Introduction
Cardiovascular disease is the leading
non-communicable disease in terms of mortality; in 2012 there were
~17.5 million moralities caused by cardiovascular diseases, among
which ~7.4 million were caused by coronary heart disease (CHD)
(1). The detrimental effects of
acute myocardial ischemia-reperfusion (IR) injury are usually
responsible for the outcome of CHD (2). During spaceflight, exposure to
microgravity may lead to a series of maladaptive alterations in a
number of organs, resulting from the redistribution of body fluids
towards the head, which is caused by the considerable stress on the
cardiovascular system (3). Data
from our previous study revealed that microgravity increases
susceptibility to myocardial IR injury in tail-suspended rats
(4); however, the underlying
mechanisms have not been fully elucidated.
The Notch gene was originally cloned from
Drosophila melanogaster >30 years ago; it encodes a
family of highly conserved transmembrane surface receptors
(5). Mammals express 4 Notch
receptors, Notch1-Notch4, and 5 Notch ligands, including Delta-like
(DLL)1, DLL3, DLL4, Jagged (JAG)1 and JAG2. The binding of a ligand
to a receptor triggers the proteolytic cleavage and release of the
Notch intracellular domain (NICD), which is the active form of
Notch (6). Following translocation
into the nucleus, NICD functions to regulate the transcription of
its downstream target genes, including HES family basic
helix-loop-helix transcription factor (HES) and HES-related with
YRPW motif (HEY), which in turn modulate cell proliferation,
apoptosis and the expression of transcription factors of the
nuclear transcription factor-κB (NF-κB) family (7). The Notch signaling pathway has been
revealed to participate in various cellular behaviors and
processes, including cell proliferation, apoptosis, differentiation
and adhesion by regulating communication between adjacent cells
(8), thus indicating its important
role in controlling the development of organs and tissues.
Notably, a previous study demonstrated that a
deficiency in Notch signaling aggravates hepatic IR injury, whereas
induced Notch activation decreases cell apoptosis following hepatic
IR (9), which suggested that Notch
may serve a crucial role in adult animals. Among the Notch
receptors and ligands, Notch1 and JAG1 are the predominant forms
expressed in adult heart (10).
Numerous previous studies have reported that the Notch1 signal
exhibits significant enhancement following myocardial IR,
suggesting that cardiac Notch1 signaling may be endogenously
activated and may exert its beneficial effects during myocardial IR
(11–13). Nevertheless, whether Notch1
protects against myocardial IR injury under simulated
weightlessness remains unclear and requires further
investigation.
Therefore, the present study was designed to
investigate the role of the Notch1 receptor in myocardial IR under
simulated weightlessness. The temporal and spatial characteristics
of Notch1 expression in myocardial IR were determined by detecting
the extracellular domain of Notch1, where the signaling pathway was
initiated. Furthermore, it was demonstrated that simulated
weightlessness aggravated myocardial IR injury by downregulating
Notch. These data may add to the understanding of the mechanisms of
myocardial IR injury in a weightless environment, which may aid in
the design of therapies for astronauts or other members who work on
the space station.
Materials and methods
Chemicals and reagents
The extracellular domain of Notch receptors consists
of 29–36 multiple epidermal growth factor-like motifs that are
responsible for ligand interaction and 3 LIN-12/Notch motifs that
are responsible for precluding receptor activation without
receptor-ligand engagement (14,15).
The NICD is the activated form of Notch1 that directly regulates
the transcription of downstream genes (7). Therefore, antibodies used against the
extracellular domain of Notch1 may indicate how and where Notch1 is
distributed. Primary antibodies against Notch1 extracellular domain
(cat. no. sc-23299) and JAG1 (cat. no. sc-8303) and connexin 43
(Cx43; cat. no. sc-13558) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA), and anti-β-actin (cat. no.
A5441) was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany). The membrane marker Texas Red®-X-conjugated
wheat germ agglutinin (WGA; cat. no. W21405) was purchased from
Molecular Probes (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Cx43 and WGA were used to distinguish between the different
zones of infarcted myocardium under microscopy. Unless otherwise
indicated, all chemicals were purchased from Sigma-Aldrich (Merck
KGaA).
Animals and samples
All Sprague-Dawley (SD) rats (n=44) were obtained
from Beijing Vital River Laboratory Animal Technology Co., Ltd.
(Beijing, China). All the animals were fed and provided with water
ad libitum in an environment with temperature of 22±1°C,
relative humidity of 50±1% and a 12-h light/dark cycle. All animal
studies, including the rat euthanasia by carbon dioxide
asphyxiation (90% CO2, 5 min), were performed in
compliance with the regulations and guidelines, and approved by the
Ethics Committee of The Fourth Military Medical University
institutional animal care and were conducted according to the
AAALAC (https://www.aaalac.org/) and the IACUC
(https://www.iacuc.org/) guidelines.
The antibody against Notch1 in the present study has
rarely been used before to detect myocardial Notch1. Therefore, the
feasibility of using this antibody to detect myocardial Notch1
expression had to be validated. As the liver is one of the organs
where Notch is abundantly expressed in the adult stage, hepatic
Notch1 expression was used as the positive control for the
validation of the experimental method, as well as for further
comparisons of Notch expression between other groups. Accordingly,
hearts and livers were sourced from male SD rats that were 14 days
embryonic (embryonic), 3 days old (neonatal) and 8 weeks old
(adult) (n=4 rats/age group) to reveal the expression patterns of
the Notch signal. Additionally, 32 adult rats were used to
elucidate the effect of simulated microgravity on the expression of
Notch signal after myocardial IR.
Tail-suspension model and IR model in
vivo
Adult rats (weight, 200–250 g) were randomly divided
into the following 6 groups prior to tail-suspension and surgery:
i) Control 3-h IR without suspension (CON-IR-3h, n=4); ii) Control
7-day IR without suspension (CON-IR-7d, n=8); iii) 3-h IR following
4-week suspension (SUS-IR-3h, n=4); iv) 7-day IR following 4-week
suspension (SUS-IR-7d, n=8); v) sham without suspension (CON-Sham,
n=4); vi) and sham following 4-week suspension (SUS-Sham, n=4)
(16). Tail-suspended rats were
maintained in a 30° head-down tilt and hindlimb unloading position
for 4 weeks (17). Rats were
anesthetized with sodium pentobarbital (40 mg/kg, intraperitoneal;
Sigma-Aldrich; Merck KGaA) prior to IR surgery. Myocardial ischemia
was induced by ligation of the left anterior descending coronary
artery (LAD) with a slipknot, which was subsequently removed 30 min
later (18). Sham-operated control
rats underwent the same surgical procedures except that the suture
placed under the LAD was not tied. SUS-IR-7d rats were returned to
the tail-suspension state post-surgery.
