Introduction
Non-mammalian vertebrates are able to regenerate
auditory and vestibular hair cells following trauma to directly
improve the functional recovery of hearing and balance (1,2).
However, in mammals, there is a limited capacity for spontaneous
hair cell regeneration in the vestibular system and no spontaneous
hair cell regeneration has been observed in the mature cochlea.
Therefore, sensory hair cell damage is a major cause of mammalian
hearing loss and balance disorder (1,3).
The development of a functionally mature cochlea
requires the engagement of a number of signaling pathways,
including Notch, bone morphogenetic protein and Wnt. The Notch
signaling pathway mediates hair and supporting cell differentiation
during the later period of inner ear development, when Notch1, hes
family basic helix-loop-helix transcription factor (Hes)1 and Hes5
are expressed in supporting cells (4–7).
Therefore, it is clear that Notch signaling is crucial for inner
ear differentiation and typical mosaic pattern formation. In mature
mammals, hair cell regeneration may fail if supporting cell
differentiation is inhibited. Hartman et al (8) and Murata et al (9) demonstrated that Notch signaling
molecules were activated in a drug-damaged cochlea mouse model.
Therefore, the Notch signaling pathway may be a climacteric pathway
for the regeneration of hair cells and the dedifferentiation of
supporting cells.
A previously identified microRNA (miR), miR-183, may
have an important role in inner ear development and function
(10). It has been previously
demonstrated that during sensory epithelial differentiation,
miR-183 is expressed in hair cells, whereas Notch1 and Hes1 are
primarily expressed in supporting cells (9,11).
The spatially exclusive expression pattern of miR-183 and Notch1
during inner ear development suggests a potential association
between miR-183 and Notch signaling.
In the current study, gentamicin-treated cells had
significantly reduced the number of myosin VI-positive hair cells
in the post-neonatal mice explanted cochlear. Notch1 signaling in
the supporting cells was also increased. Inhibition of Notch
signaling by DAPT attenuated the gentamicin-induced hair cell loss.
Conversely, the expression of the miR-183 cluster was downregulated
following gentamicin treatment. This downregulation may be reversed
by DAPT. It is of note, the increase in myosin VI-positive cells
induced by DAPT was abolished by miR-183 inhibition.
Materials and methods
Animals
Post-natal day 1 (P1) C57BL/6 mice (n=480; average
weight 1.0 g) were obtained from the Experimental Animal Center of
Sun Yat-sen University (Guangzhou, China). The study protocol was
approved by the Institution Review Board of Sun Yat-sen University
(Guangzhou, China). All animal experiments were performed within
2–3 h of the arrival of the mice and in compliance with the
guidelines of the Animal Care and Use Committee of the National
Institutes of Health of USA for experimental use of laboratory
animals.
Organ and cell culture
Hank's balanced salt solution (HBSS, pH 7.4),
supplements N2 (100×) and B27 (50×), Dulbecco's modified Eagle's
medium/F12 (DMEM/F12) were purchased from Invitrogen (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Collagen-coated cover slides,
penicillin G, heparin sulfate, and bromodeoxyuridine (BrdU) were
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
C57BL/6 mice were euthanized at postnatal day 1 and cochlear
sensory epithelium was collected and dissected in HBSS. The stria
vascularis, Reissner's membrane and the tectorial membrane were
removed prior to transfer onto the collagen-coated cover slides.
One group of organ samples from 20 mice were incubated in
serum-free DMEM/F12 media supplemented with N2, B27 and 100 U/ml
penicillin G. Culture medium was changed every other day. Following
8 days culture the incubated cochleae were then fixed with 4%
paraformaldehyde at room temperature for 30 min.
The inner ear sensory epithelial sheets were
isolated from the saccule and utricle of C57BL/6 mice. The otolith
was carefully dissected under a stereoscopic microscope in a
separate dish with ice-cold HBSS. The isolated inner ear sensory
epithelial sheets were transferred into Eppendorf tubes, digested
in 500 µl of 0.125% trypsin in phosphate-buffered saline (PBS;
Gibco; Themo Fisher Scientific, Inc.) at 37°C for 15 min. The cells
were carefully triturated with plastic 200 µl pipette tips,
centrifuged (3,000 × g, 5 min at room temperature) and suspended in
2 ml DMEM/F12 medium with N2 and B27 supplements, epidermal growth
factor (EGF; 20 ng/ml; Invitrogen; Thermo Fisher Scientific, Inc.),
insulin-like growth factor 1 (IGF-1, 20 ng/ml, PeproTech, Rocky
Hill, NJ, USA), basic fibroblast growth factor (bFGF; 20 ng/ml,
R&D Systems, Minneapolis, MN, USA). The dissociated cells were
passed through a 70 µm cell filter (BD Biosciences, Franklin Lakes,
NJ, USA) to remove cell clumps. Half of the medium was exchanged
every other day. The solid spheres were collected after 5 days of
culture, transferred into chamber slides coated with Matrigel™ (BD
Biosciences), and allowed to cultivated up to 11 days in the same
medium without growth factors. The inner ear sensory precursor
cells were fixed with 4% paraformaldehyde at room temperature for
30 min.
Drug treatment
In order to induce injury in hair cells, the
isolated organs were incubated with 150 µM gentamicin (Shanghai
DingGuo Biotech Co., Ltd., Shanghai, China) for 14 h. DAPT (5 µM,
D5942; Sigma-Aldrich; Merck KGaA) or dimethyl sulfoxide (DMSO; 15
µM; Invitrogen; Thermo Fisher Scientific, Inc.; negative control)
were subsequently added to substitute for gentamincin. The cochleae
were fixed after 3.5 days for sex determining region Y-box 2 (Sox2;
SAB2104660; 1:250; Sigma-Aldrich; Merck KGaA), anti-active Notch1
(1:200; ab8925), 4′6-diamidino-2-phenylindole (DAPI; 1:1,000;
ab104139; both Abcam, Cambridge, UK) and myosin VI (1:200; 25-6791;
rabbit polyclonal, Proteus Biosciences Inc., Ramona, CA, USA)
staining. Organs were cultured for 8 days for myosin VI, active
Notch1, anti-bromodeoxyuridine (anti-BrdU; 1:200; Sigma-Aldrich;
Merck KGaA) Rhodamine-phalloidin (1:200; Invitrogen; Thermo Fisher
Scientific, Inc.), and DAPI staining. For proliferation analysis,
BrdU was added at a final concentration of 10 µM following washes
to remove gentamycin. The organs with BrdU and the control group
were cultured for 3.5 days following the washing out of the
gentamicin prior to staining. Inner ear precursor cells were
incubated with 50 µM DAPT as a negative control reagent, with or
without small interfering RNA (siRNA) transfection of the miR-183
family (miR-183, miR-182, miR-96) by electroporation 5 days
post-culture. The inner ear precursor cells continued to be
cultured for 5 days.
