Introduction
Skin photoaging is a cosmetic concern worldwide, and
is characterized by atrophy of the skin, coarse wrinkles and
leathery skin (1). Sun exposure is
the main factor leading to photoaging, primarily due to ultraviolet
(UV) radiation. There are three categories of ultraviolet light
according to its wavelength, including UVA (320–400 nm), UVB
(290–320 nm) and UVC (200–290 nm) (2). UVB is considered to be the most
important factor associated with skin photoaging (3). UVB radiation exposure can induce the
production of reactive oxygen species (ROS) in dermal fibroblasts,
including the hydroxyl free radical, superoxide anion, singlet
oxygen and hydrogen peroxide (4).
Increased ROS levels can damage dermal fibroblasts, the most
important cell type in the dermis that produces extracellular
matrix (ECM). Subsequently, the synthesis of ECM could be inhibited
and degradation may be accelerated, resulting in skin aging
(2). Antioxidants, including
vitamins C and E, and coenzyme Q10, could inhibit and neutralize
ROS; thus, they have been suggested for the treatment and
prevention of skin photoaging (5).
Saffron has been used in traditional Chinese
medicine for a number of years. It has been traditionally used for
the treatment of many types of disease, including neurodegenerative
disorders, coronary artery diseases, respiratory diseases and
gastrointestinal diseases (6). The
therapeutic effects of saffron are associated with some of its
components. Crocin is one of the main and active constituents
isolated from saffron. It has been shown previously that crocin
exhibits multiple activities, including anti-cancer,
anti-inflammatory and anti-oxidation effects in various cell types
(7). Lv et al (8) reported that crocin exerts an
anti-oxidative effect against H2O2-induced
apoptosis in retinal ganglion cells.
Based on the anti-oxidant capacity of crocin, it was
speculated that it may prevent UVB-induced skin aging. In the
present study, the protective effects of crocin against UVB-induced
damage were investigated in cultured human dermal fibroblasts.
Materials and methods
Preparation of crocin
Crocin (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) was dissolved in sterile PBS solution at a concentration
of 10 mM, stored at −20°C and then freshly thawed at room
temperature prior to each use.
Antioxidant effects of crocin
To test the antioxidant effects of crocin,
1,1-diphenyl-2-picrylhydrazyl (DPPH; Sigma-Aldrich; Merck KGaA)
radical scavenging activity was measured as previously described
(9). Reaction mixtures (200 µl)
containing DPPH and a serial dilution of crocin (100–800 µM) were
placed in a 96-well plate at room temperature in the dark for 30
min, then the absorbance was measured at 515 nm using a Varioskan
Flash Spectral Scanning Multimode Reader (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The inhibition rate was determined using
the following equation: Inhibition rate
(%)=[1-(Asample/Acontrol)] ×100. Where A
refers to the absorbance measured at 515 nm.
Cell isolation and culture
Dermal fibroblasts were isolated from human foreskin
specimens. Samples were obtained from five donors (age, 6–12 years)
who underwent a routine circumcision procedure at Shanghai 9th
People's Hospital between January 2016 and January 2017. Written
informed consent was obtained. The present study was approved by
The Ethics Committee of Shanghai 9th People's Hospital, Shanghai
Jiao Tong University School of Medicine (Shanghai, China). A
single-cell suspension was obtained as previously described
(10). Cells were then suspended
in Dulbecco's modified Eagle's medium (Invitrogen; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA), 300 µg/ml
L-glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin (all
Sigma-Aldrich; Merck KGaA). Cells were maintained at 37°C in a
humidified atmosphere containing 5% CO2 and passaged by
trypsinization with 0.25% trypsin-EDTA (Gibco; Thermo Fisher
Scientific, Inc.) every 3–5 days. Cells at passage 3–5 were used in
the following experiments.
Crocin toxicity analysis
Fibroblasts were seeded in a 96-well plate at a
density of 2,000 cells/well and treated with different
concentrations of crocin (0, 12.5, 50 and 100 µM) and subsequently
maintained at 37°C in a humidified atmosphere containing 5%
CO2 for 72 h. Cell proliferation was then measured using
a Cell Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology,
Haimen, China), according to the manufacturer's instructions.
Untreated cells served as the control.
UVB irradiation
Fibroblasts were cultured in 96-well (2,000
cells/well) or 6-well plates (1×105 cells/well) in the
aforementioned supplemented DMEM mixture, and maintained at 37°C
for 24 h. Following a further 24 h of incubation at 37°C with or
without crocin (0, 12.5, 50 and 100 µM), culture medium was
replaced with PBS. Cells were then exposed to UVB light (Philips
311 nm, TL 20W/01; Philips Lighting Holding B.V., Eindhoven, The
Netherlands) at a total dose of 100 mJ/cm2. Following
irradiation, the medium was replaced with culture medium with or
without crocin, and cells were maintained at 37°C in a humidified
atmosphere containing 5% CO2 for 24 or 72 h for further
analyses. Cells treated with UVB irradiation and 150 µM vitamin C
(Vit C; Sigma-Aldrich; Merck KGaA) served as a positive control.
The following experiments were then performed.
Determination of cell
proliferation
Cell proliferation was measured at 72 h
post-irradiation using a CCK-8 kit (Beyotime Institute of
Biotechnology), according to the manufacturer's instructions.
Results are expressed as the relative cell proliferation (%) with
respect to the control cells (cells without UVB irradiation or
crocin treatment).
Cell cycle analysis
At 24 h post-UVB radiation, 5×105 cells
were harvested by 0.25% Trypsin-EDTA and then fixed in 70% ethanol
overnight at 4°C. Fixed cells were washed twice with PBS and then
incubated with 1.5 mg/l RNase A (Sigma-Aldrich; Merck KGaA) for 1 h
at 37°C, followed by staining with 5 µl of propidium iodide
(Sigma-Aldrich; Merck KGaA) for 20 min on ice. DNA content was
assessed using an Epics Altra Flow Cytometer (Beckman Coulter,
Inc., Brea, CA, USA), and analyzed with Modi Fit LT v2.0 software
(Verity Software House, Inc., Topsham, ME, USA).
