Introduction
Gastric cancer is one of the most common malignant
tumors worldwide. A number of factors contribute to the development
of gastric cancer, of which the pathogenesis is highly complex
(1,2). Helicobacter pylori infection
is listed as a class I carcinogenic factor for gastric cancer by
the World Health Organization, and high salt and high nitrate diets
may also be risk factors for gastric cancer development (3). Genetic factors, environmental factors
and bacterial infections ultimately affect the occurrence and
progression of gastric cancer (4,5). It
has been reported that although gastric cancer treatment and
prognosis has greatly improved in China, the incidence of gastric
cancer remains high (6). As there
is a lack of knowledge of specific symptoms, the diagnosis of
gastric cancer at an early stage is difficult. Gastrectomy is a
widely used strategy in gastric cancer therapy. However, the
prognosis of patients with gastric cancer at advanced stages is
unsatisfactory (7). Therefore, a
better understanding of the occurrence and progression of gastric
cancer is of scientific significance.
The primary target molecule of radiotherapy is DNA.
The mechanism of cell DNA damage repair is initiated by radiation
exposure, which activates cell cycle arrest, thereby promoting
repair of injury (8). If DNA fails
to repair, it may result in cell death, necrosis or senescence
(8). DNA strand breaks (DSBs)
induced by radiation exposure are closely associated with cell
death. DSB repair is associated with radiosensitivity (9). The effectiveness of therapy of
gastric cancer primarily depends on the sensitivity of the tumor to
radiotherapy (10–12). Radiation resistance has become key
to further deterioration of tumors, thus the study of
radiosensitization has become more prevalent.
Gene therapy is being increasingly recognized in
tumor therapy. Tumor radiosensitivity is associated with its
internal molecular biological mechanism. It has been demonstrated
that the abnormal expression of a number of oncogenes and tumor
suppressor genes may affect tumor cell apoptosis, radiosensitivity
and patient prognosis (13). The
potential combination of tumor gene therapy and radiotherapy has
therefore been suggested, to ultimately reduce normal tissue damage
and enhance the effects of radiotherapy (14). Numerous tumor gene therapies have
been investigated in vitro experiments and have exhibited
beneficial effects, such as cellular tumor antigen p53 (P53), which
has successful results in clinical trials, achieving desirable
treatment outcomes (15–17). Retinoblastoma-binding protein 4
(RbAp48) is a member of the WD-40 protein family and was originally
identified as a retinoblastoma protein (Rb) binding protein
(18). E2F transcription factor
(E2F) 1 and RbAp48 interaction is mediated by Rb and histone
deacetylase (HDAC) and results in the inhibition of E2F regulatory
gene transcription, which are important cell cycle regulatory
proteins (19).
The underlying mechanisms of gastric cancer
radiosensitivity remain unclear. The present study aimed to
investigate the effect and underlying mechanisms of RbAp48 on
gastric cancer cell radiosensitivity.
Materials and methods
Cell culture
The human gastric cancer cell line (AGS) was
purchased from Shanghai Gefan Biotechnology Co., Ltd. (Shanghai,
China). The cells were maintained in RPMI-1640 medium (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) in a
37°C incubator with 5% CO2.
Cell transfection and grouping
pcDNA3.1, pcDNA3.1-RbAp48, RbAp48 siRNA and
non-specific scrambled siRNA vectors were obtained from Invitrogen
(Thermo Fisher Scientific, Inc.). The vectors were transfected at a
final concentration of 100 nmol/l transfection (20). AGS cells were transfected with
pcDNA3.1 (mock), pcDNA3.1-RbAp48 (RbAp48), RbAp48 siRNA (si-RbAp48;
5′-CAGGGCATACGGCAGTAGT-3′) and non-specific scrambled siRNA (NC;
5′-ACGUGACACGUUCGGAGAATT-3′) vectors using EndoFectin™ Max
transfection reagent (GeneCopoeia, Inc., Rockville, MD, USA) at
37°C for 48 h. Following transfection, cells were lysed for western
blot analysis and RT-qPCR to verify transfection efficiency.
There were five AGS cell treatment groups: Control
(treated with PBS), mock (treated with pcDNA3.1), control+RAD
(treated with 6 Gy radiation), mock+RAD (treated with pcDNA3.1 and
radiation), and the RbAp48+RAD group (treated with pcDNA3.1-RbAp48
and radiation), in the early stage of the experiment. There were
seven treatment groups in the advanced stage of the experiment,
including: pcDNA3.1 (Mock), pcDNA3.1 and 6 Gy radiation (RAD),
si-RbAp48, si-RbAp48+RAD, pcDNA3.1-RbAp48 (RbAp48), pcDNA3.1-RbAp48
and radiation (RbAp48+RAD), as well as pcDNA3.1-RbAp48, 6 Gy
radiation and 50 ng/ml insulin-like growth factor-1 (IGF-1)
(RbAp48+RAD+IGF-1). IGF-1 was used as an agonist of the
phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway,
and was used to treat cells at 37°C for 48 h.
Cell radiation
Cells were digested with 0.25% trypsin and counted
with a hemocytometer prior to radiation exposure. Cells were
subsequently transferred into new culture bottles, each containing
106 cells. Cells were maintained in the incubator
overnight at 37°C with 5% CO2. Following this, cells
were treated with 0, 2, 4, and 6 Gy 6 MV-X ray at room temperature
for 3 h using a PRIMUS™ linear accelerator (Siemens AG, Munich,
Germany). This was a preliminary experiment to determine the
appropriate dose of radiation for the subsequent experiments.
Cell proliferation analysis
Cell proliferation was determined by an MTT assay.
Cells (6×104/ml) in the logarithmic phase were sowed
into the wells of 96-well plates and incubated for 12 h at 37°C
with 5% CO2. Along with a control group, cells were
treated with PBS, 6 Gy doses of radiation, and 100 nmol/l
pcDNA3.1-RbAp48 and pcDNA3.1 vectors. The cells were subsequently
maintained at 37°C with 5% CO2 for 12, 24 and 48 h. A
volume of 50 µl MTT solution was added into each well, and cells
were transferred to the incubator for 4 h. Subsequently, 100 µl
dimethyl sulfoxide was added into each well. The absorbance was
read at wavelengths of 570 and 630 nm using a microplate reader
(cat. no. SMR16.1; Uscn Life Sciences, Inc., Wuhan, China). Cell
proliferation was determined in terms of the percentage of cell
survival.
Flow cytometry (FCM)
FCM was used for the analysis of the cell apoptosis
and cell cycle. For cell apoptosis analysis, cells were harvested
following transfection for 48 h and fixed in 70% ethanol at room
temperature overnight. Cells prepared for assessment were first
washed with PBS and subsequently resuspended in Annexin
V-fluorescein iosthiocyanate and propidium iodide (PI; Shanghai
Yeasen Biotechnology Co., Ltd., Shanghai, China) at 37°C for 30
min. A flow cytometer (FACSCalibur; BD Biosciences, Franklin Lakes,
NJ, USA) was used to assess cell apoptosis.
Cell cycle analysis
Following digestion with 0.25% trypsin,
~5×107 cells were plated into 6-well plates and
incubated for 24 h at 37°C with 5% CO2. Following
treatment with radiation and mock or RbAp48 vector, the medium was
removed and cells were washed with PBS three times. Cells were
digested with 0.25% trypsin, placed in 15 ml centrifuge tubes and
centrifuged for 5 min at 1,000 × g at 4°C and supernatant was
subsequently discarded. Cells were resuspended following washing
with PBS, and subsequently centrifuged for 5 min at 1,000 × g at
4°C, and the supernatant was discarded. Pre-cooled 70% ethanol
(4°C; 1 ml) was added into cells and the cells were gently blown
with a pipette. Following this, cells were stored in a refrigerator
at 4°C overnight. Cells were centrifuged for 5 min at 1,000 × g at
4°C, ethanol was discarded and cells were washed with PBS three
times. PBS (500 µl) containing PI (50 µg/ml), RNase A (100 µg/ml)
and Triton X-100 (0.2%) was added to the cells, which were
incubated in the dark for 30 min at 4°C. The results were detected
by FCM and cell cycle analysis was performed with FlowJo 10
software (FlowJo LLC, Ashland, OR, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
TRIzol® reagent (Tiangen Biotech Co.,
Ltd., Beijing, China) was used to extract total RNA from cells.
According to the manufacturer's protocols, 2 µg of RNA was used for
cDNA synthesis using the First Strand cDNA Synthesis kit
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). RT-qPCR was
performed using the SYBR Green Premix reagent (TakaraBio, Inc.,
Otsu, Japan) in an ABI 7500 Thermocycler (Thermo Fisher Scientific,
Inc.). The PCR thermocycling conditions were as follows: Initial
denaturation for 10 min at 95°C, 40 cycles of 95°C for 5 sec and
65°C for 31 sec, followed by 95°C for 15 sec, 60°C for 1 min, 95°C
for 15 sec and a final extension at 72°C for 10 min and finally
held at 4°C. β-actin was used as the internal control for
normalization. The primers used for RT-qPCR were listed in Table I.
 | Table I.Sequences of the primers used in
reverse transcription-quantitative polymerase chain reaction. |
Table I.