Immunofluorescent histochemistry and
confocal analysis
Myocardial and hepatic samples sourced from rats
were cut into 1 mm3 pieces and frozen with optimum
cutting temperature compound and isopentane in liquid nitrogen
(Sigma-Aldrich; Merck kGaA). Tissue sections (10 µm) were obtained
in a freezing cryostat at −20°C. Sections were air dried at room
temperature, fixed in ice-cold acetone for 30 min, permeabilized in
the phosphate buffered saline [PBS; 135 mM NaCl, 10 mM sodium
phosphate, (pH 7.0)] including 0.1% Triton X-100 at room
temperature for 30 min, and subsequently blocked in 1% bovine serum
albumin (BSA; cat. no. 9048-46-8; Ameresco, Inc., Framingham, MA,
USA) in PBS at room temperature for 60 min. Sections were incubated
with anti-Notch1 (1:50), anti-JAG1 (1:50) or anti-Cx43 (1:50)
primary antibodies at 4°C overnight. Slides were rinsed twice in
PBS and incubated with rabbit anti-goat Alexa Fluor 488-conjugated
immunoglobulin IgG (1:800; cat. no. A27012; Invitrogen; Thermo
Fisher Scientific, Inc.), goat anti-rabbit Alexa Fluor
488-conjugated immunoglobulin IgG (1:800; cat. no. A27034;
Invitrogen; Thermo Fisher Scientific, Inc.), and goat anti-mouse
Cy3-conjugated IgG (1:1,000; cat. no. A0521; Beyotime Institute of
Biotechnology) at 37°C for 60 min. Tissue sections were
counterstained with Hoechst 33258 (1 µg/ml) and WGA (5 µg/ml) at
37°C for 8 min. Stained sections were observed using an Olympus
FV1000 laser-scanning confocal microscope (Olympus Corporation,
Tokyo, Japan) equipped with the FV10-ASW 3.1. Images were captured
using a ×60 magnification water objective. Optical densitometric
analysis of target protein expression was performed using the
FV10-ASW 3.1 (Olympus Corporation, Tokyo, Japan). For each section,
mean fluorescence was calculated from three separate fields per
heart.
Protein extraction and western blot
analysis
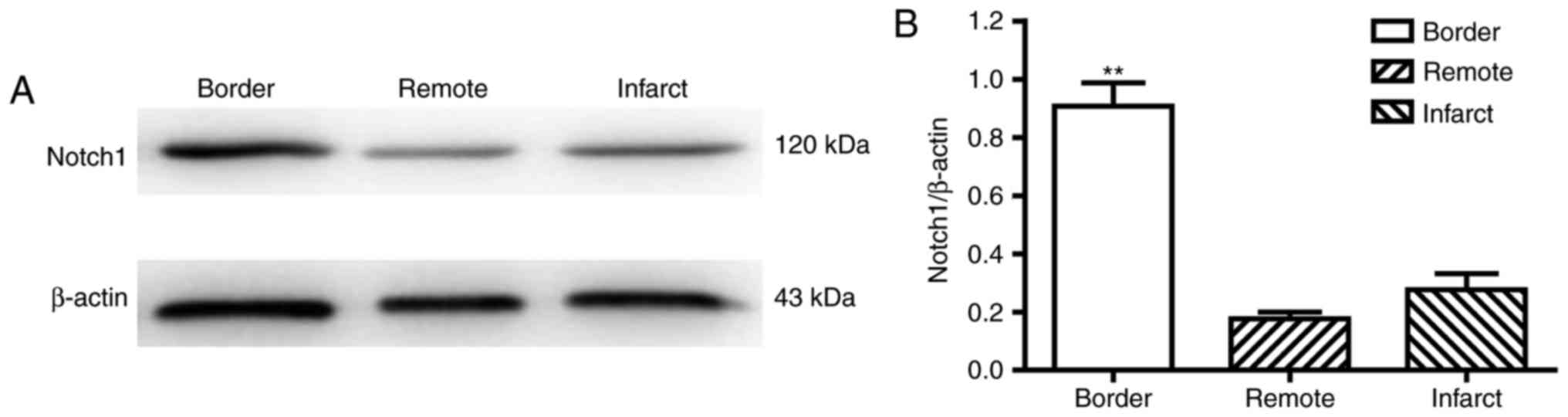
The heart samples (n=4 rats) were collected from the
CON-IR-7d group 7 days following myocardial IR surgery to confirm
and limit assessment to the spatial features of the border zone.
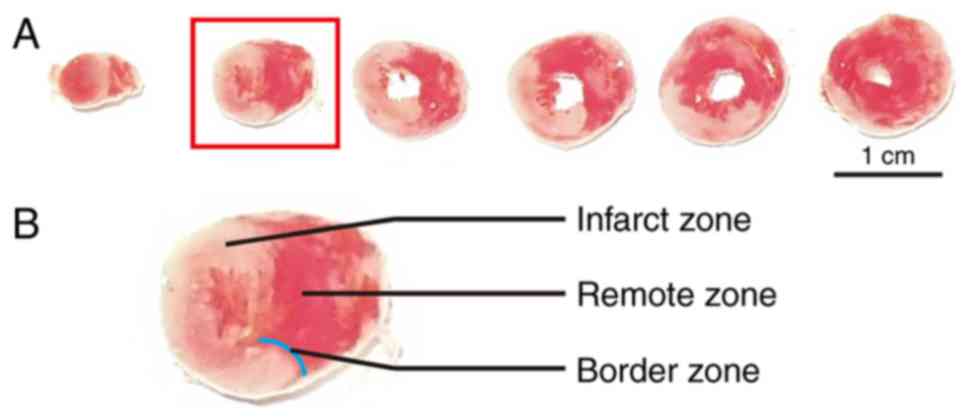
The infarcted hearts were then sectioned horizontally into 6 pieces
from apex to base, among which the second section was usually the
most typical for the border zone division (Fig. 1A). According to previous studies
(19,20), the ischemic, but still viable,
‘border zone’ was ~2 mm away from the border of the macroscopic
(pink or white colored) infarct zone (Fig. 1B). Thus, the myocardial protein
extracts from border, remote, and infarct zone of the second
section were resolved by SDS-PAGE using 12% Laemmli gels, as
described previously (21).
Following electrophoresis, proteins were electrically transferred
to polyvinylidene fluoride membrane using a Bio-Rad semi-dry
transfer apparatus (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The membranes were blocked with 1% BSA in Tris-buffered saline
[TBS; 150 mM NaCl, 50 mM Tris-HCl, (pH 7.5)] and were incubated
with a goat polyclonal antibody against the extracellular domain of
Notch1 (1:1,000) and mouse monoclonal anti-β-actin (1:4,000) in TBS
containing 0.1% BSA at 4°C overnight. The membranes were incubated
with IRDye 800CW-conjugated donkey-anti goat (1:10,000; cat. no.
926-32214; LI-COR, Inc.) or IRDye 800CW-conjugated goat-anti mouse
secondary antibodies (1:10,000; cat. no. 926-32210; LI-COR, Inc.)
for 90 min at room temperature, and were visualized using an
Odyssey scanner (LI-COR Biosciences, Lincoln, NE, USA).
Quantification analysis of blots was performed with the ImageJ
software v1.43u (National Institutes of Health, Bethesda, MD, USA).
Notch1 expression was normalized to β-actin.
Myocardial infarct size
Hearts were arrested in diastole by perfusing with
cardioplegic solution (25 mmol/l KCl, 5% dextrose in PBS) at the
end of reperfusion. To distinguish between viable and infarcted
tissue, hearts were perfused and incubated in 1%
triphenyltetrazolium chloride Sigma-Aldrich (Merck KGaA; TTC, in
PBS, (pH 7.4)] for 10 min at 37°C (22). The base and the right ventricular
free wall were dissected, and the left ventricle was frozen at
−20°C for 2 h and the heart was subsequently sectioned from the top
to the bottom into coronal sections. The sections were fixed at 4°C
overnight in 4% paraformaldehyde; the sections were weighed to
calculate the total mass of the left ventricle and the sides that
touched the bottom of the container were imaged the next day.
Following incubation with TTC, viable tissue was stained red,
whereas necrotic (infarcted) tissue turned white. The total mass of
the left ventricle was measured by adding the masses of the
individual slices. The infarct size was then expressed as the
percentage of the mass of the left ventricle. The areas of infarct
were analyzed using Image J software v1.43u.