Immunohistochemistry and
microscopy
Immunostaining of the cultured organs of Corti were
performed as previously described (12) using the following primary
antibodies: Myosin VI, mouse monoclonal anti-BrdU, mouse monoclonal
anti-active Notch1 and rabbit polyclonal anti-Sox2 (SAB2104660;
1:250; Sigma-Aldrich; Merck KGaA; all overnight at 4°C). The
following secondary antibodies were used: Alexa Fluor®
488-conjugated and Alexa Fluor® 594-conjugated donkey
anti-rabbit, Alexa Fluor® 488-conjugated and Alexa
Fluor® 594-conjugated donkey anti-mouse (1:400; R37118,
R37119, R37114, R37115 respectively; Invitrogen; Thermo Fisher
Scientific, Inc.). Following the fixation of the cultured organs,
they were permeabilized in 0.1% Triton X-100 in PBS for 15 min and
washed three times for 5 min in PBS. The organs were blocked in 5%
bovine serum albumin (AR1006; Boster Biological Technology, Ltd.,
Wuhan, China)/0.05% Triton X-100 in PBS for 1 h prior to incubation
with the primary antibodies overnight at 4°C. Incubation with the
secondary antibodies (1:400) was performed at 4°C overnight or at
room temperature for 2 h. Rhodamine-phalloidin was used to stain
the stereocilium and reticular laminae of the cochlear hair cells
for 30 min in room temperature. DAPI was used to stain the nucleus
for 15 min at room temperature. Specimens were mounted and observed
under a fluorescence microscope (Olympus IX71 Inverted Microscope;
Olympus Corporation, Tokyo, Japan) and a FluoView FV1000 confocal
microscope (Olympus Inc., Guangzhou, Guangdong, China). Specimens
were examined at ×20 or ×40 magnification. Images were processed
using Image-Pro Plus software version 6.0 (Media Cybernetics, Inc.,
Rockville, MD, USA). Cultured precursor sensory epithelia were
prepared for staining and fixed in 4% PFA/PBS for 20 min at room
temperature and then washed in PBS three times. Subsequently,
permeabilization, blocking and antibody incubation were performed
as described for organ samples.
For organ culture, the number of hair cells was
determined by calculating the number of myosin VI-positive cells
per 200 µm in the organ of Corti. For cultured precursor sensory
epithelia, the percentage of hair cells was determined by
calculating the number of myosin VI-positive cells over the total
number of cells in the whole sphere, multiplied by 100. Cells
positive for DAPI or myosin VI were counted in one representative
optical section. Cells were only counted when their nucleus
comprised >50% of the cell area. Quantification was obtained
from at least 6 random areas over the length of an entire cochlea.
At least 5 or 6 inner ear precursor spheres were counted for each
experiment.
Reverse transcription
semi-quantitative polymerase chain reaction (RT-sqPCR)
analysis
The cultured organs (at day 8) were carefully
dissected and collected in Eppendorf tubes. The total RNA was
extracted and column-purified using RNeasy purification kit (Qiagen
GmbH, Hilden, Germany) according to the manufacturer's protocol.
Total RNA was treated with RNase-free DNase (Roche Diagnostics,
Indianapolis, IN, USA). Reverse transcription was performed using
SuperScript III one-step RT-PCR system with Platinum Taq High
Fidelity DNA polymerase (1274-030; Invitrogen; Thermo Fisher
Scientific, Inc.) according to the protocol provided by
manufacturer. The PCR sequential steps are as follows: Denaturation
at 94°C for 2 min, annealing at 60°C for 30 sec, and extension at
72°C for 45 sec: 35 cycles were performed and the last
amplification was followed by a final 10-min elongation step at
72°C. The primers (Invitrogen; Thermo Fisher Scientific, Inc.) used
for PCR are presented in Table I.
The PCR products were resolved by 1% agarose gel electrophoresis
containing 0.4 µg/ml of ethidium bromide. The level of gene
expression was semi-quantified by grey scale ratio of Atoh1,
Notch1, Jagged1, Delta1, Hes1, P27 vs. GAPDH by Quantity One
(version 4.6.2; Bio-Rad Laboratories, Inc. Hercules, CA, USA). Each
experiment was performed (containing at least four different
cochleae) at least three times from each group.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sense (5′-3′) | Antisense
(5′-3′) |
|---|
| Atoh1 | AGATCT
ACATCAACGCTCTGTC |
ACTGGCCTCATCAGAGTCACTG- |
| Hes1 |
GGAGAGGCTGCCAAGGTTTT |
GCAAATTGGCCGTCAGGA |
| Notch1 |
GCTGACCTGCGCATGTCTGCCATG |
CATGTTGTCCTGGATGTTGGCATCTG |
| Jagged1 |
CAGAATGACGCCTCCTGTCG |
TGCAGCTGTCAATCACTTCG |
| Delta1 |
TACTGCACTGACCCAATCTG |
GGCACCGTTAGAACAAGG |
| p27 |
CTGGAGCGGATGGACGCCAGAC |
CGTCTGCTCCACAGTGCCAGC |
| GAPDH |
AACGGGAAGCCCATCACC |
CAGCCTTGGCAGCACCAG |
RT-quantitative (q)PCR (RT-qPCR)
analysis of miR-183 family expression
RT-qPCR was performed based on a stem-loop Rtprimer
design, as previously described (13). The cDNA was synthesized from
extracted total RNA from cultured organs as mentioned above.
Quantitative PCR (qPCR) was performed using 2 µl of SYBR-Green/ROX
qPCR Master Mix (Suzhou GenePharma Co., Ltd., Suzhou, China). All
procedures were subjected to a TaqMan miRNA assay (Applied
Biosystem 7500) according to the manufacturer's instructions. U6
was employed as an endogenous control to normalize data using the
2−ΔΔCq (14)
method.
Probe and primers specific for miR-183, miR-182,
miR-96, or for U6 RNA internal control were purchased from LC
Sciences (Hangzhou, China). RT-qPCR analyses of at least 3
independent cultures were performed.
The qPCR reaction was performed using an ABI 7500
sequence detection system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) and used the following PCR protocol: 95°C for 10
min followed by 40 cycles of 95°C for 10 sec and 55°C for 30
sec.
In situ hybridization
In situ hybridization procedure was performed
as previously described (15). The
cochleae explants were cultured in vitro for 8 days and
subsequently fixed in fresh 4% paraformaldehyde at room temperature
for 1–2 h. Then the slides of cochleae were washed for 3×5 min in
1XPBS at room temperature. Followed proteinase K treatment, the
slides were rinsed in 1XPBS at room temperature and placed
horizontally in the hybridization chamber with 70 µl of
hybridization buffer. Cochleae were prehybridized by incubating for
4–8 h at room temperature. Next, the cochleae were carefully
pipetted with probed hybridization buffer and hybridized at 50–60°C
overnight. The next day, the slides were soaked in prewarmed 60°C
5XSSC and then incubated in 0.2XSSC at 60°C for 1 h. Finally, the
slides were incubated in B1 solution (0.1 M Tris pH 7.5/0.15 M
NaCl) at room temperature for 10 min and then blocked in blocking
solution (2 ml of fetal calf serum, 18 ml of B1 and 0.1% Tween-20)
for 1 h at room temperature. The slides were carefully pipetted
with blocking solution containing anti-DIG-alkaline phosphatase
antibody (diluted 1:1,000), followed by incubation at 4°C
overnight. The next day, the slides were washed three times for 5
min each time in B1 solution at room temperature. Then, cochleae
were equilibrated for 10 min in B3 solution (0.1 M Tris pH 9.5/0.1
M NaCl/50 mM MgCl2). Finally, the developer solution
(3.4 µl of 100 mg/ml NBT, 3.5 µl of 50 mg/ml BCIP, 2.4 µl of 24
mg/ml levamisole, 5 µl of 10% Tween and 986 µl of B3) was carefully
added to the slides, which were kept in a humidified chamber. The
reaction was developed at room temperature in the dark for times
ranging from 1–3 days, depending on the microRNA expression levels.
