Introduction
Articular cartilage injury can trigger joint pain
and dysfunction. Cartilage exhibits a poor capacity for
self-repair. Cartilage degeneration after trauma or during disease
may readily trigger osteoarthritis (1). Cartilage tissue engineering is
considered to be a promising approach towards cartilage repair.
Various scaffolds promoting cartilage generation may be utilized,
and chondrocyte proliferation and differentiation can be guided by
application of suitable biological stimuli. However, maintenance of
the phenotype of regenerated cartilage, and long-term retention of
normal function, remain challenging.
Icariin is a monomeric compound identified in
extracts of Herba Epimedii; the protein exhibits a cardioprotective
effect, may be used to treat osteoporosis, and has aphrodisiac
qualities (2,3). Icariin promoted B-cell lymphoma 2
(Bcl-2) gene expression and suppressed Bax gene expression, thus
inhibiting ventricular remodeling and myocardial cell apoptosis
(4). Icariin upregulated cyclic
guanosine 3′,5′-monophosphate (cGMP) levels and inhibited
phosphodiesterase type 5, thus promoting nitric oxide (NO) release
(a property of aphrodisiacs) (2).
In the context of osteoporosis, icariin promoted osteogenic
differentiation of rat bone marrow stromal cells (BMSCs) by
activating the phosphoinositide 3-kinase (PI3K)-AKT-endothelial NO
synthase (eNOS)-NO-cGMP-protein kinase G (PKG) signaling axis,
triggering rapid phosphorylation of c-Jun N-terminal kinase (JNK),
p38 kinase, and extracellular signal-regulated kinase (ERK)
(5,6). Li et al (7) demonstrated that an
icariin-impregnated hydrogel scaffold efficiently promoted repair
of supercritically sized osteochondral defects, and enhanced the
integration of regenerated cartilage and subchondral bone in a
rabbit model.
Primary cilia are non-motile microtubular organelles
protruding from the surfaces of most eukaryotic cells. Cilia serve
as ‘antennae’, detecting mechanical stress, and they engage in
biochemical signal transduction from the extracellular environment
(8). Intraflagellar transport 88
(IFT88) is a key ciliary protein that shuttles cargos along the
ciliary axoneme (9,10). In chondrocytes, primary cilia
transmit mechanical stress and chemical signals (11–13).
Primary cilia facilitate secretion of the cartilage matrix in
association with the Golgi apparatus (14). Inhibition of IFT88 triggers ciliary
disassembly and dysplasia of the epiphyseal plate (15). However, any role for IFT88 in
maintenance of the chondrocyte phenotype remains poorly understood.
In the present study, it was hypothesized that icariin might aid in
maintenance of the chondrocyte phenotype by regulating IFT88
expression. Thus, the effects of icariin on IFT88 expression in
progenitor chondrocyte ATDC5 cells and primary chondrocytes were
examined.
Materials and methods
Cells and reagents
The progenitor chondrocytic cell line ATDC5 was
purchased from the American Type Culture Collection (Manassas, VA,
USA). Primary chondrocytes were obtained from the knee cartilage of
6 newborn (3 days old, 6–9 g) Sprague-Dawley (SD) rats of either
sex (3 male and 3 female,). These SD rats were provided by the
Experimental Animal Center (Tongji Hospital, Wuhan, China). All
animals were maintained in the same housing conditions with free
access to food and water (see below ‘Animal experiments’ section).
Cells were cultured in Dulbecco's modified Eagle's medium/nutrient
mixture F-12 (DMEM/F12) supplemented with 10% (v/v) fetal bovine
serum (FBS) and 100 U/ml penicillin/streptomycin. Icariin was
purchased from the Cayman Chemical Company (Ann Arbor, MI, USA).
The ERK inhibitor, PD0325901, was obtained from Selleckchem
(Houston, TX, USA).
Cell viability assay
The effect of icariin on ATDC5 cell proliferation
was evaluated with the aid of a CCK-8 kit (Wuhan Boster Biological
Technology, Ltd., Wuhan, China). Cells (2,000/well) were seeded
into 96-well plates and cultured in 100 µl DMEM/F12 with 10% (v/v)
FBS medium containing different concentrations of icariin (0.001–10
µmol/l). After 48 h, 10 µl amounts of CCK-8 solution were added to
the wells, followed by incubation at 37°C for 90 min. Absorbance at
450 nm was measured using a microplate reader.
Histochemical and immunohistochemical
assays
Chondrocytes (obtained from the aforementioned
newborn rats) and the knee joints of 8-week old SD rats described
below, were subjected to histochemical staining. Cells were fixed
in 4% (v/v) paraformaldehyde for 15 min, and subsequently stained
with 0.5% (v/v) toluidine blue. The knee joints were fixed in 4%
(v/v) paraformaldehyde for 2 days, decalcified for 4 weeks in 10%
(w/v) EDTA, embedded in paraffin, and stained with Safranin O-Fast
Green and toluidine blue, following standard protocols. All
immunohistochemical techniques followed were as described
previously (16). Primary
antibodies were obtained from Proteintech (Wuhan Sanying; Wuhan,
China). Aggrecan expression in tissue sections was evaluated
microscopically and its color intensity of staining was quantified
by use of Image Pro Plus 6.0 (Media Cybernetics, Inc., Rockville,
MD, USA) (17). The osteoarthritis
assessment system of O'Driscoll was used to evaluate the severity
of cartilage degeneration (18).
Immunofluorescence assay
Chondrocytes at appropriate densities were
inoculated on to coverslips. Using standard immunofluorescence
methods, the primary cilia were stained with anti-acetylated
α-tubulin antibody (cat. no. T7451; 1:300 dilution; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) or double-stained with that
antibody together with an antibody against IFT88 (cat. no.
AP11138b; 1:50 dilution; Abgent Inc., San Diego, CA, USA).
