Introduction
MicroRNAs (miRNAs/miRs) consist of a large family of
non-coding, single-stranded RNA molecules that are 18–22
nucleotides long. These RNA molecules modulate gene expression
post-transcriptionally by regulating the translation and stability
of mRNAs. This regulation is achieved through complete or
incomplete sequence matching between miRNAs and the 3′untranslated
regions of the target mRNA (1). So
far, thousands of mature miRNAs have been identified, of which
~2,600 human miRNAs have been annotated in the miRbase database
(http://www.mirbase.org/).
The incidence of papillary thyroid carcinoma (PTC)
continues to increase worldwide (2), and it has been demonstrated that
miRNAs may contribute to occurrence and progression of the disease.
Differential miRNA expression has been reported in PTC vs. normal
tissue samples (3–5); certain miRNAs, including serum
miR-34a, −155, −197, −221, −224 and −375, can be used as diagnostic
and prognostic indicators of thyroid cancer (6,7). The
mechanism of action for miR-150, −422a, −18a and −19a in human
thyroid cancer has been studied to some extent (8–10);
however, there are still a number of miRNAs that may be
biologically relevant for the development of thyroid cancer.
Human miR-486 is situated on the short arm of
chromosome 8p11 within the ankyrin 1 gene and is transcribed from
an intron (11). Genomic deletion
of miR-486-5p has been associated with various cancers, including
colorectal, lung, pancreatic and gastric cancer (12–16).
However, to the best of our knowledge, only one study to date has
investigated the function of miR-486-5p in thyroid cancer. Ma et
al (17) demonstrated that the
fibrillin 1 (FBN1) gene is a distinct molecular target for
miR-486-5p in PTC cells. In addition, reduced miR-486-5p expression
levels were identified in PTC tissues and cell lines, and
miR-486-5p was demonstrated to suppress cell growth and enhance
apoptosis in PTC. A limitation of this study was the small sample
size; therefore, it is necessary to further explore the expression
of miR-486-5p in thyroid cancer in a larger cohort to better
understand the underlying molecular mechanisms.
The Cancer Genome Atlas (TCGA; http://cancergenome.nih.gov/) is a comprehensive
database of key genetic mutations responsible for different types
of cancer, which is generated by the National Cancer Institute and
the National Human Genome Research Institute (18). The Gene Expression Omnibus (GEO;
http://www.ncbi.nlm.nih.gov/geo/) was
released by the National Center for Biotechnology Information in
2000 and is a public functional genomics data repository, which
contains gene chip expression data as well as data from non-chip
technologies, including serial analysis of gene and mass
spectrometry (19). ArrayExpress
(https://www.ebi.ac.uk/arrayexpress/)
is an archive of functional genomics data from microarray and
sequencing platforms (20). These
databases offer an excellent source of information for studying the
importance of miRNAs in PTC.
TCGA database was used to evaluate the expression
levels of miR-486-5p in 507 patients with PTC and to analyze its
association with clinical parameters. In addition, miR-486-5p
expression data were extracted, and verified by GEO and
ArrayExpress. Subsequently, the miR-486-5p target mRNAs were
identified through 12 miRNA-mRNA prediction platforms. Finally, the
potential molecular mechanisms by which miR-486-5p may contribute
to PTC were investigated using bioinformatics tools.
Materials and methods
TCGA miR-486-5p data analysis
TCGA contains miRNA sequencing data from 507 PTC and
59 normal thyroid samples (21).
In the present study, miR-486-5p expression data, as well as
clinicopathological features from PTC vs. normal thyroid samples,
were extracted from TCGA by the end of October 1, 2017. Follow-up
cases over a 5-year period from October 1, 2012 were also included.
The expression data for miR-486-5p were log2 transformed and values
<1 were censored. Student's t-test was conducted to compare the
expression levels of miR-486-5p in 507 PTC and 59 normal thyroid
samples, as well as the association between miR-486-5p expression
and the clinical parameters from TCGA. The cut-off value for high
and low miR-486-5p mRNA expression was based on median expression
in the PTC group. Receiver operating characteristic (ROC) and
summary receiver operating characteristic (SROC) curves were
generated to evaluate the accuracy of miR-486-5p in identifying
cancer and normal tissue. Kaplan-Meier (K-M) analysis and log-rank
test were used to assess the prognostic value of miR-486-5p. SPSS
version 22.0 (IBM Corp., Armonk, NY, USA) was utilized for
statistical analyses and P<0.05 was considered to indicate a
statistically significant difference.
Validation of miR-486-5p expression
using GEO and ArrayExpress
The GEO and ArrayExpress databases were searched for
PTC-relevant miRNA chip or sequencing data using the following
phrases: [(thyroid OR papillary OR follicular OR medullary) AND
(tumor OR tumour OR cancer OR carcinoma OR neopla* OR malignan*)
AND (miRNA OR miR OR microRNA) AND (profil* OR array OR chip OR
microarray OR microchip)]. ‘*’ refers to wildcard. All expression
and clinical data related to miR-486-5p were extracted.
Meta-analysis of GEO (GSE40807 (22), GSE57780 (unpublished data),
GSE62054 (unpublished data), GSE73182 (23), ArrayExpress (E-MTAB-736) (24) and TCGA miRNA data was carried out
in Stata version 12.0 (StataCorp LP, College Station, TX, USA). The
pooled standard mean difference (SMD) with 95% confidence interval
(CI) was utilized to assess miR-486-5p expression in PTC vs. normal
thyroid tissue. χ2 and I2 statistics were
calculated to measure the heterogeneity within the meta-analysis.
The Mante-Haenszel fixed-effects model was applied if there was no
obvious heterogeneity among the pooled studies (χ2 test
P>0.1 and I2 <50%); conversely, a random-effects
model was applied when obvious heterogeneity was identified
(χ2 test P<0.1 and I2>50%) (25).
Predicting miR-486-5p target
genes
To identify potential miR-486-5p target genes, 12
miR-target prediction programs, including TDIANA-microTv4.0
(26), DIANA-microT-CDS (27), miRanda-rel2010 (28), miRBridge (29), miRDB4.0 (30), miRmap (31), miRNAMap (32), doRiNA (33), PITA (34), RNA22v2 (35), RNAhybrid2.1 (36) and Targetscan6.2 (37) were used. The 80 mRNAs predicted by
at least nine programs were considered potential miR-486-5p target
mRNAs in PTC and used for further mRNA functional analysis.
Predicting miR-486-5p biological
function
To evaluate the potential molecular mechanisms that
are regulated by miR-486-5p-associated genes, FunRich version 3.0
software (http://www.funrich.org/) was used for
Gene Ontology (GO) functional enrichment analysis (38). The software supports enrichment
analysis of GO terms, including those involved in biological
process, cellular component and molecular function, which can also
be visualized. The results were considered statistically
significant when the false discovery rate (FDR) was <0.05. The
Database for Annotation, Visualization and Integrated Discovery
(DAVID) version 6.8 (https://david.ncifcrf.gov/) and STRING version 10.5
(https://string-db.org/) were used for Kyoto
Encyclopedia of Genes and Genomes (KEGG) enrichment analysis
(39,40). Genes in the top KEGG pathway were
considered to be miR-486-5p hub genes.
Validation of the hub gene expression
levels and their relationship with miR-486-5p
The cBioPortal for Cancer Genomics (http://www.cbioportal.org/), which is a public
resource for large-scale cancer genomics data, was used to explore
hub gene expression in PTC tissues (41). To ascertain the correlation between
miR-486-5p and hub genes, a Pearson's correlation analysis was
conducted; P<0.05 was considered to indicate a statistically
significant difference. The expression of the hub genes in PTC and
normal thyroid tissue was examined by immunohistochemistry via The
Human Protein Atlas (http://www.proteinatlas.org/) (42).
