Introduction
Joint immobilization (IM) is a common treatment type
in orthopedics, while clinical studies have revealed that joint IM
and stress may lead to morphological and biochemical alterations in
the articular cartilage. Animal studies have revealed that joint IM
induces alterations in the articular cartilage (1–4). A
marked decrease in cartilage thickness was observed in animals with
chronic joint IM (5–7). Alterations in proteoglycan synthesis,
fibrofatty proliferation and chondromalacia were additionally
reported in animals with joint immobilization (1–13).
Similar morphological alterations in the articular cartilage were
observed in patients with joint fixation treatment, which may in
turn induce osteoarthritis (OA). Thus, investigation into the
underlying mechanism of articular cartilage degeneration induced by
joint immobilization remains necessary to develop strategies for
the prevention and treatment of complications of joint
immobilization, including OA (8–11).
OA is characterized by articular cartilage
degeneration, osteophyte formation and a narrowed joint space
(12). Joint cartilage is involved
in the development of OA (13). A
number of studies have suggested that increased pro-inflammatory
cytokines are critical to the development of OA (14). Infiltration and activation of
immune cells (including macrophages, mast cells and T cells) in the
joints induce synovitis and damage the joint cartilage (14–16).
These results indicate that studying the role of immune factors
associated with synovitis assists in better understanding the
clinical features of joint IM induced by OA.
Complement component C5a (C5a) is an important
pro-inflammatory peptide, and is involved in complement activation,
membrane attack complex formation, chemotaxis of immune cells and
allergic responses (17,18). C5a is generated by cleavage of
complement component C5 by protease C5-convertase (17,18).
A number of studies have assessed the role of C5a and its
associated proteins in inflammatory joint diseases. The interaction
between C5a and C5aR induces the release of TNF-α in activated
synoviocytes (19,20), suggesting that the C5a/C5aR system
may serve an important role in the joint inflammation process.
Although the complement system is implicated in the inflammatory
processes of arthritic diseases, direct interaction between C5a and
joint IM induced by OA has not been demonstrated.
In the present study, it was hypothesized that C5a
may be involved in the pathogenesis of joint IM-induced OA, and
inhibition of C5a may act as a novel therapeutic target for the
prevention of OA. Therefore, to verify this hypothesis, the role of
C5a in the development of OA was studied in a chronic knee fixation
animal model.
Materials and methods
Animal model and experiments
design
A total of 72 adult male Sprague-Dawley rats,
(380–400 g; 14 weeks) were purchased from the Guangdong Medical
Laboratory Animal Center (Foshan, China) were used in the present
study. The rats were randomly assigned into three groups: Control
group (n=24), immobilization group (IM group, n=24), and
immobilization+anti-C5a antibody group (IM+anti-C5a group, n=24).
The rats in the control group did not undergo any treatment, while
rats in the IM and IM+anti-C5a groups underwent the following
procedure: A degreasing cotton patch was placed in the popliteal
fossa of unilateral (50% right and 50% left in each group) knee
joints to immobilize the joint at 180° and fixed with a plaster
bandage (Anji Wande Medical Products Co., Ltd., Anji, China) for 6
weeks. Saline (1 ml) or anti-C5a antibody (0.1 µg/ml; Wuhan Boster
Biological Technology, Ltd., Wuhan, China; cat. no. PB0184) were
given every week for 6 weeks. A total of four rats/group for 6
weeks were sacrificed at the end of each week for the subsequent
assays. All procedures were approved by the Guizhou Medical
University (Guiyang, China) Animal Care and Use Committees and were
in accordance with the National Institutes of Health Guide (1996)
for the care and use of laboratory animals (21).
Tissue preparation
Cartilage dissection
The femur was truncated at 5 cm above the femoral
condyle, and tibia was truncated at 3 mm below the tibial plateau.
Soft tissue such as muscle, fat, periosteum surrounding knee joint
were removed carefully. Patellar ligament, medial and lateral
collateral ligament and anterior cruciate ligament was cut off, and
the aponeurotic laminae on both sides were cut open. The patella is
flipped down. The knee is opened and the articular cartilage is cut
with a 15# scalpel blade. The cartilage on each joint surface is
cut off. Do not accidentally bring the subchondral bone and
periosteum.
Synovectomy
Cut the skin longitudinally along the median knee
joint until an area of about 3×3 cm centered on the knee joint is
exposed. Lift the patella with a tweezers. femur was truncated at
0.13~0.14 cm above the upper edge of patella. Split down to the
tibia along both sides of the patella. After the knee joint cavity
was opened, a layer of smooth and shiny yellowish synovial tissue
adhered to the lower edge of the patella can be seen.
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay and hematoxylin and eosin
(H&E) staining
The specimens were fixed with 4% paraformaldehyde in
0.1 M phosphate buffer, at pH 7.4 overnight at 4°C. The tissues
were dehydrated through a series graded ethanol baths (100, 95, 90,
80 and 70%), and the specimens were embedded in paraffin. The
embedded tissue was cut into 4 µm sections. The center of the
scanned region was set for histological evaluation.
TUNEL staining of fractured double-stranded DNA in
apoptotic cells was performed. Paraffin sections (4 µm) of synovial
and cartilage tissues were used for the evaluation of apoptosis by
TUNEL staining. The In Situ Cell Death Detection kit-POD
(Roche Diagnostics, Basel, Switzerland; cat. no. 11684817910) was
used to determine the number if apoptotic cells, according to the
manufacturer's protocol. Counting of TUNEL-positive cells was
conducted on ×200-magnified sections for each group. A H&E
Staining kit (Shanghai Yeasen Biotechnology Co., Ltd., Shanghai,
China, cat. no. 60524ES60) was used for staining the cardiac
muscle, according to the manufacturer's protocols. Briefly, slides
were stained for 5 min with hematoxylin, washed with diluted water
three to five times at room temperature and subsequently stained
with eosin solution for 20 sec at room temperature.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the synovial fluid and
cartilage tissue by using TRIzol reagent (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). cDNA was synthesized using 1 µg total RNA
via an RT kit (DBI Bioscience, Newark, DE, USA). The RNA solution
was incubated at 65°C for 5 min, and the temperature protocol for
the reverse transcription reaction was 37°C for 15 min, followed by
98°C for 5 min. The primers used in qPCR were as follows: C5a
sense, 5′-CTCACCTTCCTCACCAATGC-3′ and antisense,
5′-GCTTGTTCTTCCACTTTCTGAT-3′; GAPDH sense,
5′-CCTCGTCTCATAGACAAGATGGT-3′ and antisense,
5′-GGGTAGAGTCATACTGGAACATG-3′. SYBR-Green qPCR master Mix (DBI
Bioscience) was used for qPCR amplification. Cycling conditions
included denaturation at 95°C for 2 min followed by 40 cycles of
annealing at 94°C for 20 sec and extension at 58°C for 20 sec. On
the basis of exponential amplification of the target gene relative
to the housekeeping gene as an internal reference, the quantity of
amplified molecules at the quantification cycle was given by
2−ΔΔCq (22).