Sequences of the primers used in
reverse transcription-quantitative polymerase chain reaction.
| Name | NCBI gene ID | Direction | Sequence
(5′-3′) |
|---|
| RbAp48 | 5928 | Forward |
CCTCGACATGGCCTAACAGTG |
|
|
| Reverse |
TCCCCAGGACAAGTCGATGA |
| Cyclin B1 | 891 | Forward |
TCTGCTGGGTGTAGGTCCTT |
|
|
| Reverse |
ACCAATGTCCCCAAGAGCTG |
| P53 | 8273 | Forward |
CCCAGTAGGGACCCATTCATTG |
|
|
| Reverse |
CGGCTGGAGATATTGGGTGA |
| P21 | 1026 | Forward |
TGCCGAAGTCAGTTCCTTGT |
|
|
| Reverse |
CATTAGCGCATCACAGTCGC |
| Caspase-3 | 836 | Forward |
GCGGTTGTAGAAGTTAATAAAGGTA |
|
|
| Reverse |
CATGGCACAAAGCGACTGG |
| Caspase-7 | 840 | Forward |
CTCCAGGGACTATGCGTGC |
|
|
| Reverse |
GAATCCTCAACCCCCTGCTC |
| Caspase-9 | 842 | Forward |
CAGGCCCCATATGATCGAGG |
|
|
| Reverse |
TCGACAACTTTGCTGCTTGC |
| PARP | 142 | Forward |
TTCAACAAGCAGCAAGTGCC |
|
|
| Reverse |
CCTTTGGGGTTACCCACTC |
| Bcl-2 | 596 | Forward |
GGGAGGATTGTGGCCTTCTT |
|
|
| Reverse |
ACTTGTGGCCCAGATAGGCA |
| Bax | 581 | Forward |
GTCTTTTTCCGAGTGGCAGC |
|
|
| Reverse |
GGAGACAGGGACATCAGTCG |
| β-actin | 60 | Forward |
TTCTCAAGATCTGGACAGACG |
|
|
| Reverse |
TGGCAACTTCTTCCTGCAAC |
Western blotting
Treated cells were lysed in radioimmunoprecipitation
lysis and extraction buffer (Thermo Fisher Scientific, Inc.) on
ice. Cells were broken into pieces with an ultrasonic cell
disruptor. Supernatant was collected following centrifugation at
1,000 × g at 4°C for 10 min. According to the manufacturer's
protocols, protein concentration was determined using a
bicinchoninic acid assay reagent (Bio-Rad Laboratories, Inc.
Hercules, CA, USA). An equal quantity of proteins (50 µg/lane) was
separated using 10% SDS-PAGE. Proteins obtained were transferred to
nitrocellulose membranes for 1.5 h. Membranes were blocked in 5%
low-fat dried milk at room temperature for 2 h. The following
primary antibodies were incubated with membranes at 4°C overnight:
Anti-RbAp48 (1:1,000; cat. no. ab1765; Abcam, Cambridge, MA, USA),
anti-cyclin B1 (1:1,000; cat. no. ab72; Abcam), anti-cell division
control protein 2 homolog (Cdc2; 1:2,000; cat. no. ab12568; Abcam),
anti-phosphorylated (p)-Cdc2 (1:1,000; cat. no. ab258965; Abcam),
anti-p-M-phase inducer phosphatase 3 (Cdc25c; 1:500; cat. no.
ab62191), anti-Cdc25c (1:1,000, cat. no. ab32444; Abcam),
anti-cleaved caspase-3 (1:1,000; cat. no. ab2302; Abcam),
anti-cleaved caspase-7 (1:500; cat. no. ab32042; Abcam),
anti-cleaved caspase-9 (1:1,000; cat. no. ab1324; Abcam),
anti-cleaved poly (ADP-ribose) polymerase (PARP; 1:5,000; cat. no.
ab32064; Abcam) anti-p-PI3K (1:1,000; cat. no. ab182651; Abcam),
anti-PI3K (1:1,000; cat. no. ab151549; Abcam) anti-p-Akt (1:5,000;
cat. no. ab81283; Abcam), anti-Akt (1:5,000; cat. no. ab182729;
Abcam) and anti-β-actin (1:1,000; cat. no. ab8227; Abcam) at 4°C.
Rabbit anti-mouse immunoglobulin G (IgG; cat. no. 58802; 1:7,000;
CST Biological Reagents Co. Ltd., Shanghai, China), goat anti-mouse
IgG (cat. no. ab7064; 1:8,000; Abcam) and mouse anti-goat IgG (cat.
no. BA1074; 1:7,000; Invitrogen; Thermo Fisher Scientific, Inc.)
horseradish peroxidase (HRP)-conjugated secondary antibodies were
incubated with the membranes at room temperature for 2 h. Bands
were visualized using enhanced chemiluminescent reagent (EMD
Millipore, Billerica, MA, USA). Images were captured using the
Fujifilm LAS-3000 Imager imaging system (Fuji Photo Film Co., Ltd.,
Tokyo, Japan) and the software used was the LAS-3000 Image Reader
(Fuji Photo Film Co., Ltd.).
Statistical analysis
Results are presented as the mean ± standard error
of mean using SPSS 20.0 (IBM Corp., Armonk, NY, USA). Data were
analyzed by one-way analysis of variance, followed by Tukey's
post-hoc test. Each experiment was repeated three times. P<0.05
was considered to indicate a statistically significant
difference.
Results
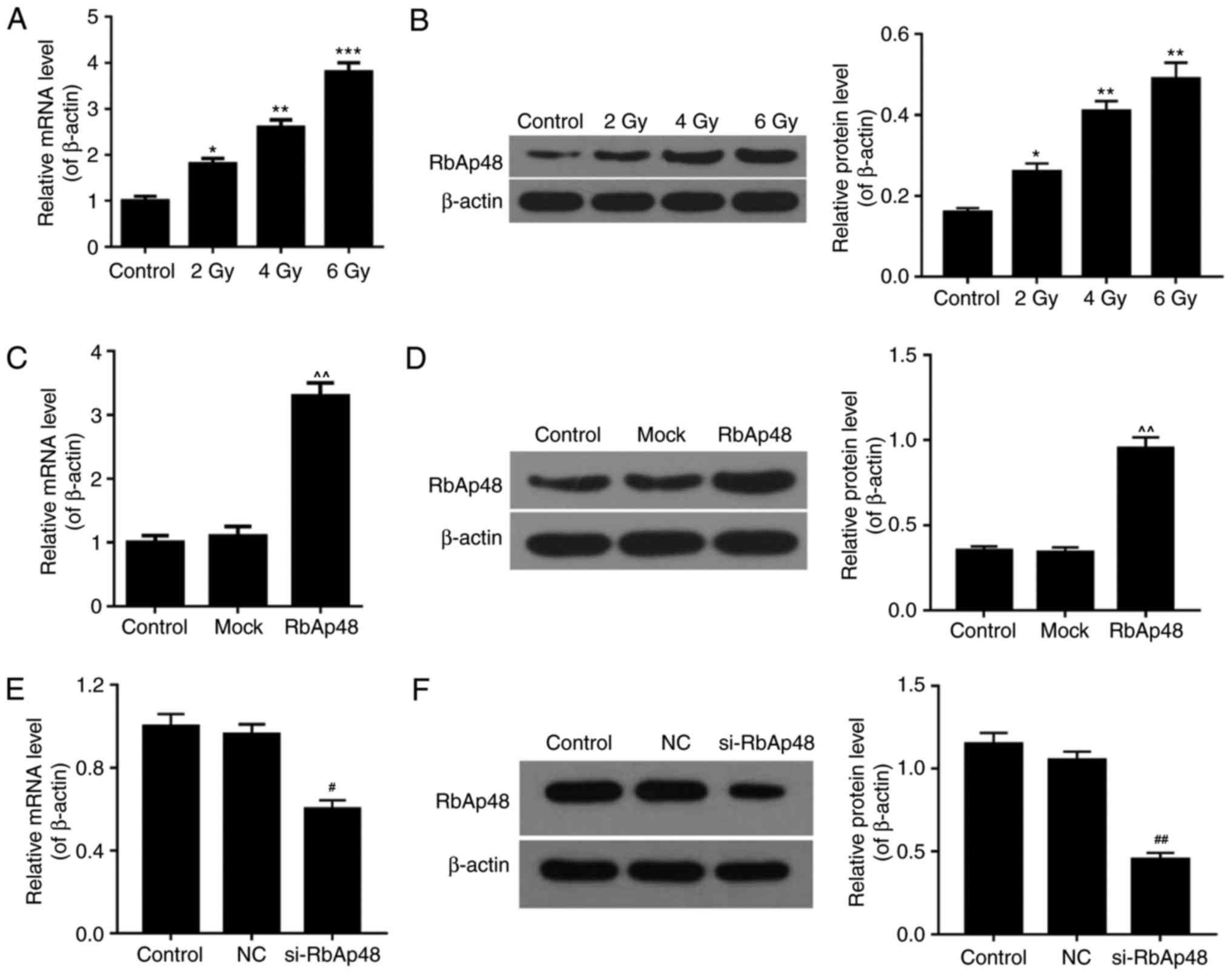
Radiation enhances the expression
level of RbAp48 in AGS cells
RT-qPCR and western blot analysis was performed to
evaluate the expression levels of RbAp48 in AGS cells under various
experimental conditions. AGS cells were treated with increasing
intensities of radiation (2, 4 and 6 Gy). The results revealed that
the mRNA (Fig. 1A) and protein
(Fig. 1B) expression of RbAp48 was
significantly higher in cells exposed to radiation compared with
the control group, in a dose-dependent manner (P<0.05).
