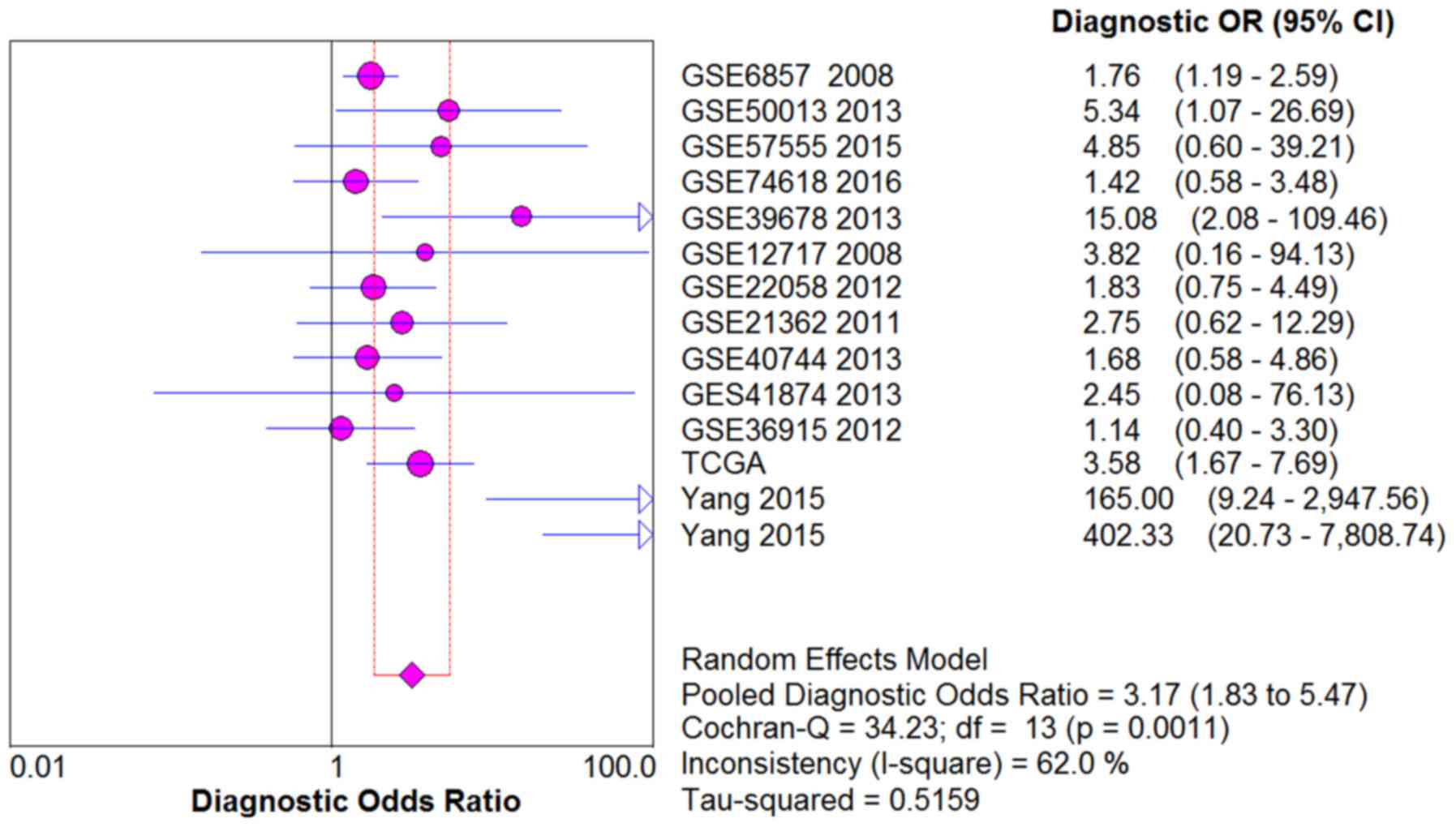
shown in Figs. 5–9, the SE, SP, PLR, NLR and DOR of all the
studies were 0.51 (95% CI: 0.48–0.54), 0.69 (95% CI: 0.65–0.73),
1.76 (95% CI: 1.17–2.66), 0.64 (95% CI: 0.52–0.80) and 3.17 (95%
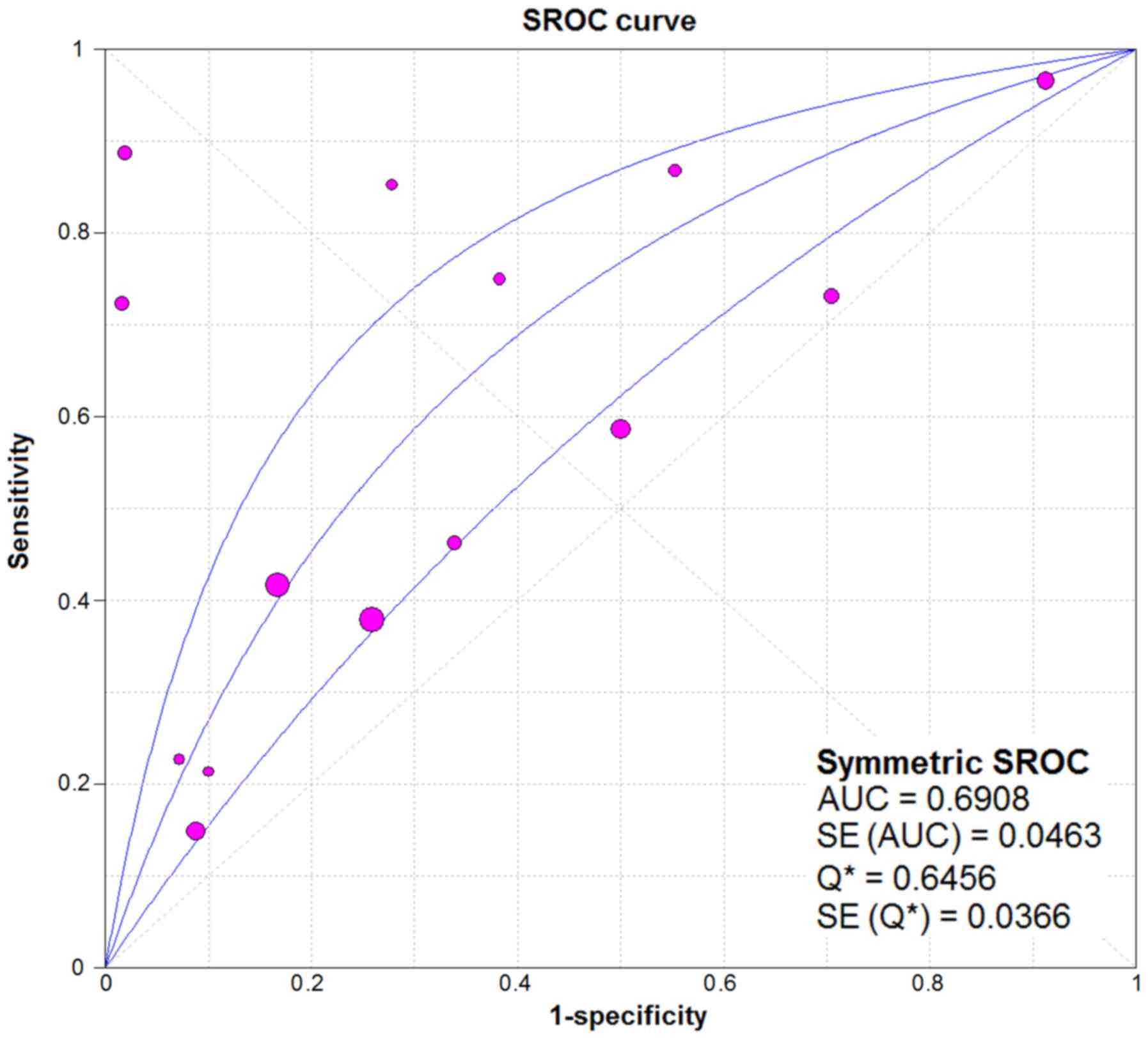
CI: 1.83–5.47). As for the result of SROC, the AUC value of
miR-338-5p was 0.691 (Fig. 10).