Results
Downregulated miR-486-5p expression in
PTC and its clinical significance
Data on miR-486-5p gene expression from 507 PTC and
59 normal thyroid samples were extracted from TCGA. The expression
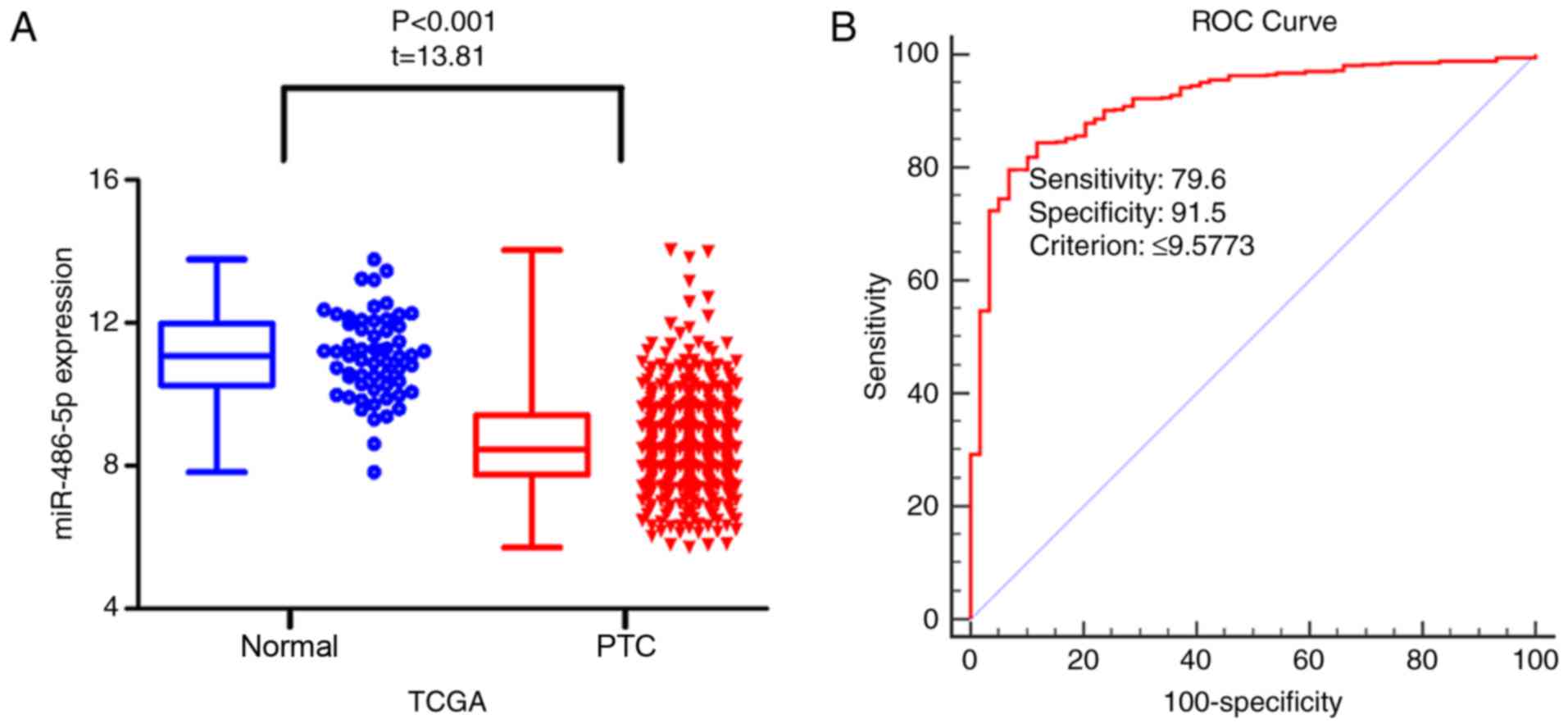
levels of miR-486-5p were significantly decreased in PTC compared
with in normal tissue [8.47±1.30 vs. 11.08±1.18; P<0.001; Fold
change (FC)=0.76; Table I;
Fig. 1A]. ROC curve analysis of
miR-486-5p in discriminating PTC from normal tissue was calculated
with an area under curve (AUC) of 0.918 (P<0.001; 95% CI:
0.892–0.939; Fig. 1B). The
miR-486-5p expression levels and clinicopathological factors are
summarized in Table I. miR-486-5p
expression levels were decreased in stage III–IV cancers
(8.36±1.26) compared with in stage I–II cancers (8.72±1.30;
P=0.003). In addition, PTC tissues with pathological lymph nodes
had decreased expression levels of miR-486-5p (8.49±1.20) compared
with PTC tissues without pathological lymph nodes (8.72±1.28;
P=0.047). Furthermore, miR-486-5p was downregulated in tissues with
metastasis (7.73±0.99) compared with in tissues without metastasis
(8.61±1.29; P=0.042). Decreased expression levels of miR-486-5p
were observed in PTC group (8.12±1.17) compared with the normal
thyroid group (8.63±1.29; P=0.012). miR-486-5p expression was also
reduced in recurrent cancer (8.15±0.90) compared with non-recurrent
cancer (8.64±1.32; P=0.016). Differential miR-486-5p expression was
not observed for the remaining clinicopathological factors analyzed
(Table I). Subsequently, the
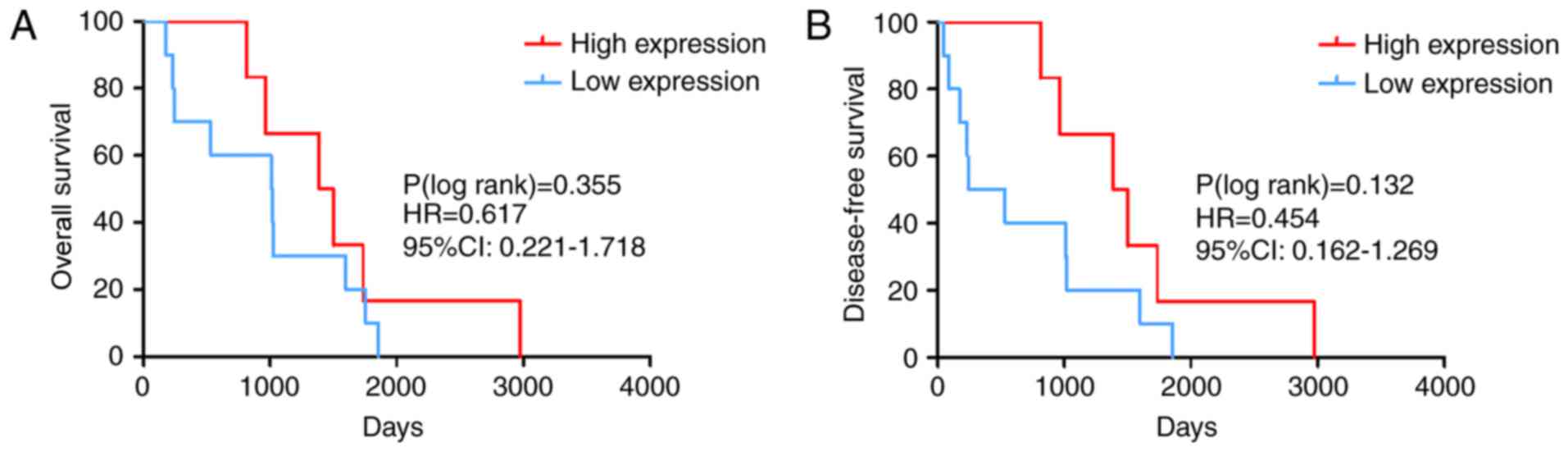
prognostic value of miR-486-5p expression was investigated. The K-M
survival curves indicated that the median overall survival (OS) for
the high expression group was 1,443 days, whereas the median OS for
the low expression group was 1,015 days. The curves suggested that
PTC cases with higher miR-486-5p expression levels were likely to
have an improved clinical outcome. However, there was no
statistically significant difference in the OS [log-rank P=0.355;
hazard ratio (HR)=0.617; 95% CI: 0.221–1.718; Fig. 2A] and the disease-free survival
(DFS) (log-rank P=0.132; HR=0.454; 95% CI: 0.162–1.269; Fig. 2B) of samples with high vs. low
expression levels of miR-486-5p (cut-off =8.469). Therefore, the
ability for miR-486-5p expression to predict OS and DFS is
limited.
 | Table I.Association between miR-486-5p
expression and clinicopathological factors in PTC. |
Table I.
Association between miR-486-5p
expression and clinicopathological factors in PTC.
|
|
| Relative miR-486-5p
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | No. | Mean ± standard
deviation | t-value | P-value |
|---|
| Tissue |
|
|
|
|
|
Normal | 59 | 11.08±1.18 | 13.81 | <0.001 |
|
PTC | 507 | 8.47±1.30 |
|
|
| Age (years) |
|
|
|
|
|
≤60 | 393 | 8.61±1.27 | 0.33 | 0.740 |
|
>60 | 113 | 8.56±1.36 |
|
|
| Sex |
|
|
|
|
|
Female | 370 | 8.64±1.31 | 1.28 | 0.203 |
|
Male | 136 | 8.48±1.24 |
|
|
| Stage |
|
|
|
|
|
I–II | 336 | 8.72±1.30 | 2.94 | 0.003 |
|
III–IV | 168 | 8.36±1.26 |
|
|
| Grade |
|
|
|
|
|
I–II | 311 | 8.69±1.29 | 1.92 | 0.056 |
|
III–V | 195 | 8.46±1.29 |
|
|
| Pathological lymph
node |
|
|
|
|
| No | 231 | 8.72±1.28 | 1.99 | 0.047 |
|
Yes | 225 | 8.49±1.20 |
|
|
| Metastasis |
|
|
|
|
| No | 496 | 8.61±1.29 | 2.04 | 0.042 |
|
Yes | 9 | 7.73±0.99 |
|
|
| Tumor |
|
|
|
|
| No | 450 | 8.63±1.29 | 2.51 | 0.012 |
|
Yes | 44 | 8.12±1.17 |
|
|
| Recurrence |
|
|
|
|
| No | 462 | 8.64±1.32 | 2.41 | 0.016 |
|
Yes | 44 | 8.15±0.90 |
|
|
Verification of miR-486-5p
downregulation in PTC using GEO and ArrayExpress data
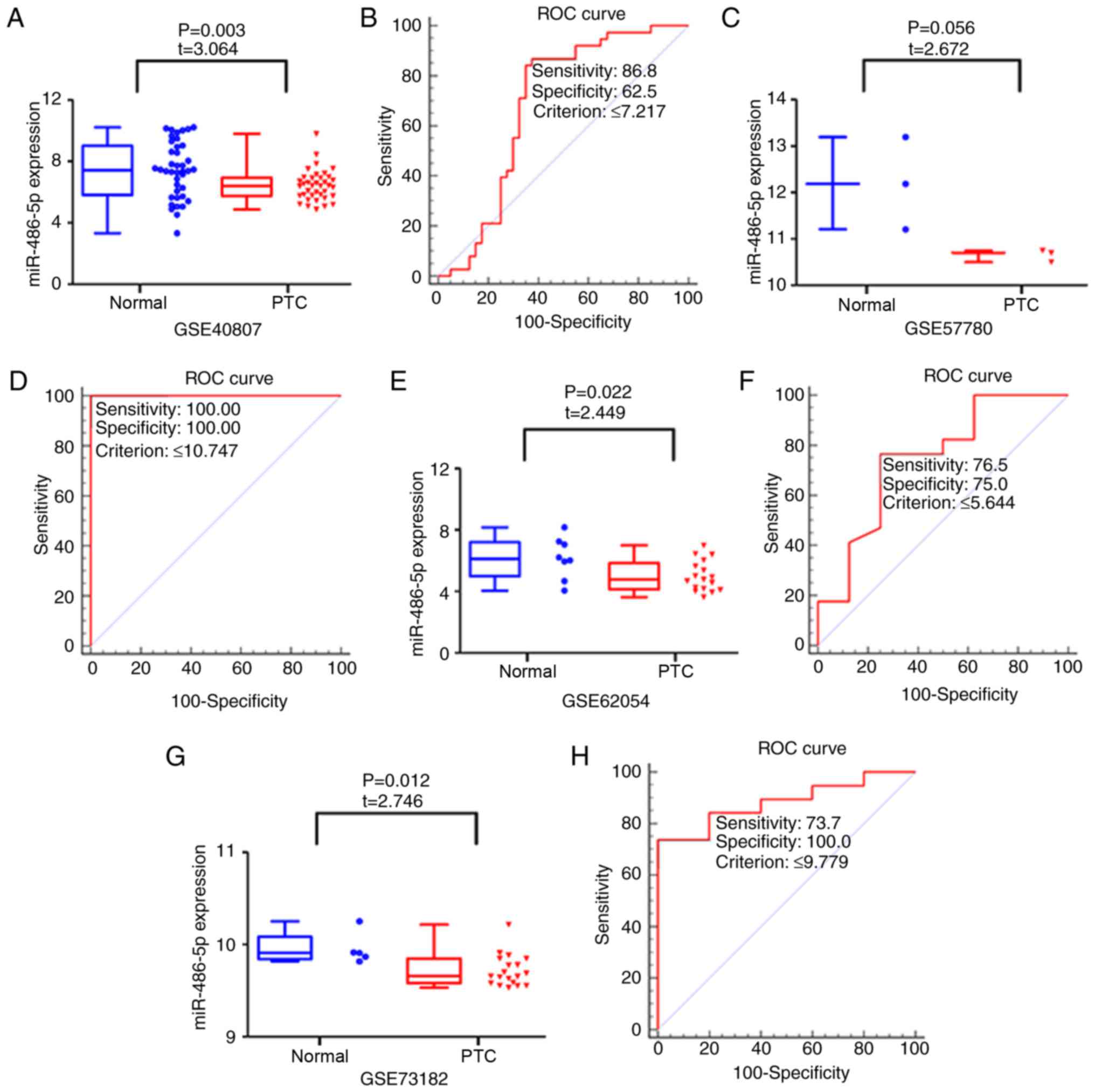
GSE40807, GSE57780, GSE62054 and GSE73182 were
obtained from the GEO database, which provided miR-486-5p
expression levels in 77 PTC and 56 normal thyroid tissues (Fig. 3). The expression levels of
miR-486-5p were lower in PTC compared with in normal samples in the
following datasets: GSE40807 (6.426±0.992 vs. 7.450±1.819; P=0.003;
FC=1.159; Fig. 3A), GSE62054
(5.000±1.003 vs. 6.176±1.349; P=0.022; FC=0.810; Fig. 3E) and GSE73182 (9.716±0.171 vs.