Additionally, following AGS cell transfection with pcDNA3.1-RbAp48
plasmids, the mRNA (Fig. 1C) and
protein (Fig. 1D) expression
levels of RbAp48 were significantly higher than control and mock
groups (P<0.05). Furthermore, the mRNA (Fig. 1E) and protein (Fig. 1F) levels of RbAp48 in the si-RbAp48
transfected group were significantly lower, compared with the
control and NC groups (P<0.05). These data demonstrated that
radiation increased RbAp48 expression and that the transfection
efficiency of RbAp48 was high in AGS cells.
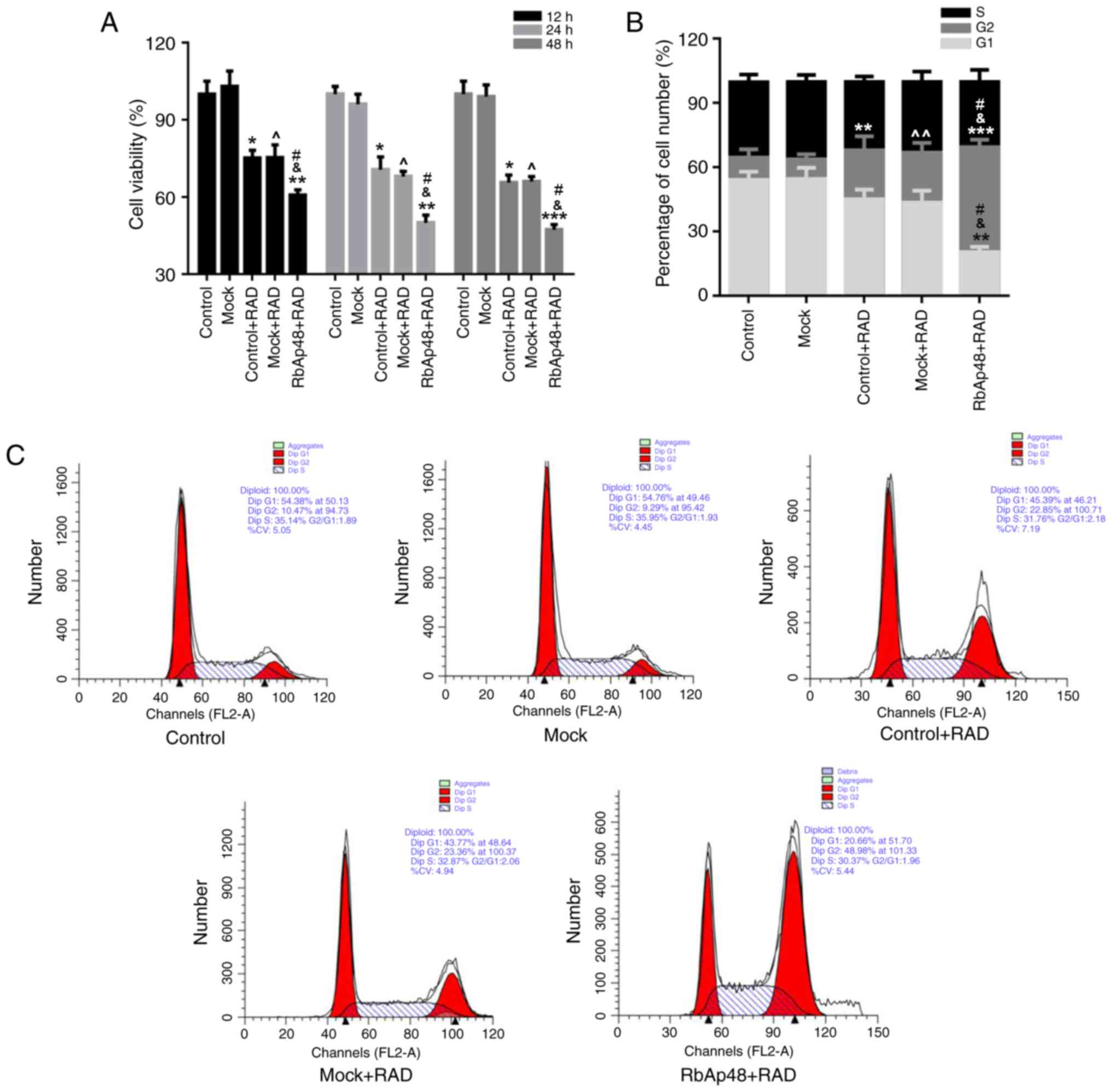
RbAp48 combined with radiation reduces
the cell proliferation of AGS cells
To investigate the effects of radiation and RbAp48
expression on AGS cell proliferation, an MTT assay was performed.
As presented in Fig. 2A, a
significant decrease in cell proliferation was observed in the
control+RAD and mock+RAD groups. The cell proliferation of AGS
cells was further decreased in the groups transfected with RbAp48
at 12, 24 and 48 h (P<0.05). These data indicated that radiation
exposure may have reduced the proliferation of AGS cells, and this
reduction was stronger when combined with RbAp48
overexpression.
RbAp48 overexpression combined with
radiation exposure causes G2 cell cycle arrest
In order to identify the mechanism of AGS cell
growth inhibition with combination of RbAp48 and radiation
exposure, cell cycle analysis was performed. As presented in
Fig. 2B and C, the percentage of
cells in the G2 phase was markedly higher in the
RbAp48+RAD group (48.98%), compared with the other groups (control,
10.47%; Mock, 9.29%; control+RAD, 22.85%; mock+RAD, 23.36%). This
phenomenon indicated that overexpression of RbAp48 in combination
with radiation exposure arrested cells in the G2
phase.
Furthermore, the expression levels of cell cycle
associated factors were evaluated in each group by RT-qPCR and
western blot analysis. It was demonstrated that radiation
significantly reduced the mRNA expression of cyclin B1, and
enhanced the expression of P53 and cyclin dependent kinase
inhibitor (P21; Fig. 3A;
P<0.05). When cells were exposed to a combination of radiation
and RbAp48 overexpression, the downregulation of cyclin B1
expression and upregulation of P53 and P21 expression was more
significant (P<0.01). As presented in Fig. 3B, western blot analysis
demonstrated that compared with the other groups, RbAp48
overexpression combined with radiation significantly decreased the
protein expression of cyclin B1 (P<0.001). Furthermore, in the
RbAp48+RAD group, the protein expression of p-Cdc2 (Tyr15) in AGS
cells was significantly increased (Fig. 3C), and p-Cdc25c expression was
significantly reduced (Fig. 3D;
P<0.01). Based on the aforementioned results, it was concluded
that RbAp48 overexpression combined with radiation exposure
resulted in G2 cell cycle arrest, through regulation of
associated cell cycle factor expression levels.
