Introduction
Irritable bowel syndrome (IBS) is one of the most
prevalent functional gastrointestinal disorders (1). It is characterised by the presence of
recurrent or chronic abdominal pain or discomfort and bloating
(2); 3 and 35% patients with IBS
develop post-infectious IBS (PI-IBS) after acute gastrointestinal
infection (3). Although it is not
a lethal disorder, PI-IBS jeopardises the quality of life and
remains a substantial burden on the health care system. The
pathological mechanism of PI-IBS remains obscure, although growing
evidence has supported that abnormal motility, low-grade
inflammation, dysfunction of enteric nervous system (ENS), and
visceral hypersensitivity are associated with the development of
PI-IBS (4–6). Of these, dysfunction of the ENS is
considered an important factor causing inflammation and increased
intestinal permeability (7). ENS
is regarded as a separate and the largest component of the
peripheral nervous system (8). The
ENS plays a central role in gastrointestinal motility, the
maintenance of the epithelial barrier and local immune response
(9). Recent findings considered
the presence of structural and functional changes in the ENS as the
underlying pathophysiology in acute gastrointestinal-stress-related
PI-IBS symptoms (10,11). However, the mechanisms by which the
ENS contributes to the development of PI-IBS are still incompletely
understood.
The erythropoietin-producing hepatocellular (Eph)
receptor family is evolutionarily conserved and the largest of the
receptor tyrosine kinase families (12). Eph receptor family can be divided
into A or B subgroups, based on their specific affinities for
different subsets of ephrin ligands (13). Eph/ephrin signalling plays a
central role in contact-dependent communication between cells,
differentiation of the ENS, inflammatory response, epithelial
barrier function and the restoration of the injured intestinal
epithelium (14–16).
In the present study, we aimed to characterise the
role of Eph/ephrin signalling in lipopolysaccharide (LPS)-induced
intestinal injury in vivo and in vitro. Our results
showed that Eph/ephrin signalling play a bidirectional role in
LPS-induced intestinal injury in vivo and in vitro.
Eph/ephrin signalling might be a novel therapeutic target for
LPS-induced intestinal injury and even PI-IBS.
Materials and methods
Reagents
Escherichia coli LPS (E. coli 0127: B8), collagenase
XI, dispaseI, sorbitol, insulin and EGF were purchased from
Sigma-Aldrich (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany);
EphA2 monoclonal antibody (mAb) and mouse IgG was purchased from
Abcam (Cambridge, MA, USA); Foetal bovine serum (FBS), Dulbecco's
modified Eagle's medium (DMEM), L-glutamine, penicillin and
streptomycin were purchased from Gibco (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). ELISA kits were purchased from Thermo
Fisher Scientific, Inc.
LPS-induced intestinal injury in
vivo
Male BALB/c mice (20–24 g) were purchased from
Beijing Vital River Laboratory Animal Technology Co., Ltd.
(Beijing, China). All animal procedures were performed in
accordance with the National Institutes of Health Guidelines on the
Use and Care of Animals, with approval from the Institutional
Animal Experiment Committee of the Second People's Hospital of
Shenzhen. All efforts have been made to minimize animal suffering
and the number of animals used. They were housed in animal care
facilities under controlled conditions of temperature (23±1°C),
humidity (50±10%), with an alternating 12 h light/dark cycle and
free access to food and water. After 7 days of environmental
adaption, 30 mice were randomly divided into three groups: i)
PBS+IgG treatment group; ii) LPS+IgG treatment group; and iii)
LPS+EphA2 mAb treatment group. LPS (100 µg/kg) was administered
intravenously. 4 µg of either mouse IgG or EphA2 mAb were
administered intravenously 6 and 12 h after LPS treatment. The
control group was given 4 µg of mouse IgG 6 and 12 h after PBS
treatment. At the end of treatment, the mice were anesthetized with
a mixture of ketamine (100 mg/kg body weight) and xylazine (10
mg/kg body weight). Mice (n=10 per group) were perfused
transcardially with 0.9% saline (pH 7.4), and the colon tissues
were removed rapidly and carefully for ELISA and western
blotting.
Cell culture and drug treatment
Small intestines were aseptically removed from
l-day-old BALB/c mice. The intestines were incubated in collagenase
XI (750 µg/ml) and dispaseI (250 µg/ml) at 37°C for 30 min. Then
the tissues were purified through an osmotic gradient with 2%
sorbitol. The cell pellet was cultured in DMEM supplemented with
10% FBS, insulin (0.25 U/ml), EGF (20 ng/ml) and 1% penicillin (100
U/ml) /streptomycin (100 mg/ml) at 37°C in 5% CO2 and
95% atmosphere. The purification of primary cultured enteric
neuronal and glial cells was confirmed by immunofluorescence using
mouse monoclonal to PGP9.5 (ab8189; Abcam) and rabbit polyclonal to
GFAP (ab7260; Abcam). The primary cultured enteric neuronal and
glial cells on culture day 7 were used. The primary cultured
enteric neuronal and glial cells were incubated with LPS (50 ng/ml)
for 24 h. Cell viability was evaluated with the Cell Counting Kit-8
(CCK-8). The release of lactate dehydrogenase (LDH) was detected
using the assay kit (Nanjing Jiancheng Bioengineering Institute,
China).
ELISA
The levels of monocyte chemoattractant protein-1
(MCP-1), tumour necrosis factor α (TNF-α), interleukin-1β (IL-1β),
IL-6, intercellular adhesion molecule 1 (ICAM-1) and vascular cell
adhesion molecule-1 (VCAM-1) in the colon tissues and primary
cultured enteric neuronal and glial cells were measured by ELISA
kits following the manufacturer's instructions.
Western blot analysis
Immunoblotting was performed by technicians who were
blinded to the experimental groups. The protein extracts were
loaded into 8-12% Bis-Tris gels in a Bio-Rad slab gel apparatus
(MiniPROTEAN Tetra cell; Bio-Rad Laboratories, Inc., Hercules, CA,
USA) and electrophoretically transferred to a nitrocellulose
membrane. Blots were probed with the following antibodies: EphA2
(ab5386), Ephrin A1 (ab199697), Ephrin B3 (ab101699), EphB1
(ab129103), Cadherin (ab6528), β-catenin (ab16051), AKT (ab8805),
AKT1 (phospho S473) (ab18206), nuclear factor (NF)-κB p65
(ab16502), Src1 (ab5407), Src (phospho Y418) (ab4816), β-actin
(ab8227) and Lamin B (ab194109) were obtained from Abcam and were
used at a 1:1,000 dilution, followed by the appropriate secondary
antibodies. Bands were visualised using the enhanced
chemiluminescence method (Super Signal CL-HRP Substrate System;
Pierce; Thermo Fisher Scientific, Inc.), scanned with a
densitometer (Bio-Rad Laboratories, Inc.) and analysed
quantitatively with commercial equipment (Multi-Analyst Macintosh
Software for Image Analysis Systems; Bio-Rad Laboratories, Inc.).
At least three independent experiments were carried out.
Stable transfection
To establish ENS cells overexpressing Ephrin B3, ENS
cells were transfected with Ephrin B3 lentiviral activation
particles (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) in
complete medium with Polybrene (5 µg/ml) after reaching 60%
confluency and incubated overnight. Stable Ephrin B3 activated
clones were selected using puromycin dihydrochloride (5 µg/ml). The
protein expression of Ephrin B3 was examined by Western
blotting.