9.952±0.173; P=0.012; FC=0.976; Fig.
3G). The expression levels of miR-486-5p did not differ
significantly between PTC and normal tissue in the GSE57780 dataset
(10.648±0.133 vs. 12.195±0.994; P=0.056; FC=0.873; Fig. 3C). Chip array data from a single
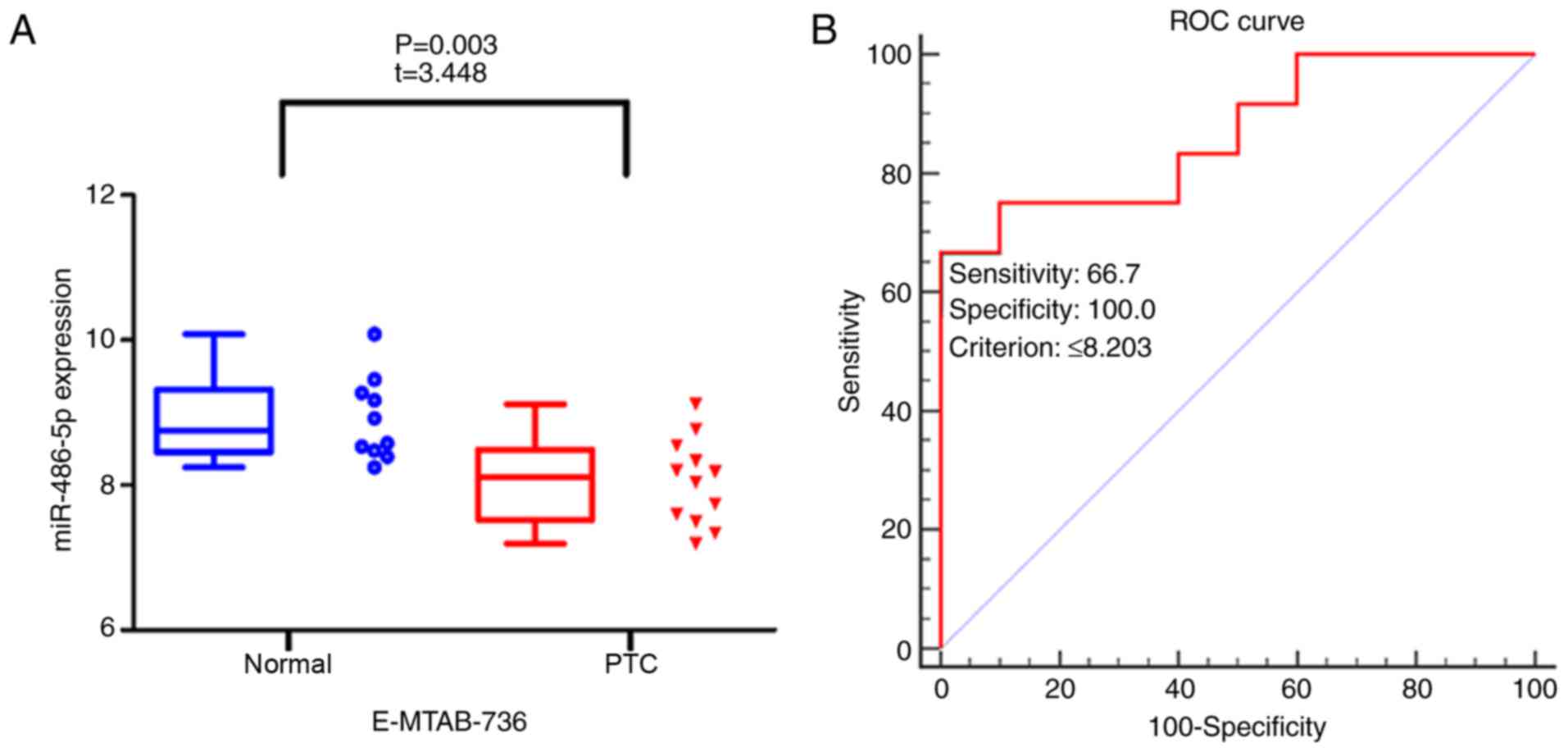
study (E-MTAB-736) was obtained from ArrayExpress and the data also
indicated that miR-486-5p expression levels were downregulated in
the 12 PTC samples compared with in the 10 normal tissue samples
(8.048±0.593 vs. 8.914±0.579; P=0.003; FC=0.903; Fig. 4A). To evaluate the ability of
miR-486-5p to distinguish cancer from normal thyroid samples, ROC
curve analyses were performed. The AUC values for miR-486-5p
calculated using the GSE40807, GSE57780, GSE62054, GSE73182 and
E-MTAB-736 datasets were 0.684 (P<0.001; cut-off ≤7.217;
Fig. 3B), 1.000 (P<0.001;
cut-off ≤10.747; Fig. 3D), 0.746
(P=0.036; cut-off ≤5.644; Fig.
3F), 0.884 (P<0.001; cut-off ≤9.779; Fig. 3H) and 0.867 (P<0.001; cut-off
≤8.203; Fig. 4B),
respectively.
Further verification of miR-486-5p
downregulation in PTC using SMD and SROC
miR-486-5p expression data from TCGA, GEO (GSE40807,
GSE57780, GSE62054 and GSE73182) and ArrayExpress (E-MTAB-736) were
combined for meta-analysis, which included a total of 603 PTC and
125 normal tissue samples (Table
II). The pooled SMD of miR-486-5p was −1.358 [95% CI,
−1.950-(−0.766); P<0.001; Fig.
5A] by the random effects model, and the P-value of the
heterogeneity test was 0.001 (I2=75.6%). From
meta-analysis, the AUC of the SROC was 0.84 (95% CI, 0.81–0.87;
P<0.001; Fig. 5B).