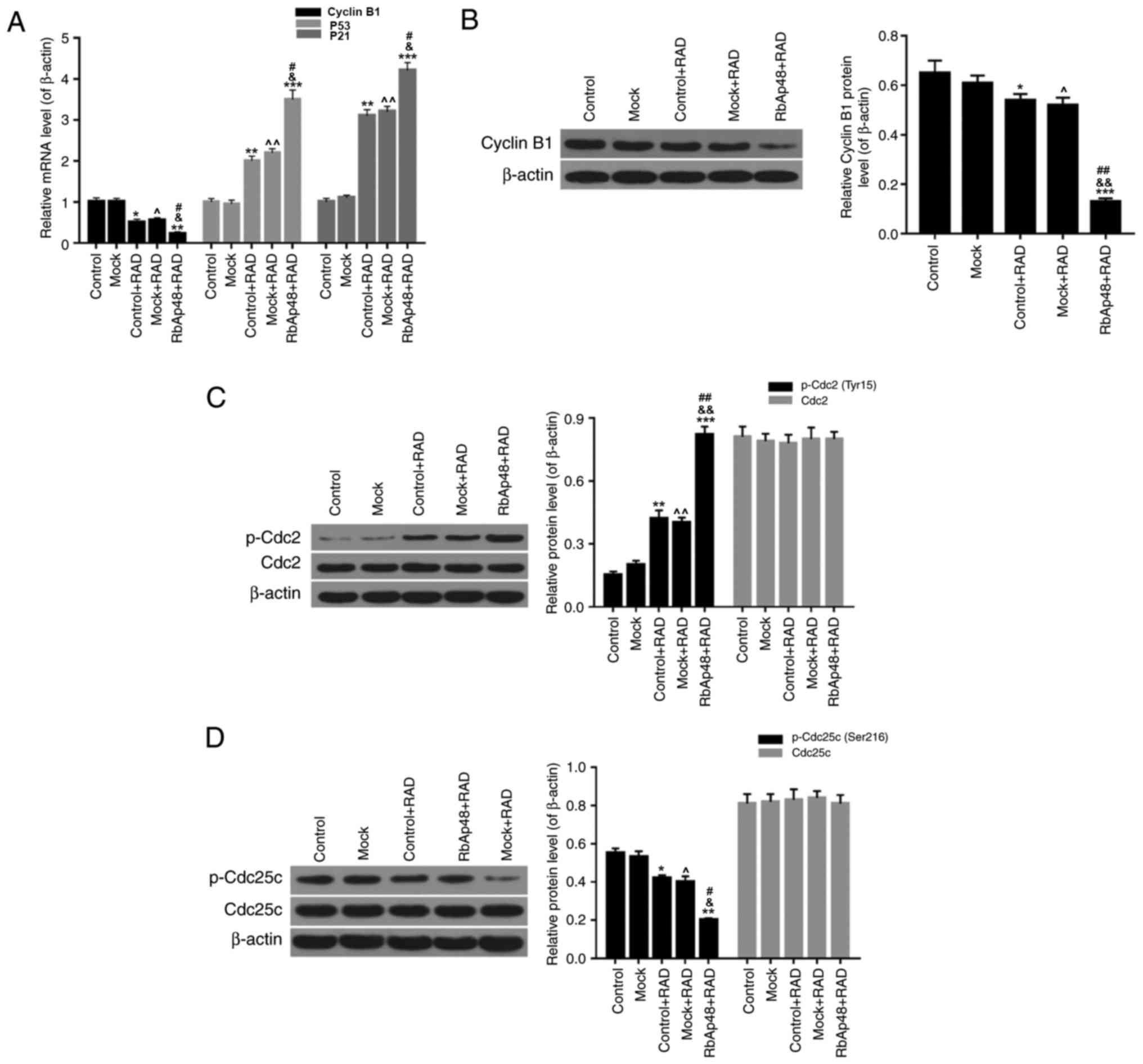 | Figure 3.RbAp48 in combination with radiation
regulates the expression of cyclin B1, P53, P21, Cdc2 and Cdc25c in
AGS cells. (A) Reverse transcription-quantitative polymerase chain
reaction was performed to determine the expression levels of cyclin
B1, P53 and P21 in AGS cells. Western blot analysis was performed
to evaluate the expression levels of (B) cyclin B1, (C) p-Cdc2/Cdc2
and (D) p-Cdc25c/Cdc25c. *P<0.05, **P<0.01, ***P<0.001 vs.
control; ^P<0.05, ^^P<0.01 vs. mock;
&P<0.05, &&P<0.01 vs.
control+RAD; #P<0.05, ##P<0.01 vs.
mock+RAD. RbAp48, retinoblastoma-binding protein 4; control, PBS;
mock, pcDNA3.1; RAD, 6 Gy radiation; P53, cellular tumor antigen
p53; P21, cyclin dependent kinase inhibitor 1A; p-, phosphorylated;
Cdc2, cell division control protein 2 homolog; Cdc25c, M-phase
inducer phosphatase 3. |
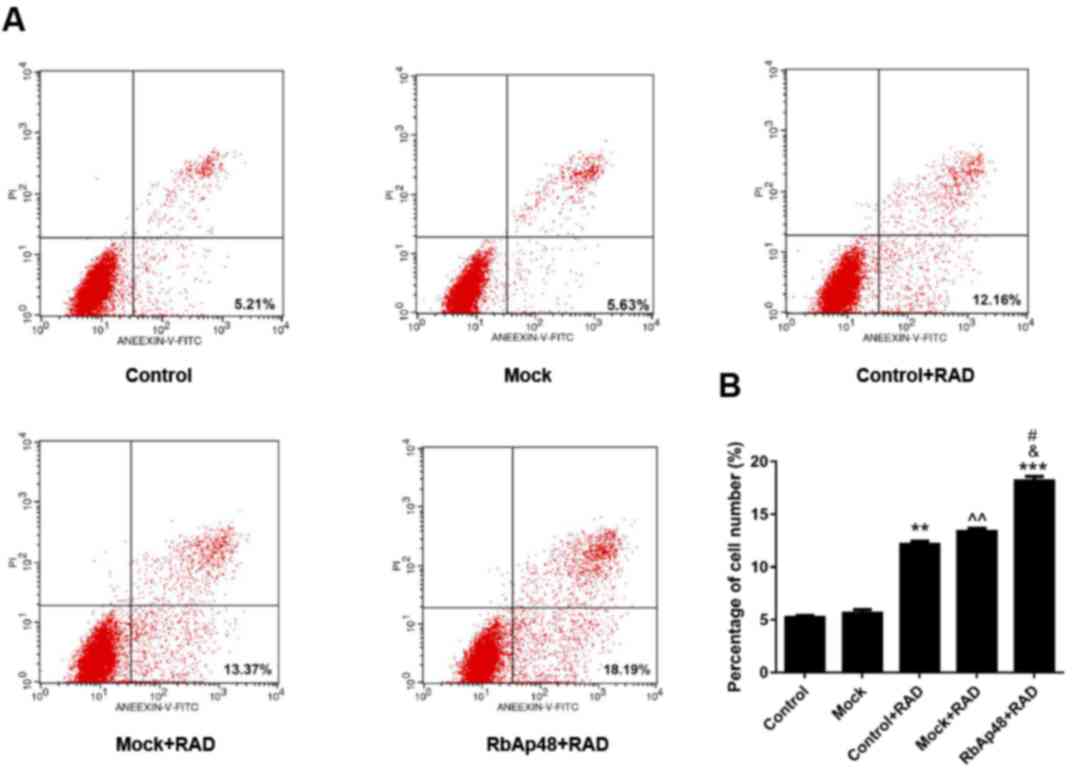
RbAp48 combined with radiation induces
AGS cell apoptosis
In order to determine the effect of RbAp48 and
radiation on cell apoptosis, FCM analysis was performed in the
present study. It was demonstrated that the percentage of apoptotic
cells was significantly increased in the control+RAD and mock+RAD
groups compared with the control (P<0.01), indicating that
radiation induced AGS cell apoptosis. Additionally, the percentage
of apoptosis in the RbAp48+RAD group was significantly higher than
the other groups (Fig. 4). These
data suggested that RbAp48 in combination with radiation further
induced AGS cell apoptosis.
The expression levels of associated apoptosis
proteins were presented in Fig. 5.
With regards to the RT-qPCR data (Fig.
5A), the expression levels of caspase-3, caspase-7, caspase-9,
PARP and Bax were significantly higher (P<0.01), while the
expression level of Bcl-2 was significantly lower (P<0.05) in
the RAD groups compared with the control. In addition, expression
levels of caspase-3, caspase-7, caspase-9, PARP and Bax were
further increased and Bcl-2 expression was further decreased in the
RbAp48+RAD group, compared with the control+RAD and mock+RAD
groups. Furthermore, the western blotting data of caspase-3,
cleaved caspase-7, cleaved caspase-9, and cleaved PARP expression
displayed a similar trend (Fig.
5B). Therefore, it was demonstrated that RbAp48 combined with
radiation induced AGS cell apoptosis through regulation of
associated apoptosis marker expression.