CY3-conjugated goat anti-mouse, fluorescein isothiocyanate
(FITC)-conjugated goat anti-rabbit immunoglobulin G (IgG) (cat. no.
BA1031, 1:200 dilution; and cat. no. BA1105, 1:100 dilution;
respectively; both from Wuhan Boster Biological Technology, Ltd.)
served as secondary antibodies, and nuclei were stained with DAPI
(1 µg/µl). Images were captured with a camera fitted to a
fluorescence microscope.
Western blotting
Total cellular lysates were prepared with the aid of
RIPA buffer. Samples of protein (20 µg) were loaded into lanes,
separated on sodium dodecyl sulfate (SDS)-polyacrylamide 10% gels,
and transferred to polyvinylidene difluoride (PVDF) membranes. The
PVDF membranes were incubated with antibodies against IFT88 (cat.
no. AP11138b; 1:200 dilution; Abgent Inc.), ERK and phosphorylated
ERK (cat. nos. 9102 and 4370, respectively; both 1:1,000; both from
Cell Signaling Technology, Inc., Danvers, MA, USA), and GAPDH (cat.
no. BM3876; 1:400 dilution; Wuhan Boster Biological Technology,
Ltd.); and subsequently with horseradish peroxidase (HRP)-labelled
goat anti-mouse or goat anti-rabbit secondary antibody (cat. nos.
BA1050 and BA1054, respectively; both 1:2,000 dilution, both from
Wuhan Boster Biological Technology, Ltd.). Protein bands were
detected using a Bio-Rad enhanced chemiluminescence system
(Bio-Rad, Philadelphia, PA, USA).
siRNA transfection
ATDC5 progenitor chondrocytic cells were transfected
with 100 nM siRNA targeting IFT88 or a negative control siRNA
(Guangzhou RiboBio Co., Ltd., Guangzhou, China), using a standard
protocol. Knockdown efficiency was evaluated by western
blotting.
Quantitative polymerase chain reaction
(qPCR)
Expression levels of genes affecting the chondrocyte
phenotype (COL-II, SOX9, and IFT88) were measured by qPCR. Total
RNA was extracted with TRIzol and cDNAs were synthesized from 2 µg
amounts of total RNA using a Toyobo cDNA synthesis kit (Toyobo,
Co., Ltd., Osaka, Japan), according to the manufacturer's protocol.
Each PCR tube contained 1 µl cDNA, 1 µl primers, 10 µl SYBR-Green
DNA polymerase (Toyobo, Co., Ltd.), and 8 µl RNAse-free water. The
primers sequences are listed in Table
I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene (rat) | Primer sequences
(5′-3′) | Gene (mouse) | Primer sequences
(5′-3′) |
|---|
| COLII | F:
TCCTCCGTCTACTGTCCA | COLII | F:
GCTCCCAACACCGCTAACG |
|
| R:
ACTTACCGGTGTGTTTCG |
| R:
GCCGCTTCGTCCAGGTAGG |
| SOX9 | F:
TCGGGGCTCTACTCCACCT | SOX9 | F:
GAGCCGGATCTGAAGAGGGA |
|
| R:
TCTGTCACCATTGCTCTTC |
| R:
GCTTGACGTGTGGCTTGTTC |
| IFT88 | F:
ACCAGGCTGTAGACACATT | IFT88 | F:
TGGCCAACGACCTGGAGATTAACA |
|
| R:
TTCTCGTAGTCACCATTTG |
| R:
ATAGCTGCTGGCTTGGGCAAATTC |
| GAPDH | F:
CTGCTCCTCCCTGTTCTA | GAPDH | F:
GCCTTCCGTGTTCCTACCC |
|
| R:
CAATGTCCACTTTGTCAC |
| R:
GCCCTCAGATGCCTGCTTC |
Animal experiments
The present study was approved by the Ethics
Committee of Tongji Hospital (Wuhan, China). A total of 24 male SD
rats (8 weeks old, 180–220 g) were divided into four groups
(control, treadmill, icariin, and icariin + treadmill). Animals
were purchased from the Experimental Animal Center, Tongji
Hospital, (Wuhan, China). All these rats were maintained with free
access to food and water, at a constant room temperature of 23±1°C
with a 12 h light/dark cycle, 50–70% humidity and 0.03%
CO2. Full-thickness cartilage defects were created, and
early treadmill exercise was used to accelerate the development of
post-traumatic osteoarthritis (PTOA). All rats underwent 1 week of
treadmill training prior to surgery. Full-thickness cartilage
defects were created by drilling a 1-mm-diameter hole through the
middle of the femoral trochlea of the right knee, as described in a
previous study (19). Rats in the
treadmill and icariin + treadmill groups commenced running
exercises (15 m/min, 30 min/day) 1 week after surgery (19), and were gavage-fed with icariin (25
mg/kg) or saline daily (20).
After 6 weeks, the rats were sacrificed and knee samples were
subjected to histomorphological and immunohistochemical
assessment.
Statistical analysis
All data are reported as the mean ± standard
deviation, and means with 95% confidence intervals were calculated.
Student's t-test or one-way analysis of variance was used to assess
the significance of between-group differences. P<0.05 was
considered to indicate a statistically significant difference.