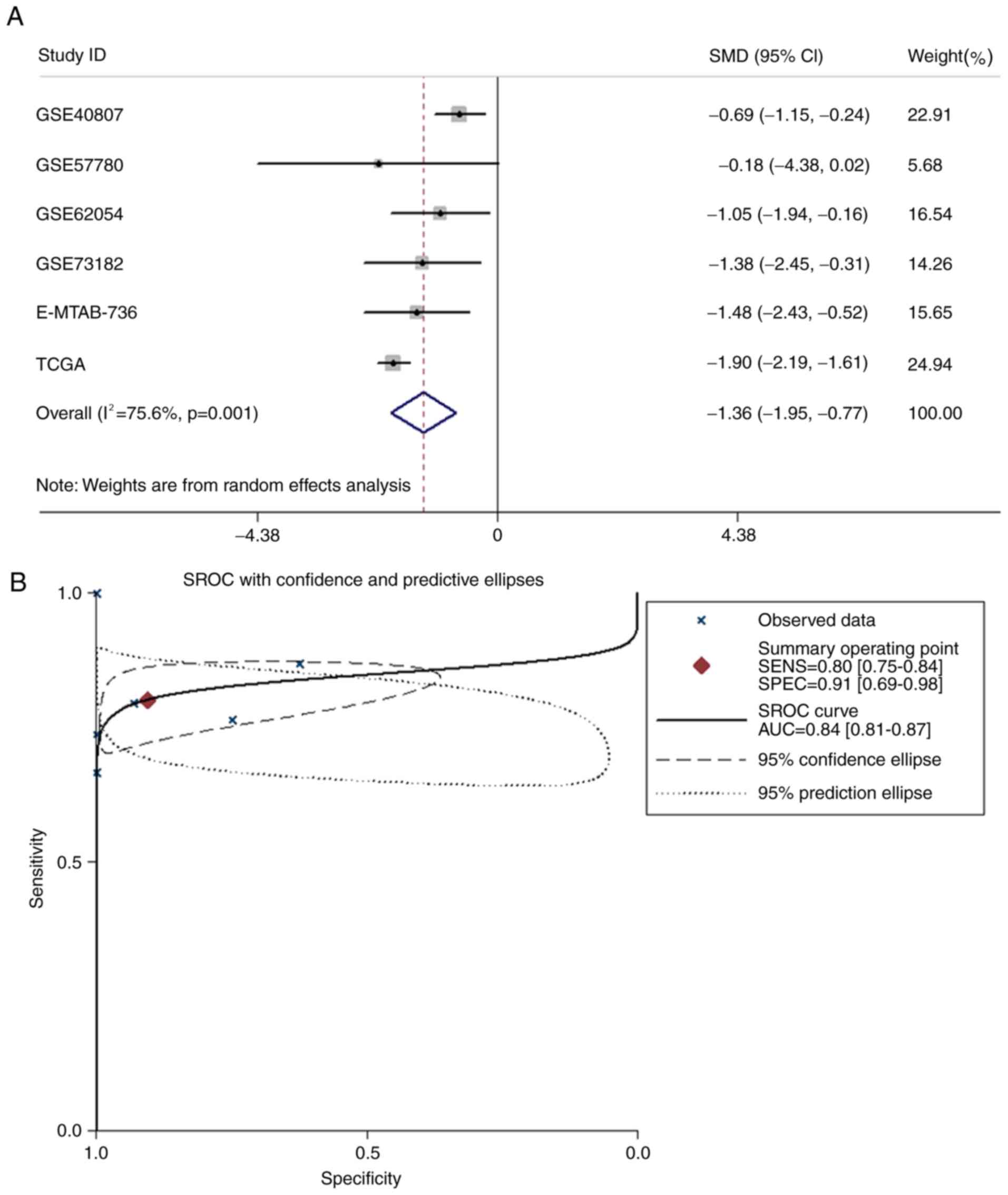 | Figure 5.Meta-analysis of miR-486-5p
expression levels in PTC. (A) Forest plot of all datasets
evaluating miR-486-5p expression between PTC and normal thyroid
tissue. (B) SROC curve for miR-486-5p in the diagnosis of PTC for
all studies. Datasets from TCGA, GSE40807, GSE57780, GSE62054,
GSE73182 and E-MTAB-736 were included in the meta-analysis. AUC,
area under curve; CI, confidence interval; miR, microRNA; PTC,
papillary thyroid carcinoma; SROC, summary receiver operating
characteristic; TCGA, The Cancer Genome Atlas. |
| Table II.Microarray and RNA sequencing
datasets. |
Table II.
Microarray and RNA sequencing
datasets.
|
|
|
|
| PTC | Normal |
|
|
|
|
|
|---|
|
|
|
|
|
|
|
|
|
|
|
|
|---|
| Contributor
(s) | Country | Year | Dataset | No. | Mean | SD | No. | Mean | SD | TP | FP | FN | TN | (Refs.) |
|---|
| Lassalle et
al | France | 2014 | GSE40807 | 38 | 6.426 | 0.992 | 40 | 7.450 | 1.819 | 33 | 15 | 5 | 25 | (22) |
| Saiselet et
al | Belgium | 2015 | GSE57780 | 3 | 10.648 | 0.133 | 3 | 12.195 | 0.994 | 3 | 0 | 0 | 3 | Unpublished
data |
| Stokowy et
al | Norway | 2014 | GSE62054 | 17 | 5.000 | 1.003 | 8 | 6.176 | 1.349 | 13 | 2 | 4 | 6 | Unpublished
data |
| Grazia Borrello
et al | Italy | 2016 | GSE73182 | 19 | 9.716 | 0.171 | 5 | 9.952 | 0.173 | 14 | 0 | 5 | 5 | (23) |
| Rossing et
al | Denmark | 2014 | E-MTAB-736 | 12 | 8.048 | 0.593 | 10 | 8.914 | 0.579 | 8 | 0 | 4 | 10 | (24) |
| TCGA | USA | 2017 | TCGA | 514 | 8.606 | 1.304 | 59 | 11.058 | 1.179 | 409 | 4 | 105 | 55 | – |
Prediction of miR-486-5p target mRNAs
and GO analysis
From at least nine of the 12 miRNA target prediction
databases, 80 mRNA targets of miR-486-5p were identified. To
understand the biological function of miR-486-5p in PTC, GO
enrichment analysis of genes encoding for miR-486-5p target mRNAs
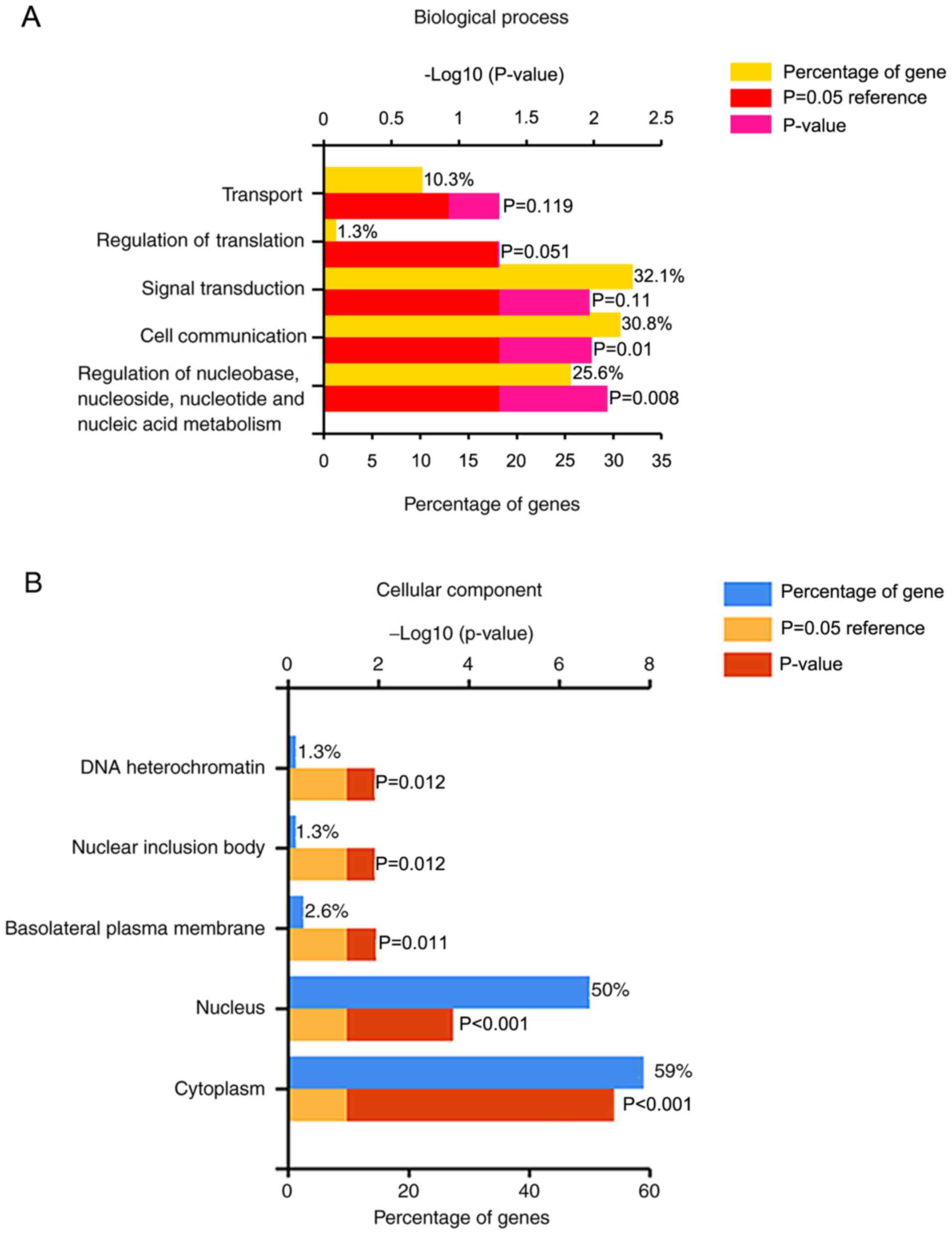
was performed in FunRich. ‘Regulation of nucleobase, nucleoside,
nucleotide and nucleic acid metabolism’ (P=0.008; Fig. 6A), ‘Cytoplasm’
(P=6.158×10−8; Fig. 6B)
and ‘RNA binding’ (P=0.004; Fig.
6C) were the top functions for biological process, cellular
component and molecular function, respectively.