ELISA analysis
Expression levels of IL-1β, C5a, TNF-α and IL-17A in
the synovial fluid and cartilage tissue were measured using an
antigen-based sandwich ELISA. The Rat IL-1β ELISA kit (cat. no.
E-EL-R0012c), Rat C5a ELISA kit (cat. no. E-EL-R0257c), Rat TNF-α
ELISA kit (cat. no. E-EL-R0019c) and Rat IL-17A ELISA Kit (cat. no.
E-EL-R0566c; all Elabscience, Houston, TX, USA) were used. The
absorbance was measured at 450 nm using a microplate
spectrophotometer (Multiskan GO; Thermo Fisher Scientific,
Inc.).
Western blot analysis
A total of 40 mg synovial fluid and cartilage tissue
was placed in 120 µl radioimmunoprecipitation assay (RIPA) lysis
buffer (Beyotime Institute of Biotechnology, Shanghai, China) with
phosphatase and protease inhibitors (BIOSS, Beijing, China) for 2 h
at room temperature. Protein concentration was determined with
bicinchoninic acid protein assay. Total protein (20 µl) was
separated on 12% Tris-glycine polyacrylamide gels. Samples were
subsequently transferred onto polyvinylidene fluoride membranes
(Pall Life Sciences, Port Washington, NY, USA). Membranes were
incubated in blocking buffer (cat. no. P0023B; Beyotime Institute
of Biotechnology) for 8 h at 4°C, and subsequently incubated with
rabbit anti-C5a (cat. no. ab11876; 1:2,000), rabbit anti-IL-1β
(cat. no. ab200478; 1:2,000), rabbit anti-TNF-α (cat. no. ab9755;
1:2,000; all Abcam, Cambridge, UK;), rabbit anti-IL-17A (cat. no.
SAB3701458-100UG; 1:2,000; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), and rabbit anti-GAPDH (cat. no. KC-5G5; 1:5,000; Kangchen
BioTech Co., Ltd., Shanghai, China) for 8 h at 4°C. Horseradish
peroxidase (HRP) goat anti-rabbit immunoglobulin G secondary
antibody (cat. no. BA1054; 1:5,000; Wuhan Boster Biological
Technology, Ltd.) was added for 1 h at 25°C. Enhanced
chemiluminescence substrate was used for protein visualization
(Pierce; Thermo Fisher Scientific, Inc.). The specific protein band
intensities were quantified by Image-Pro Plus 6.0 (Media
Cybernetics, Inc, Rockville, MD, USA).
Immunohistochemical analysis
Slices were subjected to treatment using 3% hydrogen
peroxide for 10 min at room temperature for endogenous peroxidase
inactivation. The sections were subsequently blocked using 1%
bovine serum albumin (Gibco; Thermo Fisher Scientific, Inc.) at
room temperature for 4 h to reduce non-specific staining. For
immunohistochemical staining, the sections were incubated with
primary antibodies against C5a (cat. no. ab11876; 1:1,000; Abcam)
for 8 h at 4°C. After washing with PBS, the slides were incubated
with goat anti-rabbit IgG biotinylated-modified secondary antibody
(cat. no. A27035; 1:2,000; Thermo Fisher Scientific, Inc.), and
conjugated HRP-labeled streptavidin (cat. no. K0609; 1:2,000; Dako;
Agilent Technologies, Inc., Santa Clara, CA, USA) was added.
Following staining, three high-magnification fields (×400) were
randomly selected in each slide, and the images were recorded using
an inverted light microscope (Zeiss AG, Oberkochen, Germany).
Masson staining
Paraffin sections underwent gradient dehydration by
incubating the slices in 100% xylene and a graded ethanol series
(100, 95, 90, 80 and 70%). Masson staining of Weigert's iron was
performed with a Masson Trichome Staining kit, according to the
manufacturer's protocol (cat no. M029; Shanghai Gefan Biotechnology
Co., Ltd., Shanghai, China) for 10 min at room temperature. The
stained slices underwent Ponceau S staining for 10 min at room
temperature, and treatment using phosphomolybdic acid for 5 min at
room temperature. Aniline blue staining was performed, and the
slices subsequently underwent differentiation in 1% glacial acetic
acid for 1 min. Slices were treated using dehydrated alcohol (95
and 100%) and 100% xylene. Slides were subsequently viewed under an
inverted light microscope.
Statistical analysis
All statistical analyses were performed with SPSS
19.0 software (IBM Corp., Armonk, NY, USA). The data are presented
as mean ± standard error of the mean from three separate
experiments. Statistical significance was determined by paired or
unpaired Student's t-test and one-way analysis of variance followed
by Tukey's post-hoc test in cases of standardized expression data.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Knee joint IM induces marked
destruction of the synovial fluid and cartilage
Morphological analysis was conducted to investigate
the effect of knee joint IM on the synovial fluid and cartilage.