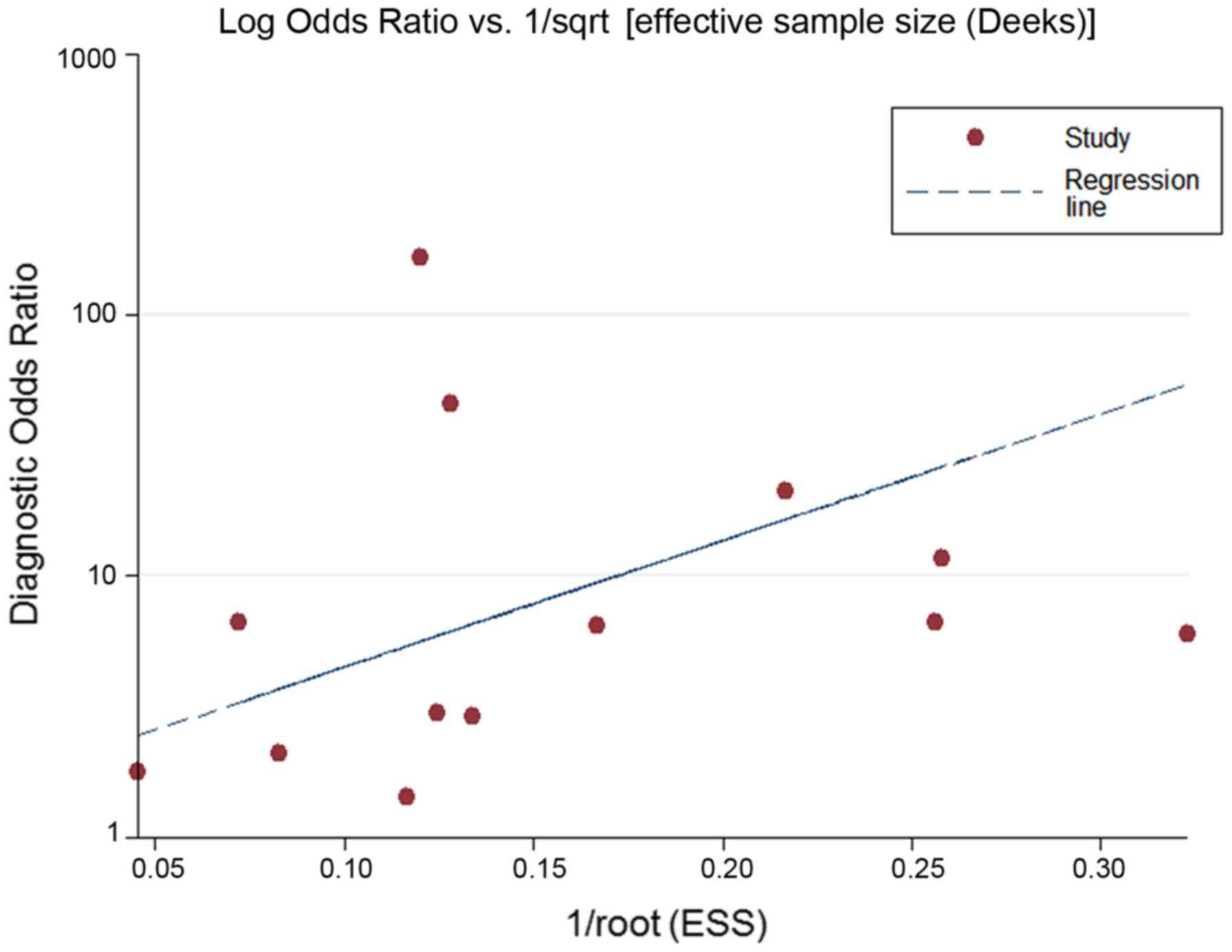
Moreover, the Deeks funnel plot asymmetry test was carried out with
Stata 12.0, and no publication bias was detected (P>0.05)
(Fig. 11).
Now that significant heterogeneity existed between
the studies, the subgroup analysis was performed to seek the
potential sources of heterogeneity. In the subgroup of sample
types, the heterogeneity decreased substantially in the pooling
estimates of NLR (49.2%) and DOR (5.6%) in the group of tissue. The
value of SE, SP, PLR and DOR were obviously higher in the plasma
group (0.83, 0.74–0.90; 0.86, 0.77–0.93; 14.02, 0.08–2,395.96;
61.30, 3.61–1,040.31) than in the tissue group (0.48, 0.45–0.51;
0.67, 0.63–0.71; 1.51, 1.08–2.11; 2.05, 1.51–2.77) and the value of
NLR was notably lower in the plasma group (0.21, 0.11–0.38) than in
the tissue group (0.81, 0.71–0.92).
With regard to the subgroup of experiment, the
heterogeneity decreased substantially in the pooling estimate of
DOR (10.3%) in the microarray group, and declined heterogeneity of
SE (0%), PLR (0%) and DOR (0.0%) were also observed in the group of
qRT-PCR. The value of SE, SP, PLR and DOR were obviously higher in
the qRT-PCR group (0.82, 0.71–0.90; 1.00, 0.94–1.00; 46.23,
6.60–323.68; 254.42, 32.2–2,010.45) than in the microarray group
(0.49, 0.46–0.52; 0.66, 0.62–0.70; 1.51, 1.11–2.06; 2.15,
1.56–2.97) and the value of NLR was notably lower in the qRT-PCR
group (0.19, 0.08–0.47) than in the microarray group (0.79,