Identification of miR-486-5p-regulated
pathways
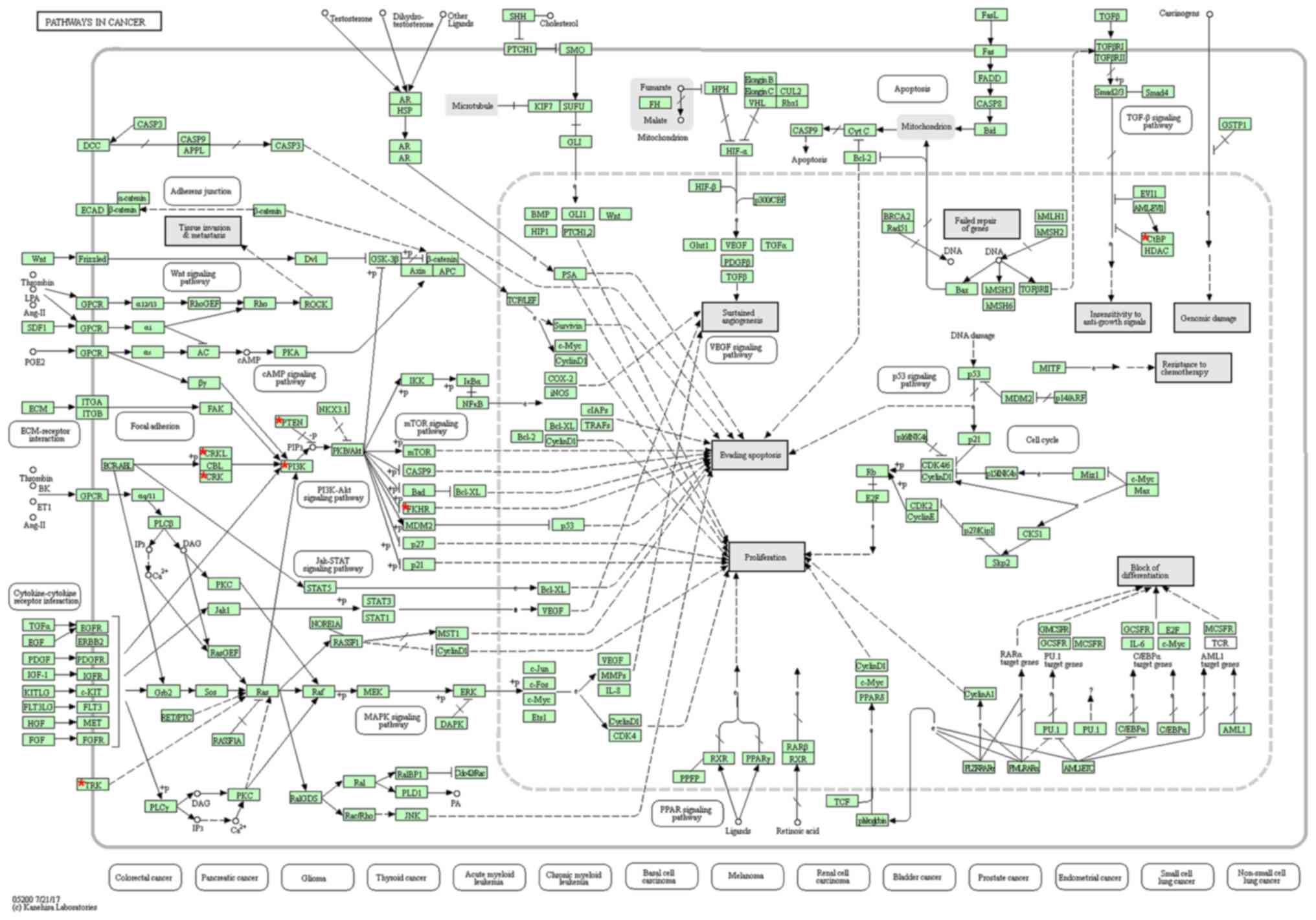
The KEGG pathway enrichment analysis was performed
on miR-486-5p-relevant genes using DAVID. The results revealed the
most significantly enriched pathway was ‘hsa05200: Pathways in
cancer’ with six genes including CRK like proto-oncogene (CRKL),
C-terminal binding protein 2, forkhead box O1, phosphatase and
tensin homolog (PTEN), phosphoinositide-3-kinase regulatory subunit
1 and tropomyosin 3 (TPM3) (P=0.010; Fig. 7). In addition, protein-protein
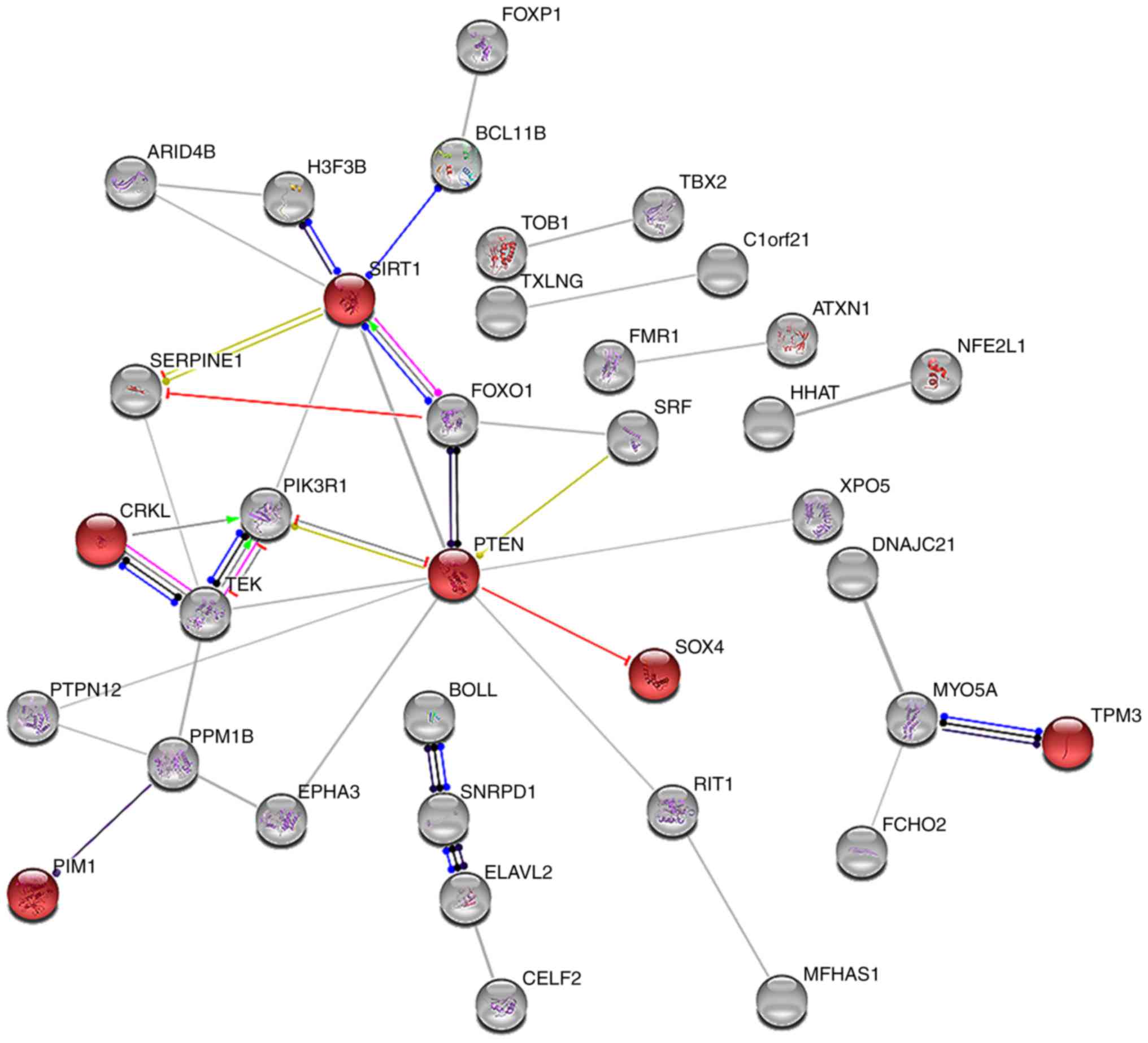
interaction network analysis was performed on the 80 relevant genes
using STRING: ‘hsa05206: MicroRNAs in cancer’ was identified, with
six genes including CRKL, pim-1 proto-oncogene, sirtuin 1, PTEN,
SRY-box 4 and TPM3 (Fig. 8). The
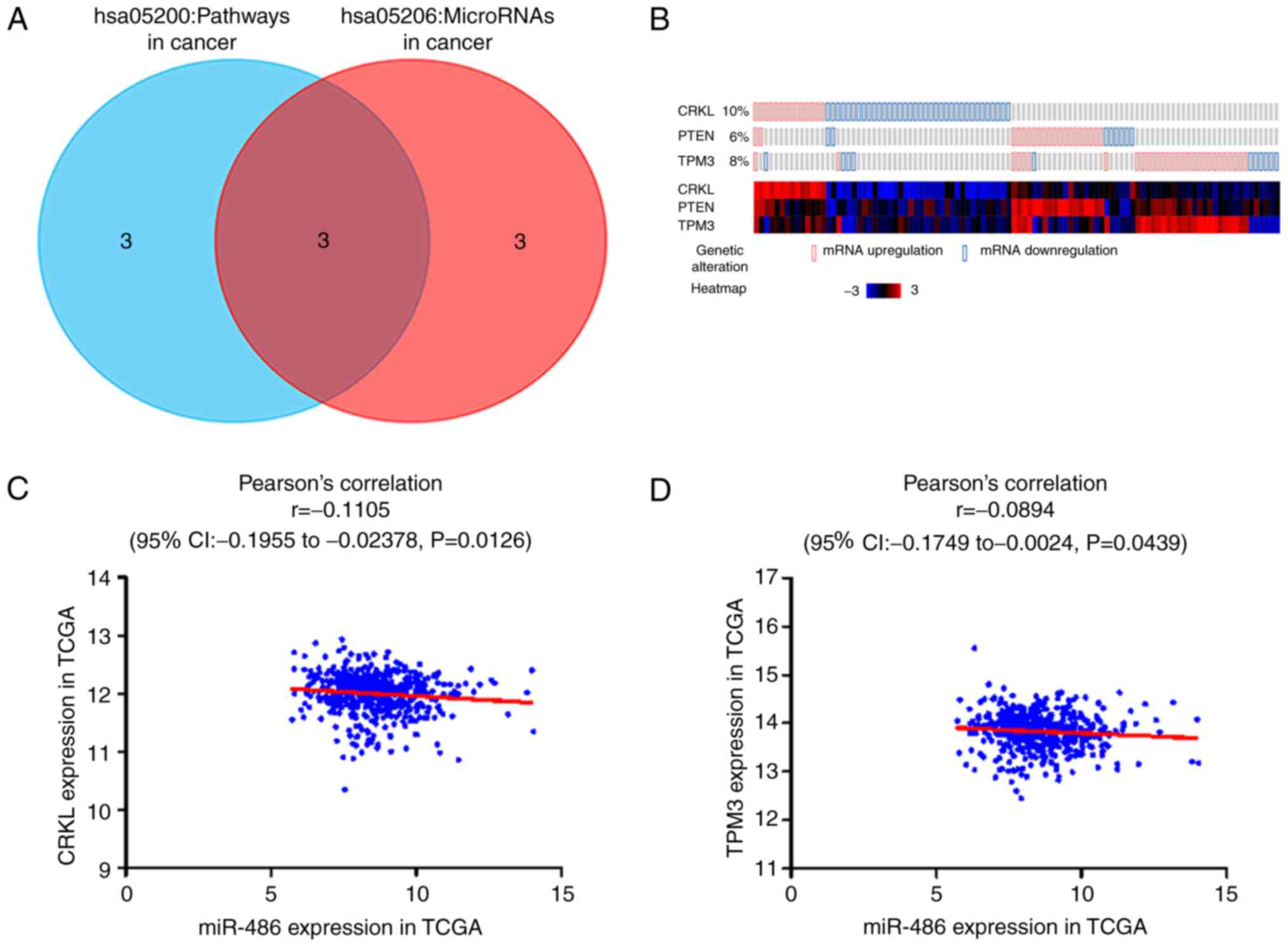
three common genes (CRKL, PTEN and TPM3) of the ‘hsa05200: Pathways
in cancer’ and ‘hsa05206: MicroRNAs in cancer’ pathways from the
analyses were considered hub genes (Fig. 9A).