β-galactosidase (SA-β-gal)
staining
To measure the cell-aging rate, SA-β-gal staining
was performed at 72 h post-irradiation using a Senescence
β-Galactosidase staining kit purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA; cat. no. 9860). Cells were
washed in PBS, fixed for 10 min at room temperature in 4%
paraformaldehyde and stained according to manufacturer's
instructions. Five random fields from each sample (n=3
samples/group) were selected to observe under a light microscope
(Nikon Eclipse 90i; Nikon Corporation, Tokyo, Japan), and the
number of SA-β-gal-positive cells was counted using Image-Pro Plus
6.0 software (Media Cybernetics, Inc., Rockville, MD, USA). The
aging rate was determined as the percentage of positive cells out
of the total number of cells.
Measurement of intracellular ROS
Intracellular ROS levels were determined by
2′,7′-dichlorodihydrofluoresce in diacetate (DCFH2-DA;
Sigma-Aldrich; Merck KGaA) staining. Briefly, immediately following
UVB irradiation, cells were incubated with DCFH2-DA (10
mM) at 37°C for 20 min. Following this, half of the cells were
observed under a fluorescent microscope (Olympus IX70-S1F2; Olympus
Corporation, Tokyo, Japan), and the other half of the cells were
collected and analyzed with an Epics Altra Flow Cytometer, as
described previously (11).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Cells (2×106) were collected at 72 h
post-irradiation. Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and cDNA was
synthesized from 2 µg total RNA using 200 U of reverse
transcriptase (MMLV-RT) and 20 pM oligo dT (Promega Corporation,
Madison, WI, USA) at 42°C for 1 h. The expression of pro-collagen I
was determined by qPCR using SYBR Green PCR Master mix (Applied
Biosystems, Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: Initial denaturation at 95°C for 10
min, followed by 40 cycles at 95°C for 30 sec, 60°C for 30 sec and
72°C for 45 sec, using the Strata Gene Mx3000p (Agilent
Technologies, Inc., Santa Clara, CA, USA). Expression was
quantified using the 2−∆∆Cq method (12). The primers employed were as
follows: Pro-collagen I, forward: 5′-CTCGAGGTGGACACCACCCT-3′ and
reverse: 5′-CAGCTGGATGGCCACATCGG-3′. All amplifications were run in
triplicate and the results were normalized to the housekeeping gene
GAPDH, the primers of which were as follows: Forward:
5′-CAAAAGGGTCATCATCTCTG-3′ and reverse: 5′-CCTGCTTCACCACCTTCTTG-3′.
Three independent experiments were performed.
Western blot analysis
Cells (2×106) were collected at 72 h
post-irradiation. Total protein was extracted for western blot
analysis and the expression of collagen type 1 (Col-1) and