Statistical analysis
All experiments were performed a minimum of three
times. Statistical analysis was performed using SPSS 19.0 software
(IBM Corp., Armonk, IL, USA). Data are presented as the mean ±
standard deviation. Independent experiments were pooled when the
coefficient of variance could be assumed identical. One-way
analysis of variance was used to assess significant differences for
multiple groups, followed by the post hoc Bonferroni's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
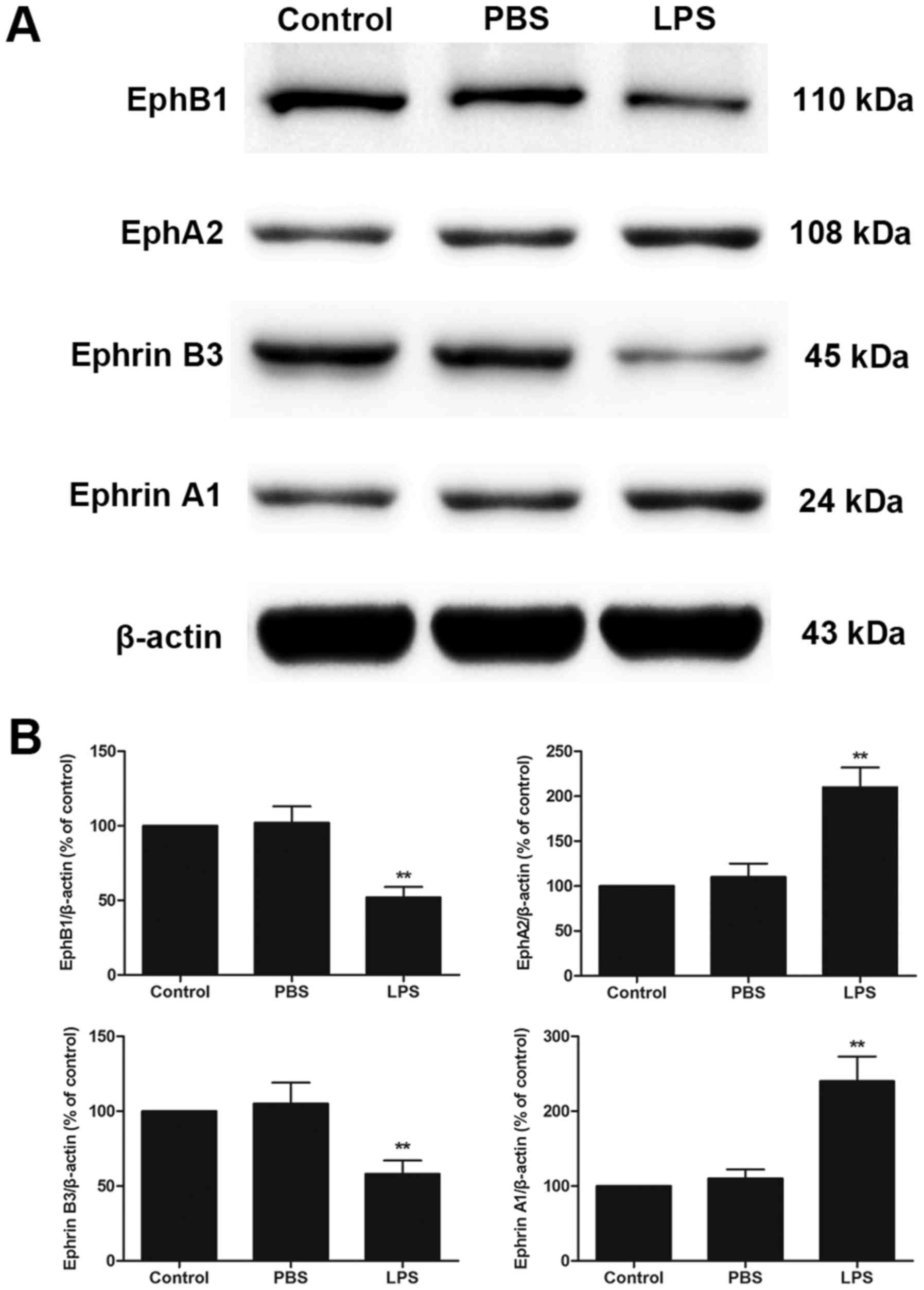
LPS treatment increased the expression
of EphA2 and Ephrin A1 but decreased the expression of EphB1 and
Ephrin B3 in colon
Western blot analysis showed that LPS treatment
increased EphA2 and Ephrin A1 protein expression but decreased
EphB1 and Ephrin B3 protein expression compared with PBS treatment
in colon (P<0.05; Fig. 1A and
B). These results implied that EphA2-Ephrin A1 signalling and
EphB1-Ephrin B3 signalling might play a central role in LPS-induced
intestinal injury.
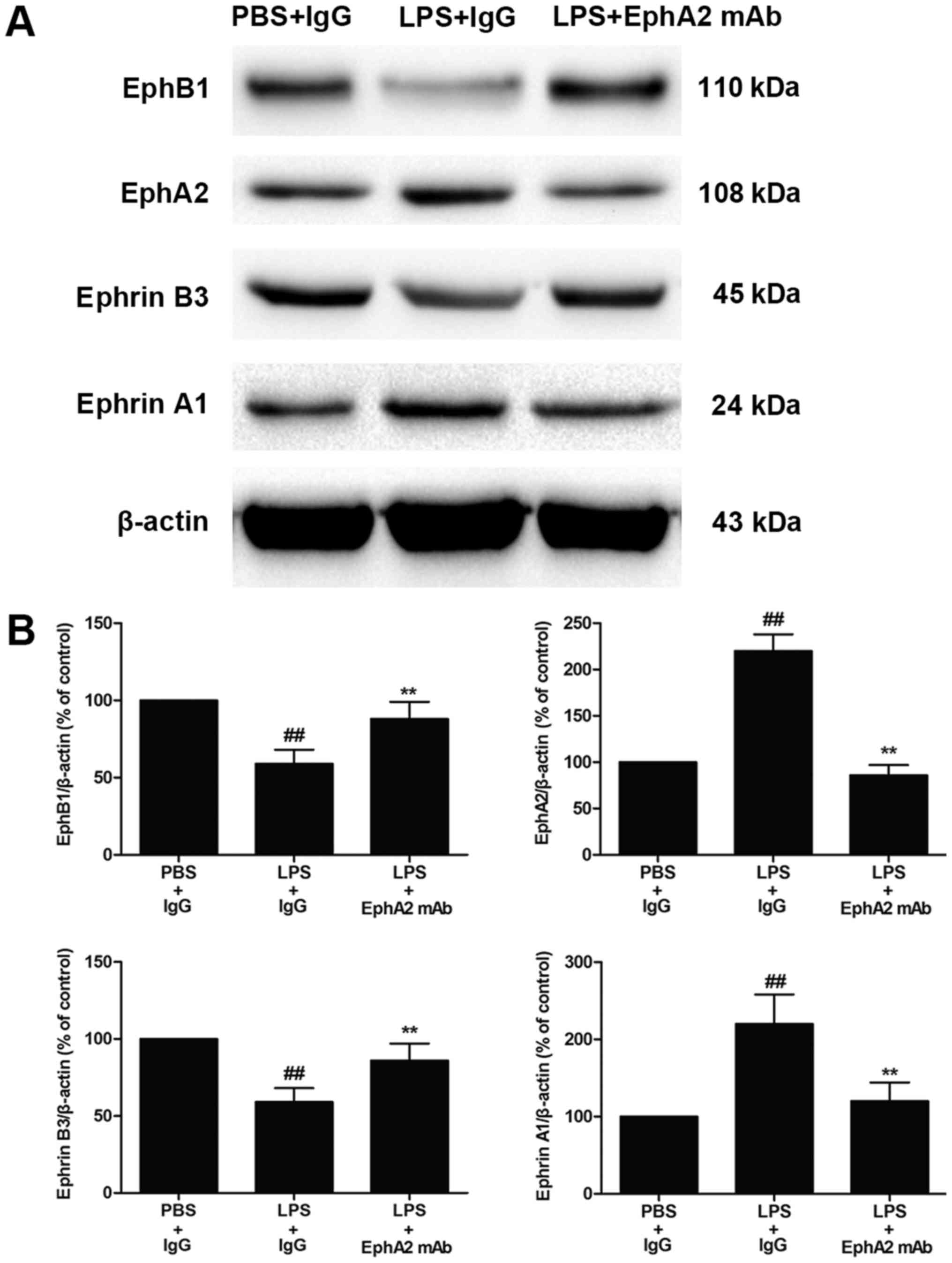
EphA2 antagonism enhanced EphB1/Ephrin
B3 signalling
After finding that LPS increased expression of EphA2
and Ephrin A1 in colon, we further investigated the effects of
EphA2 mAb on EphA2-ephrin A1 signalling and EphB1-Ephrin B3
signalling. We confirmed that the increased expression of EphA2 and
Ephrin A1 and the decreased expression of EphB1 and Ephrin B3
induced by LPS were partially inhibited by EphA2 mAb treatment
(P<0.05; Fig. 2A and B).