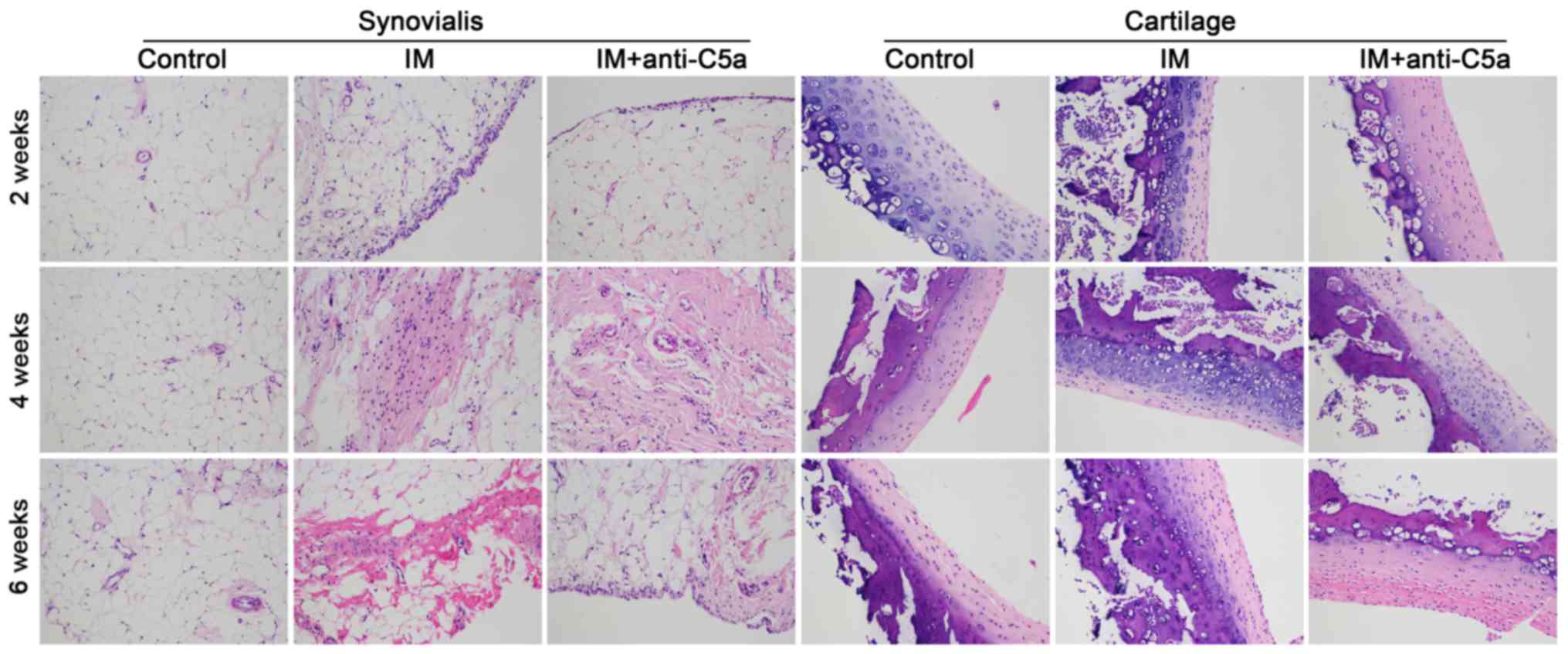
H&E and Masson staining were used. As presented in Fig. 1, prolonged knee joint IM resulted
in aggravation of the destruction of the synovial fluid and
cartilage in the IM group, while anti-C5a antibody treatment
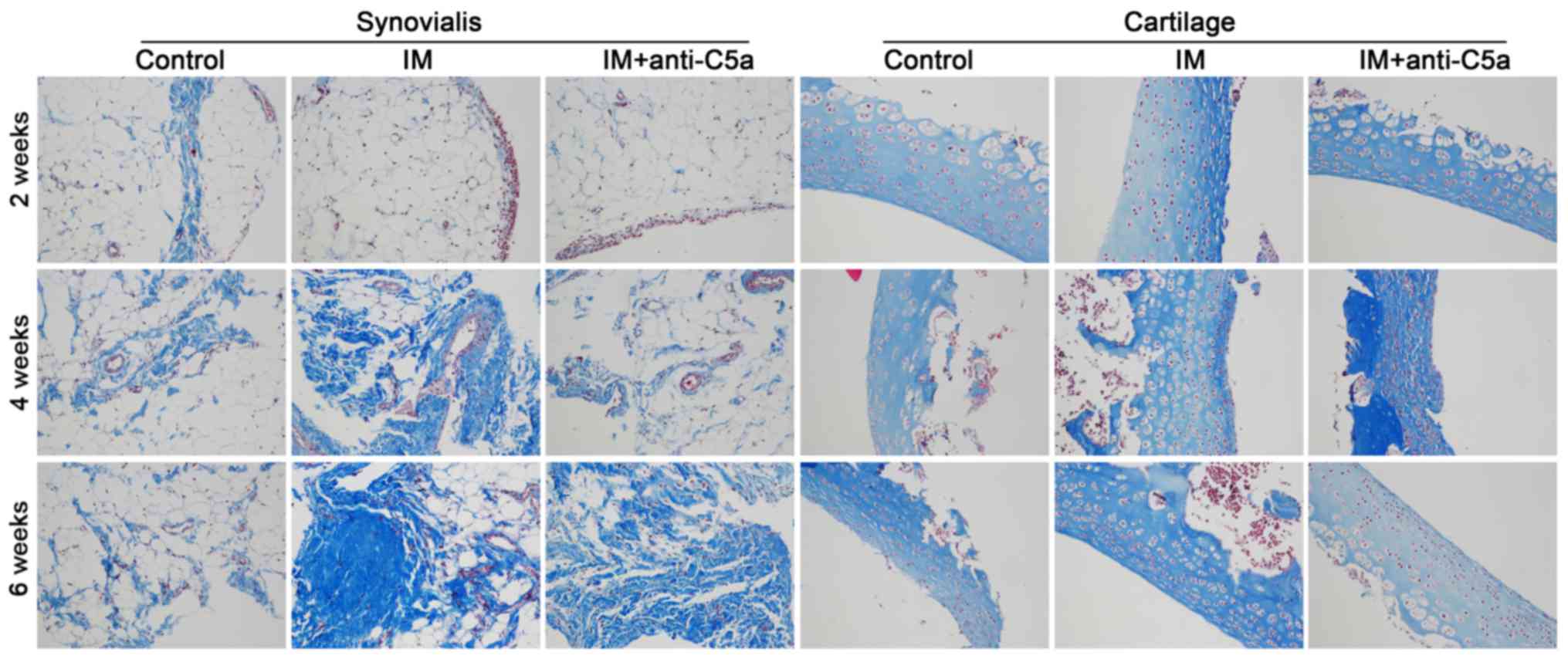
reversed these alterations. Masson staining revealed marked
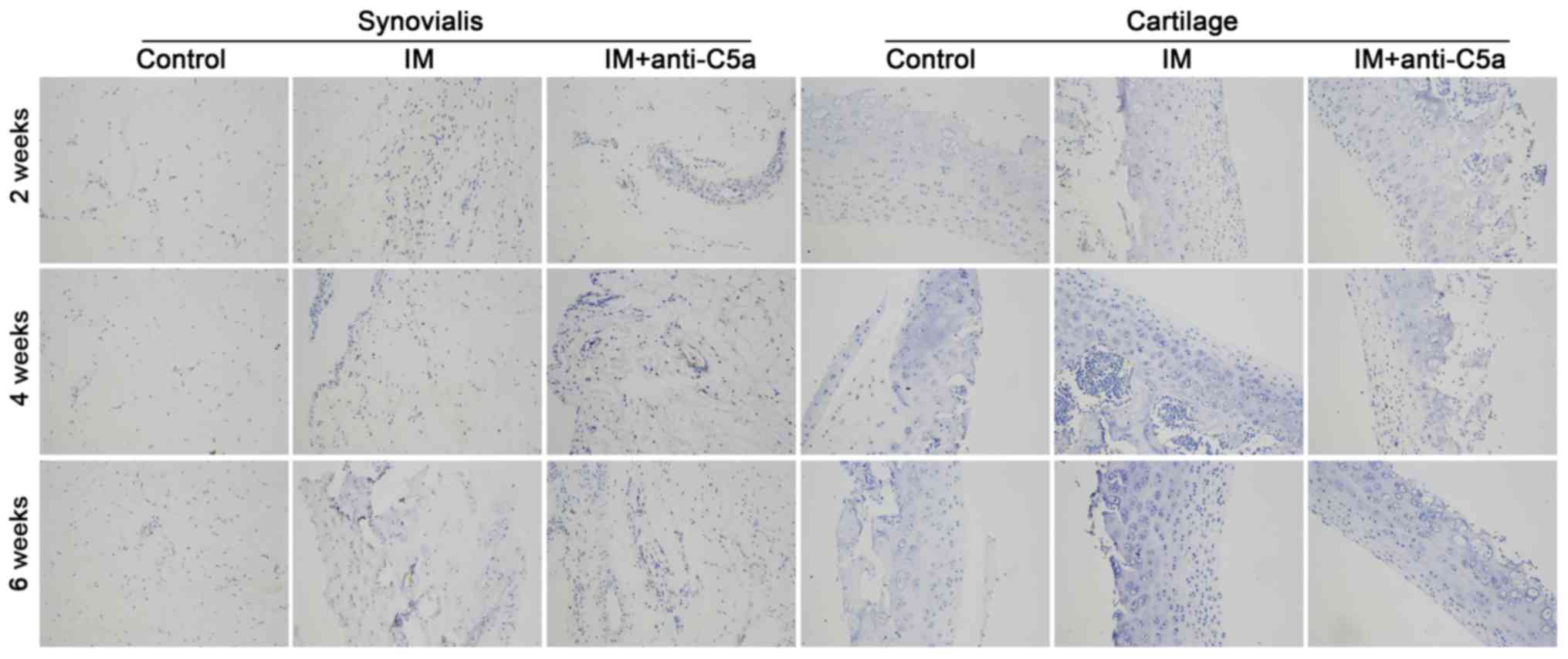
fibroplasia in the synovial membrane and cartilage (Fig. 2). TUNEL staining demonstrated
apoptosis in the synovial membrane and cartilage. As presented in
Fig. 3, the staining intensity in
the IM group was increased with prolonged treatment, indicating
increased cellular apoptosis in the synovial membrane and
cartilage, and treatment with anti-C5a mitigated these
alterations.
Increased C5a and pro-inflammatory
cytokines in the joint cavity fluid and serum in rats with knee
joint IM
The expression levels of C5a, IL-1β, TNF-α and
IL-17A in the serum (Fig. 4A) and
joint cavity fluid (Fig. 4B) were
evaluated using ELISA. The results revealed that following 1 week
of knee joint IM, the expression levels of C5a, IL-1β, TNF-α and
IL-17A in the joint cavity fluid and serum were significantly
increased compared with those in the control group (Fig. 4). These results indicated that knee
joint IM significantly induced inflammation in the knee joint. With
prolonged knee joint IM, the expression levels of C5a, IL-1β,
IL-17A and TNF-α in the IM group were gradually increased during
the experiment compared with the control and IM+anti-C5a groups.
However, the levels of C5a, IL-1β, IL-17A and TNF-α in the joint
cavity fluid of in IM+anti-C5a group were lower compared with those
in the IM group.