glutathione peroxidase 1 (GPX-1) was measured. Proteins were
harvested and collected with radioimmunoprecipitation assay lysis
buffer (Beyotime Institute of Biotechnology). Protein
concentrations were determined with a bicinchoninic acid protein
assay. Subsequently, proteins (20 µg/lane) were separated by 12%
SDS-PAGE. Following electrophoresis at 100 V for 2 h, proteins were
transferred to polyvinylidene difluoride membranes at 350 mA for 90
min. The membranes were blocked with 5% non-fat milk in
Tris-buffered saline and Tween 20 (TBST) at room temperature for 2
h, followed by incubation with the following primary antibodies
overnight at 4°C: Anti-Col-1 (cat. no. ab6308; 1:1,000; Abcam,
Cambridge, UK), anti-GPX-1 (cat. no. ab108427; 1:2,000; Abcam) and
β-actin (cat. no. 4970S; 1:2,000; Cell Signaling Technology, Inc.).
The membranes were washed three times with TBST and subsequently
incubated with horseradish peroxidase-conjugated goat anti-mouse
secondary antibody (cat. no. 115-035-062; 1:2,000; Jackson
ImmunoResearch Laboratories, Inc., West Grove, PA, USA) at room
temperature for 2 h. Protein expression levels were analyzed by
visualizing the bands with enhanced chemiluminescence (Pierce,
Rockford, USA). ImageJ 1.50i software (National Institutes of
Health USA) was used for densitometry.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Unpaired t-tests were used for direct comparisons between the UVB
and the UVB + crocin groups. For multiple comparisons, one-way
analysis of variance followed by Tukey's post hoc test was
performed. All analyses were performed using SPSS v.13.0 software
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
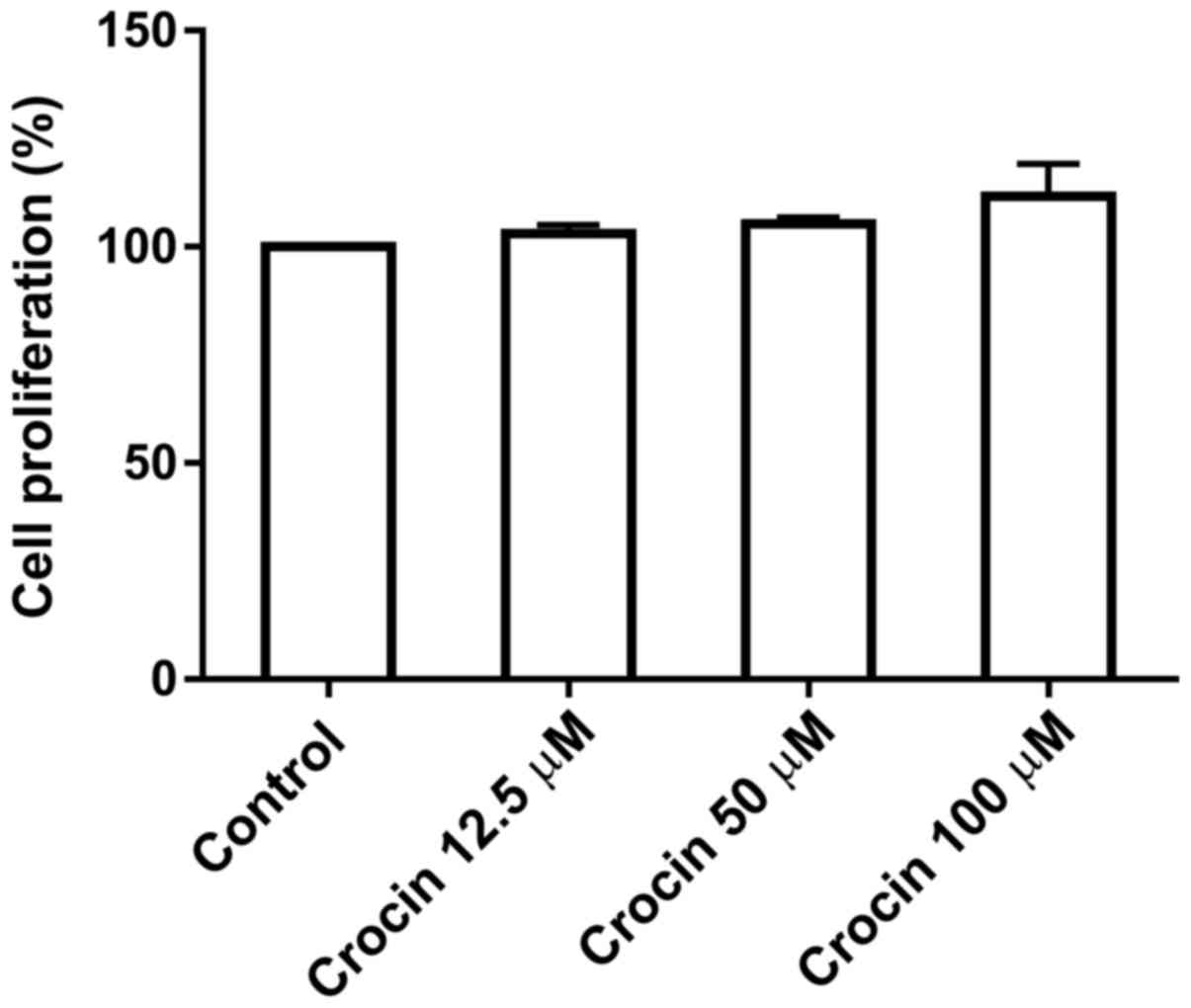
Cytotoxicity of crocin
To test the potential cytotoxicity of crocin on
dermal fibroblasts, cells were treated with various concentrations
(0, 12.5, 50 and 100 µM) of crocin for 72 h prior to measuring the
levels of proliferation using a CCK-8 assay. As shown in Fig. 1, crocin did not inhibit the
proliferation of cells at the concentrations tested, up to 100 µM,
indicating that crocin at this dose range may not be toxic to
fibroblasts.
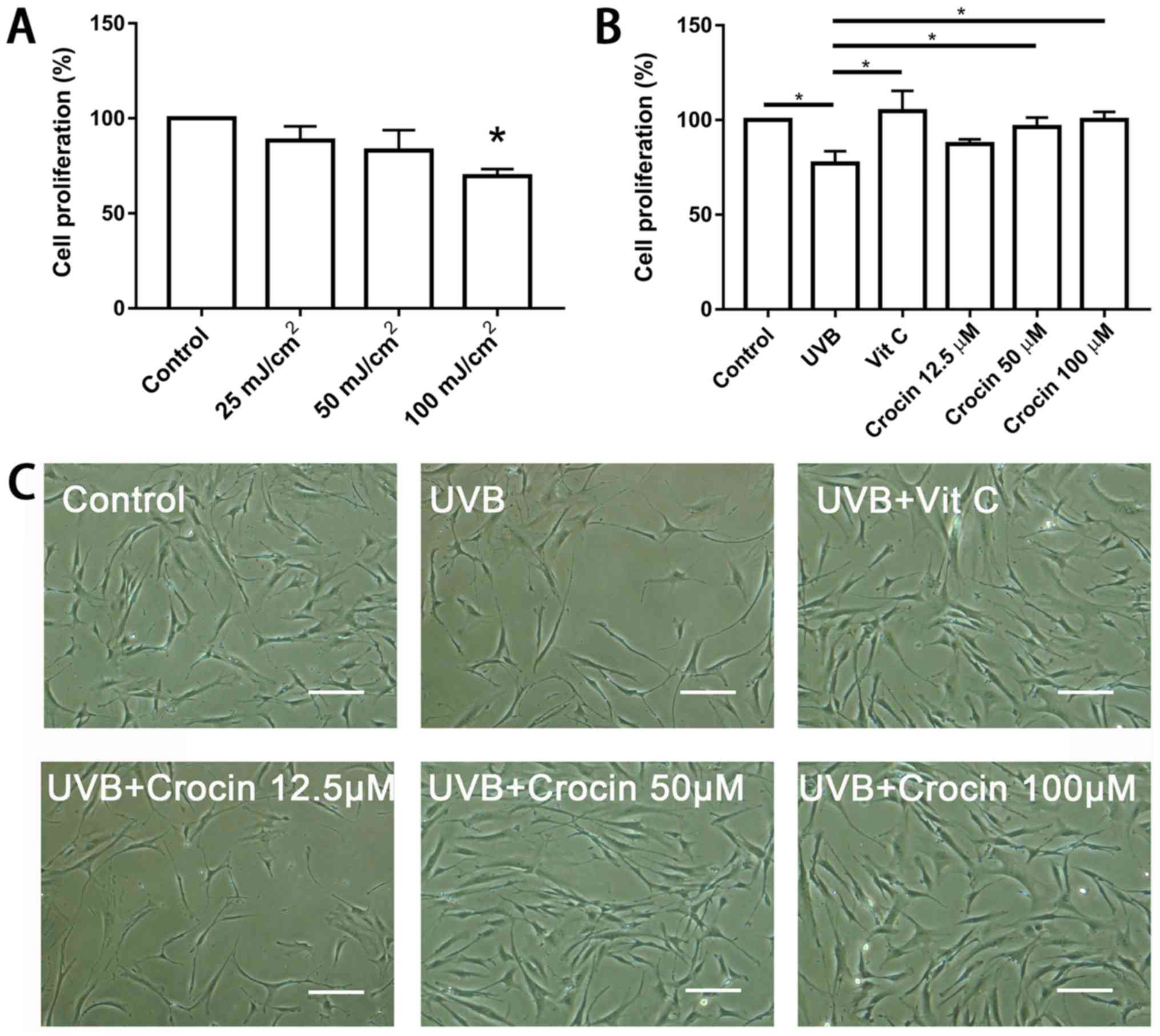
Crocin rescues the cell proliferation
inhibited by UVB irradiation
To determine the optimal dose of UVB irradiation,
fibroblasts were exposed to a range of UVB doses (0, 25, 50 and 100
mJ/cm2) and a CCK-8 kit was used to measure cell
proliferation following 72 h. UVB irradiation significantly
inhibited cell proliferation at the dose of 100 mJ/cm2
(Fig. 2A). Therefore, 100
mJ/cm2 was selected for the subsequent experiments.