Therefore, the data implied that the interaction between
EphA2-Ephrin A1 and EphB1-Ephrin B3 might be positively correlated
with LPS-induced intestinal injury.
EphA2 antagonism attenuated
LPS-enhanced expression of inflammatory cytokines
Since inflammatory cytokines are major mediators of
LPS-induced intestinal injury, we detected the levels of
inflammatory cytokines (MCP-1, TNF-α, IL-1β, IL-6, ICAM-1 and
VACM-1) in colon. Compared with mice in PBS+IgG group, the levels
of inflammatory cytokines (MCP-1, TNF-α, IL-1β, IL-6, ICAM-1 and
VACM-1) were significantly increased in colon of mice in LPS+IgG
group (P<0.05; Fig. 3).
Compared with mice with IgG treatment after LPS treatment,
decreased expression of inflammatory cytokines including MCP-1
(254.31±32.96 vs. 677.25±71.11), TNF-α (21.85±4.92 vs. 45.08±6.12),
IL-1β (2.43±0.36 vs. 5.65±0.91), IL-6 (19.19±3.05 vs. 44.44±8.18),
ICAM-1 (34.11±7.25 vs. 111.12±21.31), VACM-1 (18.41±4.55 vs.
34.19±5.09) were observed in the colon of mice with EphA2 mAb
treatment (P<0.05; Fig. 3).
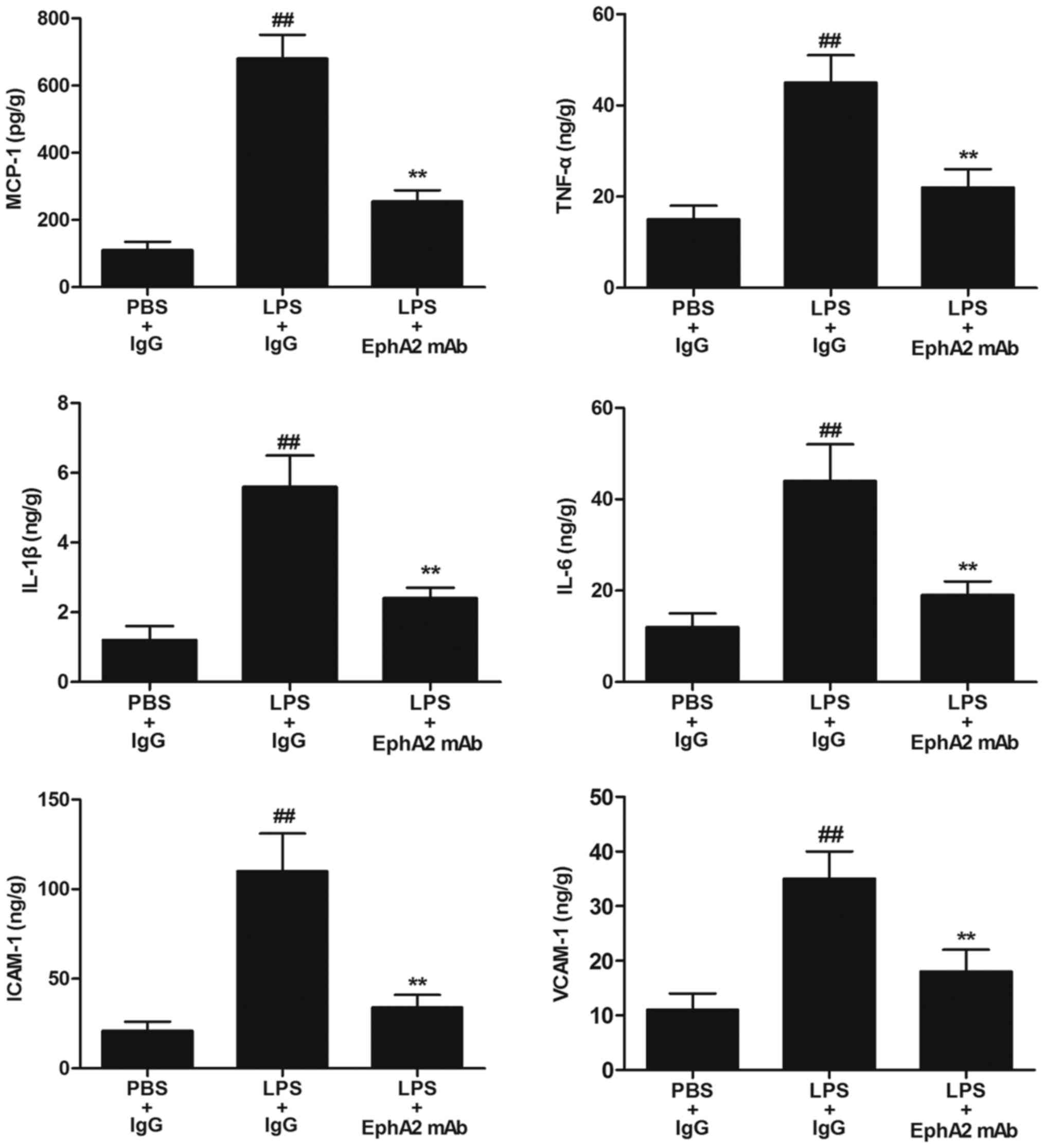 | Figure 3.Effects of EphA2 antagonism on the
levels of inflammatory cytokines in vivo. The levels of the
inflammatory cytokines (MCP-1, TNF-α, IL-1β, IL-6, ICAM-1 and
VCAM-1) in colon tissue lysates were measured by ELISA. The results
are expressed as the mean ± standard deviation (n=6/group).
##P<0.01 vs. PBS+IgG (control); **P<0.01 vs.