Results
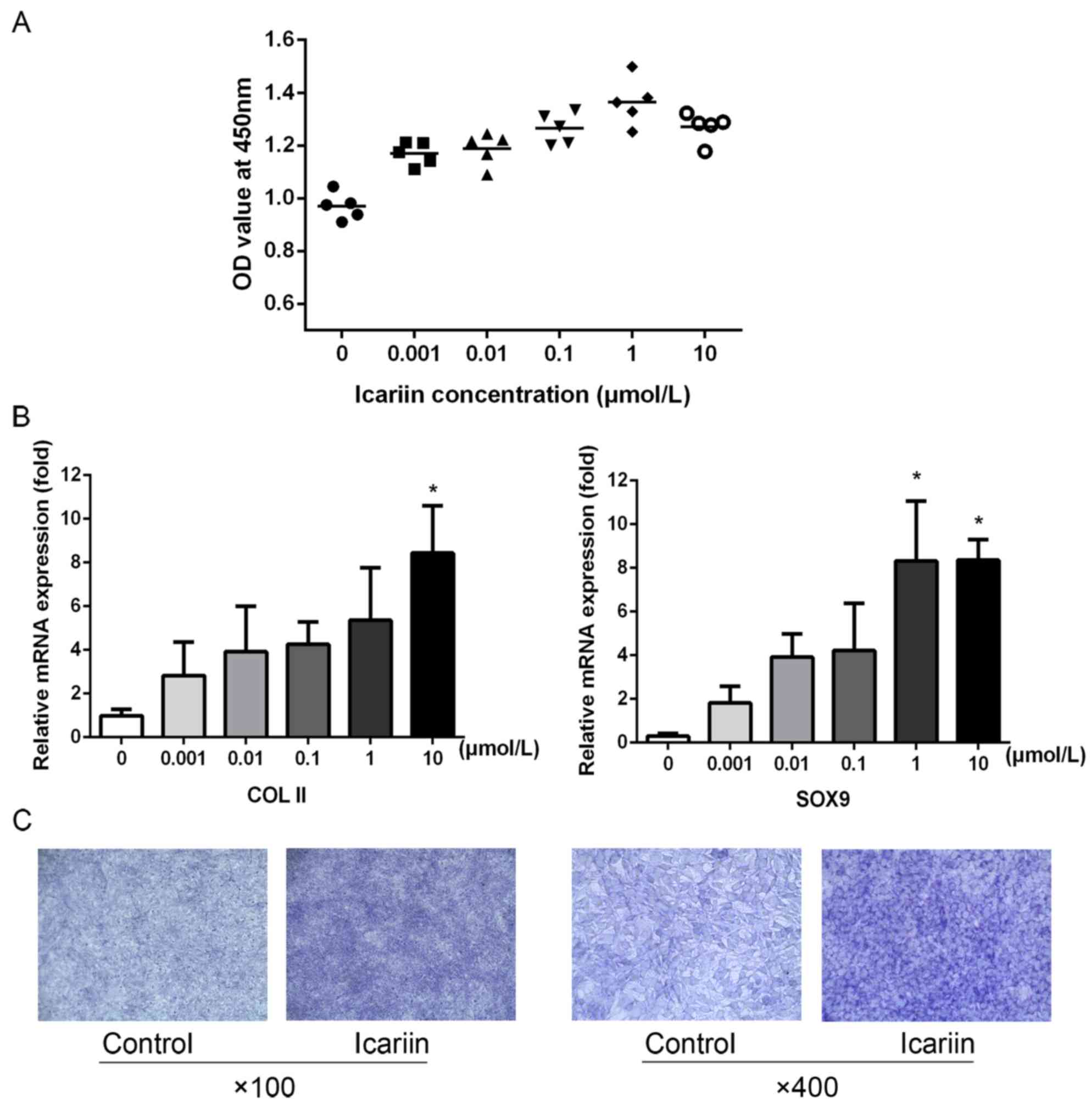
Icariin affects the proliferation and
differentiation of ATDC5 chondrocytic cells
Icariin at different concentrations, added to
growing cells, promoted ATDC5 proliferation in a
concentration-dependent manner, peaking at 1 µmol/l (Fig. 1A). Furthermore, icariin promoted
the expression of the chondrocyte phenotype-associated genes,
COL-II and SOX9, in a concentration-dependent manner, peaking at 10
µmol/l (Fig. 1B). Following
stimulation with 10 µmol/l icariin for 3 days, toluidine blue
staining confirmed that icariin significantly promoted secretion of
cartilage extracellular matrix (ECM) (Fig. 1C).
Icariin promotes ciliary assembly in,
and IFT88 expression by, ATDC5 cells
The primary cilia regulate numerous cellular
activities, particularly the balance between proliferation and
differentiation (21,22). ATDC5 cells treated with 10 µmol/l
icariin were analysed in terms of primary cilia production
(Fig. 2A and B). Icariin-treated
cells expressed a higher percentage of primary cilia (34.06±10.06%
of cells) compared with the control group (27.91±9.95%). The
average ciliary length of icariin-treated cells (3.34±1.34 µm) was
slightly greater compared with that of the control cells (3.07±0.74
µm). IFT88 was expressed at both the gene and the protein level.
RT-qPCR revealed that icariin enhanced the expression of mRNA
encoding IFT88 in a concentration-dependent manner, peaking at 10
µmol/l (Fig. 2C). Icariin promoted
the protein expression of IFT88 in a concentration- and a
time-dependent manner, peaking at 1 or 10 µmol/l. When exposed to
10 µmol/l icariin for different times, the protein expression of
IFT88 peaked at 24 h (Fig.
2D).