Following pre-incubation with crocin (12.5, 50 and 100 µM) for 24
h, cells were irradiated by UVB and cell proliferation was measured
following 72 h. High doses (50 and 100 µM) of crocin significantly
rescued the cell proliferation inhibited by UVB irradiation
(P<0.05), and the rate was similar to that following Vit C
treatment (Fig. 2B and C).
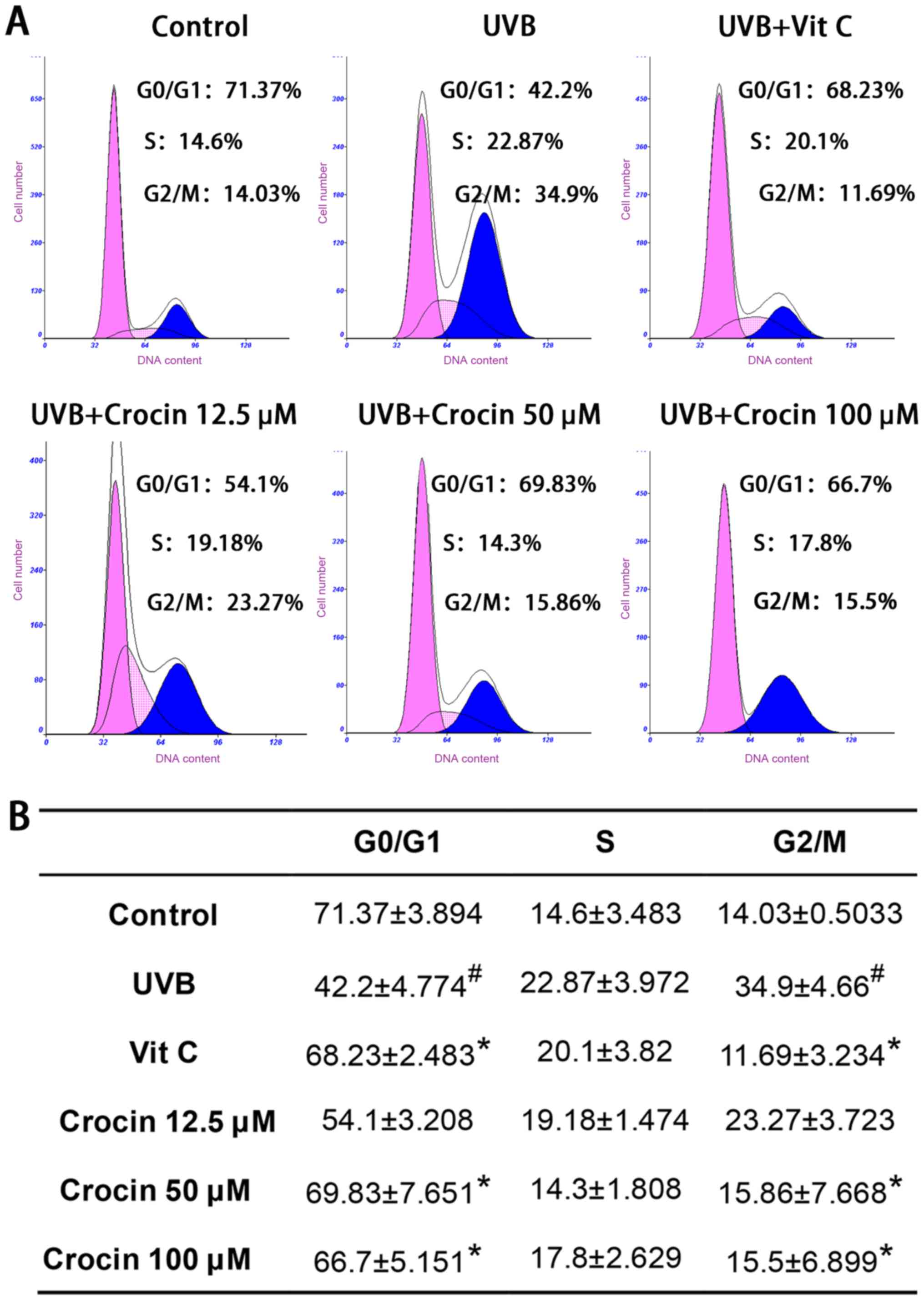
Crocin rescues UVB-induced cell cycle
arrest
Flow cytometry analyses of the cell cycle are shown
in Fig. 3. Cell populations in the
G2/M phases were significantly increased following UVB
irradiation, indicating that cells were arrested at the
G2/M phase. As a positive control, Vit C treatment
rescued cell cycle arrest as expected. High doses (50 and 100 µM)
of crocin restored cell cycle arrest to levels similar to those of
the control group (without UVB irradiation), indicating that crocin
could rescue UVB-induced cell cycle arrest.