0.69–0.91). This result confirmed that types of sample and
experiment were the possible sources of heterogeneity in this
study.
Bioinformatics study of the target
genes of miR338-5p
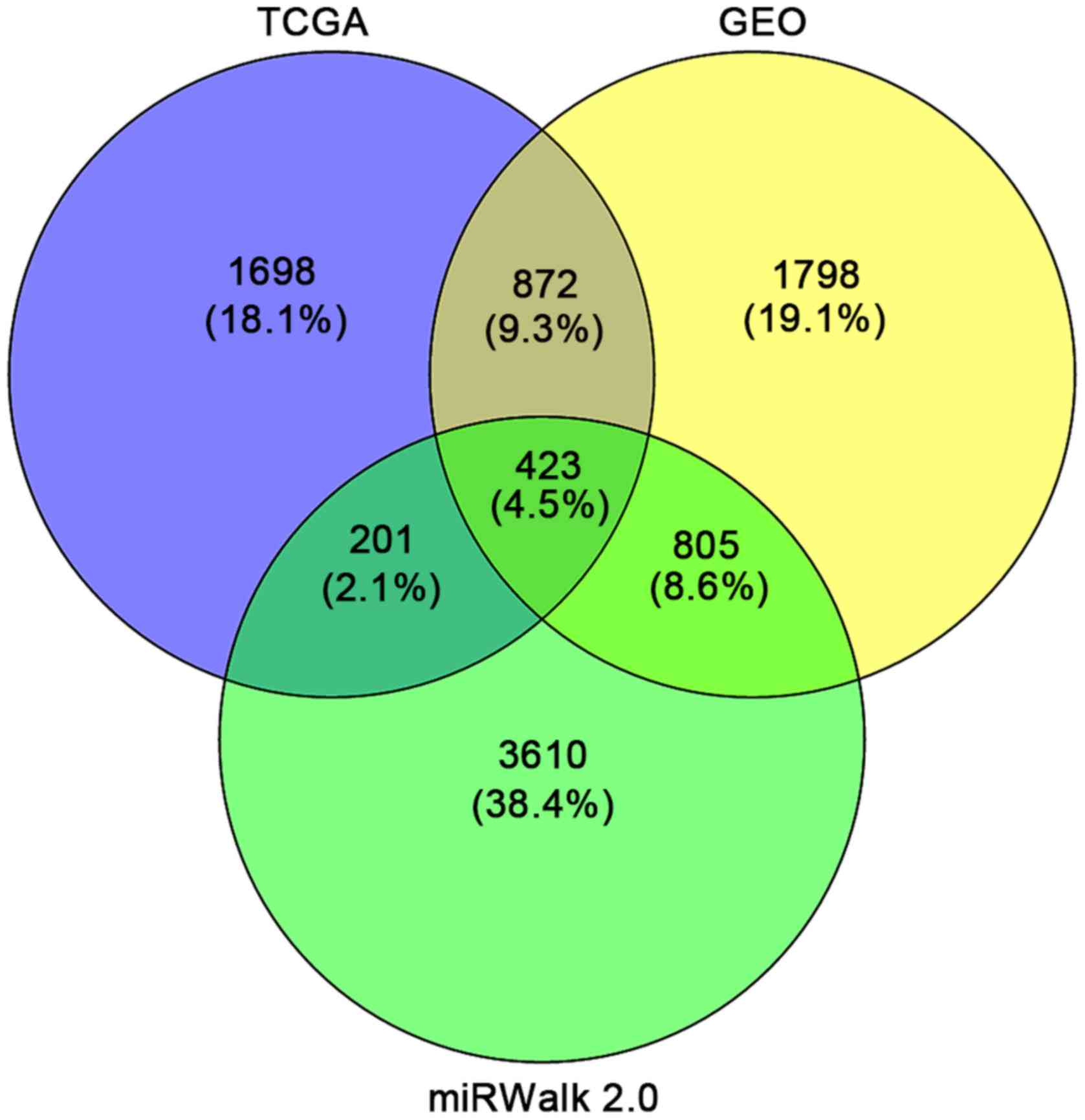
According to the results, a total of 1,698 and 1,798
genes were identified as DEGs targeted by miR-338-5p from TCGA and
GEO, respectively. Additionally, a total of 3,610 predicted target
genes that appeared in more than four times of the 12 online
software were obtained. Taking the intersection of the DEGs from
GEO and TCGA as well as the qualified predicted targets genes, we
selected 423 genes for the following bioinformatics analyses
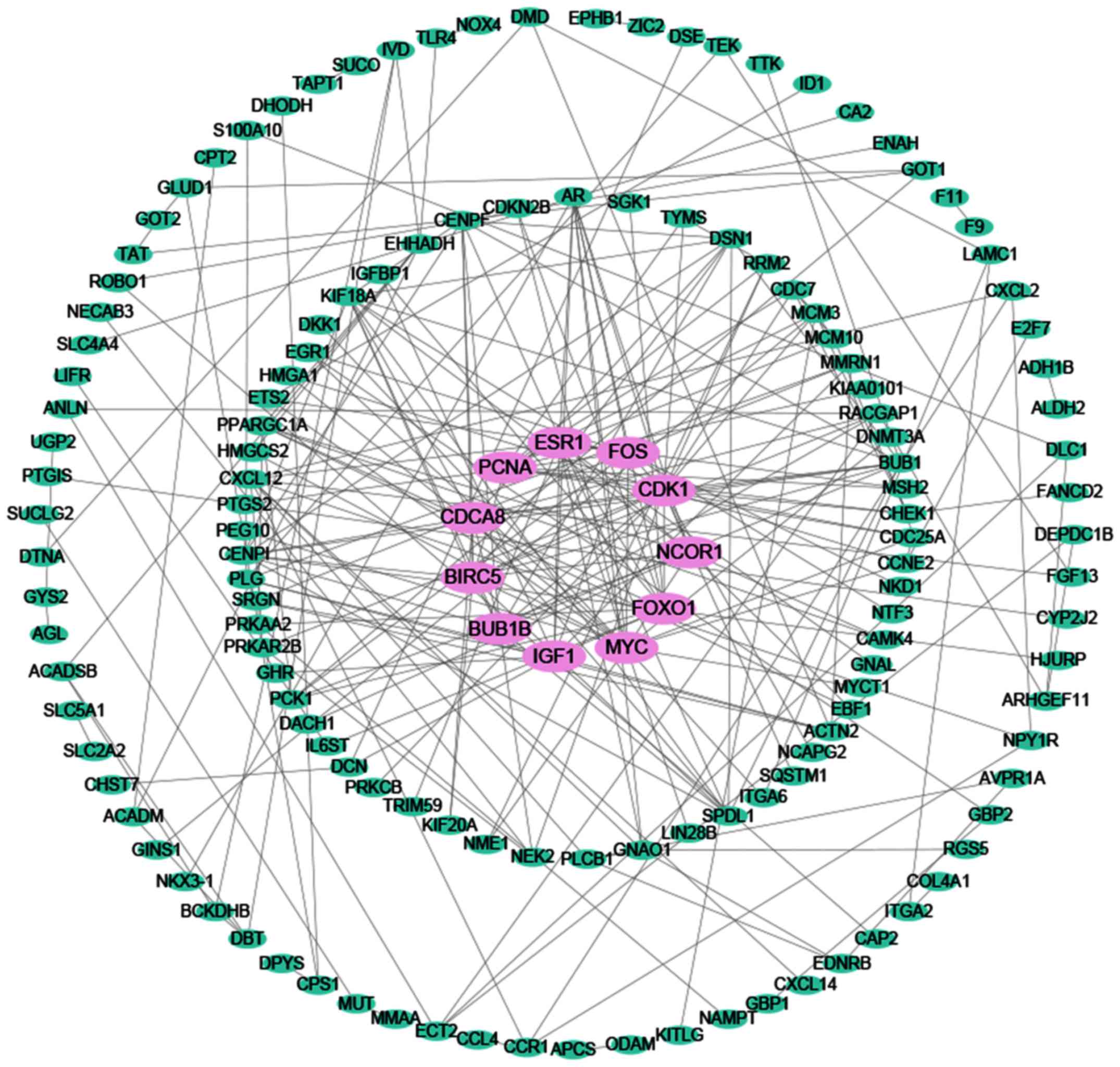
(Fig. 12). The PPI network shown
in Fig. 13 illustrated the
interactions between the target genes of miR-338-5p. There were 147
nodes and 248 edges in the network. Hub genes with a degree values
of more than 11 including NCOR1, IGF1, FOXO1, FOS, CDCA8, BUB1B,
PCNA, ESR1, BIRC5, MYC and CDK1 were emphasized in red while the
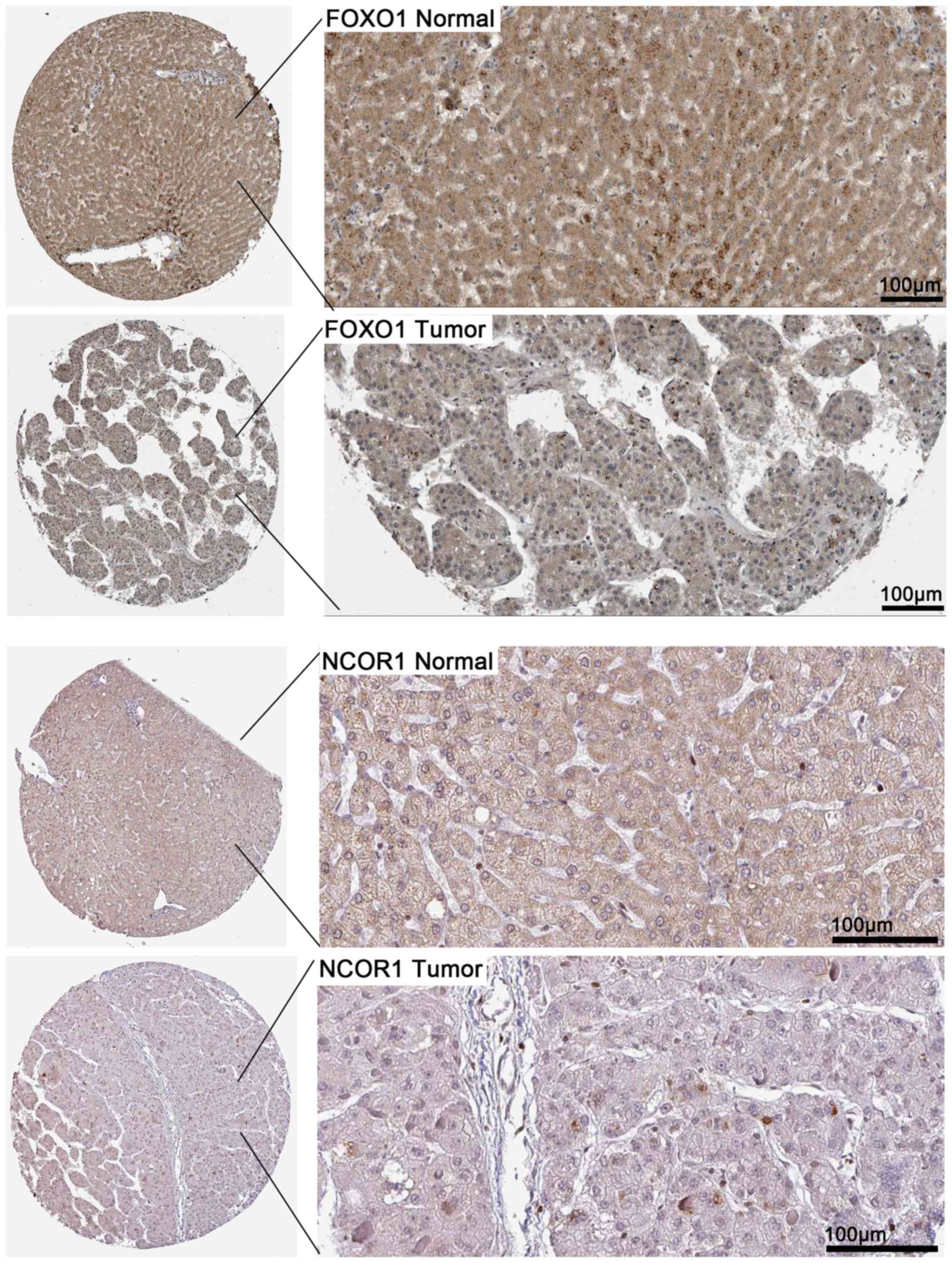
remaining were colored in green. To verify that these hub genes are
targeted by miR-338-5p, we obtained the immunohistochemical
staining of several of the hub genes including NCOR1 and FOXO1 in
HCC tissues and normal tissues. As shown in Fig. 14, NCOR1 and FOXO1 were found to
have medium staining and moderate intensity in
cytoplasmic/menbranous of normal tissues, while a lower staining
and weaker intensity of these genes were observed in HCC
tissues.