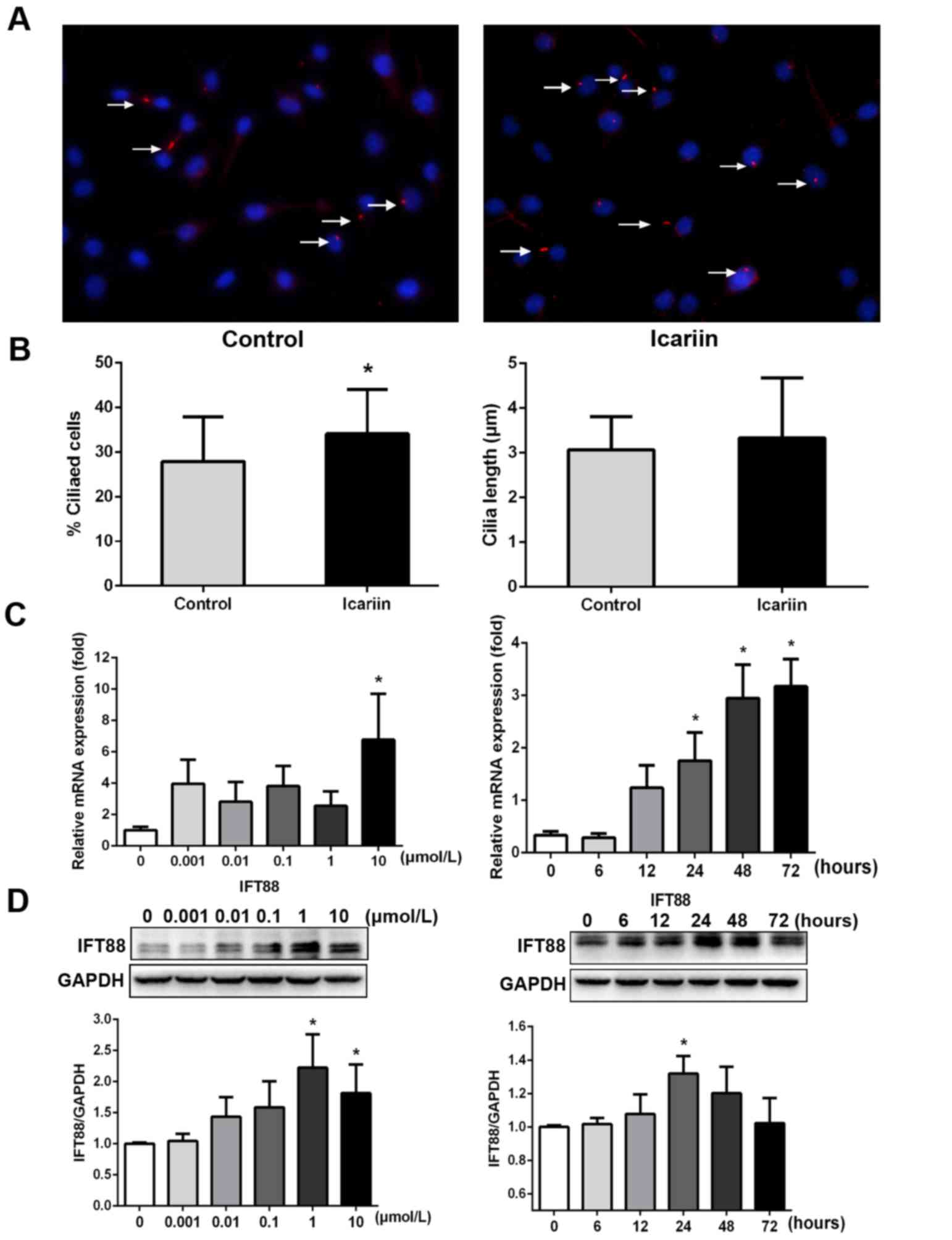 | Figure 2.Icariin promotes ciliary assembly and
IFT88 expression. (A) Primary cilia were stained for
acetylated-α-tubulin (red coloration, denoted by the white arrows;
magnification, ×200). (B) The histograms reveal that, compared with
the control group, 10 µmol/l icariin increased primary ciliary
assembly (Control group, 27.91±9.95% cf. Icariin, 34.06%±10.06;
*P<0.05), and icariin moderately increased the ciliary length
from 3.07±0.74 to 3.34±1.34 µm. (C) Icariin increased IFT88 gene
expression, peaking at 10 µmol/l (*P<0.05 cf. 0 µmol/l icariin).
(D) Icariin upregulated production of the ciliary protein, IFT88,
in a concentration- and time-dependent manner (*P<0.05 cf. 0
µmol/l icariin, or treatment at 0 h). IFT88, intraflagellar
transport protein 88. |
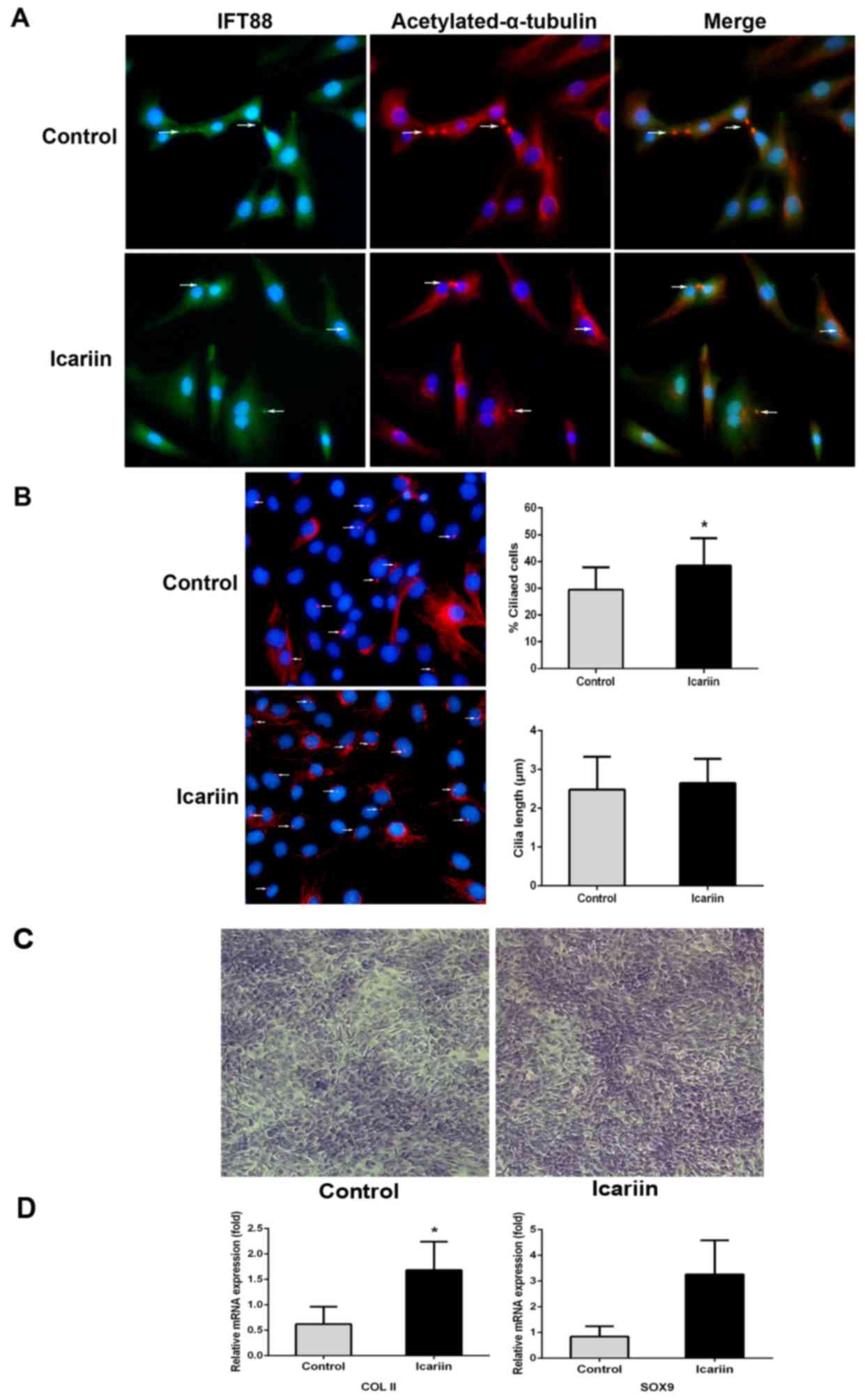
Icariin facilitates ciliary assembly
in primary chondrocytes and assists in maintenance of an
appropriate phenotype
Primary chondrocytes were treated with icariin (10