Statistical analysis
Data were statistically analyzed with the SPSS 17.0
software (SPSS Inc., Chicago, IL, USA). Data are presented as the
mean ± standard error of the mean from at least three independent
experiments. Student's t-test was used for paired observations.
Comparisons were performed using two-way ANOVA for experiments with
more than two subgroups. P<0.05 was considered to indicate a
statistically significant difference.
Results
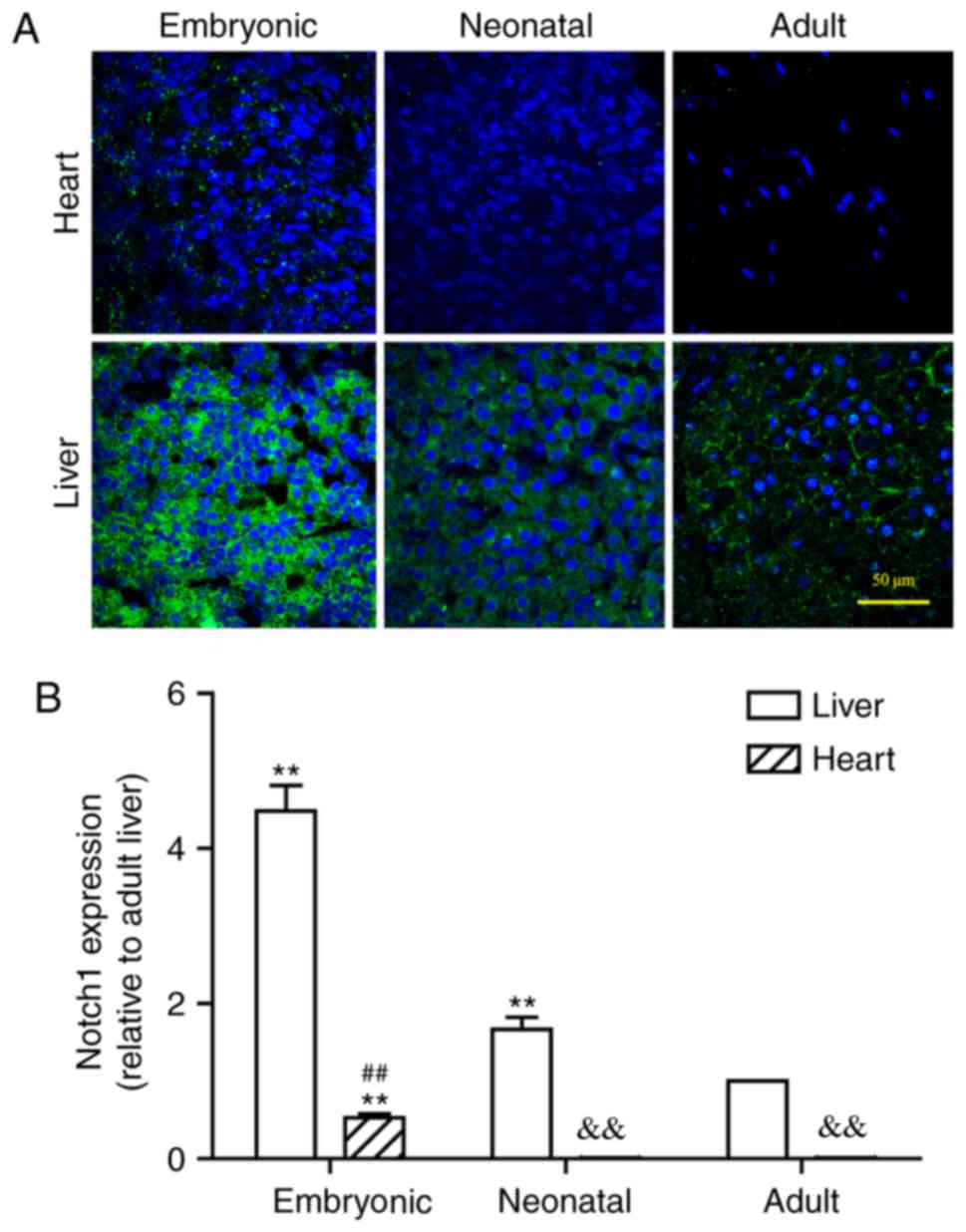
Hepatic Notch expression gradually
decreases during development
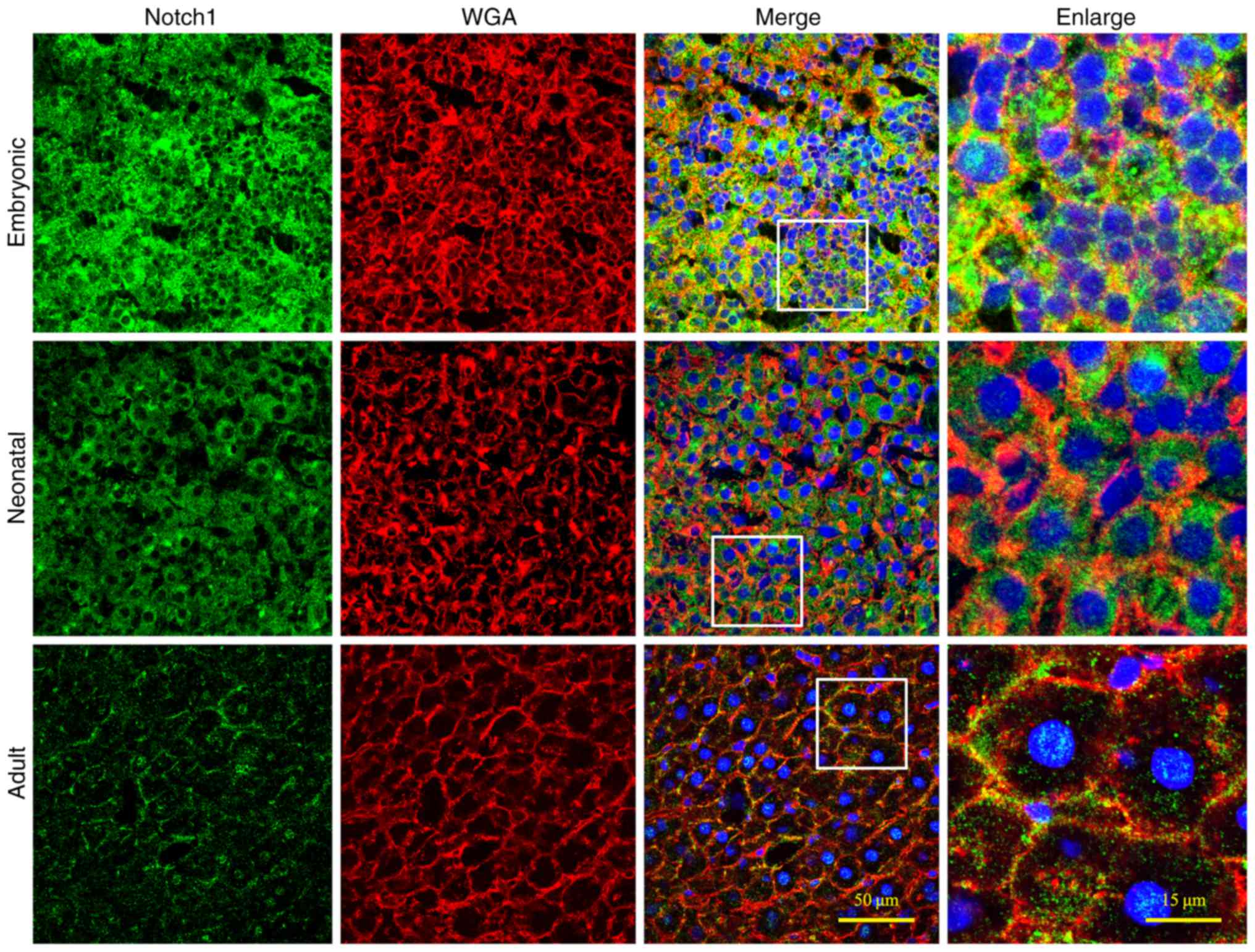
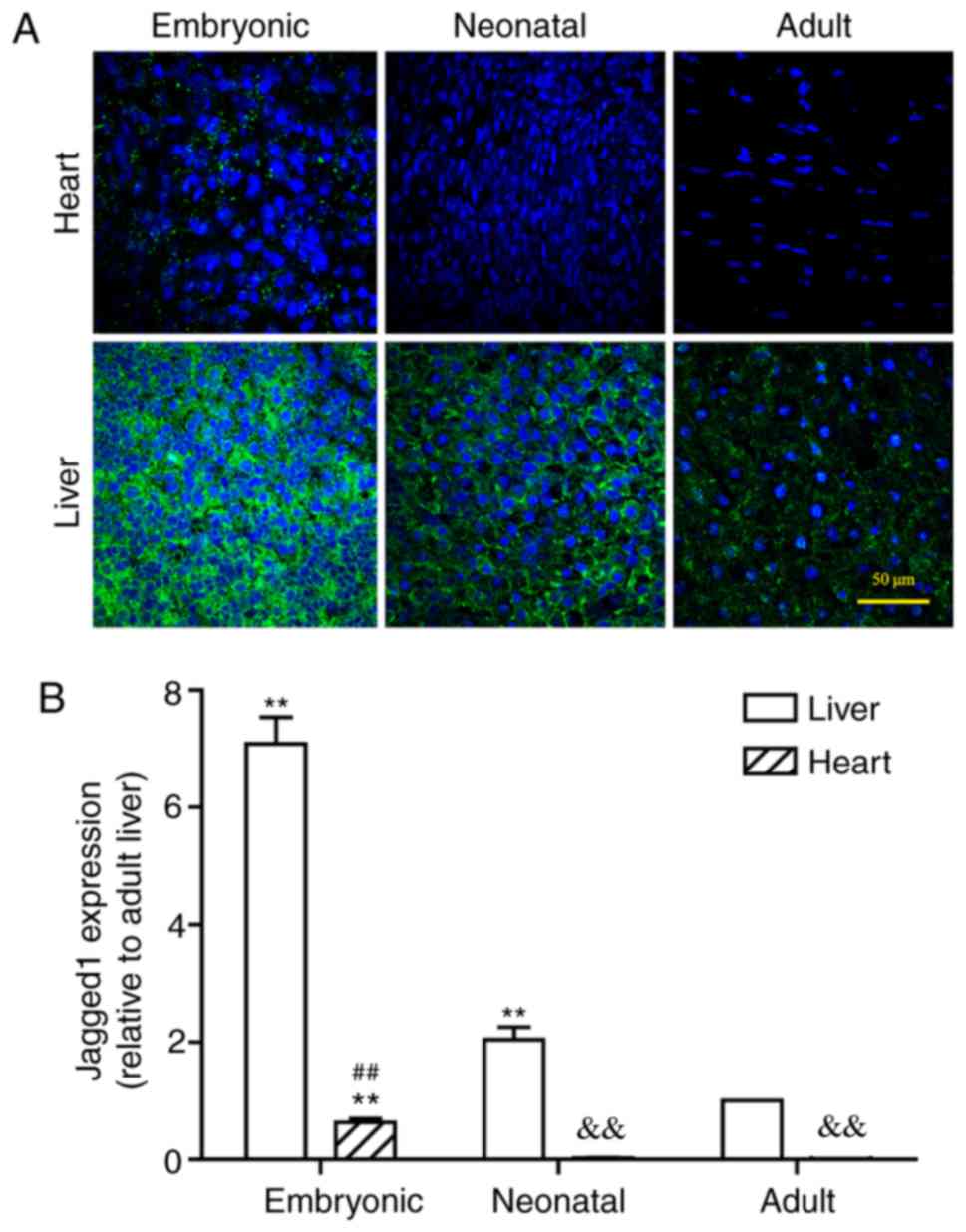
Notch1 protein was abundantly expressed in embryonic
liver, but it was notably decreased in neonatal and adult liver
tissues (Fig. 2). In the adult
liver, Notch1 expression was observed in a ring-shaped manner