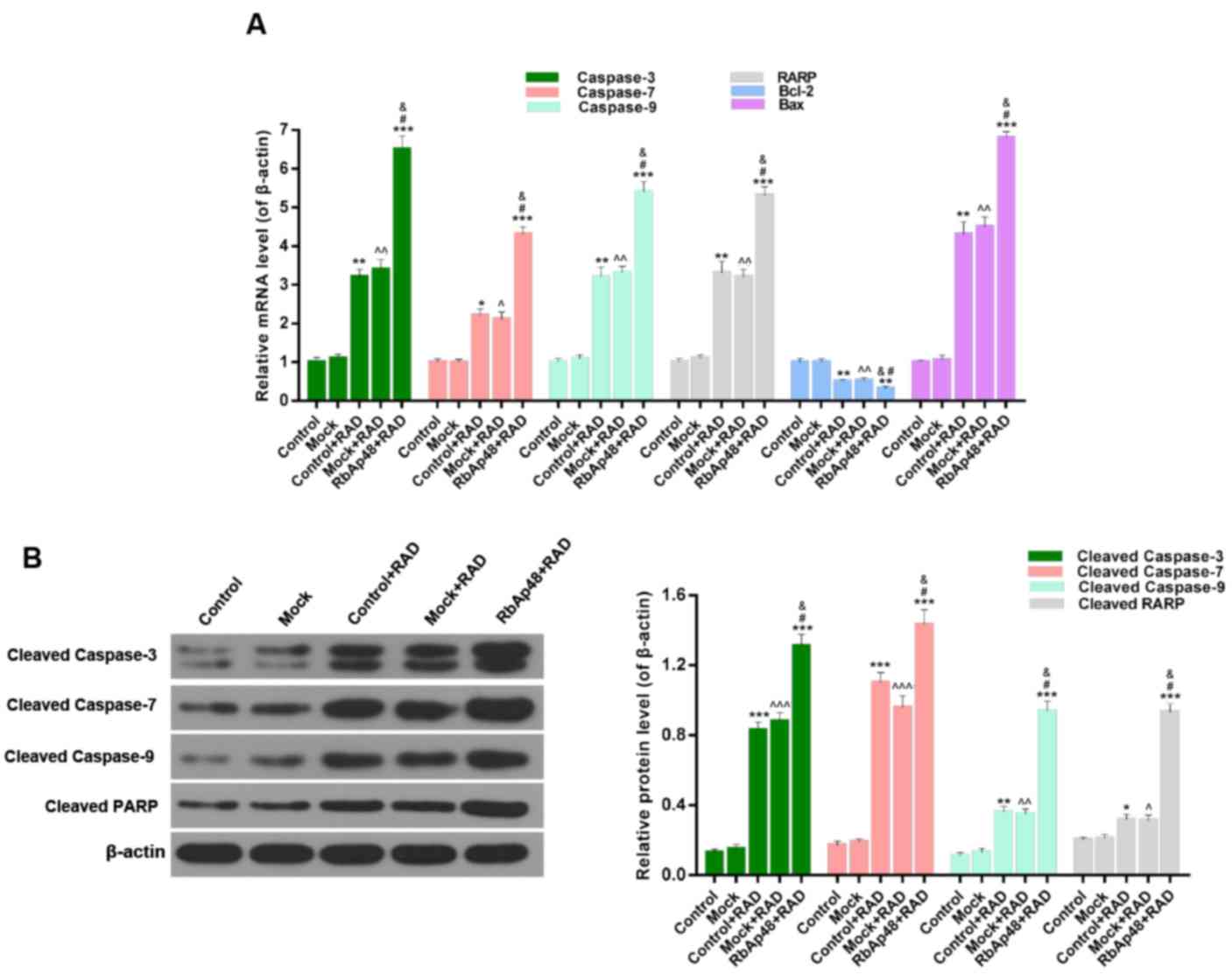 | Figure 5.RbAp48 in combination with radiation
regulates the expression of apoptosis-associated proteins. (A)
Reverse transcription-quantitative polymerase chain reaction was
performed to assess the expression levels of caspase-3, −7, −9,
PARP, Bcl-2 and Bax in AGS cells. (B) Western blot analysis was
performed to evaluate the expression levels of cleaved caspase-3,
−7, −9 and cleaved PARP. *P<0.05, **P<0.01, ***P<0.001 vs.
control; ^P<0.05, ^^P<0.01,
^^^P<0.001 vs. mock; &P<0.05 vs.
control+RAD; #P<0.05 vs. mock+RAD. RbAp48,
retinoblastoma-binding protein 4; control, PBS; mock, pcDNA3.1;
RAD, 6 Gy radiation; PARP, poly(ADP-ribose) polymerase 1; Bcl-2,
B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
RbAp48 combined with radiation
inhibits the PI3K/Akt pathway
Western blotting was also performed to explore the
functional mechanism of RbAp48 in combination with radiation in AGS
cell growth inhibition. The expression levels of p-PI3K, PI3K,
p-Akt, and Akt were detected. In the RAD groups, the protein
expression levels of p-PI3K were significantly lower than the
control and mock groups. However, the expression levels of total
PI3K were not significantly different among the five groups
(Fig. 6A). Additionally, compared
with the control and mock groups, protein expression levels of
p-Akt were markedly reduced by radiation, while the expression
levels of total Akt were not significantly different among the five
groups (Fig. 6B). Furthermore,
overexpression of RbAp48 resulted in further reductions in p-PI3K
and p-Akt expression, compared with the radiation only groups
(P<0.01). Therefore, it was confirmed that RbAp48 combined with
radiation affected the PI3K/Akt pathway activity, which may have
been involved in AGS cell growth inhibition and induction of AGS
cell apoptosis.
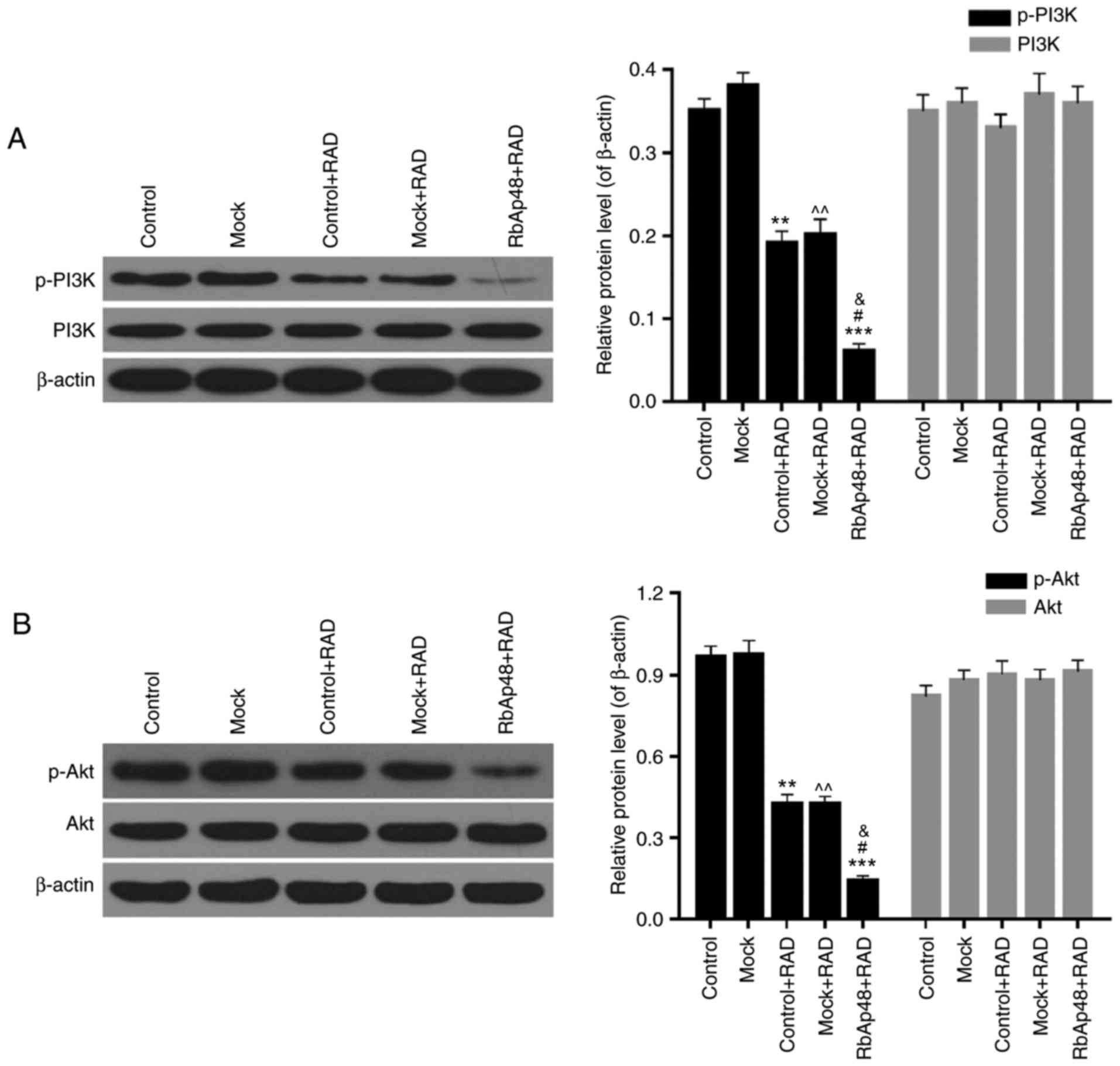 | Figure 6.RbAp48 in combination with radiation
inhibits the PI3K/Akt pathway. Western blot analysis was performed
to assess the expression levels of (A) p-PI3K, PI3K, (B) p-Akt and
Akt in AGS cells. **P<0.01, ***P<0.001 vs. control;
^^P<0.01 vs. mock; &P<0.05 vs.
control+RAD; #P<0.05 vs. mock+RAD. RbAp48,
retinoblastoma-binding protein 4; control, PBS; mock, pcDNA3.1;
RAD, 6 Gy radiation; p-, phosphorylated; PI3K, phosphoinositide
3-kinase; Akt, protein kinase B. |
si-RbAp48 represses cell apoptosis and
RbAp48 in combination with radiation promotes cell apoptosis via
PI3K/Akt pathway inhibition
FCM analysis was performed in order to analyze the
effect of si-RbAp48 on cell apoptosis, and to determine if the
combination of RbAp48 and radiation induced apoptosis via PI3K/Akt
pathway analysis. The results revealed that compared with the mock
group, the apoptotic cell number in the si-RbAp48 group was
reduced, and was significantly increased in the RbAp48
overexpression group. In addition, cells treated with a combination
of RbAp48 and radiation had the highest proportion of apoptotic
cells. Furthermore, compared with the RbAp48+RAD group, the number
of apoptotic cells in the RbAp48+RAD+IGF-1 group was significantly
lower (P<0.05; Fig. 7).