µmol/l) for 24 h, and the levels of primary cilia and IFT88 were
measured. IFT88 protein was detected in the cytoplasm, although it
was principally centralized along the axonemes of primary cilia
(Fig. 3A). Icariin-treated primary
chondrocytes exhibited enhanced ciliary assembly (38.48±10.36%)
compared with that of the control cells (29.54±8.24%), and a slight
increase in ciliary length was revealed (from 2.48±0.85 to
2.65±0.63 µm; Fig. 3B). Toluidine
blue staining confirmed that icariin-treated primary chondrocytes
secreted more cartilage matrix and expressed higher levels of the
COL-II and SOX9 genes than did the control cells (Fig. 3C and D).
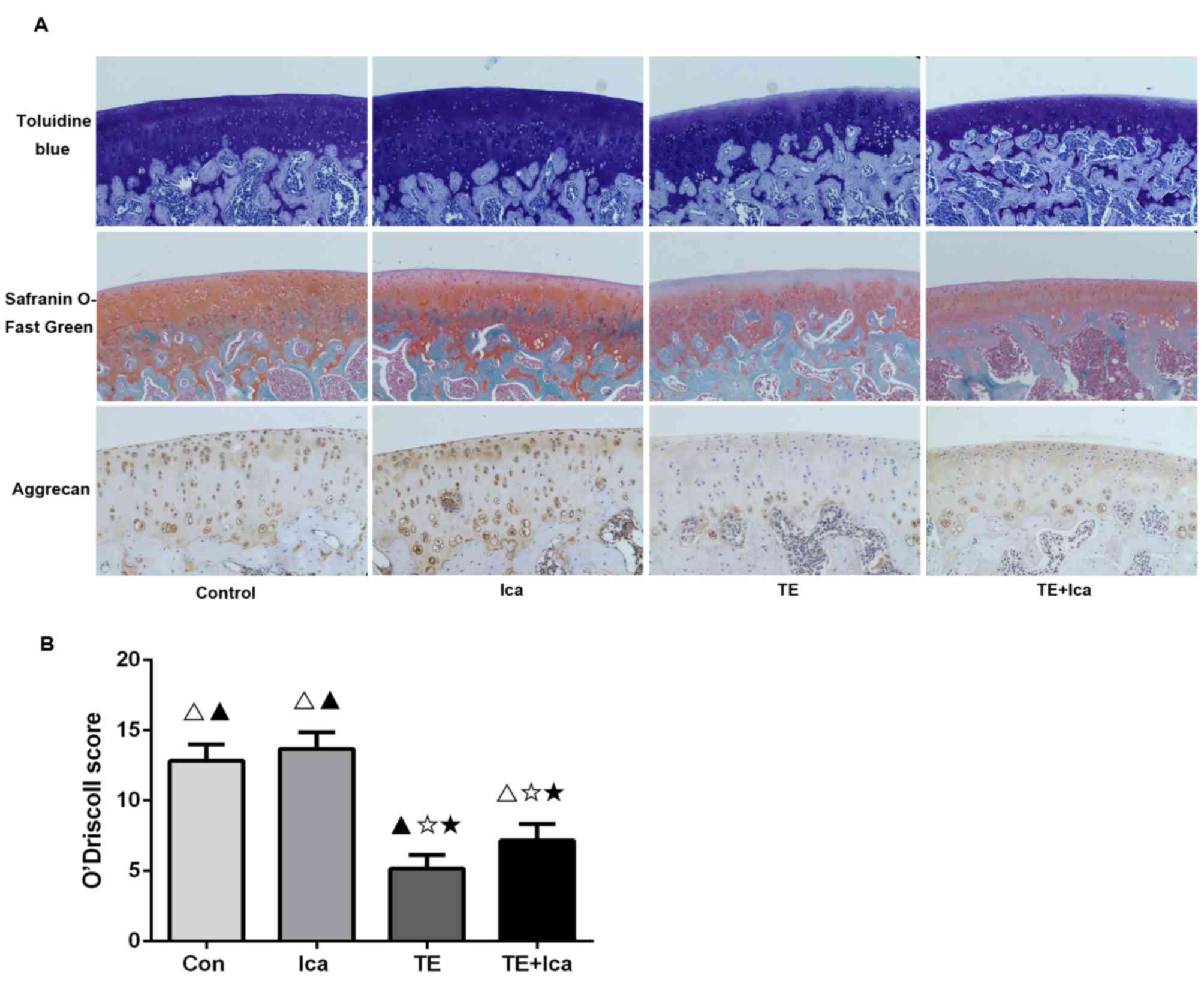
Icariin attenuates cartilage
degeneration in the PTOA rat model
Toluidine blue and Safranin O-Fast Green staining
revealed that the treadmill and treadmill + icariin groups
exhibited reductions in cartilage thickness in weight-bearing
areas, whereas the treadmill group lost more of the superficial
cartilage proteoglycans than did the treadmill + icariin group.