around each hepatocyte, which indicated that the protein was
localized to the cell membrane. This concurs with the use of the
Notch antibody in the present study against the extracellular
domain, which may better indicate how and where Notch1 signal gets
distributed and explains why Notch1 signal in the present study
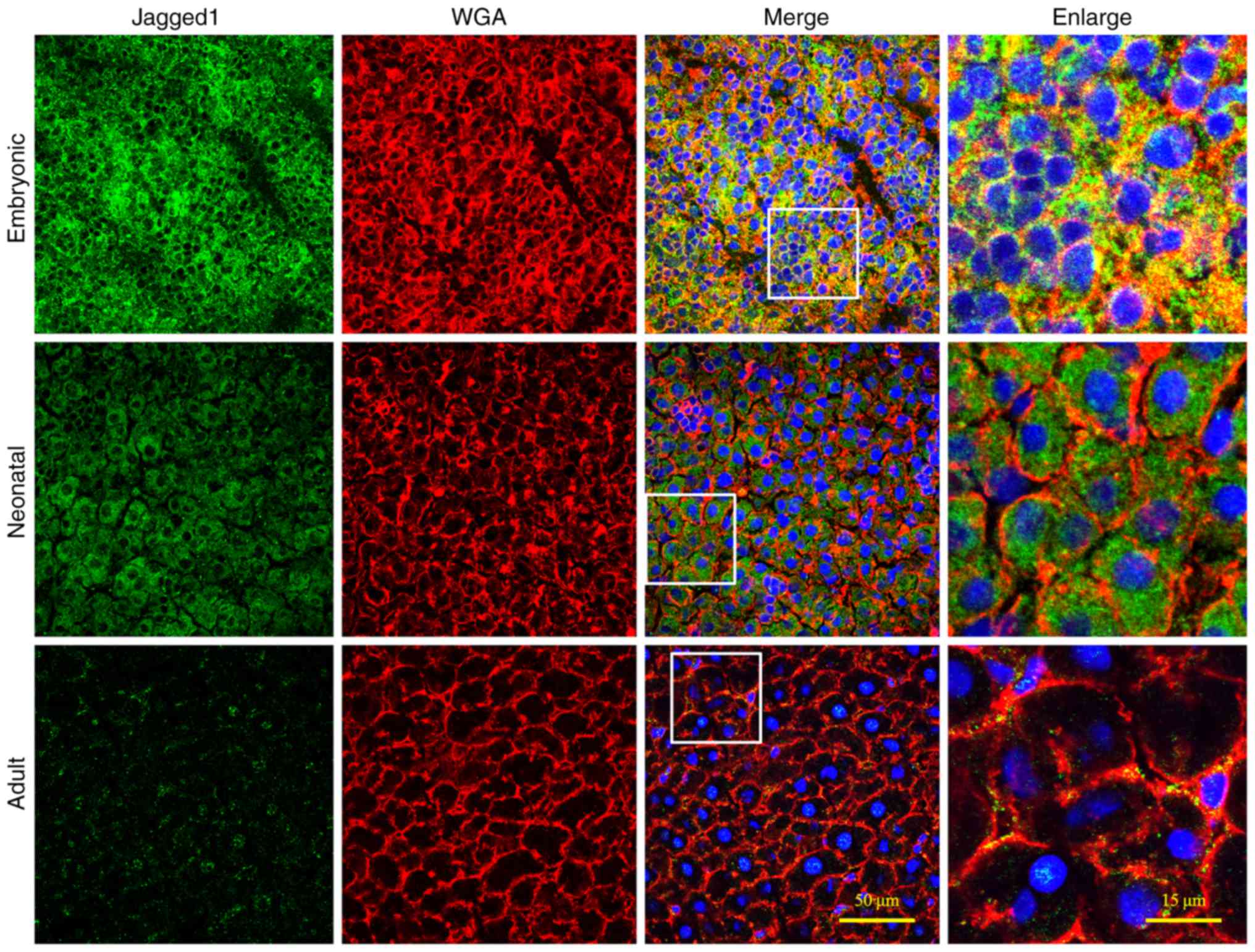
always lies near the cell membrane. Similarly, JAG1 expression was
also intensely expressed in the embryonic liver (Fig. 3), whereas the levels of hepatic
JAG1 expression were decreased in neonatal livers and became even
lower at the adult stage. In the adult liver, JAG1 expression was
distributed in a similar ring-shaped pattern to Notch1 expression
(Fig. 2). In conclusion, these
results revealed that Notch expression in rat liver gradually
decreased during development.