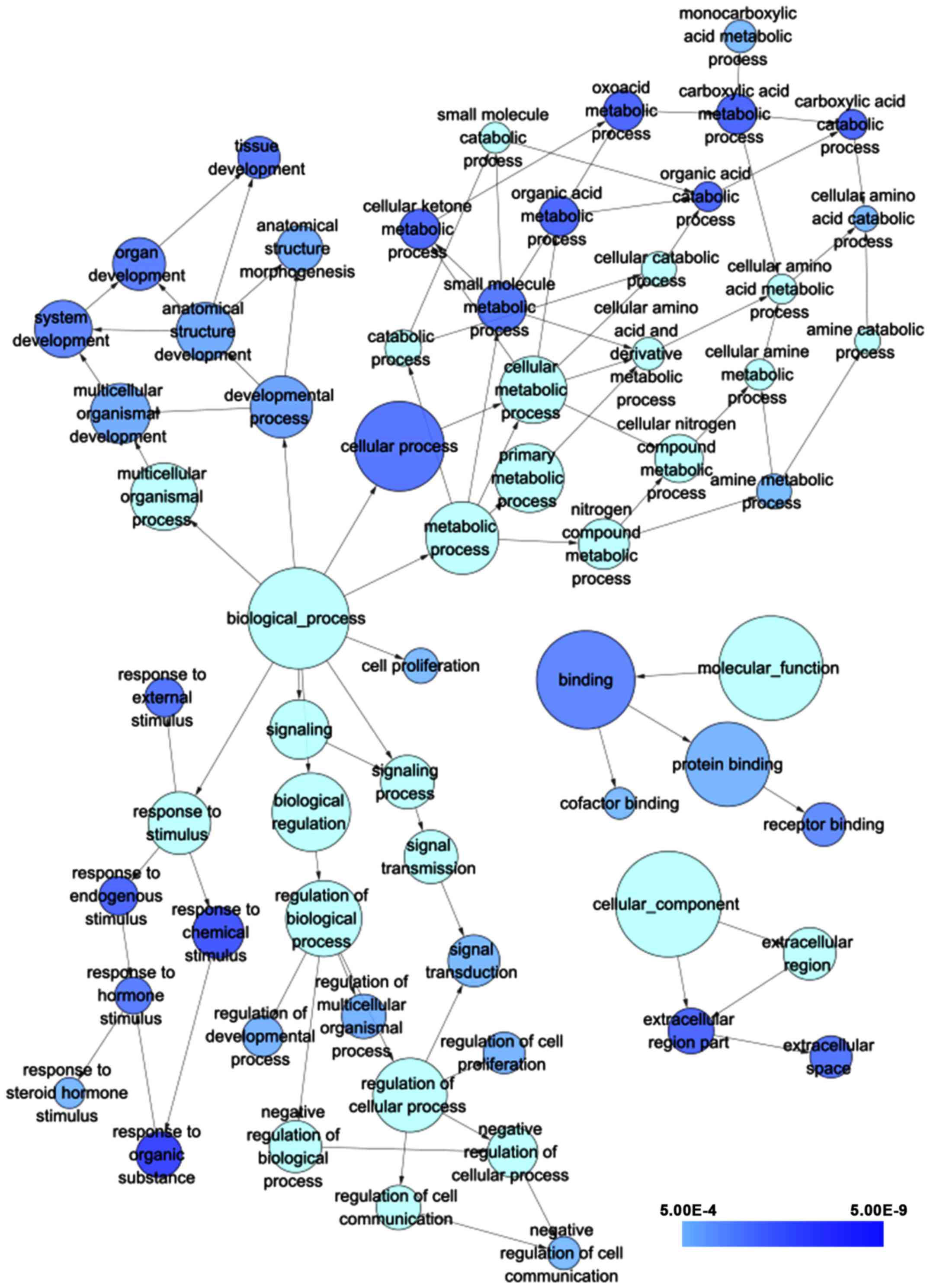
According to the results of GO analysis in
cytoskape, the target genes were found to enrich most significantly
in the following biological pathways: response to organic
substance, response to chemical stimulus and oxoacid metabolic
process. As for cellular component and molecular function, target
genes mainly assembled in extracellular region part and binding,
respectively (Table II; Fig. 15). Moreover, a total of 5
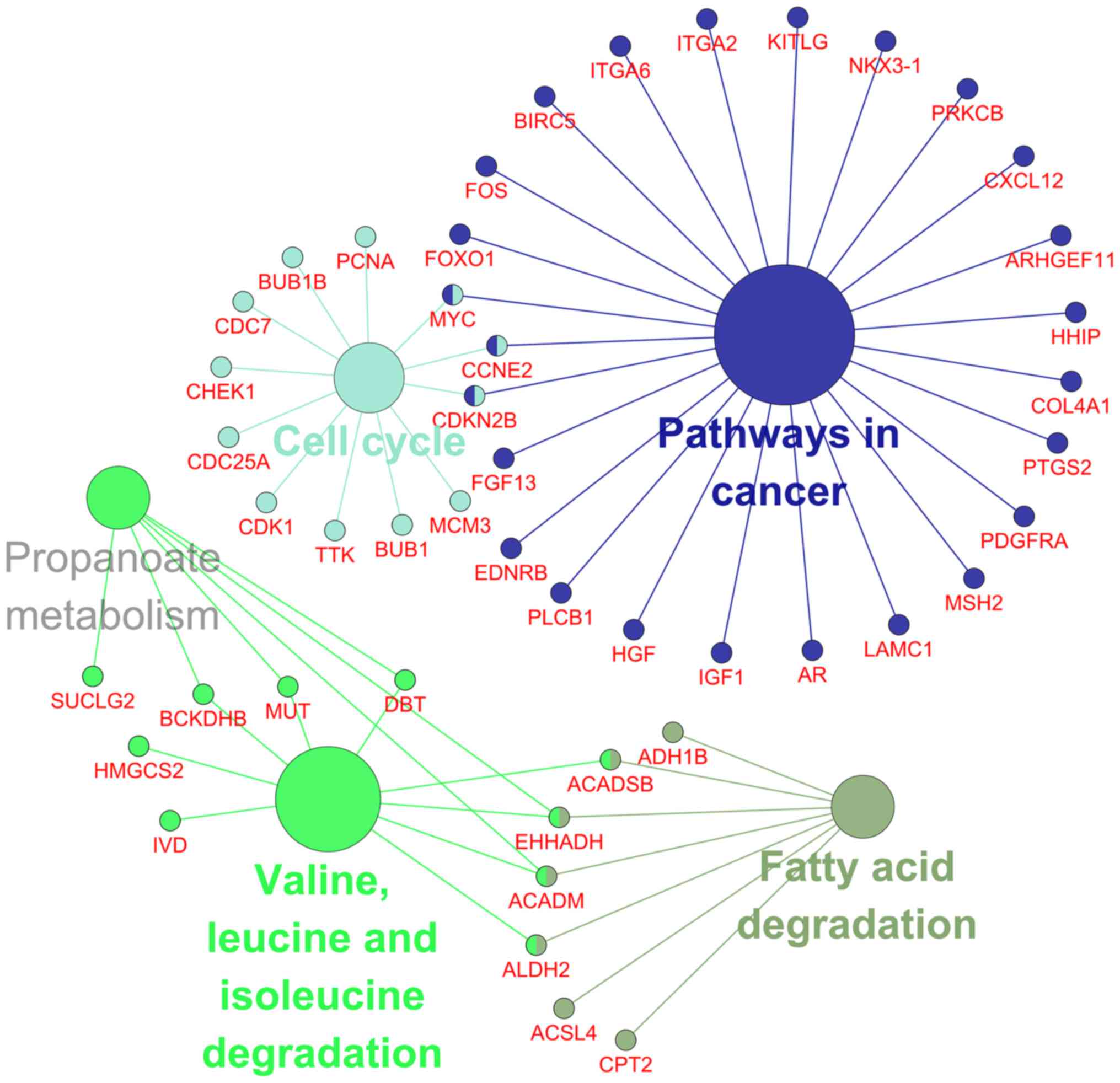
significant pathways were recorded from the KEGG pathway analysis
such as valine, leucine and isoleucine degradation, pathways in
cancer and cell cycle (Table
III; Fig. 16) were the most
significant.