Icariin-treated and control rats had thicker cartilages that were
rich in proteoglycans (Fig. 4A).
Subsequently, the expression of aggrecan (a major component of the
cartilage ECM that allows cartilage to resist compression) was
explored (23). Aggrecan was
clearly expressed in the icariin-treated group, but the treadmill
group exhibited reduced expression in the upper and middle
cartilage layers; addition of icariin partly reversed this decline
(Fig. 4A). When PTOA severity was
assessed by the O'Driscoll system (19), the scores of the control
(12.833±1.169), icariin (13.667±1.211), and treadmill + icariin
(6.833±1.472) groups were all significantly higher compared that of
the treadmill group (5.167±0.983) (Fig. 4B). Therefore, icariin was shown to
improve the histological cartilage phenotype.
ERK phosphorylation is involved in
icariin-mediated IFT88 expression
siRNA was used to knock down IFT88 gene expression,
and this revealed that icariin rescued the reductions in COL-II and
SOX9 expression levels induced by siRNA (Fig. 5A). siRNA downregulated the
expression of endogenous IFT88 and phosphorylated ERK (Fig. 5B). Icariin (10 µmol/l) promoted
IFT88 expression and ERK phosphorylation compared with the levels
noted in the presence of IFT88 siRNA (Fig. 5C). Inhibition of ERK
phosphorylation by PD0325901 clearly reduced IFT88 expression;
icariin did not rescue this decline (Fig. 5D). Therefore, ERK phosphorylation
may be involved in icariin-mediated IFT88 expression during
maintenance of the chondrocyte phenotype.
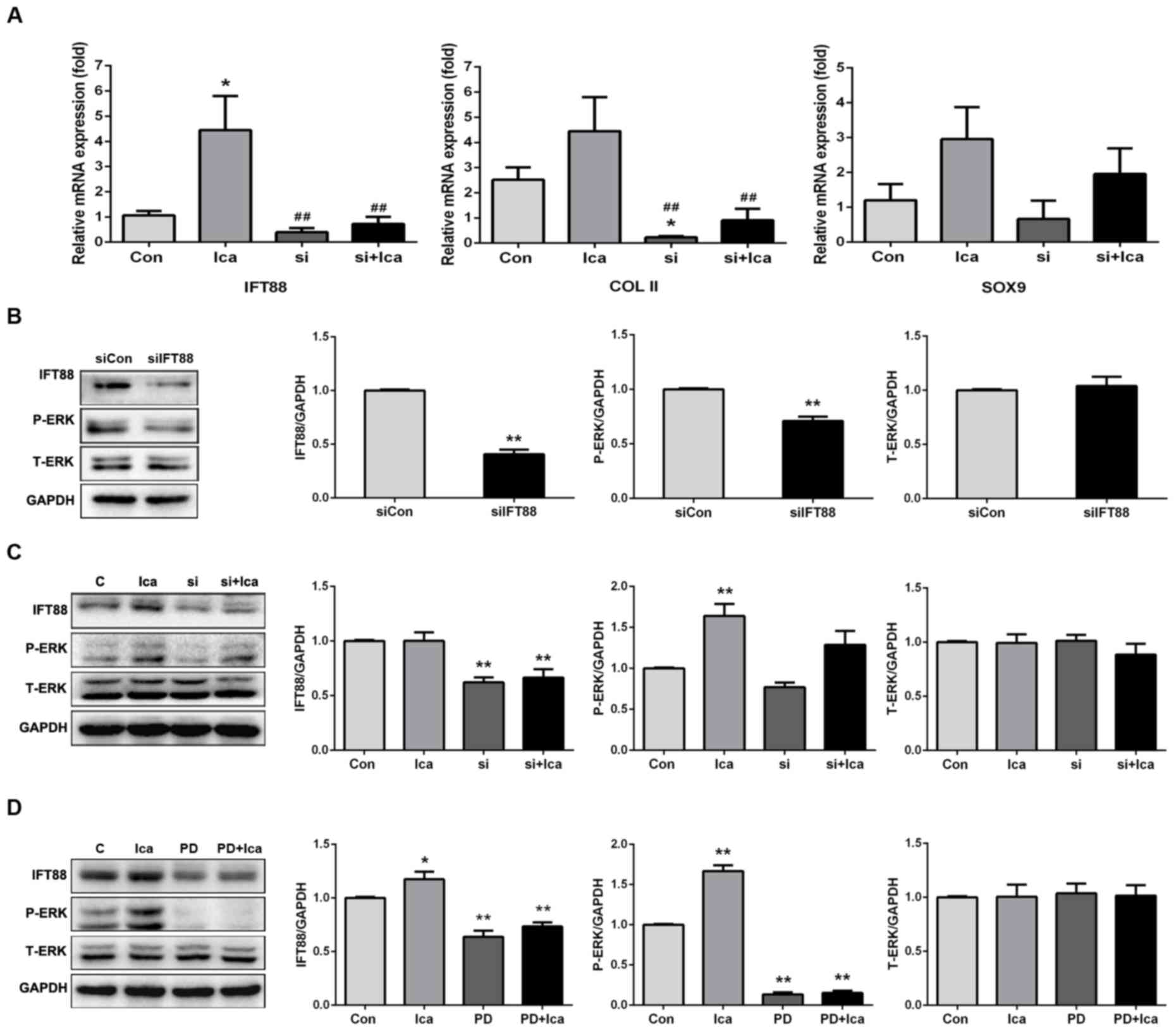 | Figure 5.Icariin regulates IFT88 expression via
the ERK signalling pathway. (A) Quantitative polymerase chain
reaction measuring the expression levels of the IFT88 and various
phenotype-associated genes after transformation of IFT88 siRNA
(*P<0.05 compared with the control group; ##P<0.01
compared with the icariin group). (B) IFT88 knockdown reduced the
expression of phosphorylated ERK (**P<0.01 compared with the
siCon group). (C) Icariin promoted expression of IFT88 and
phosphorylated ERK, and slightly increased ERK phosphorylation,
after transformation of IFT88 siRNA (**P<0.01 compared with the
control group). (D) An ERK inhibitor reduced IFT88 expression.