A light microscope (Leica Microsystems GmbH, Wetzlar, Germany) was
used to monitor the reaction, which was terminated when a strong
blue stain was observed. Washing the slides for 3×10 min in 1XPBT
terminated the color reaction. The slides were mounted in
fluorescent mounting media (S3023; Dako; Agilent Technologies,
Inc., Santa Clara, CA, USA) for further observation.
Electroporation
The siRNA oligoribonucleotides for the miR183 family
(miR-183, miR-182, miR-96) were purchased from Shanghai GenePharma
Co., Ltd., (Shanghai, China), cultured inner ear precursor cells
were dissociated into a single-cell suspension with 0.25% trypsin
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C for 3 min. Then,
1×106 cells/well were transfected by electroporation
with 5 µg miR-183 siRNA oligoribonucleotides, according to the
manufacturer's protocol. An enhanced green fluorescent protein RNA
oligoribonucleotide (Shanghai GenePharma Co., Ltd.) was used to
monitor the efficiency of the electroporation. The transfected
inner ear precursor cells were plated on cover glasses coated with
poly-L-lysine and laminin (R&D Systems, Inc.) and fixed with 4%
paraformaldehyde at room temperature for 30 min following an
additional 5 days culture. The inner ear precursor spheres with the
average diameter of 55 µm were selected for statistical
evaluation.
Statistical analysis
Data are expressed as the mean ± standard deviation
and analyzed using SPSS version 13.0 (SPSS Inc., Chicago, IL, USA).
One-way analysis of variance and Tukey's Honest Significant
Difference was used as a post-hoc test in order to determine
significant differences in multiple comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
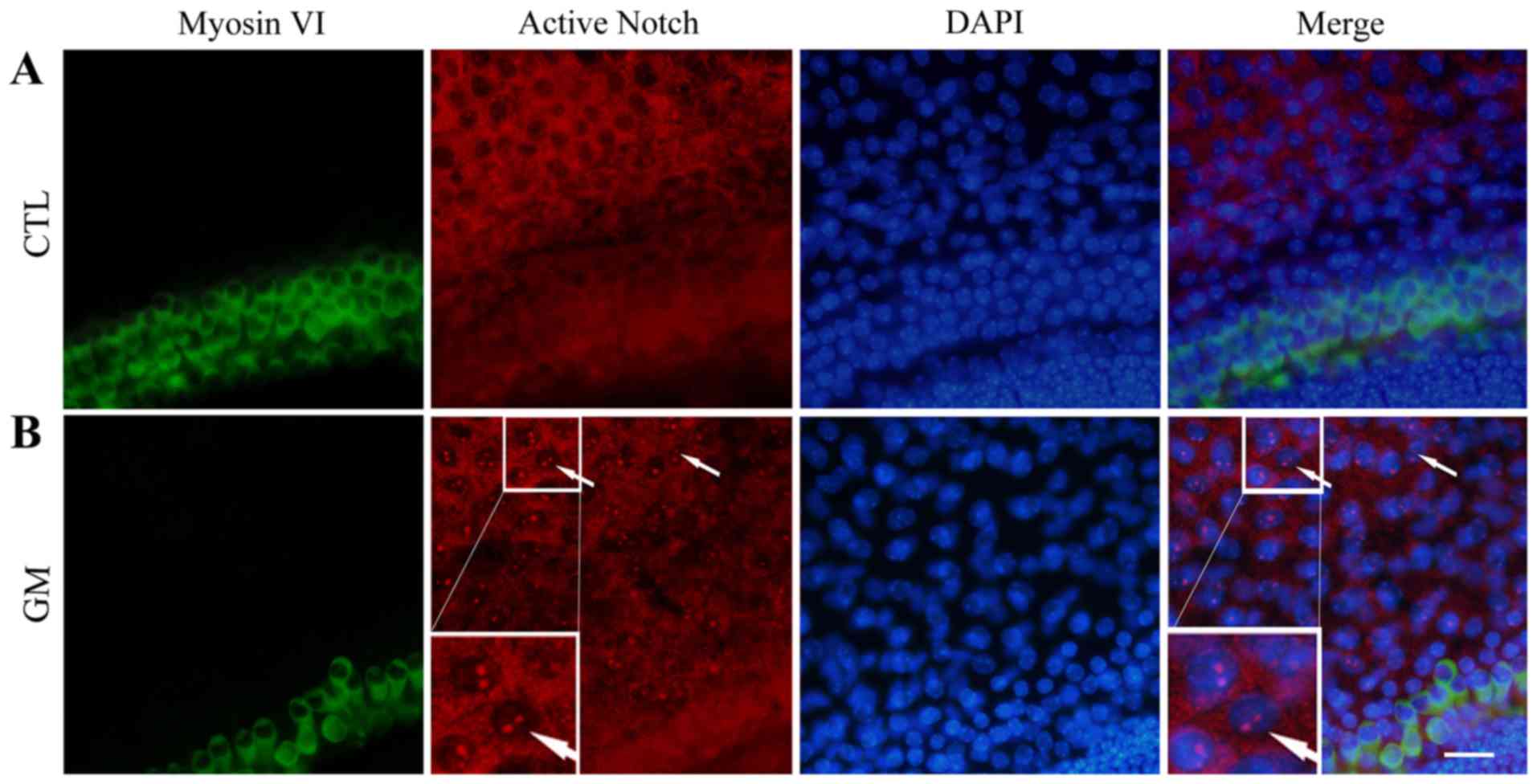
Gentamicin-induced injury of neonatal
cochlear hair cells activates Notch signaling
To investigate the expression of activated Notch in
hair and supporting cells, Notch1 expression was determined in
control and gentamycin-damaged neonatal cochlear explants. An
activated signal was not detected in control cochlear hair cells
and supporting cells (Fig. 1A).
However, fluorescence was detected in the nucleus of the supporting
cells in the gentamicin-damaged cochleae, indicating an activated
Notch signal (Fig. 1B).