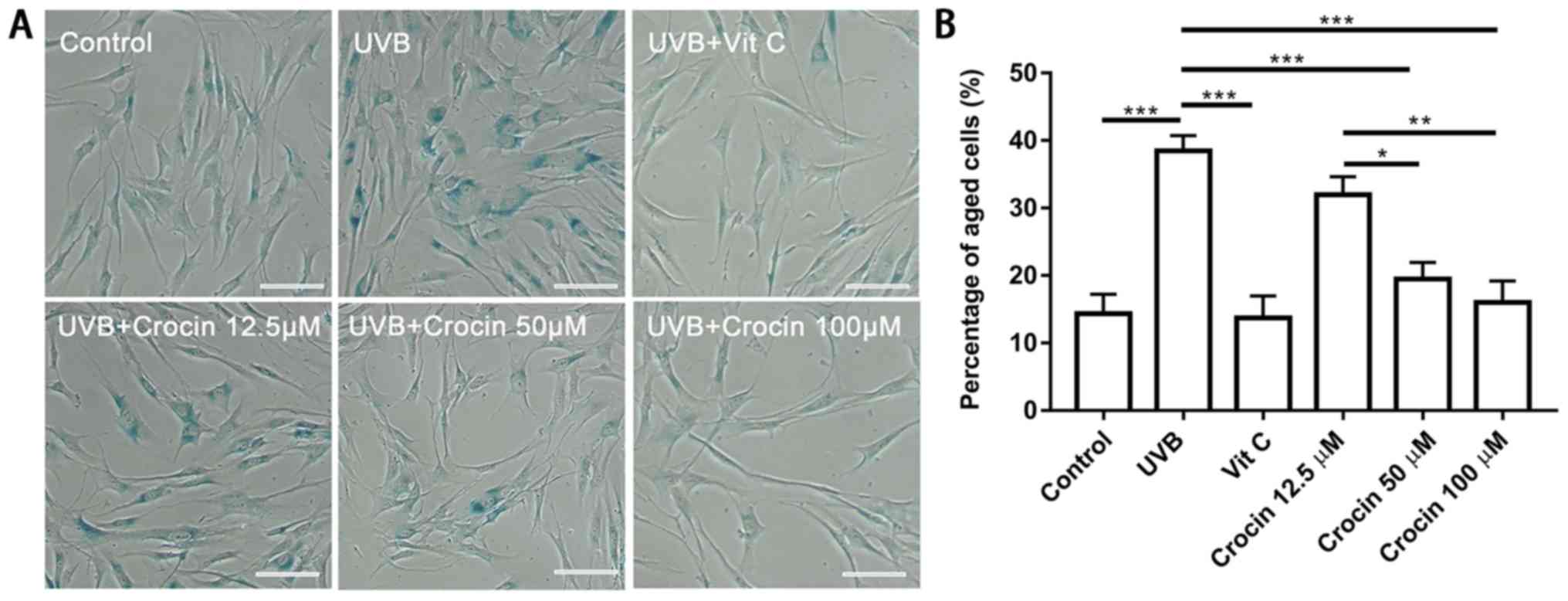
Crocin prevents UVB-induced cell
aging
To determine whether crocin could prevent cells from
UVB-induced photoaging, SA-β-gal staining was performed following
UVB irradiation (Fig. 4A). A
significant increase in SA-β-gal-positive cells was observed
following UVB irradiation (Fig.
4). A decrease in SA-β-gal-positive cells was observed in the
Vit C-treated group as well as in the high dose (50 and 100 µM)
crocin-treated groups (Fig. 4).
Statistical analysis confirmed that the percentage of
SA-β-gal-positive cells was increased in the UVB group, and
decreased following Vit C or crocin treatment (Fig. 4B).
Crocin reduces UVB-induced
intracellular ROS
An increase in ROS is one of the most important
mechanisms in photoaging (4). In
the present study, UVB irradiation significantly increased
intracellular ROS when compared with the control group, while the
ROS levels were reduced by Vit C and crocin treatment (Fig. 5A). These results were confirmed by
flow cytometric analysis of the fluorescent intensity following
DCFH2-DA staining (Fig.
5B). To verify the possible underlying mechanisms, free radical
scavenging activity of crocin was measured. Crocin inhibited DPPH
radial activity in a dose-dependent manner (Fig. 5C), indicating that it could
partially neutralize ROS. In addition, the expression of the
antioxidant protein GPX-1 was measured by western blot analysis. An
increase in GPX-1 expression was observed in the Vit C and
crocin-treated groups (Fig. 5D and
E), indicating that crocin may reduce UVB-induced ROS,
partially via the enhanced expression of antioxidant protein
GPX-1.