Validation of hub genes
To validate the relationship between miR-486-5p and
the hub genes CRKL, PTEN and TPM3, their gene expression levels
were examined using the cBioPortal for Cancer Genomics. The
expression levels of the three hub genes (CRKL, PTEN and TPM3) in
PTC were markedly higher than in non-cancerous thyroid samples
(Fig. 9B). Pearson's correlation
analysis was performed to assess the relationship between
miR-486-5p and the hub genes. A statistically significant negative
correlation between miR-486-5p and CRKL (r=−0.111; P=0.013;
Fig. 9C) and TPM3 (r=−0.089;
P=0.044; Fig. 9D) was observed.
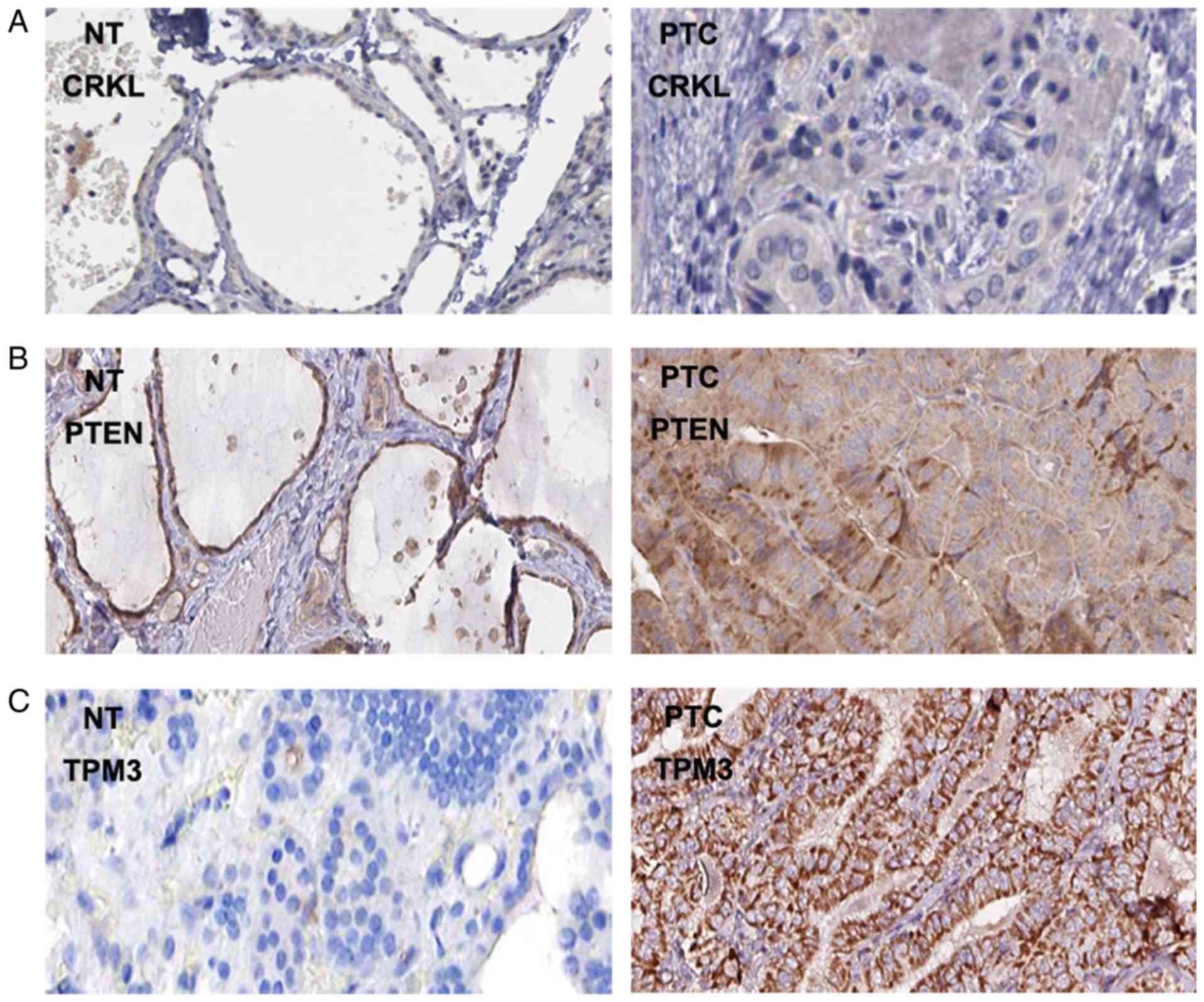
Finally, the protein expression levels of CRKL, PTEN and TPM3 in
PTC tissues were visualized using immunohistochemical staining
images from The Human Protein Atlas database. In line with the gene
expression data, protein expression of CRKL, PTEN and TPM3 was
increased in PTC compared with in normal thyroid tissue (Fig. 10).
Discussion
In the present study, significantly decreased
expression levels of miR-486-5p in PTC were demonstrated by
analysis of TCGA, GEO (GSE40807, GSE57780, GSE62054 and GSE73182)
and ArrayExpress (E-MTAB-736) data, which included 603 PTC and 125
normal thyroid tissue samples. Downregulation of miR-486-5p was
associated with cancer stage, pathological lymph node, metastasis,
neoplasm and disease recurrence, all of which are clinical
indicators of poor disease outcome. SROC curve analysis of
miR-486-5p data from TCGA, GEO (GSE40807, GSE57780, GSE62054 and
GSE73182) and ArrayExpress (E-MTAB-736) demonstrated that
expression levels could provide high diagnostic value for PTC with
an AUC of 0.84 (95% CI: 0.81–0.87). In addition, downregulation of
miR-486-5p in patients with PTC may lead to worse OS. The
biological functions of miR-486-5p in facilitating the onset and
progression of PTC were also explored. The potential miR-486-5p
target mRNAs were found to be enriched in diverse signaling
pathways, including ‘hsa05200: Pathways in cancer’ and ‘hsa05206:
MicroRNAs in cancer’. Furthermore, three hub genes, CRKL, PTEN and
TPM3, were identified, all of which were notably upregulated in PTC
samples based on analysis using the cBioPortal for Cancer Genomics.
Therefore, it may be hypothesized that downregulated miR-486-5p
serves as a clinical biomarker for PTC and exerts a potential
biological role in progression of the disease.
There is accumulating evidence suggesting that
aberrant miRNA expression levels contribute towards PTC development
and progression. Several studies have identified a crucial role for
miR-486-5p during onset and progression of various types of cancer;
for example, miR-486-5p is markedly downregulated in non-small cell
lung cancer, osteosarcoma, breast cancer, myxoid liposarcoma and
pancreatic ductal adenocarcinomas (43–47).
Conversely, miR-486-5p is upregulated in chronic myelocytic
leukemia and myeloid leukemia of Down syndrome (48,49).
Therefore, miR-486-5p may be differentially expressed depending on
cancer type. To the best of our knowledge, only one study has
explored miR-486-5p expression levels in PTC. Ma et al
(50) demonstrated that miR-486-5p
was predominantly downregulated in PTC tissues and cell lines,
whereas miR-486-5p upregulation resulted in suppression of
proliferation and growth by targeting FBN1. However, a limitation
to this study is that it only analyzed 20 paired PTC and normal
thyroid samples by reverse transcription-quantitative polymerase
chain reaction (50). In the
present study, integration of results from multiple studies via
bioinformatics analysis revealed that miR-486-5p expression levels
were decreased in PTC, and may have diagnostic and prognostic value
for patients with PTC. Further investigations in vivo and
in vitro are required to identify the underlying molecular
mechanism of miR-486-5p in PTC. Exploring miR-486-5p expression
levels in neoplastic tissues from patients with PTC will aid in
understanding its clinical value.
miR-486-5p is upregulated by miR-660-5p via the
miR-660-5p-mouse double minute 2 homolog-proto-oncogene-p53 axis in
lung cancer (51). In gastric
cancer, miR-486-5p acts as a tumor suppressor and its
downregulation facilitates tumor growth by increasing transcription
of the anti-apoptotic factor, olfactomedin 4 (52). Similarly, miR-486-5p enhances
hepatocellular carcinoma chemosensitivity to sorafenib and
regulates metastasis by targeting citron rho-interacting
serine/threonine kinase and claudin 10 (53). In vitro, Borjigin et
al (46) revealed that the FUS
RNA binding protein-DNA damage inducible transcript 3 fusion
protein suppressed miR-486-5p expression leading to induction of
plasminogen activator inhibitor-1 expression in human myxoid
liposarcoma. Overall, these results suggest that miR-486-5p may act
as a tumor suppressor in various types of cancer. However, most of
the aforementioned studies only investigated individual mRNA
targets of miR-486-5p, whereas the present study utilized pathway
enrichment and network analysis to identify possible targets of
miR-486-5p. Certain target genes were associated with ‘Regulation
of nucleobase, nucleoside, nucleotide and nucleic acid metabolism’,
‘Cytoplasm’ and ‘RNA binding’ in the GO functional enrichment
analysis. Notably, the three genes CRKL, PTEN and TPM3 were
identified in both ‘hsa05200: Pathways in cancer’ and ‘hsa05206:
MicroRNAs in cancer’ through KEGG pathway enrichment analysis;
therefore, these genes may be essential to the pathogenesis and
progression of PTC.