LPS+IgG treatment. Eph, erythropoietin-producing hepatocellular;
LPS, lipopolysaccharide; IgG, immunoglobulin G; mAb, monoclonal
antibody; MCP-1, monocyte chemoattractant protein-1; TNF-α, tumour
necrosis factor α; IL, interleukin; ICAM-1, intercellular adhesion
molecule 1; VCAM-1, vascular cell adhesion molecule-1. |
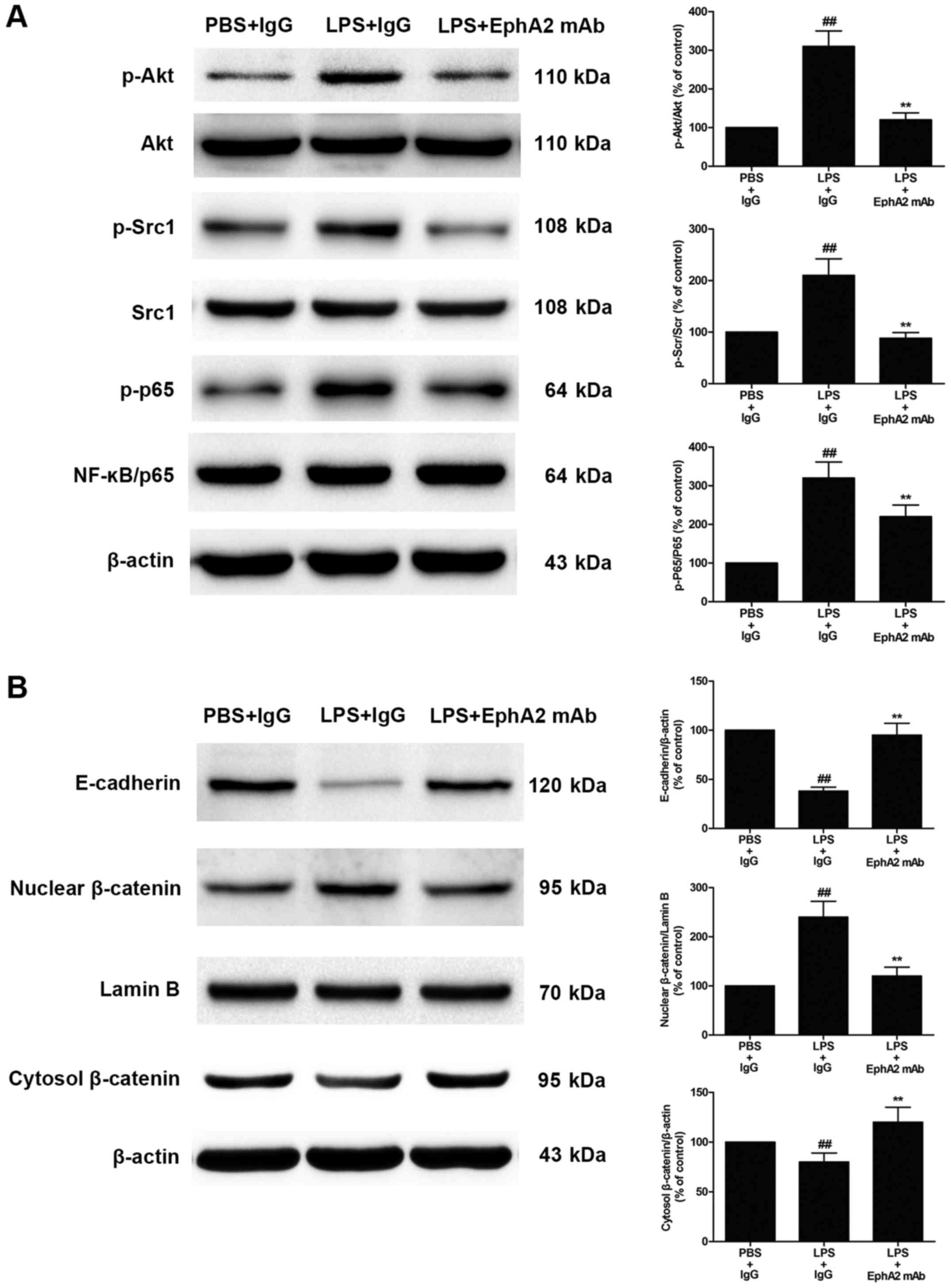
EphA2 antagonism attenuated
LPS-induced activation of Akt-NF-κB signalling, Src-NF-κB
signalling and Wnt/β-catenin signalling pathway
Since the protective effect of EphA2 antagonism in
LPS-induced intestinal injury, we wondered whether EphA2 antagonism
would affect Akt-NF-κB signalling and Src-NF-κB signalling
pathways. The expression of Akt-NF-κB signalling and Src-NF-κB
signalling were measured. As shown in Fig. 4A, compared with the PBS+IgG group,
the LPS+IgG group showed a significant increase in Akt and Src
phosphorylation in colon tissue (P<0.05). After EphA2 mAb
treatment, the phosphorylation of Akt and Src induced by LPS was
significantly reduced (P<0.05; Fig.
4A). Moreover, the phosphorylation of NF-κB p65 was increased
significantly after LPS treatment (P<0.05). However, a
significant decrease was observed in the phosphorylation of NF-κB
p65 after EphA2 mAb treatment, compared with the IgG treatment
after LPS treatment (P<0.05; Fig.
4A). This finding indicates that EphA2 signalling may be
involved in LPS-induced activation of Akt-NF-κB signalling and
Src-NF-κB signalling. Activation of the Wnt/β-catenin pathway plays
a pivotal role in LPS-induced intestinal injury. To determine
whether EphA2 antagonism could affect the Wnt/β-catenin pathway,
the expression level of Wnt/β-catenin signalling proteins were
determined by immunoblotting. As shown in Fig. 4B, compared with the PBS+IgG group,
LPS could significantly increased nuclear translocation of
β-catenin (P<0.05; Fig. 4B).
However, EphA2 mAb treatment could significantly decreased nuclear
translocation of β-catenin, compared with IgG treatment after LPS
treatment (P<0.05; Fig. 4B),
suggesting that EphA2 signalling is responsible for LPS-mediated
activation of the Wnt/β-catenin pathway. Furthermore, LPS treatment
could significantly reduce the expression of E-cadherin (P<0.05;
Fig. 4B), whereas treatment with
EphA2 mAb significantly increased E-cadherin protein expression in
colon (P<0.05; Fig. 4B). These
results demonstrate that EphA2 signaling may regulate the
expression of E-cadherin and adherens junction and epithelial
hyperpermeability.