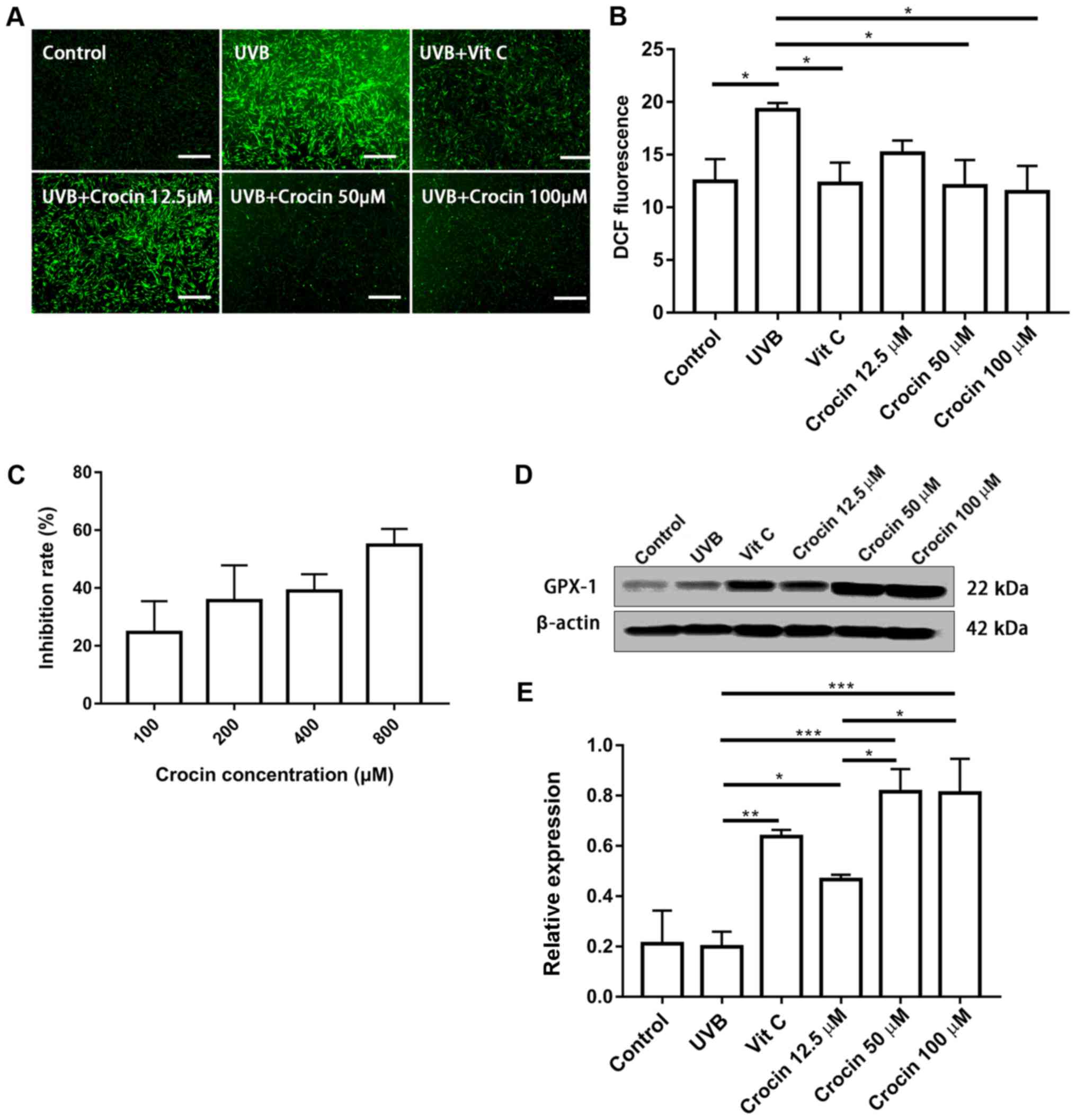 | Figure 5.Crocin reduces UVB-induced
intracellular ROS. (A) Intracellular ROS measured by
DCFH2-DA staining at 20 min post-UVB irradiation. Scale
bars, 500 µm. (B) Intracellular ROS measured by DCFH2-DA staining
and flow cytometry. (C) Antioxidant effect of crocin measured by
1,1-diphenyl-2-picrylhydrazyl radical-scavenging activity. (D)
Western blot analysis of (E) GPX-1 expression at 72 h post-UVB
irradiation. *P<0.05, **P<0.01 and ***P<0.001, as
indicated. UVB, ultraviolet B; Vit C, vitamin C; ROS, reactive
oxygen species; GPX-1, glutathione peroxidase 1;
DCFH2-DA, 2′,7′-dichlorodihydrofluoresce in
diacetate. |
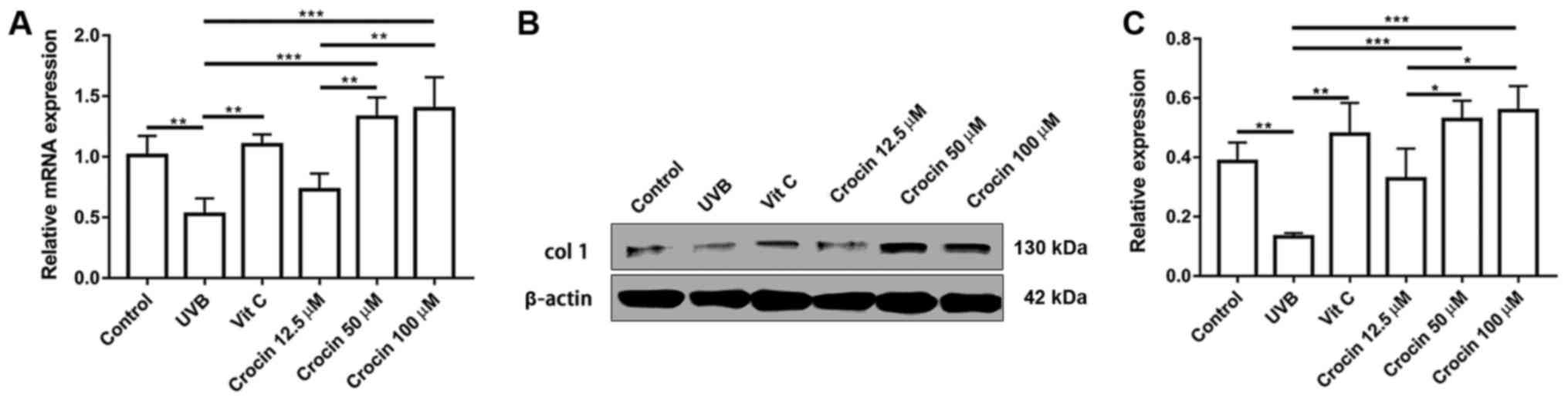
Crocin promotes the expression of ECM
protein Col-1
Secretion of ECM is one of the major functions of
dermal fibroblasts. A decrease in collagen secretion is commonly
observed during skin aging (13).
As shown in Fig. 6, a significant
decrease in Col-1 expression was observed at the mRNA and protein
level following UVB irradiation. In addition, Col-1 expression was
upregulated by Vit C and crocin treatment at the mRNA and protein
levels, suggesting that cell functions were also restored following
Vit C and crocin treatment.