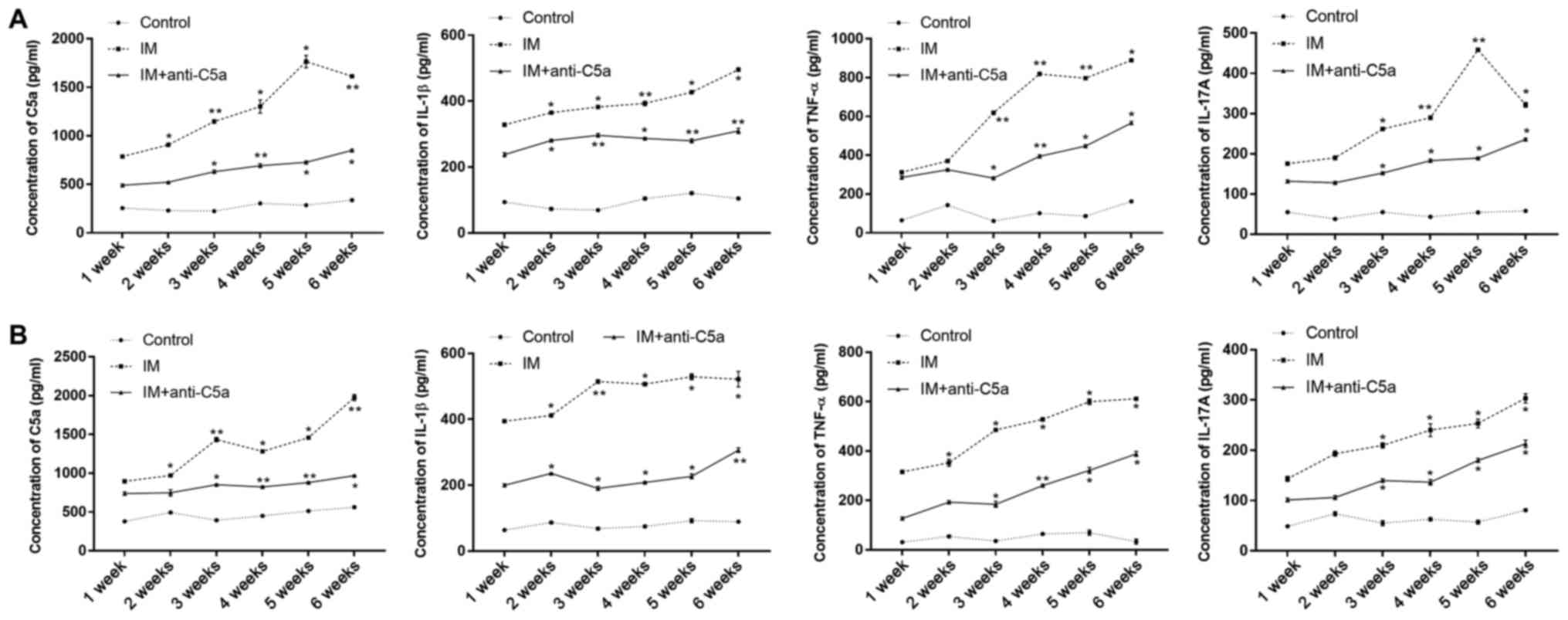 | Figure 4.Effects of joint IM on serum C5a and
pro-inflammatory cytokine expression levels. (A) Expression levels
of C5a, IL-1β, TNF-α and IL-17A in the serum; (B) expression levels
of C5a, IL-1β, TNF-α and IL-17A in joint cavity fluid. The graphs
present the mean ± SEM. *P<0.05, **P<0.01 vs. control group
at the same time point. The data are presented as the average of
triplicate values. Error bars represent the SEM. IM,
immobilization; C5a, complement component 5a; SEM, standard error
of the mean; IL, interleukin; TNF-α, tumor necrosis factor-α. |
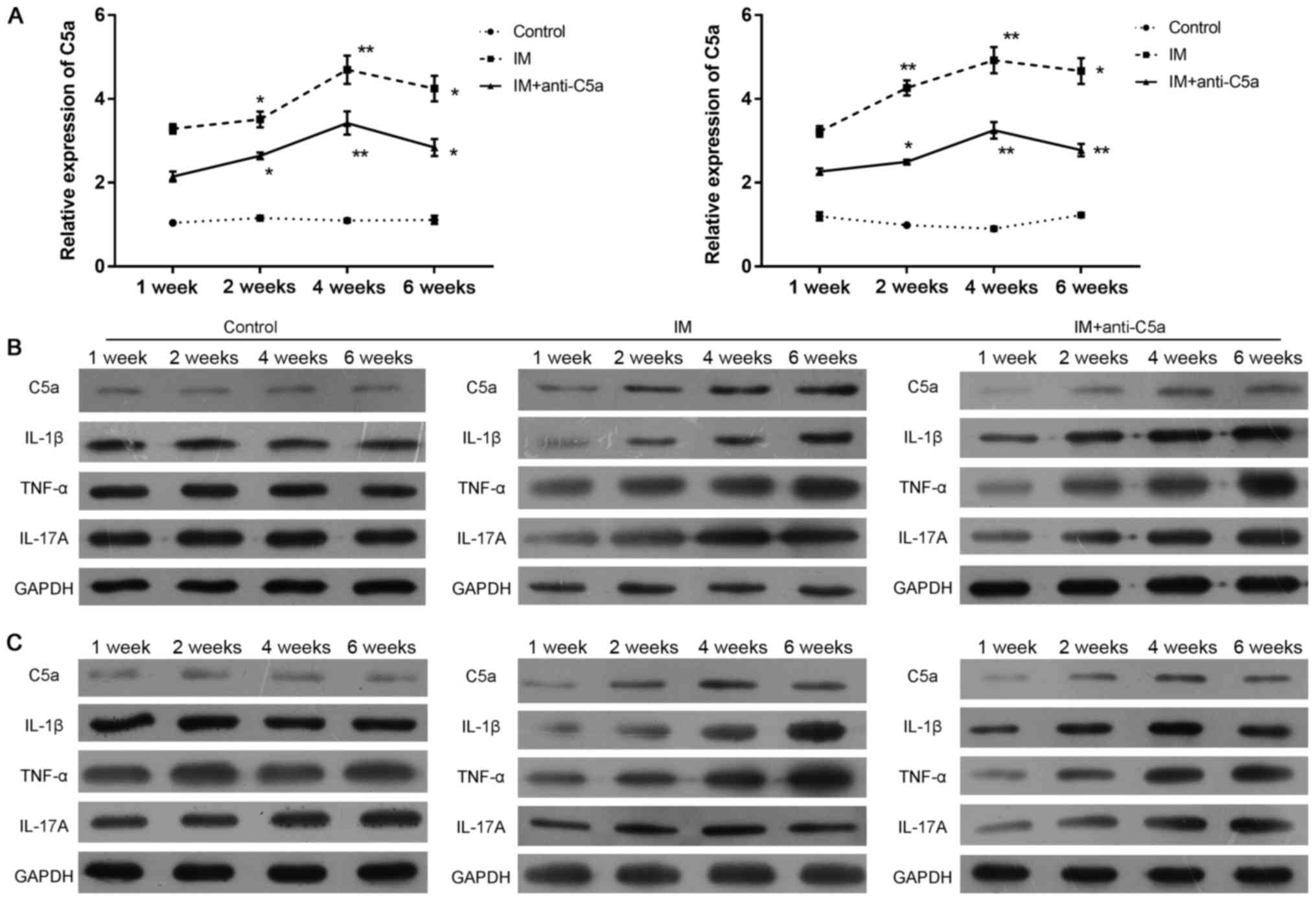
Increased levels of C5a in the
synovial membrane and cartilage in rats with knee joint
immobilization
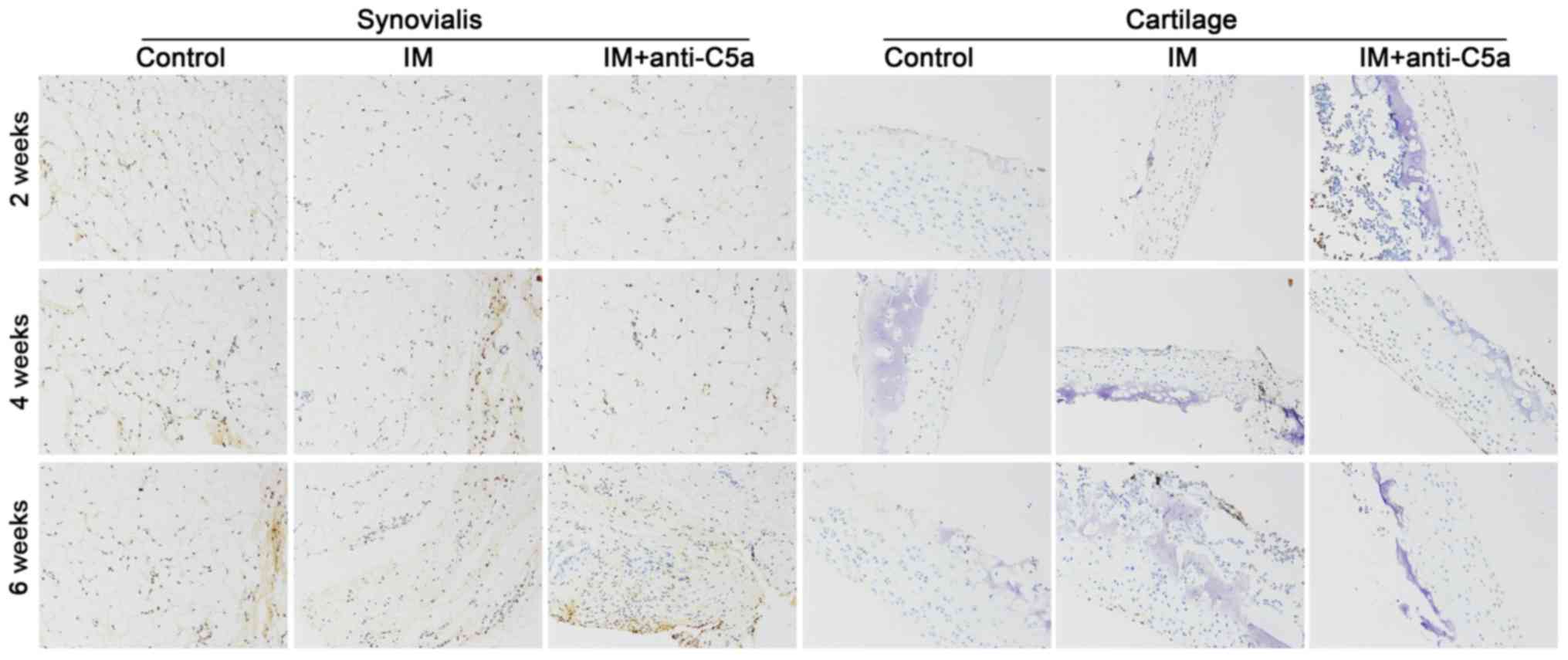
An RT-qPCR assay, western blot analysis and
immunohistochemistry were performed to assess the expression of C5a
in the synovialis and cartilage. The expression patterns of C5a in
the synovial fluid and cartilage in the rat knee joint following IM
with different treatments were evaluated by immunohistochemistry.
The results revealed that the expression of C5a was significantly
decreased in the IM+anti-C5a group compared with the IM group
(Fig. 5). In addition, the
expression of C5a mRNA was increased following 1 week of knee joint
IM in the IM and IM+anti-C5a groups in the synovial fluid (Fig. 6A, left) and cartilage (Fig. 6A, right), and western blotting
demonstrated similar results in the synovial fluid (Fig. 6B) and cartilage (Fig. 6C). These results confirmed the
findings that increased levels of C5a may serve an important role
in the development of chronic inflammation in the immobilized knee
joint, and dysfunction of the synovial fluid and cartilage induced
by OA.
Discussion
The present study investigated the role of C5a in
the development of OA in rats with joint IM. A knee cartilage
injury was induced that mimicked OA by fixing the rat knee joint at
an angle of 180°. In rats with knee joint IM, destruction of the
knee joint synovial fluid and cartilage was observed. The
expression levels of C5a in the serum and joint fluid were
assessed, and it was demonstrated that IM significantly increased
the expression levels of C5a. Immunohistochemistry, western
blotting and RT-qPCR analysis demonstrated a significant increase
in the expression of C5a. The application of anti-C5a antibody was
able to mitigate the IM-induced alterations in morphology and
cytokine expression in the knee joint synovial fluid and cartilage.
Joint IM significantly increased the expression levels of IL-1β,
IL-17A and TNF-α in the serum and joint fluid.