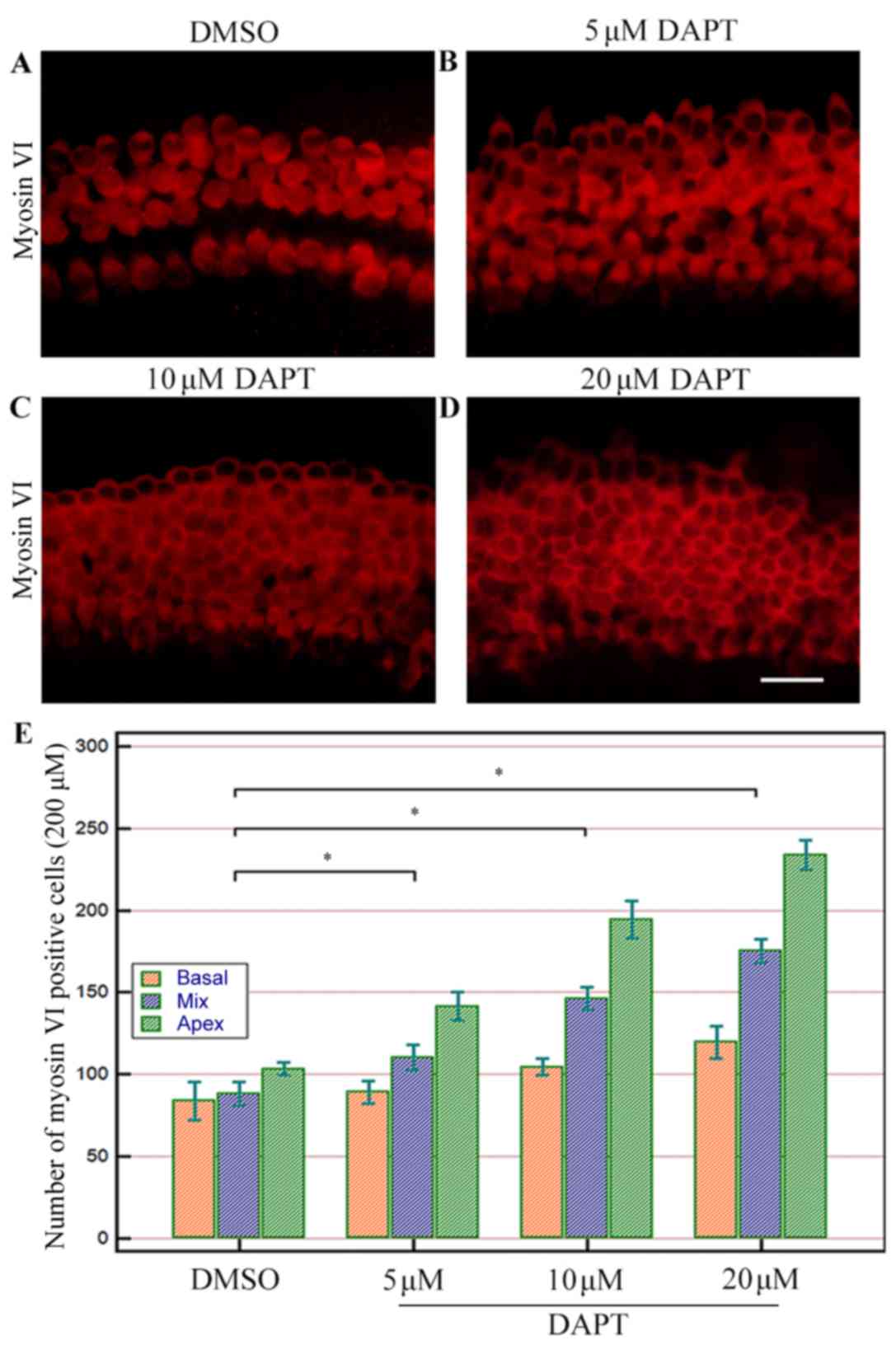
Inhibition of Notch expression
promotes postnatal cochlea hair cell differentiation and attenuates
gentamicin-induced hair cell loss
Notch inhibition by DAPT (5–20 µM) increased the
number of myosin VI-positive cells in cochlear explants (Figs. 2 and 3) in a dose-dependent manner, with the
most evident increase observed in the 20 µM DAPT group (P<0.05;
Fig. 2E). The majority of the
increase in hair cell number occurred at the apex turn, with
limited changes in the basal turn (Fig. 2E). The ability of DAPT stimulate
new hair cell growth in damaged postnatal cochlea was also
examined. The results revealed that gentamicin treatment reduced
the number of myosin VI-positive hair cells (Fig. 3C, G and J) compared with the
controls (Fig. 3A, E and I). This
reduction was mostly confined to the basal and middle turn of the
cochleae, with virtually no hair cell loss occurring in the apex
turn. Co-treatment of gentamicin and DAPT resulted in a significant
increase in the number of myosin VI positive cells (P<0.05;
Fig. 3D, H and L) compared with
gentamicin treatment alone. No hair cell loss was observed in the
gentamicin-treated apex turn of the cochleae. DAPT and gentamicin
treatment markedly increased the number of hair cells compared with
gentamicin treatment alone (Fig.
3M; P<0.05). Additionally, DAPT treatment and gentamincin
and DAPT co-treatment resulted in the blurring of the Corti's
tunnel between inner and outer hair cells and crowding in the apex
and middle turns due to new hair cell growth (Fig. 3B, D, F and H). Hair cells in the
sensory epithelium region displayed stereocilia with varying
degrees of disarrangement and random orientation (Fig. 3D, H and L).
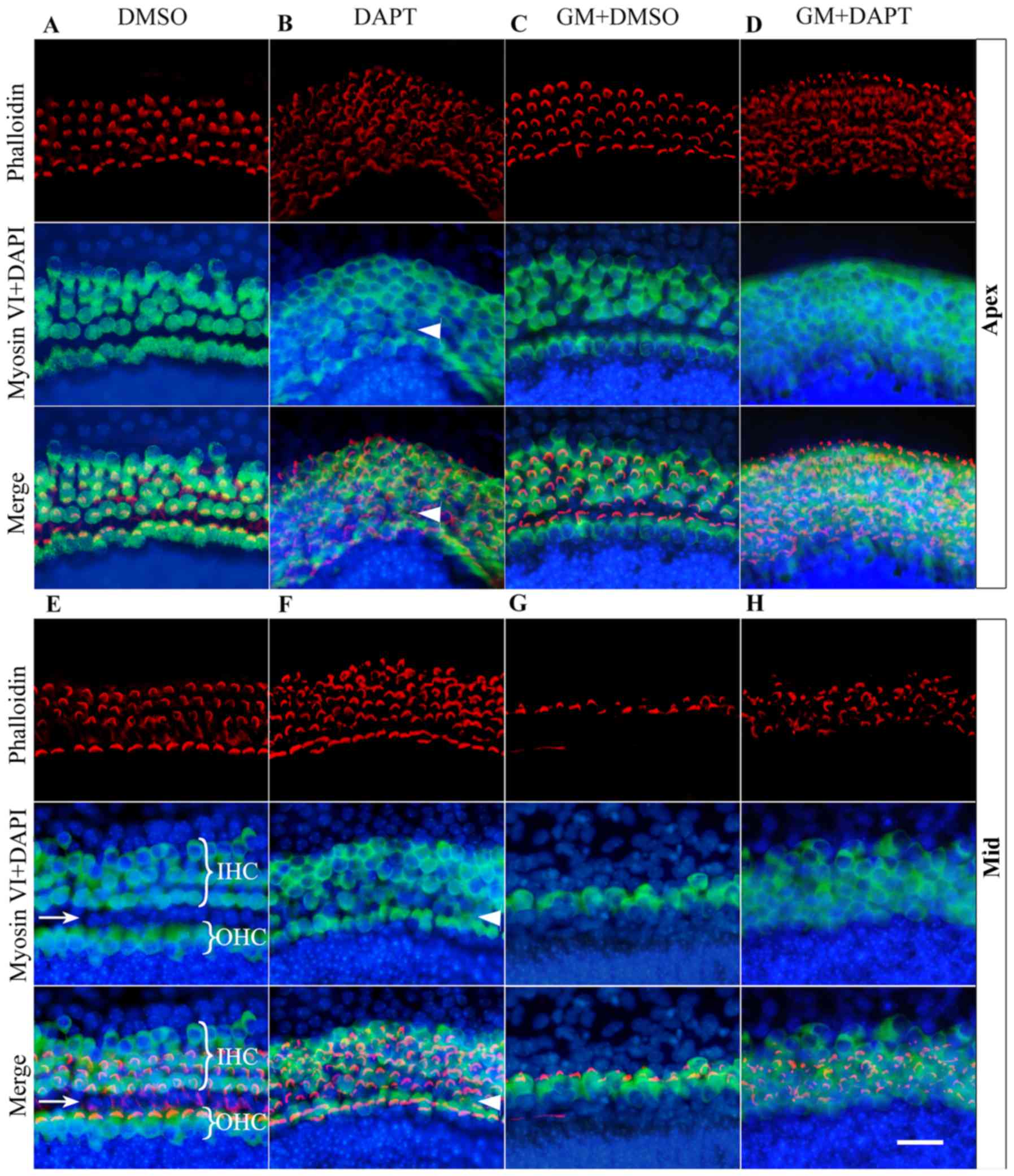 | Figure 3.Notch inhibition promoted hair cell
growth in normal and impaired cochleae. The apex turn of cochleae
of post-natal day 1 mice were treated with (A) DMSO, (B) 10 µM
DAPT, (C) 150 µM gentamicin and DMSO or (D) gentamicin and DAPT.