| Table II.GO functional annotation of the
target genes of miR-338-5p from Cytoskape. |
Table II.
GO functional annotation of the
target genes of miR-338-5p from Cytoskape.
| ID | Category | GO term | P-value | Count |
|---|
| GO:0006082 | GO_Biological
process | Organic acid
metabolic process |
3.87×10−9 | 37 |
| GO:0016054 | GO_Biological
process | Organic acid
catabolic process |
6.26×10−9 | 16 |
| GO:0008152 | GO_Biological
process | Metabolic
process |
1.29×10−3 | 159 |
| GO:0009056 | GO_Biological
process | Catabolic
process |
1.15×10−1 | 28 |
| GO:0007275 | GO_Biological
process | Multicellular
organismal development |
1.11×10−6 | 102 |
| GO:0048731 | GO_Biological
process | System
development |
9.71×10−8 | 91 |
| GO:0048519 | GO_Biological
process | Negative regulation
of biological process |
1.82×10−5 | 72 |
| GO:0048523 | GO_Biological
process | Negative regulation
of cellular process |
3.81×10−5 | 66 |
| GO:0050789 | GO_Biological
process | Regulation of
biological process |
3.62×10−4 | 176 |
| GO:0065007 | GO_Biological
process | Biological
regulation |
6.72×10−5 | 189 |
| GO:0008150 | GO_Biological
process |
Biological_process |
1.15×10−3 | 336 |
| GO:0032501 | GO_Biological
process | Multicellular
organismal process |
1.15×10−5 | 133 |
| GO:0044237 | GO_Biological
process | Cellular metabolic
process |
6.22×10−3 | 132 |
| GO:0034641 | GO_Biological
process | Cellular nitrogen
compound metabolic process |
2.19×10−2 | 59 |
| GO:0044238 | GO_Biological
process | Primary metabolic
process |
4.35×10−3 | 140 |
| GO:0050896 | GO_Biological
process | Response to
stimulus |
4.59×10−5 | 112 |
| GO:0009719 | GO_Biological
process | Response to
endogenous stimulus |
3.77×10−9 | 34 |
| GO:0050794 | GO_Biological
process | Regulation of
cellular process |
9.14×10−4 | 166 |
| GO:0023052 | GO_Biological
process | Signaling |
5.63×10−5 | 99 |
| GO:0044281 | GO_Biological
process | Small molecule
metabolic process |
3.29×10−8 | 62 |
| GO:0006519 | GO_Biological
process | Cellular amino acid
and derivative metabolic process |
5.92×10−5 | 21 |
| GO:0043436 | GO_Biological
process | Oxoacid metabolic
process |
2.77×10−9 | 37 |
| GO:0019752 | GO_Biological
process | Carboxylic acid
metabolic process |
2.77×10−9 | 37 |
| GO:0009987 | GO_Biological
process | Cellular
process |
2.40×10−8 | 258 |
| GO:0046395 | GO_Biological
process | Carboxylic acid
catabolic process |
6.26×10−9 | 16 |
| GO:0009063 | GO_Biological
process | Cellular amino acid
catabolic process |
2.78×10−6 | 10 |
| GO:0044248 | GO_Biological
process | Cellular catabolic
process |
3.54×10−2 | 25 |
| GO:0009725 | GO_Biological
process | Response to hormone
stimulus |
4.39×10−8 | 30 |
| GO:0048545 | GO_Biological
process | Response to steroid
hormone stimulus |
2.46×10−6 | 18 |
| GO:0006807 | GO_Biological
process | Nitrogen compound
metabolic process |
2.95×10−3 | 67 |
| GO:0006520 | GO_Biological
process | Cellular amino acid
metabolic process |
4.99×10−5 | 16 |
| GO:0009605 | GO_Biological
process | Response to
external stimulus |
1.81×10−8 | 35 |
| GO:0009308 | GO_Biological
process | Amine metabolic
process |
4.15×10−6 | 25 |
| GO:0009310 | GO_Biological
process | Amine catabolic
process |
9.50×10−6 | 10 |
| GO:0032502 | GO_Biological
process | Developmental
process |
5.43×10−7 | 110 |
| GO:0048856 | GO_Biological
process | Anatomical
structure development |
8.77×10−7 | 94 |
| GO:0009653 | GO_Biological
process | Anatomical
structure morphogenesis |
1.19×10−6 | 53 |
| GO:0042221 | GO_Biological
process | Response to
chemical stimulus |
3.51×10−10 | 70 |
| GO:0010033 | GO_Biological
process | Response to organic
substance |
4.09×10−11 | 52 |
| GO:0010646 | GO_Biological
process | Regulation of cell
communication |
1.30×10−5 | 48 |
| GO:0008283 | GO_Biological
process | Cell
proliferation |
3.68×10−6 | 26 |
| GO:0044282 | GO_Biological
process | Small molecule
catabolic process |
2.35×10−5 | 18 |
| GO:0023046 | GO_Biological
process | Signaling
process |
8.53×10−6 | 77 |
| GO:0023060 | GO_Biological
process | Signal
transmission |
8.53×10−6 | 77 |
| GO:0042127 | GO_Biological
process | Regulation of cell
proliferation |
1.66×10−6 | 41 |
| GO:0048513 | GO_Biological
process | Organ
development |
5.37×10−8 | 74 |
| GO:0009888 | GO_Biological
process | Tissue
development |
2.37×10−8 | 42 |
| GO:0044106 | GO_Biological
process | Cellular amine
metabolic process |
1.70×10−4 | 18 |
| GO:0032787 | GO_Biological
process | Monocarboxylic acid
metabolic process |
3.81×10−6 | 21 |
| GO:0007165 | GO_Biological
process | Signal
transduction |
2.88×10−6 | 71 |
| GO:0010648 | GO_Biological
process | Negative regulation
of cell communication |
3.44×10−6 | 22 |
| GO:0051239 | GO_Biological
process | Regulation of
multicellular organismal process |
1.60×10−6 | 48 |
| GO:0042180 | GO_Biological
process | Cellular ketone
metabolic process |
5.36×10−9 | 37 |
| GO:0050793 | GO_Biological
process | Regulation of
developmental process |
1.88×10−6 | 39 |
| GO:0005515 | GO_Molecular
function | Protein
binding |
2.06×10−6 | 223 |
| GO:0003674 | GO_Molecular
function |
Molecular_function |
4.03×10−4 | 359 |
| GO:0005488 | GO_Molecular
function | Binding |
8.63×10−8 | 316 |
| GO:0048037 | GO_Molecular
function | Cofactor
binding |
3.31×10−6 | 19 |
| GO:0005102 | GO_Molecular
function | Receptor
binding |
1.45×10−7 | 46 |
| GO:0005575 | GO_Cellular
component |
Cellular_component |
1.14×10−2 | 370 |
| GO:0005615 | GO_Cellular
component | Extracellular
space |
1.96×10−8 | 42 |
| GO:0005576 | GO_Cellular
component | Extracellular
region |
1.10×10−5 | 73 |
| GO:0044421 | GO_Cellular
component | Extracellular
region part |
3.08×10−9 | 52 |
| Table III.KEGG pathway analysis of the target
genes of miR-338-5p from Cytoskape. |
Table III.