Dysfunction of articular cartilage remains the
characteristic pathology of OA (12,13).
Articular cartilage serves an extremely important role in
maintaining the physiological function of the knee joint (6,13,14).
Alterations in the immune system in the elderly population are
associated with the development of primary OA. Decreased T and B
cell function, and elevated pro-inflammatory cytokine expression
was observed in patients with OA (23–26).
These studies indicated that abnormal immune responses may
contribute to the development of cartilage.
The present study additionally demonstrated that
rats with knee joint IM exhibited destruction of the knee joint
synovial fluid and cartilage, increasing the levels of C5a in the
serum and joint fluid significantly. A central role for complement
in OA has been determined by a series of studies (27–31).
C5a is a highly inflammatory peptide that acts as a chemotactic
agent, and additionally serves an important role in innate immunity
(17,18). C5a was able to increase the
expression of adhesion molecules and trigger mast cell
degranulation, releasing proinflammatory molecules, including
histamine and TNF-α (32,33). C5a is an effective chemoattractant,
and initiates the accumulation of complement and phagocytic cells
at sites of infection or the recruitment of antigen-presenting
cells to the lymph nodes (34,35).
The complement system is activated in the synovial fluid of
subjects with knee injury and OA (30). Increased expression of C5aR is
observed in synoviocytes in joint inflammatory diseases (16,19,35).
C5a/C5aR interaction induces the release of TNF-α in activated
synoviocytes, promoting joint inflammatory processes (16,19,35).
The present study revealed the role of C5a in the development of
OA. The levels of a number of pro-inflammatory cytokines, including
IL-1β, TNF-α and IL-17A, were significantly increased during the
experiment, and the present results provided novel insights into
the hypothesis.
In the present study, joint IM rat models were used.
Joint IM has been reported to cause degeneration or atrophy of the
articular cartilage (36–38). Loading and movement remains
important for the maintenance of morphological and functional
integrity of the joint articular cartilage (7,39).
Immobilization was able to induce increased hydration, decreased
proteoglycan content, altered PG aggregate structure and increased
collagen synthesis, and was able to maintain or elevate the
collagen content of the articular cartilage (2,11,38).
Furthermore, following joint IM, destruction of the knee joint
synovial fluid and cartilage was observed. Masson staining revealed
increased collagen in the synovial fluid and cartilage, in addition
to alterations in pro-inflammatory cytokine expression. These
results suggested that the joint IM model was a suitable animal
model for OA.
In conclusion, the present study investigated the
role of the C5a pathway in the development of OA, and demonstrated
that the expression of C5a was increased in the synoviocytes and
cartilage in OA joint diseases. Furthermore, the release of a
number of pro-inflammatory cytokines, including TNF-α and IL-1β,
was induced in the activated synoviocytes via interactions with
C5a, suggesting that the C5a system may serve an important role in
joint inflammatory processes.
Acknowledgements
Not applicable.
Funding
The present was supported by Science and Technology
Cooperation Program of Guizhou Province [GZSC LH (2015). no. 7376],
Science Program of Guizhou Province [GZSS SY (2015). no. 3043] and
the Science and Technology Bureau of Guiyang [(2017). no.
30–35).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
WL and LW conceptualized the study design. WL, LW
and JY contributed to data analyses. WL and JY prepared the
manuscript. WL, CW and YC performed the data analyses and wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures were approved by the Guizhou Medical
University (Guiyang, China) Animal Care and Use Committees.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Behrens F, Kraft EL and Oegema TR Jr:
Biochemical changes in articular cartilage after joint
immobilization by casting or external fixation. J Orthop Res.
7:335–343. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tammi M, Säämänen AM, Jauhiainen A,
Malminen O, Kiviranta I and Helminen H: Proteoglycan alterations in
rabbit knee articular cartilage following physical exercise and
immobilization. Connect Tissue Res. 11:45–55. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tammi M, Kiviranta I, Peltonen L, Jurvelin
J and Helminen HJ: Effects of joint loading on articular cartilage
collagen metabolism: Assay of procollagen prolyl 4-hydroxylase and
galactosylhydroxylysyl glucosyltransferase. Connect Tissue Res.
17:199–206. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Videman T, Eronen I and Candolin T:
[3H]proline incorporation and hydroxyproline concentration in
articular cartilage during the development of osteoarthritis caused
by immobilization. A study in vivo with rabbits. Biochem J.
200:435–440. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jurvelin J, Kiviranta I, Säämänen AM,
Tammi M and Helminen HJ: Partial restoration of
immobilization-induced softening of canine articular cartilage
after remobilization of the knee (stifle) joint. J Orthop Res.
7:352–358. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Haapala J, Arokoski JP, Hyttinen MM, Lammi
M, Tammi M, Kovanen V, Helminen HJ and Kiviranta I: Remobilization
does not fully restore immobilization induced articular cartilage
atrophy. Clin Orthop Relat Res. 218–229. 1999.PubMed/NCBI
|
|
7
|
Jortikka MO, Inkinen RI, Tammi MI,
Parkkinen JJ, Haapala J, Kiviranta I, Helminen HJ and Lammi MJ:
Immobilisation causes longlasting matrix changes both in the
immobilised and contralateral joint cartilage. Ann Rheum Dis.
56:255–261. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vanwanseele B, Eckstein F, Knecht H,
Stussi E and Spaepen A: Knee cartilage of spinal cord-injured
patients displays progressive thinning in the absence of normal
joint loading and movement. Arthritis Rheum. 46:2073–2078. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hagiwara Y, Ando A, Chimoto E, Saijo Y,
Ohmori-Matsuda K and Itoi E: Changes of articular cartilage after
immobilization in a rat knee contracture model. J Orthop Res.
27:236–242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pritzker KP: Animal models for
osteoarthritis: Processes, problems and prospects. Ann Rheum Dis.