The middle turn of cochlear of post-natal day 1 mice were treated
with (E) DMSO, (F) 10 µM DAPT, (G) 150 µM gentamicin and DMSO or
(H) gentamicin and DAPT. The basal turn of cochleae of post-natal
day 1 mice were treated with (I) DMSO, (J) 10 µM DAPT, (K) 150 µM
gentamicin and DMSO or (L) gentamicin and DAPT. Cochleae were
immunostained with phalloidin antibodies (red) and anti-myosin VI
antibodies (green). Nuclei were stained blue with DAPI. White
brackets indicate the area of OHCs and IHCs. The white arrows
indicate the tunnel of Corti. New hair cells appeared in the area
of the sensory epithelia and occupied the tunnel of Corti region,
as indicated by the white arrowheads. (M) Number of myosin
VI-positive cells were counted. Data are presented as the mean ±
standard deviation, derived from at least 3 independent
experiments. *P<0.05. Scale bar, 20 µm. GM, gentamicin; DMSO,
dimethyl sulfoxide; DAPT, 24-diamino-5-phenylthiazole; IHC, inner
hair cell; OHC, outer hair cell. |
New hair cells are derived from
transdifferentiated supporting cells
Activated Notch was revealed to be confined to the
supporting cells of gentamicin-damaged cochleae (Fig. 1B). Thus, it was investigated
whether hair cell proliferation was induced in response to DAPT
treatment. The majority of BrdU-positive nuclei were observed in
the area near outer hair cell and thus may be Hensen cells,
according to the inner ear anatomy of mice. No BrdU and myosin VI
double-positive cells were identified under any treatment
conditions (Fig. 4A-D). Sox2, a
marker of supporting cells, and myosin VI co-expressing cells were
observed within the sensory epithelium in the DAPT and gentamicin
co-treatment group (Fig. 4E). This
suggested that the newly generated hair cells were derived from the
differentiated, but not proliferating, supporting cells.
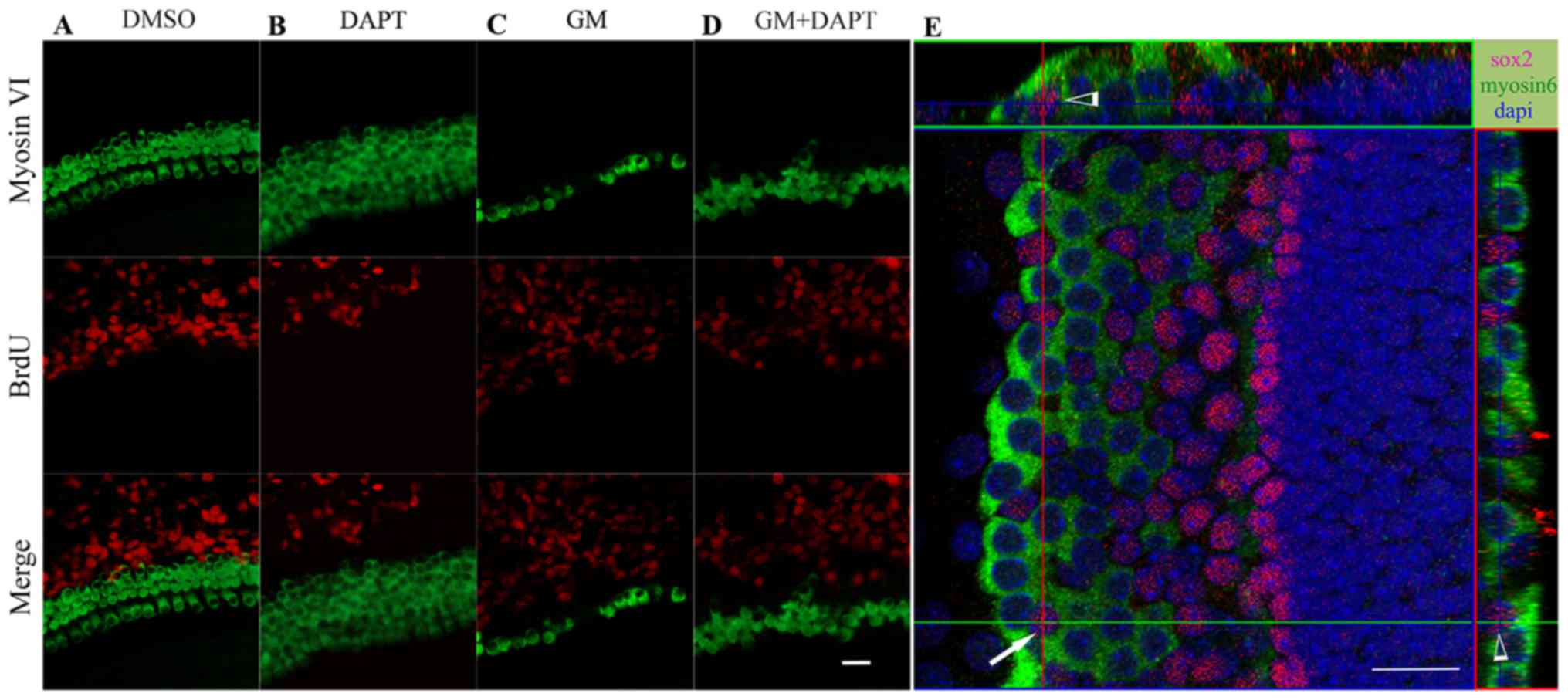 | Figure 4.New hair cells following Notch
inhibition are derived from supporting cell transdifferentiation,
but not proliferation. Cochleae of post-natal day 1 mice were
treated with (A) DMSO, (B) 10 µM DAPT, (C) 150 µM gentamicin or (D)
gentamicin and DAPT and were immunostained with myosin VI (green),
BrdU (red) and anti-Sox2 antibodies (purple). Nuclei were stained
blue with DAPI. BrdU positive nuclei were identified in supporting
cells, particularly in Hensen cells. No BrdU and myosin VI
co-expressing cells were identified. (E) Sox2 and myosin VI were
co-expressed (white arrow) in DAPT and gentamicin co-treatment
group, as indicated by the white arrow. The hollow arrowheads
indicate Sox2 and myosin VI co-expressing cells on the Y and Z
axis. Scale bar, 20 µm. GM, gentamicin; DMSO, dimethyl sulfoxide;
DAPT, 24-diamino-5-phenylthiazole; BrdU, bromodeoxyuridine; Sox2,
sex determining region Y-box 2. |
Notch signaling in the postnatal
cochlea following gentamicin and DAPT co-treatment
RT-sqPCR results revealed that gentamicin-induced
injury was associated with the upregulation of Notch1, Jagged1 and
Hes1 expression and downregulation of Atoh1 expression (Fig. 5A). The expression of Jagged1 and
Hes1 was downregulated in cochlear explants treated with DAPT. No
apparent changes were observed in Delta1 or p27 expression,
regardless of gentamicin or DAPT treatment. These results indicate
that DAPT efficiently downregulated Notch signaling.