CRKL encodes for a substrate of the BCR-ABL
proto-oncogene fusion gene, and has a pivotal role in fibroblast
transformation and may be oncogenic (54). Yang et al (55) reported that CRKL is associated with
cervical lymph node metastasis and invasion of capsules in PTC,
which has clinical implications on treatment. PTEN acts as an
oncogene and is highly prevalent in various cancers (56–58).
Fang et al (59)
demonstrated that ribonucleotide reductase catalytic subunit M1
induces PTEN expression and decreases Akt serine/threonine kinase
phosphorylation in a ribonucleotide reductase activity-independent
manner in PTC. In addition, Zhao et al (60) demonstrated that PTEN mRNA levels
may serve as a biomarker for PTC onset and act as a molecular
diagnostic index. TPM3 encodes a member of the tropomyosin family
of actin-binding proteins and is frequently involved in genetic
rearrangements resulting in fusion with the neurotrophic tyrosine
kinase receptor type 1 (NTRK1) gene, which then acts as an oncogene
(61). Numerous studies that have
demonstrated the association of NTRK1 with genetic rearrangements
in PTC (62,63).
Notably, CRKL, PTEN and TPM3 were more highly
expressed in PTC, and the results suggested that these genes may be
regulated by miR-486-5p, whose expression is lower in PTC. It may
be hypothesized that CRKL, PTEN and TPM3 have an important role in
PTC and are potential molecular targets in the clinic.
Nevertheless, in silico target gene prediction algorithms
have limited specificity, and further in vitro and in
vivo investigations are required to confirm the function of
miR-486-5p in PTC.
In conclusion, the present study revealed that the
expression of miR-486-5p was significantly downregulated in PTC.
The clinical implications of miR-486-5p were also evaluated using
results from multiple datasets. Bioinformatics analysis was
performed to elucidate the molecular mechanisms of miR-486-5p in
facilitating the onset and progression of PTC. Three hub genes
CRKL, PTEN and TPM3, were identified, which may be potential
targets of miR-486-5p in PTC. The clinical importance of miR-486-5p
in PTC and its underlying molecular mechanisms require further
investigation; however, the results indicated that miR-486-5p may
be involved in the pathogenesis of PTC.
Acknowledgements
The authors thank TCGA, GEO, ArrayExpress and The
Human Protein Atlas for providing the data.
Funding
The present study was partly supported by the
Guangxi Scientific Research and Technology Development Plan (grant
no. 1598011-4), the Fund of Guangxi Key R&D Project Plan (grant
no. AB17195020) and the Fund of National Natural Science Foundation
of China (grant nos. NSFC81060202 and NSFC81260222).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
D-YW, HY, GC and YH designed the study and performed
the experiments. D-HP, PL, Q-YM, Y-HL, J-QC, GC and Y-PW
participated in data processing and statistical analysis. D-YW and
Y-HL wrote the manuscript. D-YW, Y-HL, YH and HY critically revised
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PTC
|
papillary thyroid carcinoma
|
|
TCGA
|
the cancer genome atlas
|
|
GEO
|
gene expression omnibus
|
|
ROC
|
receiver operating characteristic
|
|
SROC
|
summary receiver operating
characteristic
|
|
OS
|
overall survival
|
|
GO
|
gene ontology
|
|
KEGG
|
Kyoto encyclopedia of genes and
genomes
|
|
PPI
|
protein-protein interaction
|
References
|
1
|
Kohlhapp FJ, Mitra AK, Lengyel E and Peter
ME: MicroRNAs as mediators and communicators between cancer cells
and the tumor microenvironment. Oncogene. 34:5857–5868. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vigneri R, Malandrino P and Vigneri P: The
changing epidemiology of thyroid cancer: Why is incidence
increasing? Curr Opin Oncol. 27:1–7. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Qiu Z, Li H, Wang J and Sun C: miR-146a
and miR-146b in the diagnosis and prognosis of papillary thyroid
carcinoma. Oncol Rep. 38:2735–2740. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Penha RCC, Sepe R, De Martino M, Esposito
F, Pellecchia S, Raia M, Del Vecchio L, Decaussin-Petrucci M, De
Vita G, Pinto LFR and Fusco A: Role of Dicer1 in thyroid cell
proliferation and differentiation. Cell Cycle. 16:2282–2289. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhuang J, Ye Y, Wang G, Ni J, He S, Hu C,
Xia W and Lv Z: MicroRNA-497 inhibits cellular proliferation,
migration and invasion of papillary thyroid cancer by directly
targeting AKT3. Mol Med Rep. 16:5815–5822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mahmoudian-Sani MR, Mehri-Ghahfarrokhi A,
Asadi-Samani M and Mobini GR: Serum miRNAs as biomarkers for the
diagnosis and prognosis of thyroid cancer: A comprehensive review
of the literature. Eur Thyroid J. 6:171–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Boufraqech M, Klubo-Gwiezdzinska J and
Kebebew E: MicroRNAs in the thyroid. Best Pract Res Clin Endocrinol
Metab. 30:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bai D, Sun H, Wang X, Lou H, Zhang J, Wang
X and Jiang L: MiR-150 inhibits cell growth in vitro and in vivo by
restraining the RAB11A/WNT/β-catenin pathway in thyroid cancer. Med
Sci Monit. 23:4885–4894. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang J, Yang H, Si Y, Hu D, Yu Y, Zhang Y,
Gao M and Zhang H: Iodine promotes tumorigenesis of thyroid cancer
by suppressing Mir-422a and Up-regulating MAPK1. Cell Physiol
Biochem. 43:1325–1336. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Paiva MM, Kimura ET and Coltri PP: miR18a
and miR19a recruit specific proteins for splicing in thyroid cancer
cells. Cancer Genomics Proteomics. 14:373–381. 2017.PubMed/NCBI
|
|
11
|
Small EM, O'Rourke JR, Moresi V,
Sutherland LB, McAnally J, Gerard RD, Richardson JA and Olson EN:
Regulation of PI3-kinase/Akt signaling by muscle-enriched
microRNA-486. Proc Natl Acad Sci USA. 107:4218–4223. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ren C, Chen H, Han C, Fu D, Zhou L, Jin G,
Wang F, Wang D, Chen Y, Ma L, et al: miR-486-5p expression pattern
in esophageal squamous cell carcinoma, gastric cancer and its
prognostic value. Oncotarget. 7:15840–15853. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu J, Zeng Y, Xu C, Qin H, Lei Z, Shen D,
Liu Z and Huang JA: Expression profile analysis of microRNAs and
downregulated miR-486-5p and miR-30a-5p in non-small cell lung
cancer. Oncol Rep. 34:1779–1786. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu C, Li M, Hu Y, Shi N, Yu H, Liu H and
Lian H: miR-486-5p attenuates tumor growth and lymphangiogenesis by
targeting neuropilin-2 in colorectal carcinoma. Onco Targets Ther.
9:2865–2871. 2016.PubMed/NCBI
|
|
15
|
Cao Z, Liu C, Xu J, You L, Wang C, Lou W,
Sun B, Miao Y, Liu X, Wang X, et al: Plasma microRNA panels to
diagnose pancreatic cancer: Results from a multicenter study.
Oncotarget. 7:41575–41583. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li CY, Liang GY, Yao WZ, Sui J, Shen X,
Zhang YQ, Peng H, Hong WW, Ye YC, Zhang ZY, et al: Identification
and functional characterization of microRNAs reveal a potential
role in gastric cancer progression. Clin Transl Oncol. 19:162–172.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma X, Wei J, Zhang L, Deng D, Liu L, Mei
X, He X and Tian J: miR-486-5p inhibits cell growth of papillary
thyroid carcinoma by targeting fibrillin-1. Biomed Pharmacother.
80:220–226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hutter C and Zenklusen JC: The cancer
genome atlas: Creating lasting value beyond its data. Cell.
173:283–285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:(Database Issue).
D991–D995. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kolesnikov N, Hastings E, Keays M,
Melnichuk O, Tang YA, Williams E, Dylag M, Kurbatova N, Brandizi M,
Burdett T, et al: ArrayExpress update-simplifying data submissions.
Nucleic Acids Res. 43:(Database Issue). D1113–D1116. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chandran UR, Medvedeva OP, Barmada MM,
Blood PD, Chakka A, Luthra S, Ferreira A, Wong KF, Lee AV, Zhang Z,
et al: TCGA expedition: A data acquisition and management system
for TCGA data. PLos One. 11:e01653952016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lassalle S, Zangari J, Popa A, Ilie M,
Hofman V, Long E, Patey M, Tissier F, Belléannée G, Trouette H, et
al: MicroRNA-375/SEC23A as biomarkers of the in vitro efficacy of
vandetanib. Oncotarget. 7:30461–30478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Minna E, Romeo P, Dugo M, De Cecco L,
Todoerti K, Pilotti S, Perrone F, Seregni E, Agnelli L, Neri A, et
al: miR-451a is underexpressed and targets AKT/mTOR pathway in
papillary thyroid carcinoma. Oncotarget. 7:12731–12747. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rossing M, Borup R, Henao R, Winther O,
Vikesaa J, Niazi O, Godballe C, Krogdahl A, Glud M, Hjort-Sørensen
C, et al: Down-regulation of microRNAs controlling tumourigenic
factors in follicular thyroid carcinoma. J Mol Endocrinol.