Discussion
Extrinsic skin aging is caused by environmental
factors including smoking, consuming alcohol, UV irradiation and
common pollutants, which can lead to dermal fibroblast damage and
aging (14,15). UVB is considered to be one of the
most important factors that contributes towards aging (16). In the present study, the results
confirmed that UVB irradiation induced the production of
intracellular ROS, arrested the cell cycle, inhibited cell
proliferation and downregulated ECM production. Antioxidants, such
as Vit C, which scavenge ROS, are able to protect cells from
UVB-induced damage (17).
Crocin, one of the main and active constituents of
Crocus sativus, has been reported to have various
bioactivities, including immunomodulatory, antitumor,
anti-inflammation and antioxidant properties (18–22).
Thus, it is not surprising that crocin could protect UVB-induced
cell damage in cultured dermal fibroblasts, which was demonstrated
by reduced intracellular ROS, rescued cell cycle and proliferation,
and upregulated ECM production in the present study. One possible
mechanism by which crocin may reduce intracellular ROS is by
neutralizing ROS directly, as crocin could inhibit DPPH radical
activity in a dose-dependent manner. Another potential mechanism
could be that crocin upregulates antioxidant gene expression, which
is supported by the upregulated expression of antioxidant protein
GPX-1 following crocin treatment observed in the present study.
However, an increased protective effect with 100 µM crocin
treatment was not observed when compared with 50 µM crocin
treatment. Intracellular ROS measurement revealed that ROS levels
returned to basal (control) levels following 50 and 100 µM crocin
treatment, indicating that 50 µM crocin may be sufficient to
neutralize the UVB-induced ROS accumulation. Although GPX-1
expression was upregulated following crocin treatment, the
regulatory pathway is still not clear. Therefore, it is worthy of
further investigation in the future.
As a natural product, crocin has been widely used as
a spice and a food colorant. Previous studies have demonstrated
that crocin does not cause damage to any major organ in
experimental animals (23,24). Mohamadpour et al (25) evaluated the safety of crocin in
healthy volunteers and confirmed that it is relatively safe. In the
present study, the proliferation of dermal fibroblasts was not
affected by crocin treatment at the tested concentrations; in fact,
slight stimulation of cell growth was observed following crocin
treatment. These results strongly suggest that crocin could be used
in the clinic without significant toxicity at pharmacological
doses.
In the present study, the dose of UVB irradiation
was optimized. Cells did not undergo apoptosis or necrosis at this
exposure dose (data not shown), and the cell cycle was arrested at
the G2/M phases. Cell senescence was observed by
SA-β-gal staining, which produced similar results to those observed
in previous studies (26–28). One limitation of the present study
was that the protective effects of crocin against a higher dose of
UVB-irradiation were not investigated. The anti-apoptotic potential
of crocin has been demonstrated in previous studies (29,30).
Jia et al (11) reported
that crocin protected retinal pigment epithelial cells from
H2O2-induced damage through the upregulation
of the anti-apoptotic genes B-cell lymphoma 2 (Bcl-2) and
Bcl-2-associated X protein. Thus, it is possible that crocin may
exhibit protective effects at higher doses of irradiation via its
anti-apoptotic activity. Another limitation of the present study
was that cells were pre-treated with crocin 24 h prior to UVB
irradiation. It remains to be investigated whether the application
of crocin following irradiation would still protect the cells.
Since crocin reduces intracellular ROS partially via the
upregulation of antioxidant gene expression, it was speculated that
the protective effects may be weaker than with pre-incubation due
to the delayed expression of antioxidant genes. In future research,
we aim to determine the anti-photoaging capacity of crocin in nude
mice.
The protective effects of plant extracts on
UVB-induced skin damage have been reported previously, including
extracts from berries and herbs (9,31).
In general, the plant extracts contain a mixture of active
components. In the present study, it was demonstrated that a single
compound with a definite chemical structure possesses
anti-photoaging activity. Furthermore, it is superior to the plant
extracts that can be industrially synthesized with better quality
control. The protective effects of crocin on photoaging fibroblasts
suggest that it has potential applications in the protection
against UVB-induced skin photoaging.
Acknowledgements
The authors deeply appreciated the contribution of
members of the Shanghai Key Laboratory of Tissue Engineering,
National Tissue Engineering Center of China (Shanghai, China) for
providing valuable suggestions and discussions.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study is
included in this published article.
Authors' contributions
GZ, WL, YC and WZ conceived and designed the
experiments. MD, DL and YZ performed the experiments. DL analyzed
the data. MD, WZ and YC wrote and revised the manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committee of Shanghai 9th People's Hospital, Shanghai Jiao Tong
University School of Medicine (Shanghai, China). Written informed
consent was obtained from the donors.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gilchrest BA: Photoaging. J Invest
Dermatol. 133:E2–E6. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kammeyer A and Luiten RM: Oxidation events
and skin aging. Ageing Res Rev. 21:16–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee YR, Noh EM, Jeong EY, Yun SK, Jeong
YJ, Kim JH, Kwon KB, Kim BS, Lee SH, Park CS and Kim JS: Cordycepin
inhibits UVB-induced matrix metalloproteinase expression by
suppressing the NF-kappaB pathway in human dermal fibroblasts. Exp
Mol Med. 41:548–554. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Poon F, Kang S and Chien AL: Mechanisms
and treatments of photoaging. Photodermatol Photoimmunol Photomed.
31:65–74. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stern RS: Clinical practice. Treatment of
photoaging. N Engl J Med. 350:1526–1534. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Boskabady MH and Farkhondeh T:
Antiinflammatory, antioxidant, and immunomodulatory effects of
Crocus sativus L. and its main constituents. Phytother Res.