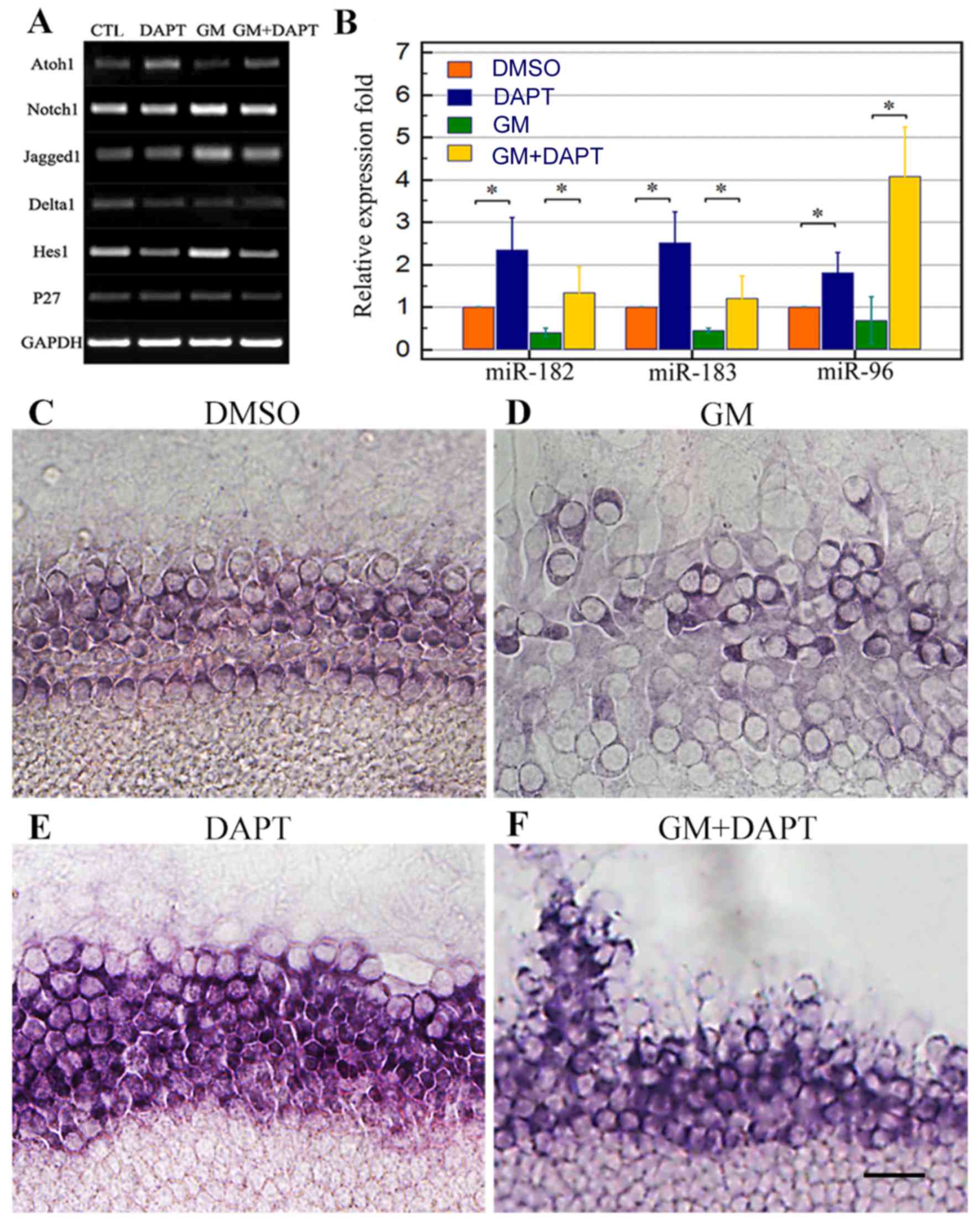 | Figure 5.DAPT treatment suppressed Notch
signaling in cochlear explants and stimulated miR-183 cluster
expression. (A) Cochleae of post-natal day 1 mice were treated with
DMSO, 10 µM DAPT, 150 µM gentamicin or gentamicin and DAPT.
Downstream Notch signaling targets, including Atoh1, Notch1,
Jagged1, Delta1, Hes1 and p27 were examined by RT-sqPCR. (B)
miR-183 cluster expression was examined by RT-qPCR. Data are
expressed as the mean ± standard deviation, derived from at least 3
independent experiments. *P<0.05. miR-183 expression was
detected by in situ hybridization in the cochleae of
post-natal day 1 mice treated with (C) DMSO, (D) 10 µM DAPT, (E)
150 µM gentamicin or (F) gentamicin and DAPT. DMSO, dimethyl
sulfoxide; DAPT, 24-diamino-5-phenylthiazole; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; RT-sqPCR,
reverse transcription-semiquantitative polymerase chain reaction;
miR, microRNA; Atoh1, atonal basic helix loop helix transcription
factor 1; Delta1, delta like canonical Notch ligand 1; Hes1, hes
family basic helix loop helix transcription factor 1; p27,
cyclin-dependent kinase inhibitor 1B; GM, gentamicin. |
DAPT attenuates the downregulation of
miR-183 family expression in gentamicin-damaged cochlear
explants
Previous studies revealed that the miR-183 family
(miR-183, miR-182, miR-96) is expressed in differentiated hair
cells in spatially exclusive pattern with Notch1 (9–11).
To investigate the association between the miR183 cluster and hair
cell differentiation and regeneration in damaged cochlea, the
expression of the miR183 cluster was examined in cochlea following
gentamicin-induced injury. The same pattern of miR-183 cluster
expression was observed in all groups (Fig. 5B). miR-183 cluster expression was
significantly downregulated (P<0.05) in gentamicin-treated
cochleae, whereas DAPT treatment significantly increased miR-183
cluster expression alone and in the presence of gentamicin
(Fig. 5B; P<0.05). Furthermore,
in situ hybridization with LNA probes revealed similar
results to the RT-qPCR analysis, demonstrating that DAPT treatment
markedly increased miR-183 cluster expression, with or without
gentamicin co-treatment (Fig.
5C-F). These findings suggest that the miR-183 cluster may play
a role in DAPT-induced hair cell differentiation and
regeneration.