Notch is expressed at lower levels in
heart compared with liver
It must be noted that myocardial Notch1 expression
has rarely been studied using the Notch1 antibody against the
extracellular domain. In addition, the expression levels of Notch
signal in the adult and neonatal hearts are too low to be visually
defined. Therefore, the expression of the Notch signal in both the
liver and heart at the three different developmental periods
(Figs. 2–5) has been compared to validate the
feasibility of using these antibodies to detect myocardial Notch
signal, with the expression level in the adult liver being used as
a baseline. Lower expression of Notch1 was observed in the hearts
of the neonatal and adult rats compared with expression in the
liver at these respective stages (Fig.
4A). Even at the embryonic stage of the rat heart, Notch1
expression level was significantly lower compared with the adult
liver. Following statistical comparison, it was demonstrated that
the expression levels of Notch1 in liver were >10 times higher
compared with the heart at the same developmental stage. Notably,
Notch1 expression levels of both neonatal and adult hearts were
significantly less than that of embryonic hearts (P<0.01;
Fig. 4B). Similarly, the levels of
JAG1 protein expression were low in the embryonic heart but
significantly higher than in neonatal or adult hearts, in both of
which JAG1 was almost undetectable (P<0.01; Fig. 5A). Furthermore, at the same stage,
cardiac JAG1 expression levels were significantly lower compared
with the hepatic JAG1 expression levels (P<0.01; Fig. 5B).
Notch1 expression is enhanced in the
border zone following myocardial IR
Previous studies have demonstrated that Notch
expression is elevated in hearts subjected to myocardial IR injury
(10,11). To further determine the area in the
infarcted heart in which Notch1 expression is the highest, the
levels of Notch1 protein within the infarct, remote and border
zones were measured 7 days following IR (CON-IR-7d). Significantly
higher expression of Notch1 was observed in border zone compared
with the infarct or remote zone (Fig.
6), which suggested that Notch1 may exhibit protective effects
in the border zone of infarcted heart, during myocardial IR.
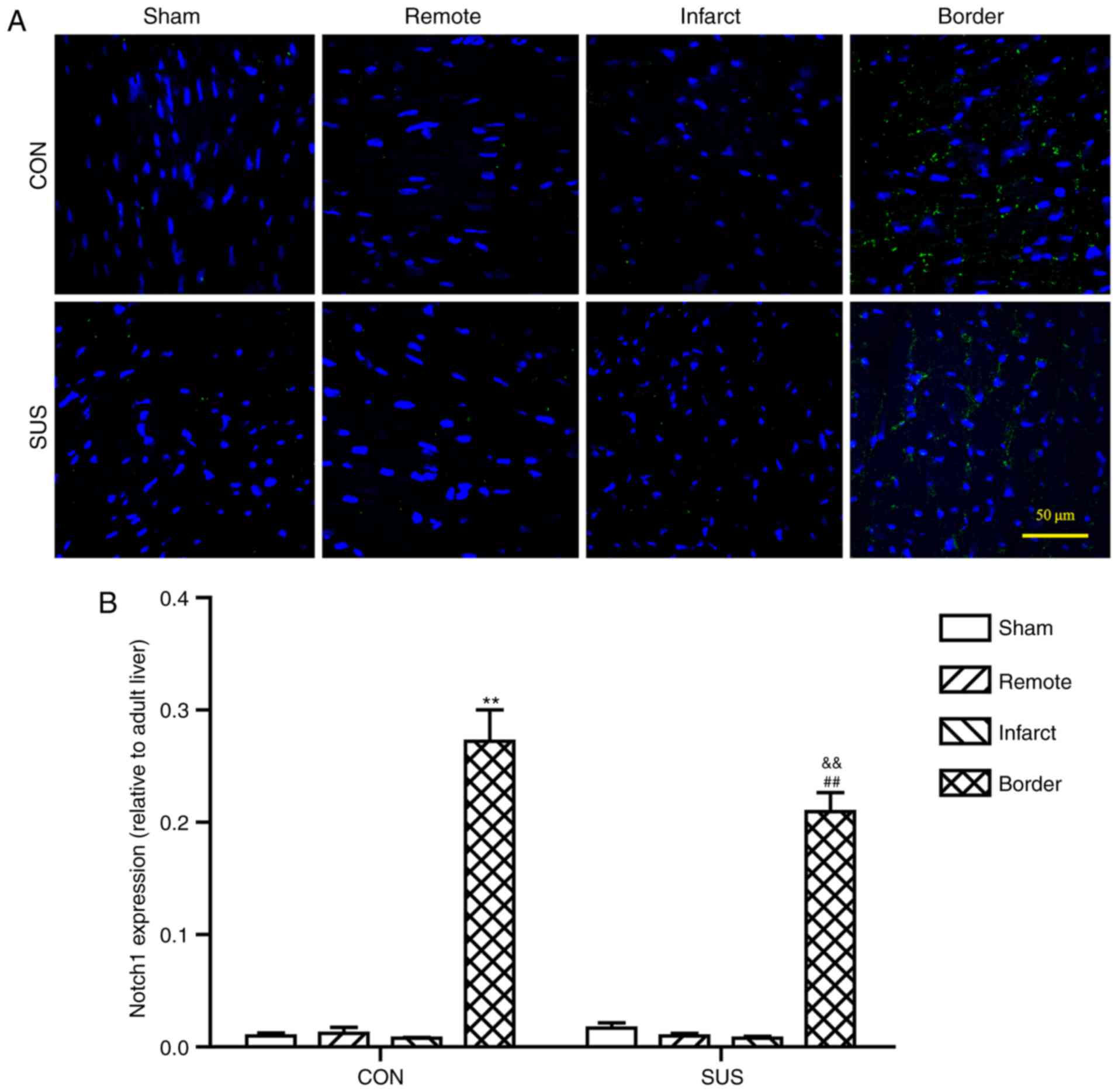
Notch1 expression levels are reduced
in the border zone of tail-suspended rat hearts following
myocardial IR injury
Notch1 protein expression levels were higher in the
border zone compared with expression levels in CON-Sham rats,
SUS-Sham rats, and in the infarcted area and the remote zone, of
the same age in both CON-IR-7d and SUS-IR-7d rats (Fig. 7). However, Notch1 expression was
significantly lower in SUS-IR-7d rats compared with expression
levels in CON-IR-7d rats (Fig.