30:1072–1094. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Samarghandian S, Azimi-Nezhad M, Borji A
and Farkhondeh T: Effect of crocin on aged rat kidney through
inhibition of oxidative stress and proinflammatory state. Phytother
Res. 30:1345–1353. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lv B, Chen T, Xu Z, Huo F, Wei Y and Yang
X: Crocin protects retinal ganglion cells against H2O2-induced
damage through the mitochondrial pathway and activation of NF-κB.
Int J Mol Med. 37:225–232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chiang HM, Lin TJ, Chiu CY, Chang CW, Hsu
KC, Fan PC and Wen KC: Coffea arabica extract and its constituents
prevent photoaging by suppressing MMPs expression and MAP kinase
pathway. Food Chem Toxicol. 49:309–318. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen FG, Zhang WJ, Bi D, Liu W, Wei X,
Chen FF, Zhu L, Cui L and Cao Y: Clonal analysis of nestin(−)
vimentin(+) multipotent fibroblasts isolated from human dermis. J
Cell Sci. 120:2875–2883. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jia C, Lu Y, Bi B, Chen L, Yang Q, Yang P,
Guo Y, Zhu J, Zhu N and Liu T: Platelet-rich plasma ameliorates
senescence-like phenotypes in a cellular photoaging model. Rsc Adv.
7:3152–3160. 2017. View Article : Google Scholar
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brenneisen P, Sies H and
Scharffetter-Kochanek K: Ultraviolet-B irradiation and matrix
metalloproteinases: From induction via signaling to initial events.
Ann N Y Acad Sci. 973:31–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bernhard D, Moser C, Backovic A and Wick
G: Cigarette smoke-an aging accelerator? Exp Gerontol. 42:160–165.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wlaschek M, Tantcheva-Poor I, Naderi L, Ma
W, Schneider LA, Razi-Wolf Z, Schüller J and Scharffetter-Kochanek
K: Solar UV irradiation and dermal photoaging. J Photochem
Photobiol B. 63:41–51. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Adil MD, Kaiser P, Satti NK, Zargar AM,
Vishwakarma RA and Tasduq SA: Effect of Emblica officinalis (fruit)
against UVB-induced photo-aging in human skin fibroblasts. J
Ethnopharmacol. 132:109–114. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Farris PK: Topical vitamin C: A useful
agent for treating photoaging and other dermatologic conditions.
Dermatol Surg. 31:814–818. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Deslauriers AM, Afkhami-Goli A, Paul AM,
Bhat RK, Acharjee S, Ellestad KK, Noorbakhsh F, Michalak M and
Power C: Neuroinflammation and endoplasmic reticulum stress are
coregulated by crocin to prevent demyelination and
neurodegeneration. J Immunol. 187:4788–4799. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Prieto MA, Vázquez JA and Murado MA:
Crocin bleaching antioxidant assay revisited: Application to
microplate to analyse antioxidant and pro-oxidant activities. Food
Chem. 167:299–310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Du J, Chi Y, Song Z, Di Q, Mai Z, Shi J
and Li M: Crocin reduces aspergillus fumigatus-induced airway
inflammation and NF-κB signal activation. J Cell Biochem.
119:1746–1754. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bolhassani A, Khavari A and Bathaie SZ:
Saffron and natural carotenoids: Biochemical activities and
anti-tumor effects. Biochim Biophys Acta. 1845:20–30.
2014.PubMed/NCBI
|
|
22
|
Xu GL, Li G, Ma HP, Zhong H, Liu F and Ao
GZ: Preventive effect of crocin in inflamed animals and in
LPS-challenged RAW 264.7 cells. J Agric Food Chem. 57:8325–8330.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hosseinzadeh H and Jahanian Z: Effect of
Crocus sativus L. (saffron) stigma and its constituents,
crocin and safranal, on morphine withdrawal syndrome in mice.
Phytother Res. 24:726–730. 2010.PubMed/NCBI
|
|
24
|
Hosseinzadeh H, Karimi G and Niapoor M:
Antidepressant effects of Crocus sativus L. stigma extracts
and its constituents, crocin and safranal, in mice. Acta
Horticulturae. 650:435–445. 2004. View Article : Google Scholar
|
|
25
|
Mohamadpour AH, Ayati Z, Parizadeh MR,
Rajbai O and Hosseinzadeh H: Safety evaluation of crocin (a
constituent of saffron) tablets in healthy volunteers. Iran J Basic
Med Sci. 16:39–46. 2013.PubMed/NCBI
|
|
26
|
Zhang JA, Yin Z, Ma LW, Yin ZQ, Hu YY, Xu
Y, Wu D, Permatasari F, Luo D and Zhou BR: The protective effect of
baicalin against UVB irradiation induced photoaging: An in vitro
and in vivo study. PLoS One. 9:e997032014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park J, Seok JK, Suh HJ and Boo YC:
Gardenia jasminoides extract attenuates the UVB-induced expressions
of cytokines in keratinocytes and indirectly inhibits matrix
metalloproteinase-1 expression in human dermal fibroblasts. Evid
Based Complement Alternat Med. 2014:4292462014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zeng Q, Zhou F, Lei L, Chen J, Lu J, Zhou
J, Cao K, Gao L, Xia F, Ding S, et al: Ganoderma lucidum
polysaccharides protect fibroblasts against UVB-induced photoaging.
Mol Med Rep. 15:111–116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
El-Maraghy SA, Rizk SM and Shahin NN:
Gastroprotective effect of crocin in ethanol-induced gastric injury
in rats. Chem Biol Interact. 229:26–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thushara RM, Hemshekhar M, Santhosh MS,
Jnaneshwari S, Nayaka SC, Naveen S, Kemparaju K and Girish KS:
Crocin, a dietary additive protects platelets from oxidative
stress-induced apoptosis and inhibits platelet aggregation. Mol
Cell Biochem. 373:73–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bravo K, Duque L, Ferreres F, Moreno DA
and Osorio E: Passiflora tarminiana fruits reduce UVB-induced
photoaging in human skin fibroblasts. J Photochem Photobiol B.
168:78–88. 2017. View Article : Google Scholar : PubMed/NCBI
|