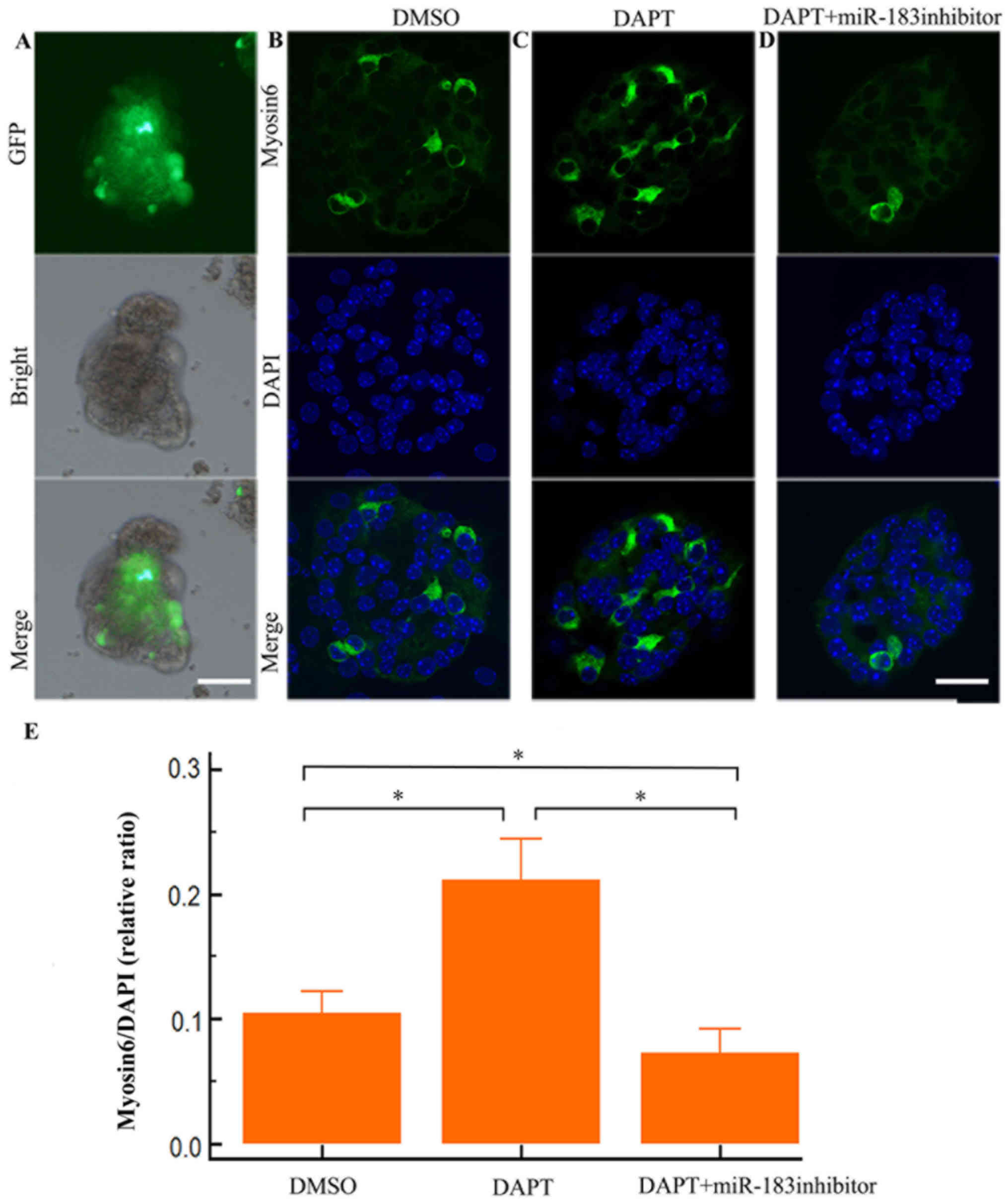
MiR-183 is required for hair cell
differentiation when the Notch signal is inhibited
Electroporation transfection efficiency was detected
by eGFP protein with fluorescence microscopy 24 h after eGFP
transfection (Fig. 6A). The
cultured organs of Corti were repeatedly electroporated with
miR-183 cluster inhibitors and eGFP fluorescence was not observed
24 h post-transfection. Therefore, inner ear precursor cells from
neonatal mice were cultured. Cultured inner ear precursor cells
were subsequently transfected with miR-183 cluster siRNA
oligoribonucleotides. Transfection efficiency was determined to be
40–50% by eGFP. After 5 days of DAPT treatment, the number of
myosin VI-positive cells was significantly increased (Fig. 6C) compared with controls
(P<0.05; Fig. 6B). However, the
number of myosin VI-positive cells observed in the inner ear
precursor cells pretreated with DAPT and miR-183 siRNA (Fig. 6D) was below that of the controls
(Fig. 6B; P<0.05), indicating
that the DAPT-induced increase in myosin VI-positive cell numbers
was abolished by miR-183 inhibition (Fig. 6, P<0.05). These results suggest
that miR-183 is required for hair cell differentiation in Notch
signaling-prohibited inner ear precursor cells.
Discussion
In mature mammals, hair cell regeneration may fail
if supporting cells are inhibited from dedifferentiating. The Notch
signaling pathway participates in the dedifferentiation of
supporting cells via multiple mechanisms (16–19).
In current study, the expression levels of activated Notch1,
Jagged1 and Hes1 were increased, whereas Atoh1 was decreased in
gentamicin-damaged cochleae. Activated Notch1 expression was also
identified to be confined to the supporting cells of
gentamicin-damaged cochleae. The new hair cells which were
identified when Notch1 was inhibited by DAPT were derived from the
supporting cells, rather than the mitosis or proliferation of
existing hair cells. These findings suggest that Notch signaling
plays an important role as a repressor of inner ear hair cell
regeneration following gentamicin-induced damage. It is believed
that hair cell regeneration in avian species occurs in two ways, by
quiescent supporting cells directly differentiating into new hair
cells or cells may divide first and subsequently differentiate into
new hair and supporting cells (1,6,20).
The BrdU incorporation assay in the present study revealed that no
BrdU expression was present in regenerated hair cells. Furthermore,
Sox2, a protein marker of supporting cells and myosin VI, a marker
of hair cells, were co-expressed in certain hair cell-like cells.
These findings support the hypothesis that regenerated hair cells
do not come from mitosis but originate from supporting cell
transdifferentiation in impaired mice cochlea in vitro. Hair
cells and supporting cells share common precursors during inner ear
development and avian hair cell regeneration comes from the
supporting cells (1,16). However, the specific regulatory
mechanisms underlying these processes are not clear. Hair cell
production in the tunnel of Corti following DAPT treatment in the
present study suggested that pillar cells may be latent precursor
cells. Pillar cells are a type of supporting cell which are found
adjacent to hair cells and may be able to easily substitute the
damaged hair cell following successful transdifferentiation.
Misaligned regenerated hair cells, disordered stereocilia and no
exact mosaic structure were identified in explanted cochlear
co-treated with gentamicin and DAPT. Additionally, the number of
regenerated hair cells seemed to be proportional to the number of
residual hair cells. Taken together, the results of the current
study indicate that multiple signaling pathways may be involved in
hair cell regeneration following cochlea injury. Future studies
should be focused on the interaction between multiple signaling
pathways that render structure formation and innervation between
hair cells, the cochlear nerve and functional hair cell
regeneration.
The miR-183 family regulates the sensorineural fates
in the inner ear and is important for hair cell maintenance and
survival (10). miR-183 expression
is upregulated during inner ear progenitor cell differentiation
into a hair cell-like fate. Ectopic miR-182 promotes inner ear
progenitor cell differentiation into a hair cell-like fate
(21). In the current study, Notch
signaling suppression with DAPT promoted the numbers of myosin VI
positive hair cells and upregulated miR-183 in cultured cochlea.