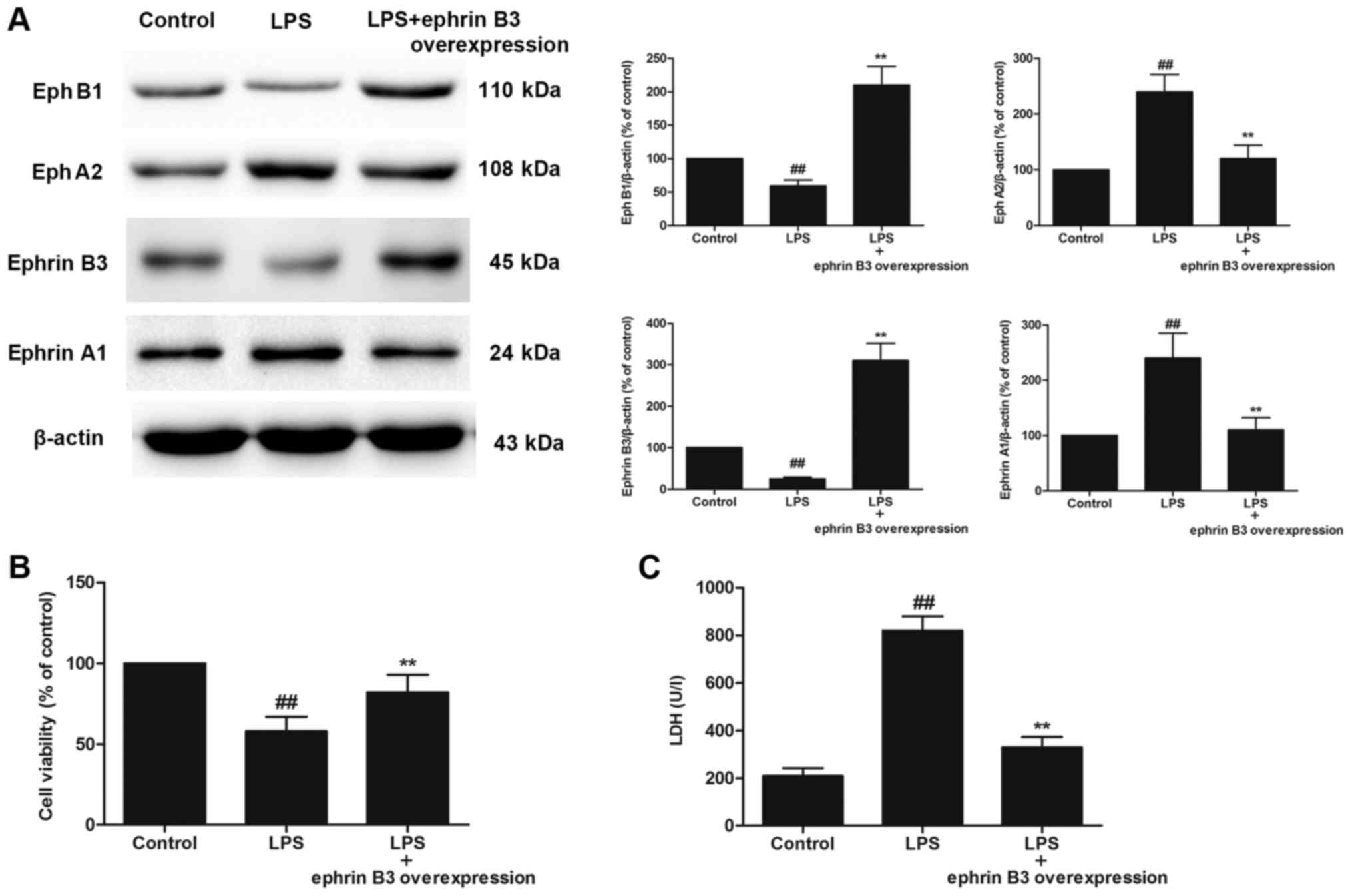
EphB1/Ephrin B3 signalling antagonised
the EphA2/Ephrin A1-dependent pathway
The primary cultured enteric neuronal and glial
cells were confirmed by immunofluorescence using mouse monoclonal
to PGP9.5 and rabbit polyclonal to GFAP. The purification of
primary cultured enteric neuronal and glial cells was more than 90%
(data not shown). We further investigated the effects of Ephrin B3
overexpression on LPS-induced injury in primary cultured enteric
neuronal and glial cells. A significant increase in the expression
of EphA2 and Ephrin A1 and a significant decrease in EphB1 and
Ephrin B3 were observed in LPS-treated primary cultured enteric
neuronal and glial cells, compared with the control cells
(P<0.05; Fig. 5A). However,
Ephrin B3 overexpression reversed LPS-induced up-regulation of
EphA2 and Ephrin A1, and enhanced expression of EphB1 compared with
the control group (P<0.05; Fig.
5A). Importantly, Ephrin B3 overexpression significantly
increased the cell viability and significantly decreased LDH
leakage (P<0.05; Fig. 5B).
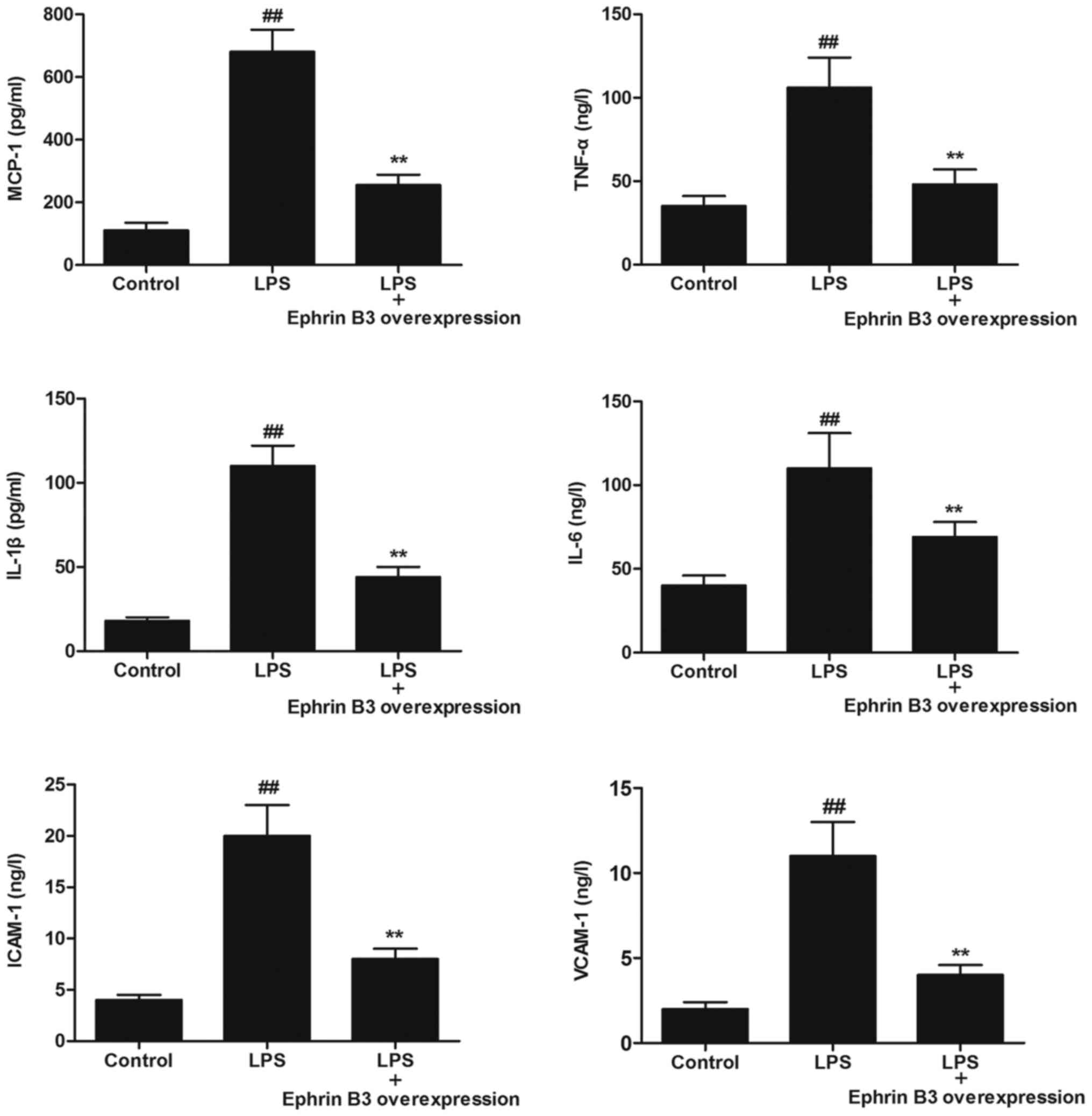
Ephrin B3 overexpression reversed
LPS-enhanced expression of inflammatory cytokines in primary
cultured enteric neuronal and glial cells
To examine whether Ephrin B3 overexpression
negatively regulated LPS-induced inflammatory responses, we
detected the levels of inflammatory cytokines (MCP-1, TNF-α, IL-1β,
IL-6, ICAM-1 and VACM-1) in primary cultured enteric neuronal and
glial cells. We found that the levels of inflammatory cytokines
(MCP-1, TNF-α, IL-1β, IL-6, ICAM-1 and VACM-1) were significantly
increased after LPS treatment, compared with the control
(P<0.05; Fig. 6). However,
Ephrin B3 overexpression significantly reversed LPS-induced
up-regulation of inflammatory cytokines (P<0.05; Fig. 6).