KEGG pathway analysis of the target
genes of miR-338-5p from Cytoskape.
| ID | Name | Category | Term P-value | Count |
|---|
| GO:0000280 | Valine, leucine and
isoleucine degradation | KEGG |
5.1×10−6 | 9 |
| GO:0005200 | Pathways in
cancer | KEGG |
9.5×10−5 | 25 |
| GO:0004110 | Cell cycle | KEGG |
1.4×10−4 | 12 |
| GO:0000071 | Fatty acid
degradation | KEGG |
1.7×10−4 | 7 |
| GO:0000640 | Propanoate
metabolism | KEGG |
2.0×10−4 | 6 |
Discussion
Considerable attention has been attracted to miRNAs
as promising diagnostic targets for the early screening of human
cancers. Prior to our study, several researches have reported some
miRNAs had diagnostic value in HCC. A 3-miRNA panel: miR-92-3p,
miR-107, and miR-3126-5p discovered by Zhang et al were
claimed to distinguish HCC patients in early stage and HCC patients
with low-level AFP from their corresponding controls with high
accuracy (39). Additionally, some
single miRNAs including miR-21 and miR-224 also exhibited prominent
diagnostic potential for HCC (40,41)
and so far, only one study referred to the diagnostic value of
miR-338-5p in HCC with the method of miRNA array. Chen et al
(21) reported a moderate ability
of miR-338-5p to differentiate HCC from liver cirrhosis with the
AUC of 0.799. Furthermore, an extremely strong diagnostic value of
miR-338-5p (AUC=0.909) was observed when diagnosing HCC from
healthy controls. Despite some advances has been made in exploring
the diagnostic capacity of miRNAs for HCC, the diagnostic
significance of miR-338-5p in HCC was indefinite, and the relative
molecular mechanism has not been elucidated in these studies;
therefore our study was the first one to comprehensively assess the
diagnostic value of miR-338-5p in HCC with the data from GEO, TCGA
and literature as well as to investigate the underlying molecular
mechanism through bioinformatics study.
From the meta-analysis result from our collected
literature and the integrated meta-analysis, miR-338-5p may serve
as a possible diagnostic target for HCC with fair sensitivity and
specificity, which enlightened us that miR-338-5p might play an
essential role in the occurrence and progression. Previous studies
have pointed out that miR-338-5p exerted a tumor suppressive
function in a wide range of cancers. In glioblastoma, miR-338-5p
was discovered to inhibit the proliferation, invasion and promote
apoptosis by targeting EFEMP1 (42); similarly, miR-338-5p significantly
attenuated the malignant potential of gastric cancer cells through
regulating BMI1 (13). In
contrast, miR-338-5p was increased in both blood and tissue of
coloreactal cancer (CRC), and represented high area under ROC curve
(AUC) of 0.871. The performance of miR-338-5p indicated that it
could be a potential biomarker in CRC (19). A similar trend was observed in CRC
compared with HCC. Furthermore, according to the subgroup analysis
in the integrated meta-analysis, studies with samples from plasma
and the method of qRT-PCR were more precise in diagnosing HCC than
studies with the controlled conditions, which hinted that the
sample type and experiment type may also influence the accuracy of
the diagnosis. Although the overall diagnostic ability of
miR-338-5p in HCC was the same, which was reflected by the
integrated meta-analysis and the meta-analysis from our literature
there were still some differences between them. The sensitivity,
specificity, diagnostic odds ratio and the area under SROC of the
result from the literature meta-analysis were higher than those
from the integrated meta-analysis, especially in the evaluation of
sensitivity; miR-338-5p showed a poor sensitivity of only 0.51 in
the integrated meta-analysis. This might be attributed to the
difference in sample type and experiment type as well as the
sources of the data. The integrated meta-analysis included GSE
datasets from different platforms and TCGA data based on the
literature meta-analysis, the samples of which were different.
Moreover, due to the limited number of literature, we failed to
trace the heterogeneity by carrying out subgroup analysis for our
literature meta-analysis. Expanding the sample size was necessary
for a more reliable assessment of the diagnostic value of
miR-338-5p in HCC.
The results from meta-analysis only provided a
superficial hint that miR-338-5p possessed significant diagnostic
capacity in HCC and the molecular mechanism underlying it needed
further exploration. Thus, we emphasized on the network and
functional analysis of the target genes.
We firstly identified the potential target genes of
miR-338-5p and further defined the hub genes from PPI network. The
11 hub genes were assumed to correlate closely with miR-338-5p and
play essential roles in the miR-338-5p relevant pathogenesis of
HCC. Among the hub genes, CDK1 was important protein for the
regulation of cell cycles belonging to the cyclin-dependent kinases
family (43). The overexpression
of CDK1 was detected in various cancers, and a poor prognosis of
renal cell carcinoma patients was associated with the high
expression of CDK1 and CDK2 (44–48).
We hypothesized that CDK1 deregulated by miR-338-5p might promote
the deterioration of HCC by affecting the cell cycle of HCC cells.
Apart from CDK1, several hub genes such as MYC, BIRC5, IGF1, NCOR1
and FOXO1 participate in the regulation of a wide range of
biological processes including cell proliferation, apoptosis and
migration (49–57). These genes were reported to be
aberrantly expressed in various cancers (58–62)
and they were also involved in the malignant progression of HCC
(49,50,52,63).
It was conceived that miR-338-5p might interact with these
molecules through potential signaling pathways to influence the
development of HCC. PCNA, a protein that acted as DNA sliding
clamp, was found to engage in DNA duplication and repair with its
overexpressed in HCC. Further study was necessary to probe into the
association between PCNA and miR-338-5p in HCC. In this study, we
analyzed 11 hub genes protein expression by HPA database. The
result indicated that the expression of FOXO1 and NCOR1 was
downregulated in HCC and most likely regulated by miR-338-5p.
GO enrichment analysis was indicative of the
possible functions of the target genes in HCC and the results from
three GO terms hinted that the target genes were mainly assembled
in response to organic substance. Most of the potential functions
of the target genes from the GO analysis were accomplished in
signaling pathways. Therefore, it is of great importance to
investigate the signaling pathways gathered by the target genes of
miR-338-5p. From the results of the KEGG pathway analysis, the most
significant ten pathways such as pathways in cancer and cell cycle
were closely associated with cancer.
Despite the valuable findings acquired from the
meta-analysis and bioinformatics study, there were still some
limitations in our study. The sample size of our literature was too
small for further analysis to identify heterogeneity, which
weakened the reliability of our results. Since the samples are from
different types, including tissue and plasma, a bias and
sensitivity problems might originate from sample types in analysis.
Additionally, we only included studies published in Chinese or
English, which might cause bias of selection to the meta-analysis.
A plausible way to address these issues is to conduct future
studies with larger samples and fewer language restrictions to
further verify the diagnostic value of miR-338-5p for HCC.