Therefore, it was demonstrated that si-RbAp48 repressed cell
apoptosis, and RbAp48 in combination with radiation promoted cell
apoptosis via PI3K/Akt pathway inhibition.
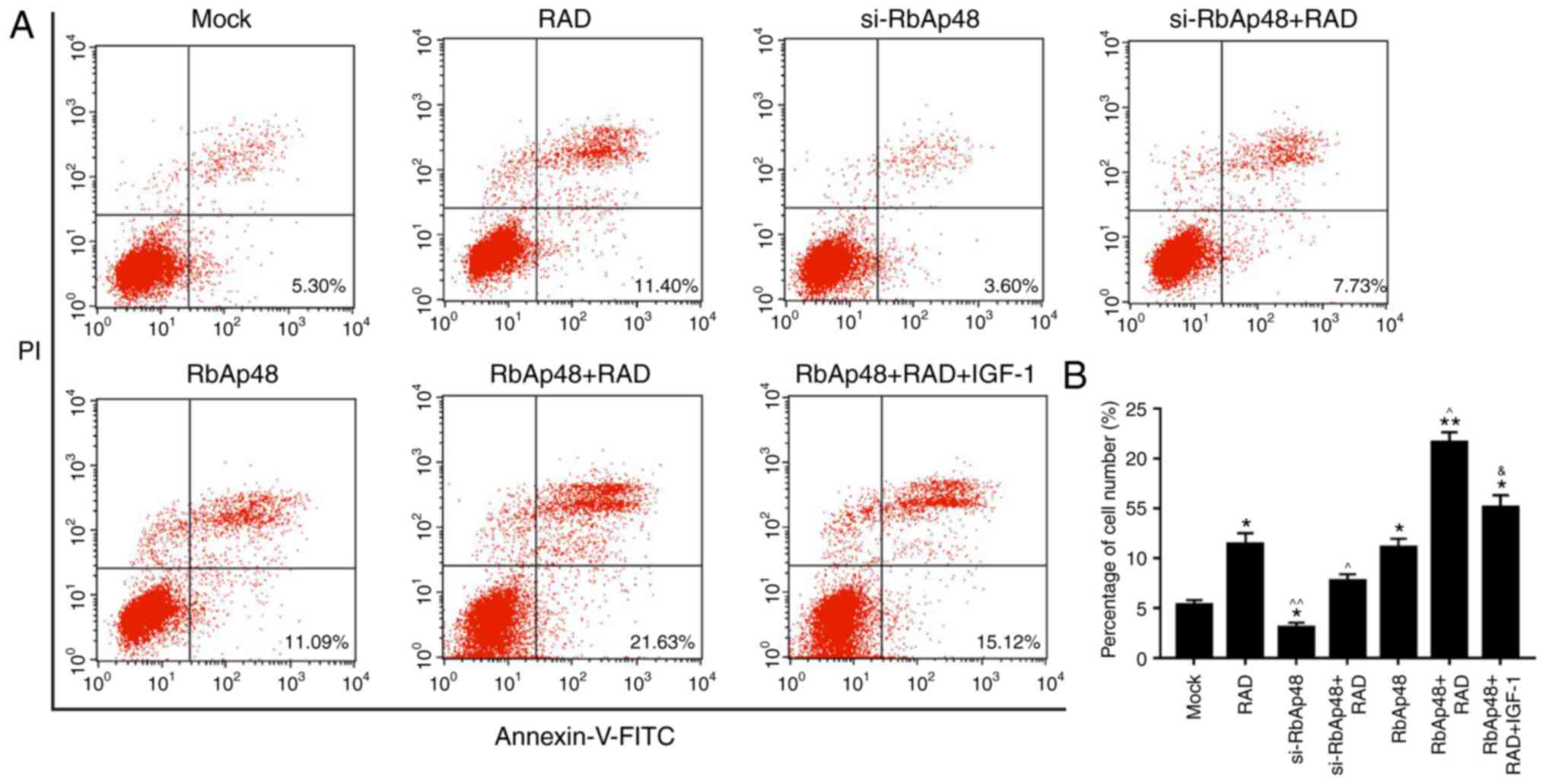 | Figure 7.si-RbAp48 represses cell apoptosis,
and RbAp48 in combination with radiation promotes cell apoptosis
via PI3K/Akt pathway inhibition. (A) Flow cytometry analysis was
performed and (B) the percentage of apoptotic cells was calculated.
IGF-1 was used as a PI3K/Akt pathway agonist. *P<0.05,
**P<0.01 vs. mock; ^P<0.05, ^^P<0.01
vs. RAD; &P<0.05 vs. RbAp48+RAD. RbAp48,
retinoblastoma-binding protein 4; si-RbAp48, RbAp48 small
interfering RNA; mock, pcDNA3.1; RAD, 6 Gy radiation; IGF-1,
insulin-like growth factor 1; PI3K, phosphoinositide 3-kinase; Akt,
protein kinase B; FITC, fluorescein isothiocyanate; PI, propidium
iodide. |
Discussion
RbAp48, as a binding protein of tumor suppressor
protein Rb, is involved in various biological functions, including
chromatin assembly, histone modification and nucleosome remodeling
(21,22). Studies have demonstrated that the
Rb/RbAp48-associatedhistone acetyl transferase complex participates
in the inhibition of E2F regulatory gene transcription (23–27).
It has been reported that RbAp48 promotes radiosensitivity in
breast, melanoma and cervical cancer cells (28,29).
However, the effects of RbAp48 in gastric cancer remain
unclear.
AGS gastric cancer cells were used in the present
study. Radiation treatment was administered and the mRNA and
protein expression of RbAp48 had significantly increased in the AGS
cells, suggesting that RbAp48 is a radiation-inducible protein in
gastric cancer. This finding provided the basis for further
investigation. Furthermore, compared with the cells treated with
radiation only, cell proliferation was significantly inhibited in
cells overexpressing RbAp48 in addition to radiation exposure. The
mechanism underlying this phenomenon may be associated with the
inhibition of oncogene expression and increased cancer suppressor
gene expression (30). In the
present study, gastric cancer cell proliferation was significantly
inhibited following treatment with RbAp48 in combination with
radiation. However, the mechanism underlying the inhibition of AGS
cell proliferation by this combination may be complex. The present
preliminary study suggested that RbAp48 arrested the AGS cells at
the G2 phase and induced apoptosis. These processes may
serve a critical role in RbAp48-induced radiosensitivity.
Radiation-induced cell responses include cell cycle
arrest, apoptosis and DNA repair, as a complex result of multiple
gene involvement (31). In
response to radiation-induced DNA impairment, DNA repair,
G2 phase arrest and apoptosis predominantly occur
(32,33). In the present study, it was
demonstrated that the G2 phase arrest was increased by
radiation, and this effect was further increased by RbAp48
overexpression. Typically, the G1 and G2
phases are more sensitive to radiation, whereas the S phase is the
least sensitive. Thus, the G2 phase is considered a key
point in cancer radiotherapy (31). Previous research has reported that
RbAp48 may enhance the proportion of HS-578T cells in the
G2 phase (28), which
was in accordance with the results of the present study.
Radiation leads to apoptosis in cancer cells,
indicating that apoptosis is a key mechanism in tumor cell
radiotherapy. The detection and signaling system of DNA impairment
controls cell cycle checkpoints and results in subsequent cell
death. The present study revealed that overexpression of RbAp48
significantly increased the apoptotic rate in combination with
radiotherapy. Additionally, RbAp48 silencing markedly attenuated
apoptosis in radiotherapy-treated AGS cells. These results
demonstrated that RbAp48 is a radiotherapy-induced protein in
gastric cancer cells, which promoted the radiosensitivity of
gastric cancer cells through increased cell apoptosis.
Cell cycle arrest, DNA repair and apoptosis
initially occur in response to DNA impairment. Cyclin B1, Cdc2 and
Cdc25c are critical to the radiographic response and mediate the
cell cycle when DNA is impaired (34–38).