48:11–23. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lau J, Ioannidis JP and Schmid CH:
Quantitative synthesis in systematic reviews. Ann Intern Med.
127:820–826. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maragkakis M, Vergoulis T, Alexiou P,
Reczko M, Plomaritou K, Gousis M, Kourtis K, Koziris N, Dalamagas T
and Hatzigeorgiou AG: DIANA-microT Web server upgrade supports Fly
and Worm miRNA target prediction and bibliographic miRNA to disease
association. Nucleic Acids Res. 39:W145–W148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Paraskevopoulou MD, Georgakilas G,
Kostoulas N, Vlachos IS, Vergoulis T, Reczko M, Filippidis C,
Dalamagas T and Hatzigeorgiou AG: DIANA-microT web server v5.0:
Service integration into miRNA functional analysis workflows.
Nucleic Acids Res. 41:W169–W173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36:(Database Issue). D149–D153. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsang JS, Ebert MS and van Oudenaarden A:
Genome-wide dissection of microRNA functions and cotargeting
networks using gene set signatures. Mol Cell. 38:140–153. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wong N and Wang X: miRDB: An online
resource for microRNA target prediction and functional annotations.
Nucleic Acids Res. 43:(Database Issue). D146–D152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vejnar CE, Blum M and Zdobnov EM: miRmap
web: Comprehensive microRNA target prediction online. Nucleic Acids
Res. 41:W165–W168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hsu SD, Chu CH, Tsou AP, Chen SJ, Chen HC,
Hsu PW, Wong YH, Chen YH, Chen GH and Huang HD: miRNAMap 2.0:
Genomic maps of microRNAs in metazoan genomes. Nucleic Acids Res.
36:(Database Issue). D165–D169. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Blin K, Dieterich C, Wurmus R, Rajewsky N,
Landthaler M and Akalin A: DoRiNA 2.0-upgrading the doRiNA database
of RNA interactions in post-transcriptional regulation. Nucleic
Acids Res. 43:(Database Issue). D160–D167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weingarten-Gabbay S, Elias-Kirma S, Nir R,
Gritsenko AA, Stern-Ginossar N, Yakhini Z, Weinberger A and Segal
E: Comparative genetics. Systematic discovery of cap-independent
translation sequences in human and viral genomes. Science. 351:pii:
aad4939. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Miranda KC, Huynh T, Tay Y, Ang YS, Tam
WL, Thomson AM, Lim B and Rigoutsos I: A pattern-based method for
the identification of MicroRNA binding sites and their
corresponding heteroduplexes. Cell. 126:1203–1217. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rehmsmeier M, Steffen P, Hochsmann M and
Giegerich R: Fast and effective prediction of microRNA/target
duplexes. RNA. 10:1507–1517. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cheng Y, Chen L, Cao X, Ha S and Xie X:
Expression profiling and functional analysis of hsa-miR-125b and
its target genes in drug-resistant cell line of human gastric
cancer. Yi Chuan. 36:119–126. 2014.(In Chinese). View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pathan M, Keerthikumar S, Ang CS, Gangoda
L, Quek CY, Williamson NA, Mouradov D, Sieber OM, Simpson RJ, Salim
A, et al: FunRich: An open access standalone functional enrichment
and interaction network analysis tool. Proteomics. 15:2597–2601.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
da Huang W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Szklarczyk D, Morris JH, Cook H, Kuhn M,
Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, et al:
The STRING database in 2017: Quality-controlled protein-protein
association networks, made broadly accessible. Nucleic Acids Res.
45:D362–D368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Uhlen M, Fagerberg L, Hallstrom BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li W, Wang Y, Zhang Q, Tang L, Liu X, Dai
Y, Xiao L, Huang S, Chen L, Guo Z, et al: MicroRNA-486 as a
biomarker for early diagnosis and recurrence of non-small cell lung
cancer. PLos One. 10:e01342202015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
He M, Wang G, Jiang L, Qiu C, Li B, Wang J
and Fu Y: miR-486 suppresses the development of osteosarcoma by
regulating PKC-δ pathway. Int J Oncol. 50:1590–1600. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang G, Liu Z, Cui G, Wang X and Yang Z:
MicroRNA-486-5p targeting PIM-1 suppresses cell proliferation in
breast cancer cells. Tumour Biol. 35:11137–11145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Borjigin N, Ohno S, Wu W, Tanaka M, Suzuki
R, Fujita K, Takanashi M, Oikawa K, Goto T, Motoi T, et al:
TLS-CHOP represses miR-486 expression, inducing upregulation of a
metastasis regulator PAI-1 in human myxoid liposarcoma. Biochem
Biophys Res Commun. 427:355–360. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mees ST, Mardin WA, Sielker S, Willscher
E, Senninger N, Schleicher C, Colombo-Benkmann M and Haier J:
Involvement of CD40 targeting miR-224 and miR-486 on the
progression of pancreatic ductal adenocarcinomas. Ann Surg Oncol.
16:2339–2350. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang LS, Li L, Li L, Chu S, Shiang KD, Li
M, Sun HY, Xu J, Xiao FJ, Sun G, et al: MicroRNA-486 regulates
normal erythropoiesis and enhances growth and modulates drug
response in CML progenitors. Blood. 125:1302–1313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shaham L, Vendramini E, Ge Y, Goren Y,
Birger Y, Tijssen MR, McNulty M, Geron I, Schwartzman O, Goldberg
L, et al: MicroRNA-486-5p is an erythroid oncomiR of the myeloid
leukemias of Down syndrome. Blood. 125:1292–1301. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ma X, Wei J, Zhang L, Deng D, Liu L, Mei
X, He X and Tian J: miR-486-5p inhibits cell growth of papillary
thyroid carcinoma by targeting fibrillin-1. Biomed Pharmacother.
80:220–226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Borzi C, Calzolari L, Centonze G, Milione
M, Sozzi G and Fortunato O: mir-660-p53-mir-486 Network: A new key
regulatory pathway in lung tumorigenesis. Int J Mol Sci. 18:pii:
E222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Oh HK, Tan AL, Das K, Ooi CH, Deng NT, Tan
IB, Beillard E, Lee J, Ramnarayanan K, Rha SY, et al: Genomic loss
of miR-486 regulates tumor progression and the OLFM4 antiapoptotic
factor in gastric cancer. Clin Cancer Res. 17:2657–2667. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sun H, Cui C, Xiao F, Wang H, Xu J, Shi X,
Yang Y, Zhang Q, Zheng X, Yang X, et al: miR-486 regulates
metastasis and chemosensitivity in hepatocellular carcinoma by
targeting CLDN10 and CITRON. Hepatol Res. 45:1312–1322. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Frietsch JJ, Kastner C, Grunewald TG,
Schweigel H, Nollau P, Ziermann J, Clement JH, La Rosée P, Hochhaus
A and Butt E: LASP1 is a novel BCR-ABL substrate and a
phosphorylation-dependent binding partner of CRKL in chronic
myeloid leukemia. Oncotarget. 5:5257–5271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yang X, Lv W, Shi R, Cheng S, Zhang J and
Xu Z: The clinical implications of Crk-like adaptor protein
expression in papillary thyroid microcarcinoma. Tumour Biol.
35:12435–12440. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang J, Chen D, Liang S, Wang J, Liu C,
Nie C, Shan Z, Wang L, Fan Q and Wang F: miR-106b promotes cell
invasion and metastasis via PTEN mediated EMT in ESCC. Oncol Lett.
15:4619–4626. 2018.PubMed/NCBI
|
|
57
|
Gao ZJ, Yuan WD, Yuan JQ, Yuan K and Wang
Y: miR-486-5p functions as an oncogene by targeting PTEN in
non-small cell lung cancer. Pathol Res Pract. 214:700–705. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wu S, Wang J and Li F: Dysregulation of
PTEN caused by the underexpression of microRNA130b is associated
with the severity of lupus nephritis. Mol Med Rep. 17:7966–7972.
2018.PubMed/NCBI
|
|
59
|
Fang Z, Song R, Gong C, Zhang X, Ren G, Li
J, Chen Y, Qiu L, Mei L, Zhang R, et al: Ribonucleotide reductase
large subunit M1 plays a different role in the invasion and
metastasis of papillary thyroid carcinoma and undifferentiated
thyroid carcinoma. Tumour Biol. 37:3515–3526. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhao Y, Liu X, Zhong L, He M, Chen S, Wang
T and Ma S: The combined use of miRNAs and mRNAs as biomarkers for
the diagnosis of papillary thyroid carcinoma. Int J Mol Med.
36:1097–1103. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Geeves MA, Hitchcock-DeGregori SE and
Gunning PW: A systematic nomenclature for mammalian tropomyosin
isoforms. J Muscle Res Cell Motil. 36:147–153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bastos AU, de Jesus AC and Cerutti JM:
ETV6-NTRK3 and STRN-ALK kinases fusions are recurrent events in
papillary thyroid cancer of adult population. Eur J Endocrinol.
178:85–93. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Iyama K, Matsuse M, Mitsutake N,
Rogounovitch T, Saenko V, Suzuki K, Ashizawa M, Ookouchi C, Suzuki
S, Mizunuma H, et al: Identification of three novel fusion
oncogenes, SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET, in thyroid
cancers of young patients in fukushima. Thyroid. 27:811–818. 2017.
View Article : Google Scholar : PubMed/NCBI
|