53:406–420. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wong K, Sun F, Trudel G, Sebastiani P and
Laneuville O: Temporal gene expression profiling of the rat knee
joint capsule during immobilization-induced joint contractures. BMC
Musculoskelet Disord. 16:1252015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Misra D, Felson DT, Silliman RA, Nevitt M,
Lewis CE, Torner J and Neogi T: Knee osteoarthritis and frailty:
Findings from the Multicenter Osteoarthritis Study and
Osteoarthritis Initiative. J Gerontol A Biol Sci Med Sci.
70:339–344. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goldring MB and Goldring SR: Articular
cartilage and subchondral bone in the pathogenesis of
osteoarthritis. Ann N Y Acad Sci. 1192:230–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Haseeb A and Haqqi TM: Immunopathogenesis
of osteoarthritis. Clin Immunol. 146:185–196. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Berenbaum F: Osteoarthritis as an
inflammatory disease (osteoarthritis is not osteoarthrosis!).
Osteoarthritis Cartilage. 21:16–21. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu-Bryan R: Synovium and the innate
inflammatory network in osteoarthritis progression. Curr Rheumatol
Rep. 15:3232013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nishiura H: The alternative C5a receptor
function. Adv Exp Med Biol. 735:111–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Woodruff TM, Nandakumar KS and Tedesco F:
Inhibiting the C5-C5a receptor axis. Mol Immunol. 48:1631–1642.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan G, Wei J, Zhou J, Hu H, Tang Z and
Zhang G: Expression of C5aR (CD88) of synoviocytes isolated from
patients with rheumatoid arthritis and osteoarthritis. Chin Med J
(Engl). 116:1408–1412. 2003.PubMed/NCBI
|
|
20
|
Pujol JP and Loyau G: Interleukin-1 and
osteoarthritis. Life Sci. 41:1187–1198. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
National Institute of Allergy and
Infectious Diseases NIoH. Bethesda, Md: National Institutes of
Health (1996) Guide for the care and use of laboratory animals;
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ponchel F, Burska AN, Hensor EM, Raja R,
Campbell M, Emery P and Conaghan PG: Changes in peripheral blood
immune cell composition in osteoarthritis. Osteoarthritis
Cartilage. 23:1870–1878. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nykula TD, Bychkov OA and Bychkova NH: The
dynamics of indices of immune and cytokine status for patients with
essential hypertension in combination with osteoarthritis. Lik
Sprava. 90–93. 2013.(In Ukrainian). PubMed/NCBI
|
|
25
|
Kandahari AM, Yang X, Dighe AS, Pan D and
Cui Q: Recognition of immune response for the early diagnosis and
treatment of osteoarthritis. J Immunol Res. 2015:1924152015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
de Lange-Brokaar BJ, Ioan-Facsinay A, van
Osch GJ, Zuurmond AM, Schoones J, Toes RE, Huizinga TW and
Kloppenburg M: Synovial inflammation, immune cells and their
cytokines in osteoarthritis: A review. Osteoarthritis Cartilage.
20:1484–1499. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Q, Rozelle AL, Lepus CM, Scanzello
CR, Song JJ, Larsen DM, Crish JF, Bebek G, Ritter SY, Lindstrom TM,
et al: Identification of a central role for complement in
osteoarthritis. Nat Med. 17:1674–1679. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Buckland J: Osteoarthritis:
Complement-mediated inflammation in OA progression. Nat Rev
Rheumatol. 8:22011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu-Bryan R and Terkeltaub R: Emerging
regulators of the inflammatory process in osteoarthritis. Nat Rev
Rheumatol. 11:35–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Struglics A, Okroj M, Swärd P, Frobell R,
Saxne T, Lohmander LS and Blom AM: The complement system is
activated in synovial fluid from subjects with knee injury and from
patients with osteoarthritis. Arthritis Res Ther. 18:2232016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang X, Hunter D, Xu J and Ding C:
Metabolic triggered inflammation in osteoarthritis. Osteoarthritis
Cartilage. 23:22–30. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kuna P, Reddigari SR, Schall TJ, Rucinski
D, Sadick M and Kaplan AP: Characterization of the human basophil
response to cytokines, growth factors, and histamine releasing
factors of the intercrine/chemokine family. J Immunol.
150:1932–1943. 1993.PubMed/NCBI
|
|
33
|
Stone SF, Isbister GK, Shahmy S, Mohamed
F, Abeysinghe C, Karunathilake H, Ariaratnam A, Jacoby-Alner TE,
Cotterell CL and Brown SG: Immune response to snake envenoming and
treatment with antivenom; complement activation, cytokine
production and mast cell degranulation. PLoS Negl Trop Dis.
7:e23262013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Michel O, Ginanni R, Le Bon B, Content J,
Duchateau J and Sergysels R: Inflammatory response to acute
inhalation of endotoxin in asthmatic patients. Am Rev Respir Dis.
146:352–357. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cavaillon JM, Fitting C and
Haeffner-Cavaillon N: Recombinant C5a enhances interleukin 1 and
tumor necrosis factor release by lipopolysaccharide-stimulated
monocytes and macrophages. Eur J Immunol. 20:253–257. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kiviranta I, Tammi M, Jurvelin J, Arokoski
J, Saamanen AM and Helminen HJ: Articular cartilage thickness and
glycosaminoglycan distribution in the young canine knee joint after
remobilization of the immobilized limb. J Orthop Res. 12:161–167.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Maldonado DC, Silva MC, Neto Sel-R, de
Souza MR and de Souza RR: The effects of joint immobilization on
articular cartilage of the knee in previously exercised rats. J
Anat. 222:518–525. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nomura M, Sakitani N, Iwasawa H, Kohara Y,
Takano S, Wakimoto Y, Kuroki H and Moriyama H: Thinning of
articular cartilage after joint unloading or immobilization. An
experimental investigation of the pathogenesis in mice.
Osteoarthritis Cartilage. 25:727–736. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Müller FJ, Setton LA, Manicourt DH, Mow
VC, Howell DS and Pita JC: Centrifugal and biochemical comparison
of proteoglycan aggregates from articular cartilage in experimental
joint disuse and joint instability. J Orthop Res. 12:498–508. 1994.
View Article : Google Scholar : PubMed/NCBI
|