The P53 gene is a tumor suppressor gene (39), and the P21 gene is an important
member of the cyclin dependent kinase inhibitor family (40). P53 and P21 coordinate the
association between cell cycle, DNA replication and repair, and
thus closely associate tumor inhibition with cell cycle control
(39–42). The present results demonstrated
that overexpression of RbAp48 in combination with radiation
significantly reduced the expression of cyclin B1 and p-Cdc25c, and
enhanced the expression of p-Cdc2, P53 and P21.
Caspases are key elements in the apoptotic pathway
and participate in apoptosis signal transduction and execution.
Research has indicated that caspase-3, −7 and −9 are involved in
radiation-induced cell apoptosis (43,44).
In the present study, overexpression of RbAp48 in combination with
radiotherapy markedly enhanced AGS cell apoptosis. Furthermore,
RT-qPCR and western blotting data indicated that overexpression of
RbAp48 in combination with radiation enhanced the expression levels
of cleaved caspase-3, −7, −9 and cleaved PARP.
Furthermore, the functional mechanism underlying
RbAp48 in combination with radiation in increasing AGS cell
apoptosis was explored. PI3K is a class of kinases that generates
second messenger molecules via activation of its substrates PIP2
and PIP3 (45). As a downstream
effector of PI3K, Akt is a critical tumor factor (46). Activated Akt may regulate specific
families of proteins (including the Bcl-2 family, caspase family
and IKK family) associated with apoptosis and participate in the
regulation of biological behavior of tumor cells (47–49).
Previous studies have demonstrated that apoptosis is induced in
gastric cancer cells via the PI3K/Akt pathway (50,51).
Therefore, it was hypothesized that AGS cell apoptosis, induced by
RbAp48 overexpression in combination with radiation, was associated
with PI3K/Akt pathway regulation. In the present study, the results
of the RT-qPCR and western blot analysis demonstrated that
overexpression of RbAp48 in combination with radiation
significantly downregulated the expression levels of p-PI3K and
p-Akt. In addition, the percentage of apoptotic cells in the
RbAp48+RAD+IGF-1 group was significantly lower than that of the
RbAp48+RAD group. Therefore, it was confirmed that overexpression
of RbAp48 in combination with radiation increased apoptosis via
inhibition of PI3K/Akt pathway activity.
Radiotherapy is one of the major treatment options
for malignant tumors. Though tumor cells cannot be completely
removed by surgery, surgery in combination with radiotherapy
increases the possibility of patient survival (52,53).
However, certain tumor cells (glioma and pancreatic tumor) are not
sensitive to radiation, which consequently results in treatment
failure (54,55). Therefore, improving
radiosensitivity is key to improving the clinical outcomes of
radiotherapy. Current clinical radiotherapy sensitization drugs,
including cisplatin, 5-fluorouracil and gemcitabin, are effective;
however, these drugs may additionally result in damaging effects in
normal tissues (56,57). Therefore, developing specific
radiation-sensitive agents for tumors should be the direction of
future radiotherapy research.
In conclusion, overexpression of RbAp48 in
combination with radiation arrested AGS cells in G2
phase and induced apoptosis via regulation of the PI3K/Akt pathway.
The present study established an experimental basis for clinical
radiotherapy in combination with gene therapy to enhance
radiosensitivity in gastric cancer. The results suggest that RbAp48
may be a potential genetic therapeutic target in gastric cancer.
However, the specific effects of RbAp48 overexpression in healthy
tissues remain unknown and require further investigation.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated and analyzed during this study
are included in this published article.
Authors' contributions
XJ, RJ, YX, BY and YY made substantial contributions
to the conception and designed the experimental scheme. ZF, ZW, LY
and SW analyzed and interpreted data. XJ was a principal
contributor in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declared that they had no competing
interests.
References
|
1
|
Danaei G, Vander Hoorn S, Lopez AD, Murray
CJ and Ezzati M: Comparative Risk Assessment collaborating group
(Cancers): Causes of cancer in the world: Comparative risk
assessment of nine behavioural and environmental risk factors.
Lancet. 366:1784–1793. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guggenheim DE and Shah MA: Gastric cancer
epidemiology and risk factors. J Surg Oncol. 107:230–236. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang X, Tseng CC, Bernstein L and Wu AH:
Family history of cancer and gastroesophageal disorders and risk of
esophageal and gastric adenocarcinomas: A case-control study. BMC
Cancer. 14:602014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yaghoobi M, Bijarchi R and Narod SA:
Family history and the risk of gastric cancer. Br J Cancer.
102:237–242. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Catalano V, Labianca R, Beretta GD, Gatta
G, de Braud F and Van Cutsem E: Gastric cancer. Crit Rev Oncol
Hematol. 71:127–164. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wright JD and Herzog TJ: Human
papillomavirus: Emerging trends in detection and management. Curr
Womens Health Rep. 2:259–265. 2002.PubMed/NCBI
|
|
9
|
Franco EL: Epidemiology of anogenital
warts and cancer. Obstet Gynecol Clin North Am. 23:597–623.
1996.PubMed/NCBI
|
|
10
|
Chun-Zhi Z, Lei H, An-Ling Z, Yan-Chao F,
Xiao Y, Guang-Xiu W, Zhi-Fan J, Pei-Yu P, Qing-Yu Z and Chun-Sheng
K: MicroRNA-221 and microRNA-222 regulate gastric carcinoma cell
proliferation and radioresistance by targeting PTEN. BMC Cancer.
10:3672010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hamada M, Fujiwara T, Hizuta A, Gochi A,
Naomoto Y, Takakura N, Takahashi K, Roth JA, Tanaka N and Orita K:
The p53 gene is a potent determinant of chemosensitivity and
radiosensitivity in gastric and colorectal cancers. J Cancer Res
Clin Oncol. 122:360–365. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qiu H, Yashiro M, Shinto O, Matsuzaki T
and Hirakawa K: DNA methyltransferase inhibitor 5-aza-CdR enhances
the radiosensitivity of gastric cancer cells. Cancer Sci.
100:181–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Furuyama T, Tie F and Harte PJ: Polycomb
group proteins ESC and E(Z) are present in multiple distinct
complexes that undergo dynamic changes during development. Genesis.
35:114–124. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Müller J, Hart CM, Francis NJ, Vargas ML,
Sengupta A, Wild B, Miller EL, O'Connor MB, Kingston RE and Simon
JA: Histone methyltransferase activity of a Drosophila Polycomb
group repressor complex. Cell. 111:197–208. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Döhner H, Fischer K, Bentz M, Hansen K,
Benner A, Cabot G, Diehl D, Schlenk R, Coy J, Stilgenbauer S, et
al: p53 gene deletion predicts for poor survival and non-response
to therapy with purine analogs in chronic B-cell leukemias. Blood.
85:1580–1589. 1995.PubMed/NCBI
|
|
16
|
Eastham JA, Hall SJ, Sehgal I, Wang J,
Timme TL, Yang G, Connell-Crowley L, Elledge SJ, Zhang WW, Harper
JW, et al: In vivo gene therapy with p53 or p21 adenovirus for
prostate cancer. Cancer Res. 55:5151–5155. 1995.PubMed/NCBI
|
|
17
|
Lang FF, Bruner JM, Fuller GN, Aldape K,
Prados MD, Chang S, Berger MS, McDermott MW, Kunwar SM, Junck LR,
et al: Phase I trial of adenovirus-mediated p53 gene therapy for
recurrent glioma: Biological and clinical results. J Clin Oncol.
21:2508–2518. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Parthun MR, Widom J and Gottschling DE:
The major cytoplasmic histone acetyltransferase in yeast: Links to
chromatin replication and histone metabolism. Cell. 87:85–94. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Loyola A and Almouzni G: Histone
chaperones, a supporting role in the limelight. Biochim Biophys
Acta. 1677:3–11. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Scuto A, Zhang H, Zhao H, Rivera M,
Yeatman TJ, Jove R and Torres-Roca JF: RbAp48 regulates
cytoskeletal organization and morphology by increasing K-Ras
activity and signaling through mitogen-activated protein kinase.
Cancer Res. 67:10317–10324. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brehm A, Miska EA, McCance DJ, Reid JL,
Bannister AJ and Kouzarides T: Retinoblastoma protein recruits
histone deacetylase to repress transcription. Nature. 391:597–601.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Luo RX, Postigo AA and Dean DC: Rb
interacts with histone deacetylase to repress transcription. Cell.
92:463–473. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bae SM, Lee CH, Cho YL, Nam KH, Kim YW,
Kim CK, Han BD, Lee YJ, Chun HJ and Ahn WS: Two-dimensional gel
analysis of protein expression profile in squamous cervical cancer
patients. Gynecol Oncol. 99:26–35. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng Q, Lau WM, Tay SK, Chew SH, Ho TH
and Hui KM: Identification and characterization of genes involved
in the carcinogenesis of human squamous cell cervical carcinoma.