7B).
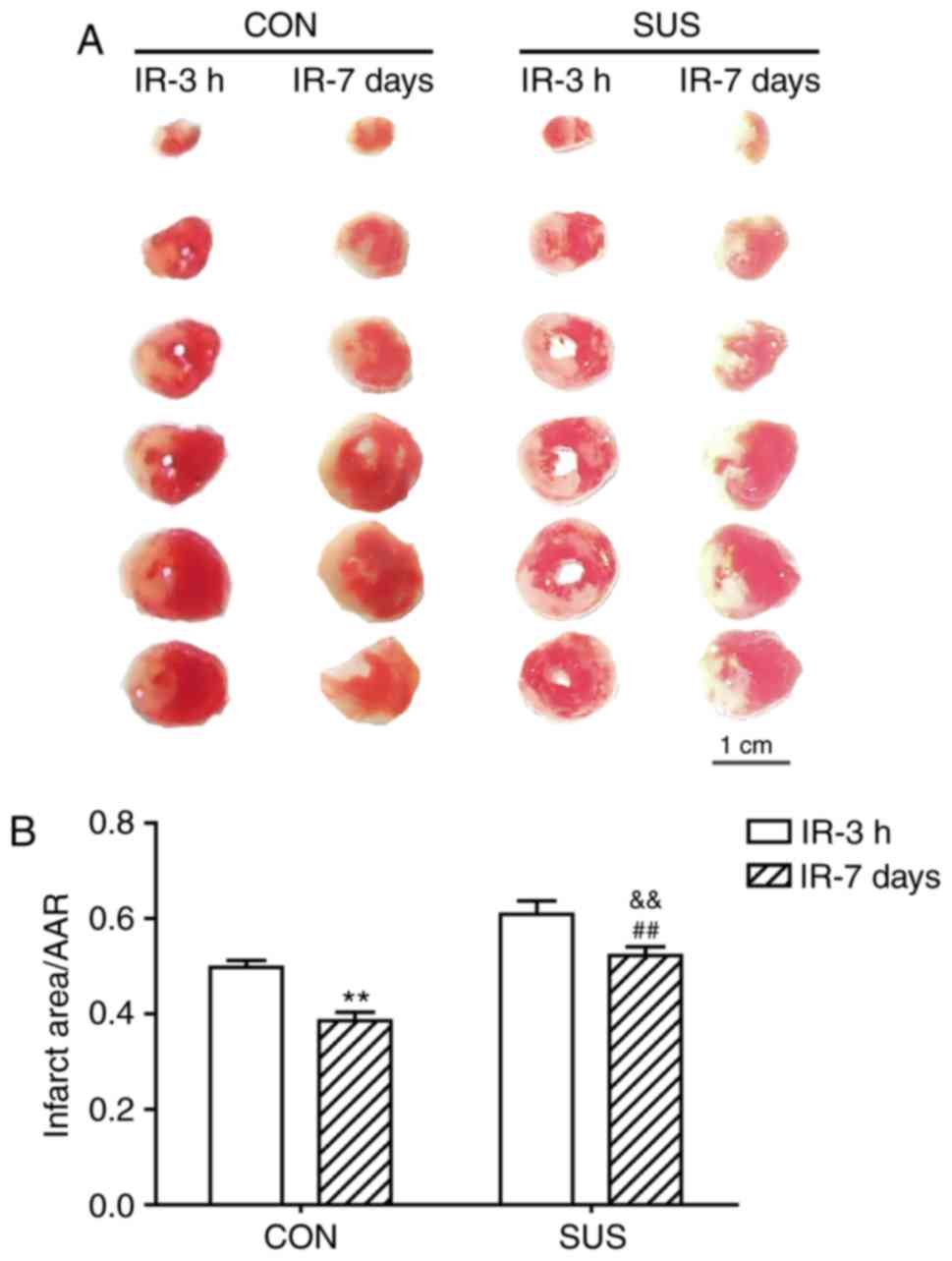
Infarcted zone size is reduced
following myocardial IR
The myocardial infarct size was significantly
smaller in rats that underwent 7 days of reperfusion compared to
those who underwent 3 h of reperfusion in both CON and SUS groups
(Fig. 8). In addition, the
myocardial infarct size was significantly larger in both SUS groups
compared with both of the corresponding CON groups post-IR.
Discussion
The gravitational field is crucial for regulating
cardiovascular function. It has been reported that astronauts
exhibit decreased cardiac preload, circulating blood volume and red
cell mass following long-term exposure to microgravity, which in
turn may jeopardize the ability of the body to fight against
cardiovascular injury (23). Our
previous data have demonstrated that the myocardium is more
sensitive to IR injury under microgravity exposure (4). In the present study, a 4-week tail
suspension rat model was used to simulate microgravity and to
further investigate the detailed mechanisms behind this change.
Notch signaling pathway is a conserved cell-cell
interaction system that controls tissue formation and homeostasis
during embryonic and adult life (6). Previous studies have demonstrated
that Notch serves multifaceted roles in cardiac development,
including ventricular chamber development (24) and embryonic vasculature remodeling
(25). Additionally, Notch
signaling participates in the maintenance of adult heart tissue
integrity (10,26). Notch1 is also involved in heart
diseases, including myocardial infarction (27) and hypertrophic cardiomyopathy
(10). In particular, Notch
activation mediates cardioprotection provided by ischemic
preconditioning and ischemic postconditioning (28). Previous studies have reported that
Notch1 signaling also contributes to the suppression of
IR-injury-induced cardiac oxidative/nitrative stress (12,13).
Therefore, Notch1 upregulation may represent a potential
pharmacologic target for the cardioprotection of ischemic heart
disease. However, it remains unknown whether Notch1 functions
properly in myocardial IR under microgravity exposure. In the
present study, the expression and distribution of Notch1 expression
was investigated. Subsequently, the differences in Notch signaling
expression following myocardial IR were compared between control
and tail-suspended rats following IR injury.
Previous studies have demonstrated the expression of
Notch1 and JAG1 in fetal and postnatal rat and human liver tissues
(29,30). Since most studies that examined the
myocardial Notch1 expression by the NICD levels (12), the present study is the first to
the best of the authors' knowledge to confirm that the Notch1
antibody against the extracellular domain can detect myocardial
Notch1 levels. Data from the present study also indicated that the
hepatic expression of Notch1 and JAG1 declined remarkably from
embryo to the adult stage. Importantly, the expression of Notch1
and JAG1 in normal heart is far lower compared with the expression
in liver at any developmental stage.
Notch1 has been previously reported to exert
cardioprotective effects in myocardial IR injury (13). Owing to the negligible regenerative
capacity of the heart, myocardial infarction ultimately leads to
the replacement of necrotic cardiomyocytes with a collagen-based
scar. A previous study divided the cellular and molecular events
involved in the reparative response into three overlapping phases
(16). During the inflammatory
phase, the activation of cytokines and chemokines induces a
protease-rich environment, which in turn leads to extensive
degradation of the cardiac matrix (31–33).
The transition to the proliferative phase undergoes the inhibition
of pro-inflammatory signals and the promotion of fibrous tissue
deposition and angiogenesis (34).