Similarly, an increase in hair cell numbers was also observed with
DAPT in inner ear precursor cells. Meanwhile, miR-183 siRNA was
demonstrated to abolish the increase in hair cells induced by DAPT.
No evident changes were detected in hair cells treated with miR-96
and miR-182 siRNA (data not shown). As Notch suppression may induce
the formation of novel hair cells through the transdifferentiation
of supporting cells, the aforementioned results suggested that
miR-183 may be involved in the transdifferentiation of Notch
signaling-inhibited hair cells. Furthermore, the activity of
miR-183 in DAPT-induced hair cell regeneration was examined. The
results revealed that Notch1 signaling was activated in
gentamicin-injured hair cells and was accompanied by a reduction of
miR-183 cluster expression. Suppression of Notch signaling by DAPT
not only inhibited Notch signaling activation, but also upregulated
miR-183 cluster expression which encouraged hair cell regeneration,
indicating that the miR-183 cluster may be involved in hair cell
regeneration induced by Notch suppression in gentamicin-treated
cochleae. miR-183 cluster expression decreased following
gentamicin-induced injury of the cochlea and was accompanied by
Notch signaling activation. DAPT inhibition of Notch attenuated
these effects. Additionally, miR-183 inhibition blocked the
increase in hair cells induced by DAPT. The Notch signal pathway
was not directly identified as a target of miR-183 in the present
study, but a previous study revealed that the miR-183 family is
associated with the Notch signaling pathway (22). Thus, further experiments to
determine the detailed mechanism of miR-183 in hair cell
differentiation are required. The present in vitro study
used cultured neonatal mouse cochlea and inner ear precursor to
investigate hair cell regeneration and differentiation. In
vitro study is simple and convenient but many factors,
including the influence of the environment and temperature are
quite different from an in vivo experiment. As the neonatal
cochlea, particularly the apex, still retains the ability to
differentiate into hair cells (16), the data from the present study may
not be strong enough to confirm that the hair cells in apex turn
were regenerated from the gentamicin damaged inner ear cells.
Therefore, future in vivo experiments are required.
In summary, the Notch signaling pathway was
demonstrated to have an important role in the negative regulation
of hair cell regeneration. Additionally, gentamicin-induced Notch
activation reduced miR-183 expression. Inhibition of the Notch
signaling pathway by DAPT attenuated these effects and promoted the
transdifferentiation of supporting cells into hair cells. miR-183
may be a crucial factor in the differentiation and regeneration of
the Notch signaling-inhibited hair cells from mouse cochlea.
Inhibition of the Notch signaling pathway induced miR-183
expression, which participated in hair cell differentiation and
regeneration. These findings suggest a therapeutic approach to
promote mammalian hair cell regeneration by modulating Notch or
miR-183 function.
Acknowledgements
The present study was supported by the grants from
the National Basic Research Program of China (grant no.
2011CB504502), National Natural Science Foundation of China (grant
no. 30973306, 81271076, 81200748 and 81500802), China Post-Doctoral
Science Foundation (grant no. 2014M562329) and Key Natural Science
Foundation of Guangdong Province (grant no. 8251008901000016).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Corwin JT and Cotanche DA: Regeneration of
sensory hair cells after acoustic trauma. Science. 240:1772–1774.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ryals BM and Rubel EW: Hair cell
regeneration after acoustic trauma in adult Coturnix quail.
Science. 240:1774–1776. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Warchol ME, Lambert PR, Goldstein BJ,
Forge A and Corwin JT: Regenerative proliferation in inner ear
sensory epithelia from adult guinea pigs and humans. Science.
259:1619–1622. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Adam J, Myat A, Le Roux I, Eddison M,
Henrique D, Ish-Horowicz D and Lewis J: Cell fate choices and the
expression of Notch, Delta and Serrate homologues in the chick
inner ear: Parallels with drosophila sense-organ development.
Development. 125:4645–4654. 1998.PubMed/NCBI
|
|
5
|
Daudet N and Lewis J: Two contrasting
roles for Notch activity in chick inner ear development:
Specification of prosensory patches and lateral inhibition of
hair-cell differentiation. Development. 132:541–551. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Daudet N, Gibson R, Shang J, Bernard A,
Lewis J and Stone J: Notch regulation of progenitor cell behavior
in quiescent and regenerating auditory epithelium of mature birds.
Dev Biol. 326:86–100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kelley MW: Regulation of cell fate in the
sensory epithelia of the inner ear. Nat Rev Neurosci. 7:837–849.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hartman BH, Basak O, Nelson BR, Taylor V,
Bermingham-McDonogh O and Reh TA: Hes5 expression in the postnatal
and adult mouse inner ear and the drug-damaged cochlea. J Assoc Res
Otolaryngol. 10:321–340. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murata J, Ikeda K and Okano H: Notch
signaling and the developing inner ear. Adv Exp Med Biol.
727:161–173. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Weston MD, Pierce ML, Jensen-Smith HC,
Fritzsch B, Rocha-Sanchez S, Beisel KW and Soukup GA: MicroRNA-183
family expression in hair cell development and requirement of
microRNAs for hair cell maintenance and survival. Dev Dyn.
240:808–819. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang XR, Zhang XM, Du J and Jiang H:
MicroRNA-182 regulates otocyst-derived cell differentiation and
targets T-box1 gene. Hea Res. 286:55–63. 2012. View Article : Google Scholar
|
|
12
|
Sage C, Huang M, Karimi K, Gutierrez G,
Vollrath MA, Zhang DS, García-Añoveros J, Hinds PW, Corwin JT,
Corey DP and Chen ZY: Proliferation of functional hair cells in
vivo in the absence of the retinoblastoma protein. Science.
307:1114–1118. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee
DH, Nguyen JT, Barbisin M, Xu NL, Mahuvakar VR, Andersen MR, et al:
Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic
Acids Res. 33:e1792005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kloosterman WP, Wienholds E, de Bruijn E,
Kauppinen S and Plasterk RH: In situ detection of miRNAs in animal
embryos using LNA-modified oligonucleotide probes. Nat Methods.
3:27–29. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stone JS and Cotanche DA: Hair cell
regeneration in the avian auditory epithelium. Int J Dev Biol.
51:633–647. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stone JS and Rubel EW: Delta1 expression
during avian hair cell regeneration. Development. 126:961–973.
1999.PubMed/NCBI
|
|
18
|
Eddison M, Le Roux I and Lewis J: Notch
signaling in the development of the inner ear: Lessons from
Drosophila. Proc Natl Acad Sci USA. 97:11692–11699. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Żak M, Klis SF and Grolman W: The Wnt and
Notch signalling pathways in the developing cochlea: Formation of
hair cells and induction of regenerative potential. Int J Dev
Neurosci. 47:247–258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roberson DW, Alosi JA and Cotanche DA:
Direct transdifferentiation gives rise to the earliest new hair
cells in regenerating avian auditory epithelium. J Neurosci Res.
78:461–471. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang XR, Zhang XM, Du J and Jiang H:
MicroRNA-182 regulates otocyst-derived cell differentiation and
targets T-box1 gene. Hear Res. 286:55–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheung CC, Lun SW, Chung GT, Chow C, Lo C,
Choy KW and Lo KW: MicroRNA-183 suppresses cancer stem-like cell
properties in EBV-associated nasopharyngeal carcinoma. BMC Cancer.
16:4952016. View Article : Google Scholar : PubMed/NCBI
|