Int J Cancer. 98:419–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pardo M, García A, Thomas B, Piñeiro A,
Akoulitchev A, Dwek RA and Zitzmann N: Proteome analysis of a human
uveal melanoma primary cell culture by 2-DE and MS. Proteomics.
5:4980–4993. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Verreault A, Kaufman PD, Kobayashi R and
Stillman B: Nucleosome assembly by a complex of CAF-1 and
acetylated histones H3/H4. Cell. 87:95–104. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xue Y, Wong J, Moreno GT, Young MK, Côté J
and Wang W: NURD, a novel complex with both ATP-dependent
chromatin-remodeling and histone deacetylase activities. Mol Cell.
2:851–861. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Torres-Roca JF, Eschrich S, Zhao H, Bloom
G, Sung J, McCarthy S, Cantor AB, Scuto A, Li C, Zhang S, et al:
Prediction of radiation sensitivity using a gene expression
classifier. Cancer Res. 65:7169–7176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng L, Tang W, Wei F, Wang H, Liu J, Lu
Y, Cheng Y, Bai X, Yu X and Zhao W: Radiation-inducible protein
RbAp48 contributes to radiosensitivity of cervical cancer cells.
Gynecol Oncol. 130:601–608. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kong L, Yu XP, Bai XH, Zhang WF, Zhang Y,
Zhao WM, Jia JH, Tang W, Zhou YB and Liu CJ: RbAp48 is a critical
mediator controlling the transforming activity of human
papillomavirus type 16 in cervical cancer. J Biol Chem.
282:26381–26391. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Teyssier F, Bay JO, Dionet C and Verrelle
P: Cell cycle regulation after exposure to ionizing radiation. Bull
Cancer. 86:345–357. 1999.PubMed/NCBI
|
|
32
|
Iliakis G, Wang Y, Guan J and Wang H: DNA
damage checkpoint control in cells exposed to ionizing radiation.
Oncogene. 22:5834–5847. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miyata H, Doki Y, Yamamoto H, Kishi K,
Takemoto H, Fujiwara Y, Yasuda T, Yano M, Inoue M, Shiozaki H, et
al: Overexpression of CDC25B overrides radiation-induced G2-M
arrest and results in increased apoptosis in esophageal cancer
cells. Cancer Res. 61:3188–3193. 2001.PubMed/NCBI
|
|
34
|
Bulavin DV, Higashimoto Y, Popoff IJ,
Gaarde WA, Basrur V, Potapova O, Appella E and Fornace AJ Jr:
Initiation of a G2/M checkpoint after ultraviolet radiation
requires p38 kinase. Nature. 411:102–107. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jin P, Gu Y and Morgan DO: Role of
inhibitory CDC2 phosphorylation in radiation-induced G2 arrest in
human cells. J Cell Biol. 134:963–970. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kao GD, McKenna WG, Maity A, Blank K and
Muschel RJ: Cyclin B1 availability is a rate-limiting component of
the radiation-induced G2 delay in HeLa cells. Cancer Res.
57:753–758. 1997.PubMed/NCBI
|
|
37
|
Maity A, McKenna WG and Muschel RJ:
Evidence for post-transcriptional regulation of cyclin B1 mRNA in
the cell cycle and following irradiation in HeLa cells. EMBO J.
14:603–609. 1995.PubMed/NCBI
|
|
38
|
Porter LA, Singh G and Lee JM: Abundance
of cyclin B1 regulates gamma-radiation-induced apoptosis. Blood.
95:2645–2650. 2000.PubMed/NCBI
|
|
39
|
Sui X, Cai J, Li H, He C, Zhou C, Dong Y,
Chen L, Zhang B, Wang Y, Zhang Y, et al: p53-dependent CD51
expression contributes to characteristics of cancer stem cells in
prostate cancer. Cell Death Dis. 9:5232018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pérez-Yépez EA, Saldívar-Cerón HI,
Villamar-Cruz O, Pérez-Plasencia C and Arias-Romero LE: p21
Activated kinase 1: Nuclear activity and its role during DNA damage
repair. DNA Repair (Amst). 65:42–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Galanos P, Pappas G, Polyzos A, Kotsinas
A, Svolaki I, Giakoumakis NN, Glytsou C, Pateras IS, Swain U,
Souliotis VL, et al: Mutational signatures reveal the role of RAD52
in p53-independent p21-driven genomic instability. Genome Biol.
19:372018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Y, Qiu C, Lu N, Liu Z, Jin C, Sun C,
Bu H, Yu H, Dongol S and Kong B: FOXD1 is targeted by miR-30a-5p
and miR-200a-5p and suppresses the proliferation of human ovarian
carcinoma cells by promoting p21 expression in a p53-independent
manner. Int J Oncol. 52:2130–2142. 2018.PubMed/NCBI
|
|
43
|
Kim KW, Moretti L, Mitchell LR, Jung DK
and Lu B: Endoplasmic reticulum stress mediates radiation-induced
autophagy by perk-eIF2alpha in caspase-3/7-deficient cells.
Oncogene. 29:3241–3251. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kuida K, Haydar TF, Kuan CY, Gu Y, Taya C,
Karasuyama H, Su MS, Rakic P and Flavell RA: Reduced apoptosis and
cytochrome c-mediated caspase activation in mice lacking caspase 9.
Cell. 94:325–337. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Franke TF, Kaplan DR and Cantley LC: PI3K:
Downstream AKTion blocks apoptosis. Cell. 88:435–437. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Larue L and Bellacosa A:
Epithelial-mesenchymal transition in development and cancer: Role
of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene.
24:7443–7454. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sun G, Wang X, Li T, Qu S and Sun J:
Taurine attenuates acrylamide-induced apoptosis via a
PI3K/AKT-dependent manner. Hum Exp Toxicol. Jan 1–2018.(Epub ahead
of print). View Article : Google Scholar
|
|
48
|
Weng HY, Hsu MJ, Wang CC, Chen BC, Hong
CY, Chen MC, Chiu WT and Lin CH: Zerumbone suppresses IKKa, Akt,
and FOXO1 activation, resulting in apoptosis of GBM 8401 cells. J
Biomed Sci. 19:862012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu DD, Gao YR, Li T, Wang DY, Lu D, Liu
SY, Hong Y, Ning HB, Liu JP, Shang J, et al: PEST-containing
nuclear protein mediates the proliferation, migration, and invasion
of human neuroblastoma cells through MAPK and PI3K/AKT/mTOR
signaling pathways. BMC Cancer. 18:4992018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kwon MJ and Nam TJ: A polysaccharide of
the marine alga Capsosiphon fulvescens induces apoptosis in AGS
gastric cancer cells via an IGF-IR-mediated PI3K/Akt pathway. Cell
Biol Int. 31:768–775. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li D, Qu X, Hou K, Zhang Y, Dong Q, Teng
Y, Zhang J and Liu Y: PI3K/Akt is involved in bufalin-induced
apoptosis in gastric cancer cells. Anticancer Drugs. 20:59–64.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lukoseviciene V, Tikuisis R, Dulskas A,
Miliauskas P and Ostapenko V: Surgery for triple-negative breast
cancer-does the type of anaesthesia have an influence on oxidative
stress, inflammation, molecular regulators, and outcomes of
disease? J BUON. 23:290–295. 2018.PubMed/NCBI
|
|
53
|
Wei X, Liu M, Ding Y, Li Q, Cheng C, Zong
X, Yin W, Chen J and Gu W: Setup errors and effectiveness of
Optical Laser 3D Surface imaging system (Sentinel) in postoperative
radiotherapy of breast cancer. Sci Rep. 8:72702018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang Y, Xu H, Liu T, Huang M, Butter PP,
Li C, Zhang L, Kao GD, Gong Y, Maity A, et al: Temporal DNA-PK
activation drives genomic instability and therapy resistance in
glioma stem cells. JCI Insight. 3:pii: 98096. 2018. View Article : Google Scholar
|
|
55
|
Zhou P, Li B, Liu F, Zhang M, Wang Q, Liu
Y, Yao Y and Li D: The epithelial to mesenchymal transition (EMT)
and cancer stem cells: Implication for treatment resistance in
pancreatic cancer. Mol Cancer. 16:522017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tang X, Hu YJ, Ju WT, Fu Y, Sun WW, Liu Y,
Tan YR, Wang LZ, Li J, Tu YY, et al: Elevated growth
differentiating factor 15 expression predicts long-term benefit of
docetaxel, cisplatin and 5-fluorouracil induction chemotherapy in
patients with oral cancer. Oncol Lett. 15:8118–8124.
2018.PubMed/NCBI
|
|
57
|
Uitterhoeve AL, Koolen MG, van Os RM,
Koedooder K, van de Kar M, Pieters BR and Koning CC: Accelerated
high-dose radiotherapy alone or combined with either concomitant or
sequential chemotherapy; treatments of choice in patients with
non-small cell lung cancer. Radiat Oncol. 2:272007. View Article : Google Scholar : PubMed/NCBI
|