Icariin upregulated IFT88 expression, but could not restore such
expression in the presence of the ERK inhibitor, PD0325901
(*P<0.05; **P<0.01 compared with the control group). ERK,
extracellular signal-regulated kinase; IFT88, intraflagellar
transport protein 88; Ica, icariin; PD, PD0325901, Con, control;
T-ERK, total ERK; P-ERK, phosphorylated ERK. |
Discussion
Maintenance of the cartilage phenotype is a major
concern during cartilage tissue engineering. Although various
biological materials and recombinant cytokines may be of
assistance, these materials degrade rapidly, exhibit unpredictable
side-effects, and are very costly (24–26).
In the present study, a Herba Epimedii extract, icariin, was shown
to aid in maintenance of the cartilage phenotype, and the IFT88
protein of primary cilia served an important role in this context.
Icariin exerts multiple functions. Li et al (7) demonstrated that, in animal models,
icariin-impregnated hydrogel scaffolds efficiently promoted the
repair of defects in superficial cartilage and osteochondral
regions (7). Zhang et al
(27) showed that icariin promoted
chondrocyte clustering and ECM secretion (27). In the present study, icariin was
shown to promote cellular proliferation, and expression of the
cartilage phenotype genes, COL-II and SOX9, in progenitor
chondrocytic cells and primary chondrocytes was enhanced. These
findings suggested that icariin may be valuable in cartilage tissue
engineering. However, the mechanism by which icariin promotes
cartilage repair has yet to be fully elucidated.
IFT88, an intraflagellar transport protein, carries
cargos into or out of primary cilia (9). This protein has been shown to be very
important in the development of cartilage (10,28).
IFT88 affects actin organisation within chondrocytes, and also the
biomechanical properties of cartilage (28). IFT88 mutations render cartilage
‘osteoarthritis-like’, with reduction of the compressive modulus
(29). IFT88 fulfils a major role
in the columnar organisation of growth plate cartilage (15). In the present study, IFT88 was
shown to be widely distributed in the cytoplasm, although it was
principally concentrated along the ciliary axoneme. Icariin
promoted ciliary assembly, but did not affect ciliary length.
Icariin upregulated IFT88 expression at both the gene and the
protein level, promoted ECM secretion, and increased the expression
of COL-II and SOX9. Therefore, icariin enhanced maintenance of the
cartilage phenotype and IFT88 exerted an important role in
icariin-mediated ECM secretion. IFT88 knockdown suppressed ERK
phosphorylation, and icariin partially rescued this reduction.
Thus, there may be feedback between the IFT88 expression level and
ERK phosphorylation in operation, and icariin contributes
positively to such regulation. It was therefore confirmed that
icariin increased ciliary assembly, enhanced IFT88 expression,
promoted ciliary transportation, stimulated ERK phosphorylation,
and induced SOX9 and COL-II gene expression to promote cartilage
matrix secretion. Primary cilia and the IFT88 protein may serve
important roles during cartilage development. However, the gene
expression profile and the signaling systems involved require
further study.
In conclusion, the present study explored the role
exerted by the ciliary protein IFT88 in icariin-mediated
maintenance of cartilage phenotype in progenitor cells and in
primary chondrocytes. Icariin promoted ciliary assembly, enhanced
intraflagellar transportation, induced ERK phosphorylation, and
stimulated cartilage matrix secretion. Icariin thus aids in the
maintenance of cartilage phenotype, and the present study has
provided the theoretical basis for the use of icariin in cartilage
tissue engineering.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81572094 and 81371915).
References
|
1
|
Varady NH and Grodzinsky AJ:
Osteoarthritis year in review 2015: Mechanics. Osteoarthritis
Cartilage. 24:27–35. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jiang Z, Hu B, Wang J, Tang Q, Tan Y,
Xiang J and Liu J: Effect of icariin on cyclic GMP levels and on
the mRNA expression of cGMP-binding cGMP-specific phosphodiesterase
(PDE5) in penile cavernosum. J Huazhong Univ Sci Technolog Med Sci.
26:460–462. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schluesener JK and Schluesener H: Plant
polyphenols in the treatment of age-associated diseases: Revealing
the pleiotropic effects of icariin by network analysis. Mol Nutr
Food Res. 58:49–60. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Song YH, Li BS, Chen XM and Cai H: Ethanol
extract from Epimedium brevicornum attenuates left ventricular
dysfunction and cardiac remodeling through down-regulating matrix
metalloproteinase-2 and −9 activity and myocardial apoptosis in
rats with congestive heart failure. Int J Mol Med. 21:117–124.