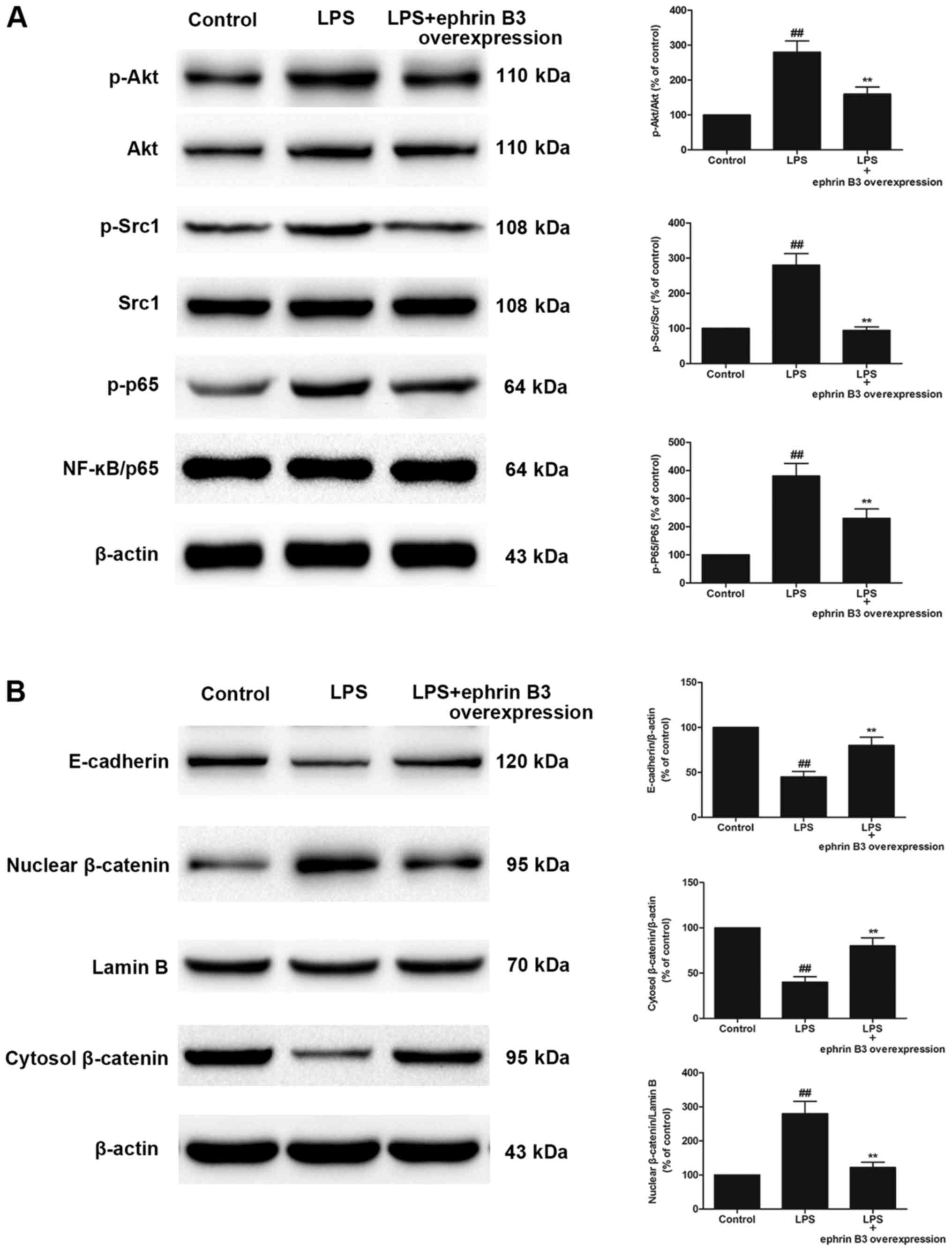
Ephrin B3 overexpression attenuated
LPS-induced activation of Akt-NF-κB signalling, Src-NF-κB
signalling and Wnt/β-catenin signalling pathway in primary cultured
enteric neuronal and glial cells
Because EphA2 signalling was shown to be crucial in
LPS-induced activation of Akt-NF-κB signalling, Src-NF-κB
signalling and Wnt/β-catenin pathway, we examined the influence of
Ephrin B3 overexpression in LPS-induced activation of Akt-NF-κB
signalling, Src-NF-κB signalling and Wnt/β-catenin pathway, thus
exploring the potential link between bidirectional signalling
mediated by Eph-Ephrins. Ephrin B3 overexpression significantly
reversed the up-regulated phosphorylation of Akt, Src and NF-κB p65
in LPS-treated primary cultured enteric neuronal and glial cells
(P<0.05; Fig. 7A). In addition,
Ephrin B3 overexpression significantly inhibited the nuclear
translocation of β-catenin and increased expression of E-cadherin
in LPS-treated primary cultured enteric neuronal and glial cells
(P<0.05; Fig. 7B).
Discussion
In recent years, the role of ENS in PI-IBS has
gained increasing attention; however, there are still only a few
studies addressing how the ENS contributes to PI-IBS. The data from
our present work indicated that ENS responded to LPS-induced injury
by activating proinflammatory mediators, and integrating the
Eph-ephrin signalling-dependent downstream pathways (including the
Akt-NF-κB, Src-NF-κB and Wnt/β-catenin pathways), which facilitated
the development of PI-IBS. Importantly, we hypothesised that Eph
receptors and ephrins activated bidirectional signalling networks
which played a crucial role in PI-IBS.
Our studies demonstrated that EphA2-Ephrin A1
signalling was activated and EphB1-Ephrin B3 signalling was
inactivated in LPS-induced injury in vivo and in
vitro. Both the expression of the EphA2 receptor and Ephrin A1
were markedly increased in LPS-treated colon and ENS, while the
expression levels of EphB1 receptor and Ephrin B3 were decreased.