During the proliferative phase, the restricted expressions of
fibronectin and matricellular proteins regulate the phenotype,
survival and gene expression of cardiac cells (35). Finally, the formation of a mature
cross-linked matrix may ‘shield’ fibroblasts and further lead to
their deactivation during the maturation phase (36,37).
Of the three phases, myocardial reparative response mainly occurs
during the maturation phase, which in rodents is between 5 and 25
days, and in large mammals is between 14 days and 2 months.
Therefore, the present study focused on Notch1 expression during
the maturation phase of myocardial IR injury. Notable increases of
Notch1 expression were observed in the border zone at 7 days
post-IR injury, which was consistent with the study by Gude et
al (27).
The border zone surrounding a healed myocardial
infarction is a region where the normal organization of cardiac
cells gets disrupted (20).
Tremendous effort has been devoted to the research on limiting
infarct size. Thereinto, the size of the ischemic but still viable
border zone has received increasing attention in experimental
animals (38,39). The present data indicated a
significant increase in Notch1 expression levels at the border zone
compared with the remote and the infarcted zones, 7 days
post-myocardial IR injury (CON-IR-7d) and may indicate a positive
role of Notch signaling in the repair phase of myocardial IR
process.
Tail-suspended rat models have been widely used to
simulate microgravity to study the influence of weightlessness on
the cardiovascular system (17).
In the present study, increased Notch1 expression was observed
following myocardial IR injury in the border zone compared with the
remote zone and infarct zone of tail-suspended rats. However,
Notch1 expression levels were reduced in the tail-suspension group
compared with the control group. It may be hypothesized that the
normal function of Notch1 in protection against myocardial IR
injury may be interrupted when exposed to simulated microgravity;
however, further in vivo studies are required.
A link between the function of the infarcted heart
and the decreased expression of Notch1 under simulated microgravity
was also examined. According to Dobaczewski et al (16), the rat models that underwent 3 h of
reperfusion (IR-3h) were used to represent the situation without
repair responses to myocardial IR injury. The IR-3h groups served
as the control for comparing the extent of myocardial reparation at
7 d after reperfusion. The results indicated that the infarcted
area was significantly decreased in rats that underwent 7 days of
reperfusion compared with those who underwent only 3 h of
reperfusion. Additionally, the infarction area at time points
between 3 h and 7 days, including 6, 12 h, 1 and 4 days, and even 1
h and 14 days (unpublished data) was evaluated. Among them, the
damage of the rat heart (infarcted area) peaked at 3 h following IR
injury, when the heart was still in the inflammatory phase. Later,
the infarction area gradually decreased and it finally stabilized
at 7 d after reperfusion (Jiang et al, unpublished data). In
agreement with the immunohistochemical results, which demonstrated
that Notch1 expression was decreased in tail-suspended rats
(CON-IR-7d), the myocardial infarct size was significantly
increased in tail-suspended rats (SUS-IR-7d) compared with the
control rats (CON-IR-7d), which provided further evidence for the
presumption that the proper function of Notch signaling pathway was
hampered under simulated microgravity.
However, this study has few limitations. First,
myocardial Notch signaling pathway mainly involves JAG1, Notch1 and
several downstream target genes, including HES1, HEY1 and cyclin D
(40,41). In the present study, the main focus
was on the distribution of Notch1 expression, and therefore
proteins involved in the intracellular action of signaling were not
evaluated. Furthermore, the presented data mainly provide
morphological evidence that validates the antibody choice and
depicts the distribution of Notch. Additional experiments by
knockdown or overexpression of Notch1 should be conducted to
confirm whether Notch1 signaling is dependent upon simulated
microgravity and to what extent simulated microgravity hampers the
protective effects of Notch signaling against myocardial IR.
In conclusion, by establishing and validating a
tail-suspended rat model with myocardial IR, novel features of
Notch1 expression in the repair phase of border zone were observed.
The present data confirmed that Notch1 and its endogenous ligand
JAG1 are poorly expressed in normal adult heart. In addition, it
was demonstrated that there is significantly higher Notch1
expression in border zone compared to infarct zone and remote zone
following myocardial IR. Notably, it was observed for the first
time that simulated microgravity may jeopardize the normal function
of Notch1 in the fight against myocardial IR. These data may
improve our knowledge of the underlying mechanisms of ischemic
heart disease in weightless environment, but further studies
evaluating the role of Notch1 in myocardial IR injury and the
reduction of cardiac ischemic risk in astronauts are needed.
Acknowledgements
This study was supported by The National Natural
Science Foundation of China (grant no. 81571844). We would also
like to thank Mr. Cheng-Fei Li (Department of Aerospace
Biodynamics, The Fourth Military Medical University, Xi'an, China)
for the kind help with Western blot detection.
Glossary
Abbreviations
Abbreviations:
|
BSA
|
bovine serum albumin
|
|
CHD
|
coronary heart disease
|
|
CON
|
control
|
|
CON-IR-3h
|
3-h reperfusion without suspension
|
|
CON-IR-7d
|
7-day reperfusion without
suspension
|
|
Cx43
|
connexin 43
|
|
IR
|
ischemia-reperfusion
|
|
LAD
|
left anterior descending coronary
artery
|
|
NF-κB
|
nuclear transcription factor-κB
|
|
NICD
|
Notch intracellular domain
|
|
PBS
|
phosphate buffered saline
|
|
PVDF
|
polyvinylidene fluoride
|
|
SD
|
Sprague-Dawley
|
|
SEM
|
standard error of the mean
|
|
SUS
|
tail-suspended
|
|
SUS-IR-3h
|
3-h reperfusion following 4-week
suspension
|
|
SUS-IR-7d
|
7-day reperfusion following 4-week
suspension
|
|
TBS
|
Tris-buffered saline
|
|
TTC
|
triphenyltetrazolium chloride
|
|
WGA
|
wheat germ agglutinin
|
References
|
1
|
World Health Organization, . World Health
Statistics 2016-Monitoring health for the SDGs. Sustainable
development goals. http://www.who.int/gho/publications/world_health_statistics/2016/EN_WHS2016_TOC.pdfApril
16–2016
|
|
2
|
Ibáñez B, Heusch G, Ovize M and Van de
Werf F: Evolving therapies for myocardial ischemia/reperfusion
injury. J Am Coll Cardiol. 65:1454–1471. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
White RJ and Blomqvist CG: Central venous
pressure and cardiac function during spaceflight. J Appl Physiol
(1985). 85:738–746. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu YM, Jiao B, Lee J, Zhang L and Yu ZB:
Simulated microgravity increases myocardial susceptibility to
ischemia-reperfusion injury via a deficiency of AMP-activated
protein kinase. Can J Physiol Pharmacol. 95:59–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wharton KA, Johansen KM, Xu T and
Artavanis-Tsakonas S: Nucleotide sequence from the neurogenic locus
notch implies a gene product that shares homology with proteins
containing EGF-like repeats. Cell. 43:567–581. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Niessen K and Karsan A: Notch signaling in
cardiac development. Circ Res. 102:1169–1181. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wiese C, Heisig J and Gessler M: Hey bHLH
factors in cardiovascular development. Pediatr Cardiol. 31:363–370.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ferrari R and Rizzo P: The Notch pathway:
A novel target for myocardial remodelling therapy? Eur Heart J.
35:2140–2145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu HC, Qin HY, He F, Wang L, Fu W, Liu D,
Guo FC, Liang L, Dou KF and Han H: Canonical notch pathway protects
hepatocytes from ischemia/reperfusion injury in mice by repressing
reactive oxygen species production through JAK2/STAT3 signaling.