In conclusion, we anticipated that miR-338-5p may
serve as a promising diagnostic marker for HCC and miR-338-5p could
affect the development of HCC by targeting certain downstream genes
and pathways. The future research will be concentrated on
validating the target genes of miR-338-5p and its function in the
significant signaling pathways mentioned before.
References
|
1
|
Nault JC, De Reyniès A, Villanueva A,
Calderaro J, Rebouissou S, Couchy G, Decaens T, Franco D, Imbeaud
S, Rousseau F, et al: A hepatocellular carcinoma 5-gene score
associated with survival of patients after liver resection.
Gastroenterology. 145:176–187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu AX: Molecularly targeted therapy for
advanced hepatocellular carcinoma in 2012: Current status and
future perspectives. Semin Oncol. 39:493–502. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Llovet JM, Villanueva A, Lachenmayer A and
Finn RS: Advances in targeted therapies for hepatocellular
carcinoma in the genomic era. Nat Rev Clin Oncol. 12:408–424. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lages E, Ipas H, Guttin A, Nesr H, Berger
F and Issartel JP: MicroRNAs: Molecular features and role in
cancer. Front Biosci (Landmark Ed). 17:2508–2540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Neilson JR and Sharp PA: Small RNA
regulators of gene expression. Cell. 134:899–902. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:pp. 2257–2261.
2006; View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee EJ, Gusev Y, Jiang J, Nuovo GJ, Lerner
MR, Frankel WL, Morgan DL, Postier RG, Brackett DJ and Schmittgen
TD: Expression profiling identifies microRNA signature in
pancreatic cancer. Int J Cancer. 120:1046–1054. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yanaihara N, Caplen N, Bowman E, Seike M,
Kumamoto K, Yi M, Stephens RM, Okamoto A, Yokota J, Tanaka T, et
al: Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Murakami Y, Yasuda T, Saigo K, Urashima T,
Toyoda H, Okanoue T and Shimotohno K: Comprehensive analysis of
microRNA expression patterns in hepatocellular carcinoma and
non-tumorous tissues. Oncogene. 25:2537–2545. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Calin GA, Liu CG, Sevignani C, Ferracin M,
Felli N, Dumitru CD, Shimizu M, Cimmino A, Zupo S, Dono M, et al:
MicroRNA profiling reveals distinct signatures in B cell chronic
lymphocytic leukemias. Proc Natl Acad Sci USA. 101:pp. 11755–11760.
2004; View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tong D, Zhao L, He K, Sun H, Cai D, Ni L,
Sun R, Chang S, Song T, Huang C, et al: MECP2 promotes the growth
of gastric cancer cells by suppressing miR-338-mediated
antiproliferative effect. Oncotarget. 7:34845–34859. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen JS, Liang LL, Xu HX, Chen F, Shen SL,
Chen W, Chen LZ, Su Q, Zhang LJ, Bi J, et al: miR-338-3p inhibits
epithelial-mesenchymal transition and metastasis in hepatocellular
carcinoma cells. Oncotarget. 8:71418–71429. 2016.PubMed/NCBI
|
|
15
|
Chen X, Wei L and Zhao S: miR-338 inhibits
the metastasis of lung cancer by targeting integrin β3. Oncol Rep.
36:1467–1474. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Weng HL and Wang MJ: Effects of
microRNA-338-3p on morphine-induced apoptosis and its underlying
mechanisms. Mol Med Rep. 14:2085–2092. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhuang Y, Dai J and Wang Y, Zhang H, Li X,
Wang C, Cao M, Liu Y, Cai H, Zhang D and Wang Y: miR-338*
suppresses fibrotic pathogenesis in pulmonary fibrosis through
targeting LPA1. Am J Transl Res. 8:3197–3205. 2016.PubMed/NCBI
|
|
18
|
Xing Z, Yu L, Li X and Su X: Anticancer
bioactive peptide-3 inhibits human gastric cancer growth by
targeting miR-338-5p. Cell Biosci. 6:532016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yong FL, Law CW and Wang CW: Potentiality
of a triple microRNA classifier: miR-193a-3p, miR-23a and
miR-338-5p for early detection of colorectal cancer. BMC Cancer.
13:2802013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Besse A, Sana J, Lakomy R, Kren L, Fadrus
P, Smrcka M, Hermanova M, Jancalek R, Reguli S, Lipina R, et al:
miR-338-5p sensitizes glioblastoma cells to radiation through
regulation of genes involved in DNA damage response. Tumour Biol.
37:7719–7727. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Y, Chen J, Liu Y, Li S and Huang P:
Plasma miR-15b-5p, miR-338-5p, and miR-764 as biomarkers for
hepatocellular carcinoma. Med Sci Monit. 21:1864–1871. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Deeks JJ, Macaskill P and Irwig L: The
performance of tests of publication bias and other sample size
effects in systematic reviews of diagnostic test accuracy was
assessed. J Clin Epidemiol. 58:882–893. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Glas AS, Lijmer JG, Prins MH, Bonsel GJ
and Bossuyt PM: The diagnostic odds ratio: A single indicator of
test performance. J Clin Epidemiol. 56:1129–1135. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Harbord RM, Deeks JJ, Egger M, Whiting P
and Sterne JA: A unification of models for meta-analysis of
diagnostic accuracy studies. Biostatistics. 8:239–251. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Higgins JP, Thompson SG, Deeks JJ and
Altman DG: Measuring inconsistency in meta-analyses. BMJ.
327:557–560. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jackson D, White IR and Thompson SG:
Extending DerSimonian and Laird's methodology to perform
multivariate random effects meta-analyses. Stat Med. 29:1282–1297.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Uhlen M, Oksvold P, Fagerberg L, Lundberg
E, Jonasson K, Forsberg M, Zwahlen M, Kampf C, Wester K, Hober S,
et al: Towards a knowledge-based Human protein atlas. Nat
Biotechnol. 28:1248–1250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen Y, Huang P, Chen J, Liu Y, Li SL,
Wang Z and Song D: Plasma circulating miR-338-5p, miR-21-5p and
miR-15b-5p are potential biomarkers for screening hepatocellular
carcinoma. J Third Mil Med Univ. 37:1720–1726. 2015.(In
Chinese).
|
|
29
|
Budhu A, Jia HL, Forgues M, Liu CG,
Goldstein D, Lam A, Zanetti KA, Ye QH, Qin LX, Croce CM, et al:
Identification of metastasis-related microRNAs in hepatocellular
carcinoma. Hepatology. 47:897–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Su H, Yang JR, Xu T, Huang J, Xu L, Yuan Y
and Zhuang SM: MicroRNA-101, down-regulated in hepatocellular
carcinoma, promotes apoptosis and suppresses tumorigenicity. Cancer
Res. 69:1135–1142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Burchard J, Zhang C, Liu AM, Poon RT, Lee
NP, Wong KF, Sham PC, Lam BY, Ferguson MD, Tokiwa G, et al:
microRNA-122 as a regulator of mitochondrial metabolic gene network
in hepatocellular carcinoma. Mol Syst Biol. 6:4022010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Diaz G, Melis M, Tice A, Kleiner DE,
Mishra L, Zamboni F and Farci P: Identification of microRNAs
specifically expressed in hepatitis C virus-associated
hepatocellular carcinoma. Int J Cancer. 133:816–824. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen J, Wang A, Wang Q, Gurvich I, Siegel
AB, Remotti H and Santella RM: Exploration of genome-wide
circulating microRNA in hepatocellular carcinoma: miR-483-5p as a
potential biomarker. Cancer Epidemiol Biomarkers Prev.
22:2364–2373. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Murakami Y, Kubo S, Tamori A, Itami S,
Kawamura E, Iwaisako K, Ikeda K, Kawada N, Ochiya T and Taguchi YH:
Comprehensive analysis of transcriptome and metabolome analysis in
Intrahepatic cholangiocarcinoma and hepatocellular carcinoma. Sci
Rep. 5:162942015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Martinez-Quetglas I, Pinyol R, Dauch D,
Torrecilla S, Tovar V, Moeini A, Alsinet C, Portela A,
Rodriguez-Carunchio L, Solé M, et al: IGF2 is up-regulated by
epigenetic mechanisms in hepatocellular carcinomas and is an
actionable oncogene product in experimental models.