In addition, blocking of EphA2 receptor by EphA2 mAb partially
ameliorated LPS-triggered injury and inflammatory responses in a
mouse model. Moreover, Ephrin B3 overexpression reversed
LPS-induced damage in primary cultured enteric neuronal and glial
cells. In our experiments, we demonstrated that the Eph/ephrins
singnaling played an opposing role and exerted different effects in
LPS-induced intestial injury. We found that EphA2-Ephrin A1
mediated ‘forward’ signalling might contribute to LPS-induced
injury, while Ephrin B3 dependent ‘reverse’ signalling may have a
role in generating the negative regulation function after LPS
treatment.
Signal transduction activated by ephrin binding to
Eph receptors is associated with their interaction with specific
intracellular pathways (17–19).
Herein, our results suggested that increased Ephrin A1 might induce
ligand-dependent EphA2 signalling, which further induced
phosphorylation of Akt and Src. The activation of Akt and Src would
trigger nuclear translocation of NF-κB, which would lead to
enhanced proinflammatory cytokine production. The role of Akt and
Src pathways in modulating NF-κB activation has been demonstrated
in numerous cell populations (20,21).
Activation of Akt is required for the efficient localisation of p65
to the promoter regions of a specific subset of the NF-κB-targeted
genes (22). In addition, it has
been demonstrated that Src tyrosine kinases mediate the activation
of NF-κB in LPS-induced injury, and that selective Src tyrosine
kinase inhibitors could prevent this damage (23). In the present study, antagonising
the EphA2-Ephrin A1 pathway by EphA2 mAb treatment partially
attenuated the phosphorylation of Akt-Src and NF-κB, which
suggested that LPS-induced activation of the Akt-NF-κB and
Src-NF-κB pathways were mediated by EphA2-Ephrin A1 signalling.
Furthermore, our findings also indicated the
involvement of EphA2 signalling in LPS-induced NF-κB activation via
Wnt/β-catenin as an upstream pathway. Enhanced EphA2 would
facilitate the nuclear translocation of β-catenin protein, which is
known to be a marker for hyperactivation of Wnt signalling
(24). Many studies have
demonstrated that constitutive activation of Wnt/β-catenin
signalling promoted the activation of NF-Κb (25,26).
Conversely, downregulation of the nuclear translocation of
β-catenin and abrogation of Wnt signalling was exactly associated
with inactivation of NF-κB (27).
Our study demonstrated that EphA2 mAb partially inhibited Wnt
signalling by inhibiting the nuclear translocation of β-catenin in
LPS-induced injury. This suggests that EphA2 might promote NF-κB
activation via the Wnt/β-catenin pathway.
Interestingly, our studies also demonstrated that
EphB1-Ephrin B3 signalling might act to antagonise the
EphA2-dependent pathway after LPS treatment. We found Ephrin B3
overexpression could reverse LPS-triggered injury, increased
concentrations of cytokines, and activation of Akt-Src and NF-κB,
which implied that enhanced Ephrin B3-dependent ‘reverse’
signalling served as a potentially negative regulator to counteract
the EphA2-Ephrin A1 pathway. Previous studies have provided
evidence that binding of EphB1 to Ephrin B3 led to a reduction and
inactivation of Src in striatal neurons (28). In contrast, in cortical
interneurons binding of EphB1 to Ephrin B3 would enhance the
phosphorylation of Src (29).
Consistent with our data, it has been demonstrated that the
activation of EphrinB-dependent ‘reverse’ signalling could
downregulate β-catenin level in the cytoplasm by recruiting Axin
protein, but in the meantime Wnt signalling could also suppress the
EphB-ephrinB pathway by inhibiting the transcription of ephrinB
ligands (30). Therefore, we
hypothesised that the maladjustment of negative feedback loops
between EphrinB-dependent ‘reverse’ signalling and the
Wnt/β-catenin pathway might act as a crucial factor which
influenced the excessive activation of the EphA2-Ephrin A1 pathway
and led to LPS-induced persistent inflammation and injury.
Furthermore, we speculated that there might be more interacting
links between the Eph-ephrin ‘forward’ and ‘reverse’ pathways, and
these bidirectional signalling networks often meditated opposing
events. This may explain why LPS stimulation in a cellular context
can trigger dramatically different outcomes of Eph/ephrin
response.
Previous studies have proved that Eph/ephrin
signaling directly activated Akt, NF-κB and Wnt/β-catenin
signalling pathways (18).
Crosstalk between Wnt/β-Catenin and NF-κB Signaling has been
reported (25). In this study, we
found that inhibition of EphA2 or Ephrin B3 overexpression could
partially alleviate LPS-triggered activation of Akt-NF-κB,
Src-NF-κB and Wnt/β-catenin signalling pathways. However, the
precise mechanism underlying Eph/ephrin signaling regulates Akt,
NF-κB and Wnt/β-catenin signalling pathways and the crosstalk
needed to be intensively investigated in future studies.
In conclusion, we demonstrated that EphA2 signalling
contributes to inflammation in LPS-induced injury and that it may
regulate several signalling pathways, including the Akt-NF-κB,
src-NF-κB and Wnt/β-catenin pathways. Nevertheless, EphA2 mAb
treatment partially attenuated LPS-induced activation of
EphA2-Ephrin A1 signalling. In addition, with regard to
bidirectional signalling of the Eph-ephrins complex, we suspect
that binding of EphB1 to Ephrin B3 mediated the counter-directed
effect, which reduced activation of the EphA2-Ephrin A1 pathway. In
the case of Ephrin B3 overexpression, two mechanisms may counteract
EphA2-dependent signalling. First, Ephrin B3 overexpression
decreased the expression of EphA2 and Ephrin A1. Second, the
enhanced Ephrin B3-dependent'reverse’ signalling led to an
inactivation of the Wnt/β-catenin pathway. Our results suggest that
EphA2 antagonism or reinforcing Ephrin B3 binding to EphB1 might be
a potentially therapeutic avenue for LPS-induced intestinal injury,
and even PI-IBS. Further work regarding Eph/ephrin signalling in
the PI-IBS is required to establish a therapeutic strategy with
clinical potential.
Acknowledgements
Not applicable.
Funding
The present study was supported by the International
Scientific and Technological Cooperation Projects of Shenzhen
Collaborative Innovation Technology Plan (grant no.
GJHZ20150316141713255).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YX, KL, HW and LZ contributed to the conception and
design of the study. YX, KL, HW, LJ, SY and LZ performed the
experiments to acquire the data. YX, KL, LJ, SY and LZ performed
data analyses and interpreted the data.
Ethics approval and consent to
participate
The present study was approved by The Institutional
Animal Experiment Committee of the Second People's Hospital of
Shenzhen (Guangdong, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Oświęcimska J, Szymlak A, Roczniak W,
Girczys-Połedniok K and Kwiecień J: New insights into the
pathogenesis and treatment of irritable bowel syndrome. Adv Med
Sci. 62:17–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mearin F, Ciriza C, Mínguez M, Rey E,
Mascort JJ, Peña E, Cañones P and Júdez J: en nombre de la SEPD; la
semFYC, et al: Clinical practice guidelines: Irritable bowel
syndrome with constipation and functional constipation in adults:
Concept, diagnosis, and healthcare continuity. (Part 1 of 2).
Atencion Primaria. 49:42–55. 2017.(In Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sundin J, Rangel I, Repsilber D and
Brummer RJ: cytokine response after stimulation with key commensal
bacteria differ in Post-Infectious Irritable Bowel Syndrome
(PI-IBS) patients compared to healthy controls. PLoS One.