Hepatology. 54:979–988. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Croquelois A, Domenighetti AA, Nemir M,
Lepore M, Rosenblatt-Velin N, Radtke F and Pedrazzini T: Control of
the adaptive response of the heart to stress via the Notch1
receptor pathway. J Exp Med. 205:3173–3185. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Boccalini G, Sassoli C, Formigli L, Bani D
and Nistri S: Relaxin protects cardiac muscle cells from
hypoxia/reoxygenation injury: Involvement of the Notch-1 pathway.
FASEB J. 29:239–249. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pei H, Song X, Peng C, Tan Y, Li Y, Li X,
Ma S, Wang Q, Huang R, Yang D, et al: TNF-α inhibitor protects
against myocardial ischemia/reperfusion injury via Notch1-mediated
suppression of oxidative/nitrative stress. Free Radic Biol Med.
82:114–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pei H, Yu Q, Xue Q, Guo Y, Sun L, Hong Z,
Han H, Gao E, Qu Y and Tao L: Notch1 cardioprotection in myocardial
ischemia/reperfusion involves reduction of oxidative/nitrative
stress. Basic Res Cardiol. 108:3732013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Greenwald I and Seydoux G: Analysis of
gain-of-function mutations of the lin-12 gene of Caenorhabditis
elegans. Nature. 346:197–199. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rebay I, Fleming RJ, Fehon RG, Cherbas L,
Cherbas P and Artavanis-Tsakonas S: Specific EGF repeats of Notch
mediate interactions with Delta and Serrate: Implications for Notch
as a multifunctional receptor. Cell. 67:687–699. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dobaczewski M, Gonzalez-Quesada C and
Frangogiannis NG: The extracellular matrix as a modulator of the
inflammatory and reparative response following myocardial
infarction. J Mol Cell Cardiol. 48:504–511. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu ZB, Zhang LF and Jin JP: A proteolytic
NH2-terminal truncation of cardiac troponin I that is up-regulated
in simulated microgravity. J Biol Chem. 276:15753–15760. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao E, Lei YH, Shang X, Huang ZM, Zuo L,
Boucher M, Fan Q, Chuprun JK, Ma XL and Koch WJ: A novel and
efficient model of coronary artery ligation and myocardial
infarction in the mouse. Circ Res. 107:1445–1453. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Reimer KA, Lowe JE, Rasmussen MM and
Jennings RB: The wavefront phenomenon of ischemic cell death. 1.
Myocardial infarct size vs. duration of coronary occlusion in dogs.
Circulation. 56:786–794. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rutherford SL, Trew ML, Sands GB, LeGrice
IJ and Smaill BH: High-resolution 3-dimensional reconstruction of
the infarct border zone: Impact of structural remodeling on
electrical activation. Circ Res. 111:301–311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu ZB, Gao F, Feng HZ and Jin JP:
Differential regulation of myofilament protein isoforms underlying
the contractility changes in skeletal muscle unloading. Am J
Physiol Cell Physiol. 292:C1192–C1203. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maass K, Chase SE, Lin X and Delmar M:
Cx43 CT domain influences infarct size and susceptibility to
ventricular tachyarrhythmias in acute myocardial infarction.
Cardiovasc Res. 84:361–367. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Herault S, Fomina G, Alferova I,
Kotovskaya A, Poliakov V and Arbeille P: Cardiac, arterial and
venous adaptation to weightlessness during 6-month MIR spaceflights
with and without thigh cuffs (bracelets). Eur J Appl Physiol.
81:384–390. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Grego-Bessa J, Luna-Zurita L, del Monte G,
Bolós V, Melgar P, Arandilla A, Garratt AN, Zang H, Mukouyama YS,
Chen H, et al: Notch signaling is essential for ventricular chamber
development. Dev Cell. 12:415–429. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xue Y, Gao X, Lindsell CE, Norton CR,
Chang B, Hicks C, Gendron-Maguire M, Rand EB, Weinmaster G and
Gridley T: Embryonic lethality and vascular defects in mice lacking
the Notch ligand Jagged1. Hum Mol Genet. 8:723–730. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nemir M and Pedrazzini T: Functional role
of Notch signaling in the developing and postnatal heart. J Mol
Cell Cardiol. 45:495–504. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gude NA, Emmanuel G, Wu W, Cottage CT,
Fischer K, Quijada P, Muraski JA, Alvarez R, Rubio M, Schaefer E
and Sussman MA: Activation of Notch-mediated protective signaling
in the myocardium. Circ Res. 102:1025–1035. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou XL, Wan L, Xu QR, Zhao Y and Liu JC:
Notch signaling activation contributes to cardioprotection provided
by ischemic preconditioning and postconditioning. J Transl Med.
11:2512013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Köhler C, Bell AW, Bowen WC, Monga SP,
Fleig W and Michalopoulos GK: Expression of Notch-1 and its ligand
Jagged-1 in rat liver during liver regeneration. Hepatology.
39:1056–1065. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Louis AA, Van Eyken P, Haber BA, Hicks C,
Weinmaster G, Taub R and Rand EB: Hepatic jagged1 expression
studies. Hepatology. 30:1269–1275. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cleutjens JP, Kandala JC, Guarda E,
Guntaka RV and Weber KT: Regulation of collagen degradation in the
rat myocardium after infarction. J Mol Cell Cardiol. 27:1281–1292.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gaggar A, Jackson PL, Noerager BD,
O'Reilly PJ, McQuaid DB, Rowe SM, Clancy JP and Blalock JE: A novel
proteolytic cascade generates an extracellular matrix-derived
chemoattractant in chronic neutrophilic inflammation. J Immunol.
180:5662–5669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Spinale FG: Myocardial matrix remodeling
and the matrix metalloproteinases: Influence on cardiac form and
function. Physiol Rev. 87:1285–1342. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sottile J and Hocking DC: Fibronectin
polymerization regulates the composition and stability of
extracellular matrix fibrils and cell-matrix adhesions. Mol Biol
Cell. 13:3546–3559. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shimazaki M, Nakamura K, Kii I, Kashima T,
Amizuka N, Li M, Saito M, Fukuda K, Nishiyama T, Kitajima S, et al:
Periostin is essential for cardiac healing after acute myocardial
infarction. J Exp Med. 205:295–303. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hinz B: Formation and function of the
myofibroblast during tissue repair. J Invest Dermatol. 127:526–537.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tomasek JJ, Gabbiani G, Hinz B, Chaponnier
C and Brown RA: Myofibroblasts and mechano-regulation of connective
tissue remodelling. Nat Rev Mol Cell Biol. 3:349–363. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Albrecht-Schgoer K, Schgoer W, Holfeld J,
Theurl M, Wiedemann D, Steger C, Gupta R, Semsroth S,
Fischer-Colbrie R, Beer AG, et al: The angiogenic factor
secretoneurin induces coronary angiogenesis in a model of
myocardial infarction by stimulation of vascular endothelial growth
factor signaling in endothelial cells. Circulation. 126:2491–2501.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
MacArthur JW Jr, Purcell BP, Shudo Y,
Cohen JE, Fairman A, Trubelja A, Patel J, Hsiao P, Yang E, Lloyd K,
et al: Sustained release of engineered stromal cell-derived factor
1-alpha from injectable hydrogels effectively recruits endothelial
progenitor cells and preserves ventricular function after
myocardial infarction. Circulation. 128 11 Suppl 1:S79–S86. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bray SJ: Notch signalling: A simple
pathway becomes complex. Nat Rev Mol Cell Biol. 7:678–689. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Miele L: Notch signaling. Clin Cancer Res.
12:1074–1079. 2006. View Article : Google Scholar : PubMed/NCBI
|