2008.PubMed/NCBI
|
|
5
|
Wu Y, Xia L, Zhou Y, Xu Y and Jiang X:
Icariin induces osteogenic differentiation of bone mesenchymal stem
cells in a MAPK-dependent manner. Cell Prolif. 48:375–384. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhai YK, Guo XY, Ge BF, Zhen P, Ma XN,
Zhou J, Ma HP, Xian CJ and Chen KM: Icariin stimulates the
osteogenic differentiation of rat bone marrow stromal cells via
activating the PI3K-AKT-eNOS-NO-cGMP-PKG. Bone. 66:189–198. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li D, Yuan T and Zhang X, Xiao Y, Wang R,
Fan Y and Zhang X: Icariin: A potential promoting compound for
cartilage tissue engineering. Osteoarthritis Cartilage.
20:1647–1656. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Muhammad H, Rais Y, Miosge N and Ornan EM:
The primary cilium as a dual sensor of mechanochemical signals in
chondrocytes. Cell Mol Life Sci. 69:2101–2107. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Scholey JM: Intraflagellar transport. Annu
Rev Cell Dev Biol. 19:423–443. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Haycraft CJ, Zhang Q, Song B, Jackson WS,
Detloff PJ, Serra R and Yoder BK: Intraflagellar transport is
essential for endochondral bone formation. Development.
134:307–316. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thompson CL, Chapple JP and Knight MM:
Primary cilia disassembly down-regulates mechanosensitive hedgehog
signalling: A feedback mechanism controlling ADAMTS-5 expression in
chondrocytes. Osteoarthritis Cartilage. 22:490–498. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shao YY, Wang L, Welter JF and Ballock RT:
Primary cilia modulate Ihh signal transduction in response to
hydrostatic loading of growth plate chondrocytes. Bone. 50:79–84.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hoey DA, Tormey S, Ramcharan S, O'Brien FJ
and Jacobs CR: Primary cilia-mediated mechanotransduction in human
mesenchymal stem cells. Stem Cells. 30:2561–2570. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Poole CA, Jensen CG, Snyder JA, Gray CG,
Hermanutz VL and Wheatley DN: Confocal analysis of primary cilia
structure and colocalization with the Golgi apparatus in
chondrocytes and aortic smooth muscle cells. Cell Biol Int.
21:483–494. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song B, Haycraft CJ, Seo HS, Yoder BK and
Serra R: Development of the post-natal growth plate requires
intraflagellar transport proteins. Dev Biol. 305:202–216. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ho L, Ali SA, Al-Jazrawe M, Kandel R,
Wunder JS and Alman BA: Primary cilia attenuate hedgehog signalling
in neoplastic chondrocytes. Oncogene. 32:5388–5396. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu W, Shi J, Zhang J, Lv Z, Guo F, Huang
H, Zhu W and Chen A: CXCL12/CXCR4 Axis regulates aggrecanase
activation and cartilage degradation in a post-traumatic
osteoarthritis rat model. Int J Mol Sci. 17(pii): E15222016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
O'Driscoll SW, Keeley FW and Salter RB:
Durability of regenerated articular cartilage produced by free
autogenous periosteal grafts in major full-thickness defects in
joint surfaces under the influence of continuous passive motion. A
follow-up report at one year. J Bone Joint Surg Am. 70:595–606.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Song JQ, Dong F, Li X, Xu CP, Cui Z, Jiang
N, Jia JJ and Yu B: Effect of treadmill exercise timing on repair
of full-thickness defects of articular cartilage by bone-derived
mesenchymal stem cells: An experimental investigation in rats. PLoS
One. 9:e908582014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nian H, Ma MH, Nian SS and Xu LL:
Antiosteoporotic activity of icariin in ovariectomized rats.
Phytomedicine. 16:320–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
de Andrea CE, Zhu JF, Jin H, Bovée JV and
Jones KB: Cell cycle deregulation and mosaic loss of Ext1 drive
peripheral chondrosarcomagenesis in the mouse and reveal an
intrinsic cilia deficiency. J Pathol. 236:210–218. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ke YN and Yang WX: Primary cilium: An
elaborate structure that blocks cell division? Gene. 547:175–185.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gibson BG and Briggs MD: The
aggrecanopathies; an evolving phenotypic spectrum of human genetic
skeletal diseases. Orphanet J Rare Dis. 11:862016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hunziker EB, Lippuner K, Keel MJ and
Shintani N: An educational review of cartilage repair: Precepts
& practice - myths & misconceptions-progress &
prospects. Osteoarthritis Cartilage. 23:334–350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Demoor M, Ollitrault D, Gomez-Leduc T,
Bouyoucef M, Hervieu M, Fabre H, Lafont J, Denoix JM, Audigié F,
Mallein-Gerin F, et al: Cartilage tissue engineering: Molecular
control of chondrocyte differentiation for proper cartilage matrix
reconstruction. Biochim Biophys Acta. 1840:2414–2440. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Johnstone B, Alini M, Cucchiarini M, Dodge
GR, Eglin D, Guilak F, Madry H, Mata A, Mauck RL, Semino CE and
Stoddart MJ: Tissue engineering for articular cartilage repair-the
state of the art. Eur Cell Mater. 25:248–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang L, Zhang X, Li KF, Li DX, Xiao YM,
Fan YJ and Zhang XD: Icariin promotes extracellular matrix
synthesis and gene expression of chondrocytes in vitro. Phytother
Res. 26:1385–1392. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Z, Wann AK, Thompson CL, Hassen A,
Wang W and Knight MM: IFT88 influences chondrocyte actin
organization and biomechanics. Osteoarthritis Cartilage.
24:544–554. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Irianto J, Ramaswamy G, Serra R and Knight
MM: Depletion of chondrocyte primary cilia reduces the compressive
modulus of articular cartilage. J Biomech. 47:579–582. 2014.
View Article : Google Scholar : PubMed/NCBI
|