10:e01348362015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mondelaers SU, Theofanous SA, Florens MV,
Perna E, Aguilera-Lizarraga J, Boeckxstaens GE and Wouters MM:
Effect of genetic background and postinfectious stress on visceral
sensitivity in Citrobacter rodentium-infected mice.
Neurogastroenterol Motil. 28:647–658. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lan C, Sun XN, Zhou XC, Yang B, Huang BL,
Deng TZ, He ZT and Han XY: Preinduced intestinal HSP70 improves
visceral hypersensitivity and abnormal intestinal motility in
PI-IBS mouse model. Asian Pac J Trop Med. 9:302–305. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Polster AV, Friberg P, Neve BL, Törnblom H
and Simren M: Autonomic nervous system function in patients with
Irritable Bowel Syndrome (IBS): Relevance for symptoms?
Gastroenterology. 152 Suppl 1:S7242017. View Article : Google Scholar
|
|
7
|
Menzies V, Jallo N, Kinser P, Robins JL,
An K, Driscoll C, Starkweather A, Bajaj JS and Lyon DE: Shared
symptoms and putative biological mechanisms in chronic liver
disease: Implications for biobehavioral research. Biol Res Nurs.
17:222–229. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hyland NP and Cryan JF: Microbe-host
interactions: Influence of the gut microbiota on the enteric
nervous system. Dev Biol. 417:182–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rao M, Rastelli D, Dong L, Chiu S, Setlik
W, Gershon MD and Corfas G: Enteric glia regulate gastrointestinal
motility but are not required for maintenance of the epithelium in
mice. Gastroenterology. 153:1068–1081.e7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cashman MD, Martin DK, Dhillon DK and Puli
SR: Irritable bowel syndrome: A clinical review. Curr Rheumatol
Rev. 12:13–26. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li S, Fei G, Fang X, Yang X, Sun X, Qian
J, Wood JD and Ke M: Changes in enteric neurons of small intestine
in a rat model of irritable bowel syndrome with diarrhea. J
Neurogastroenterol Motil. 22:310–320. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park I and Lee HS: EphB/ephrinB signaling
in cell adhesion and migration. Mol Cells. 38:14–19. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barquilla A and Pasquale EB: Eph receptors
and ephrins: Therapeutic opportunities. Annu Rev Pharmacol Toxicol.
55:465–487. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen X, Cheng ZR, Zhang SJ, Werling D and
Wathes DC: Combining genome wide association studies and
differential gene expression data analyses identifies candidate
genes affecting mastitis caused by two different pathogens in the
dairy cow. Open J Anim Sci. 5:358–393. 2015. View Article : Google Scholar
|
|
15
|
Kang M, Jeong W, Bae H, Lim W, Bazer FW
and Song G: Bifunctional role of ephrin A1-Eph system in
stimulating cell proliferation and protecting cells from cell death
through the attenuation of ER stress and inflammatory responses in
bovine mammary epithelial cells. J Cell Physiol. 233:2560–2571.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han J, Xu Y, Yang D, Yu N, Bai Z and Bian
L: Effect of polysaccharides from acanthopanax senticosus on
intestinal mucosal barrier of escherichia coli lipopolysaccharide
challenged mice. Asian-Australas J Anim Sci. 29:134–141. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dun MD, Chalkley RJ, Faulkner S, Keene S,
Avery-Kiejda KA, Scott RJ, Falkenby LG, Cairns MJ, Larsen MR,
Bradshaw RA and Hondermarck H: PProteotranscriptomic profiling of
231-BR breast cancer cells: Identification of potential biomarkers
and therapeutic targets for brain metastasis. Mol Cell Proteomics.
14:2316–2330. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hong JY, Shin MH, Douglas IS, Chung KS,
Kim EY, Jung JY, Kang YA, Kim SK, Chang J, Kim YS and Park MS:
Inhibition of EphA2/EphrinA1 signal attenuates
lipopolysaccharide-induced lung injury. Clin Sci (Lond).
130:1993–2003. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dunne PD, Dasgupta S, Blayney JK, McArt
DG, Redmond KL, Weir JA, Bradley CA, Sasazuki T, Shirasawa S, Wang
T, et al: EphA2 expression is a key driver of migration and
invasion and a poor prognostic marker in colorectal cancer. Clin
Cancer Res. 22:230–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Whitman E and Barber A: NKG2D receptor
activation of NF-κB enhances inflammatory cytokine production in
murine effector CD8(+) T cells. Mol Immunol. 63:268–278. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng CY, Huang WR, Chi PI, Chiu HC and
Liu HJ: Cell entry of bovine ephemeral fever virus requires
activation of Src-JNK-AP1 and PI3K-Akt-NF-κB pathways as well as
Cox-2-mediated PGE2/EP receptor signalling to enhance
clathrin-mediated virus endocytosis. Cell Microbiol. 17:967–987.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Daniel AR, Gaviglio AL, Knutson TP,
Ostrander JH, D'Assoro AB, Ravindranathan P, Peng Y, Raj GV, Yee D
and Lange CA: Progesterone receptor-B enhances estrogen
responsiveness of breast cancer cells via scaffolding PELP1- and
estrogen receptor-containing transcription complexes. Oncogene.
34:506–515. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu Y, Zhang Y, Wang L, Diao Z and Liu W:
The role of autophagy in kidney inflammatory injury via the NF-κB
route induced by LPS. Int J Med Sci. 12:655–667. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang J, He Y, Mcleod HL, Xie Y, Xiao D,
Hu H, Chen P, Shen L, Zeng S, Yin X, et al: miR-302b inhibits
tumorigenesis by targeting EphA2 via Wnt/β-catenin/EMT signaling
cascade in gastric cancer. BMC Cancer. 17:8862017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma B and Hottiger MO: Crosstalk between
Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation.
Front Immunol. 7:3782016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma B, Fey M and Hottiger MO: WNT/β-catenin
signaling inhibits CBP-mediated RelA acetylation and expression of
proinflammatory NF-κB target genes. J Cell Sci. 128:2430–2436.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Voutilainen M, Lindfors PH, Trela E,
Lönnblad D, Shirokova V, Elo T, Rysti E, Schmidt-Ullrich R,
Schneider P and Mikkola ML: Ectodysplasin/NF-κB promotes mammary
cell Fate via Wnt/β-catenin pathway. PLoS Genet. 11:e10056762015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang H, Li R, Yuan J, Zhou X, Liu X, Ou
S, Xu T and Chen Y: Up-regulated ephrinB3/EphB3 expression in
intractable temporal lobe epilepsy patients and pilocarpine induced
experimental epilepsy rat model. Brain Res. 1639:1–12. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dong LD, Gao F, Wang XH, Miao Y, Wang SY,
Wu Y, Li F, Wu J, Cheng XL, Sun XH, et al: GluA2 trafficking is
involved in apoptosis of retinal ganglion cells induced by
activation of EphB/EphrinB reverse signaling in a rat chronic
ocular hypertension model. J Neurosci. 35:5409–5421. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Eusemann TN, Willmroth F, Fiebich B, Biber
K and van Calker D: Adenosine receptors differentially regulate the
expression of regulators of G-protein signalling (RGS) 2, 3 and 4
in astrocyte-like cells. PLoS One. 10:e01349342015. View Article : Google Scholar : PubMed/NCBI
|