Gastroenterology. 151:1192–1205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shih TC, Tien YJ, Wen CJ, Yeh TS, Yu MC,
Huang CH, Lee YS, Yen TC and Hsieh SY: MicroRNA-214 downregulation
contributes to tumor angiogenesis by inducing secretion of the
hepatoma-derived growth factor in human hepatoma. J Hepatol.
57:584–591. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Noh JH, Chang YG, Kim MG, Jung KH, Kim JK,
Bae HJ, Eun JW, Shen Q, Kim SJ, Kwon SH, et al: miR-145 functions
as a tumor suppressor by directly targeting histone deacetylase 2
in liver cancer. Cancer Lett. 335:455–462. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sato F, Hatano E, Kitamura K, Myomoto A,
Fujiwara T, Takizawa S, Tsuchiya S, Tsujimoto G, Uemoto S and
Shimizu K: MicroRNA profile predicts recurrence after resection in
patients with hepatocellular carcinoma within the Milan Criteria.
PLoS One. 6:e164352011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Y, Li T, Qiu Y, Zhang T, Guo P, Ma
X, Wei Q and Han L: Serum microRNA panel for early diagnosis of the
onset of hepatocellular carcinoma. Medicine (Baltimore).
96:e56422017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Okajima W, Komatsu S, Ichikawa D, Miyamae
M, Kawaguchi T, Hirajima S, Ohashi T, Imamura T, Kiuchi J, Arita T,
et al: Circulating microRNA profiles in plasma: Identification of
miR-224 as a novel diagnostic biomarker in hepatocellular carcinoma
independent of hepatic function. Oncotarget. 7:53820–53836. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yan SR, Liu ZJ, Yu S and Bao YX:
Investigation of the value of miR-21 in the diagnosis of early
stage HCC and its prognosis: A meta-analysis. Genet Mol Res.
14:11573–11586. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lei D, Zhang F, Yao D, Xiong N, Jiang X
and Zhao H: miR-338-5p suppresses proliferation, migration,
invasion, and promote apoptosis of glioblastoma cells by directly
targeting EFEMP1. Biomed Pharmacother. 89:957–965. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang W, Cho H, Shin HY, Chung JY, Kang ES,
Lee EJ and Kim JH: Accumulation of cytoplasmic Cdk1 is associated
with cancer growth and survival rate in epithelial ovarian cancer.
Oncotarget. 7:49481–49497. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tsaur I, Makarević J, Hudak L, Juengel E,
Kurosch M, Wiesner C, Bartsch G, Harder S, Haferkamp A and Blaheta
RA: The cdk1-cyclin B complex is involved in everolimus triggered
resistance in the PC3 prostate cancer cell line. Cancer Lett.
313:84–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sung WW, Lin YM, Wu PR, Yen HH, Lai HW, Su
TC, Huang RH, Wen CK, Chen CY, Chen CJ and Yeh KT: High
nuclear/cytoplasmic ratio of Cdk1 expression predicts poor
prognosis in colorectal cancer patients. BMC Cancer. 14:9512014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Willder JM, Heng SJ, McCall P, Adams CE,
Tannahill C, Fyffe G, Seywright M, Horgan PG, Leung HY, Underwood
MA and Edwards J: Androgen receptor phosphorylation at serine 515
by Cdk1 predicts biochemical relapse in prostate cancer patients.
Br J Cancer. 108:139–148. 2013.PubMed/NCBI
|
|
47
|
Banerjee SK, Weston AP, Zoubine MN,
Campbell DR and Cherian R: Expression of cdc2 and cyclin B1 in
Helicobacter pylori-associated gastric MALT and MALT lymphoma:
Relationship to cell death, proliferation, and transformation. Am J
Pathol. 156:217–225. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hongo F, Takaha N, Oishi M, Ueda T,
Nakamura T, Naitoh Y, Naya Y, Kamoi K, Okihara K, Matsushima T, et
al: CDK1 and CDK2 activity is a strong predictor of renal cell
carcinoma recurrence. Urol Oncol. 32:1240–1246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tian J, Hu X, Gao W, Zhang J, Chen M,
Zhang X, Ma J and Yuan H: Identification a novel tumor-suppressive
hsa-miR-599 regulates cells proliferation, migration and invasion
by targeting oncogenic MYC in hepatocellular carcinoma. Am J Transl
Res. 8:2575–2584. 2016.PubMed/NCBI
|
|
50
|
Cao L, Li C, Shen S, Yan Y, Ji W, Wang J,
Qian H, Jiang X, Li Z, Wu M, et al: OCT4 increases BIRC5 and CCND1
expression and promotes cancer progression in hepatocellular
carcinoma. BMC Cancer. 13:822013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Le Coz V, Zhu C, Devocelle A, Vazquez A,
Boucheix C, Azzi S, Gallerne C, Eid P, Lecourt S and Giron-Michel
J: IGF-1 contributes to the expansion of melanoma-initiating cells
through an epithelial-mesenchymal transition process. Oncotarget.
7:82511–82527. 2016.PubMed/NCBI
|
|
52
|
Yang XW, Shen GZ, Cao LQ, Jiang XF, Peng
HP, Shen G, Chen D and Xue P: MicroRNA-1269 promotes proliferation
in human hepatocellular carcinoma via downregulation of FOXO1. BMC
Cancer. 14:9092014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ko YS, Cho SJ, Park J, Kim Y, Choi YJ, Pyo
JS, Jang BG, Park JW, Kim WH and Lee BL: Loss of FOXO1 promotes
gastric tumour growth and metastasis through upregulation of human
epidermal growth factor receptor 2/neu expression. Br J Cancer.
113:1186–1196. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ren JW, Li ZJ and Tu C: miR-135
post-transcriptionally regulates FOXO1 expression and promotes cell
proliferation in human malignant melanoma cells. Int J Clin Exp
Pathol. 8:6356–6366. 2015.PubMed/NCBI
|
|
55
|
Martinez-Iglesias OA, Alonso-Merino E,
Gómez-Rey S, Velasco-Martín JP, Martín Orozco R, Luengo E, García
Martín R, Ibáñez de Cáceres I, Fernández AF, Fraga MF, et al:
Autoregulatory loop of nuclear corepressor 1 expression controls
invasion, tumor growth, and metastasis. Proc Natl Acad Sci USA.
113:pp. E328–E337. 2016; View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yu JJ, Wu YX, Zhao FJ and Xia SJ: miR-96
promotes cell proliferation and clonogenicity by down-regulating of
FOXO1 in prostate cancer cells. Med Oncol. 31:9102014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang B, Gui LS, Zhao XL, Zhu LL and Li
QW: FOXO1 is a tumor suppressor in cervical cancer. Genet Mol Res.
14:6605–6616. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Li K, Chen MK, Situ J, Huang WT, Su ZL, He
D and Gao X: Role of co-expression of c-Myc, EZH2 and p27 in
prognosis of prostate cancer patients after surgery. Chin Med J
(Engl). 126:82–87. 2013.PubMed/NCBI
|
|
59
|
Liu Z, Jiang Y, Hou Y, Hu Y, Cao X, Tao Y,
Xu C, Liu S, Wang S, Wang L, et al: The IκB family member Bcl-3
stabilizes c-Myc in colorectal cancer. J Mol Cell Biol. 5:280–282.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Duffy MJ, O'Donovan N, Brennan DJ,
Gallagher WM and Ryan BM: Survivin: A promising tumor biomarker.
Cancer Lett. 249:49–60. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Grimberg A: Mechanisms by which IGF-I may
promote cancer. Cancer Biol Ther. 2:630–635. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xie L, Ushmorov A, Leithäuser F, Guan H,
Steidl C, Färbinger J, Pelzer C, Vogel MJ, Maier HJ, Gascoyne RD,
et al: FOXO1 is a tumor suppressor in classical Hodgkin lymphoma.
Blood. 119:3503–3511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chun YS, Huang M, Rink L and Von Mehren M:
Expression levels of insulin-like growth factors and receptors in
hepatocellular carcinoma: A retrospective study. World J Surg
Oncol. 12:2312014. View Article : Google Scholar : PubMed/NCBI
|