Introduction
Atherosclerosis (AS) is a common complication of
diabetes. Arteriosclerosis in diabetic patients has higher
mortality than that among the non-diabetic (1,2).
Hyperglycemia is a primary phenotype of diabetic patients (3). Vascular endothelial cells are
critical targets of hyperglycemia injury in diabetes mellitus
(4). The injury of vascular
endothelial cells is the initial event of AS (5). Moreover, oxidative stress caused by
hyperglycemia is a main inducement of vascular endothelial cells
injury (6), which is resulted from
the imbalance between production and removal of reactive oxygen
species (ROS) (7). In normal
circumstances, intracellular antioxidant systems, for instance,
superoxide dismutase (SOD) and catalase (CAT), maintain the cell
redox self-steady state (8).
Nevertheless, ROS accumulation will induce lipid peroxidation,
resulting in the production of MDA (9,10).
Moreover, excessive exposure of ROS can lead to the dysfunction of
the vascular endothelial cells (11). Vascular endothelium, a complex
endocrine organ (12), could
secrete cytoactive factors in response to the change of signals and
stress (13). Nitric oxide (NO)
plays a critical role in maintaining the vasoconstrictor function,
which is a primary endogenous vascular diastolic factor (14). Additionally, NO is helpful to the
inhibition of the production of oxygen free radicals (15). It has been documented that
oxidative stress and the production or activation of NO were
essential for the AS (8).
Furthermore, studies have pointed out that cell
apoptosis was closely linked to the ROS accumulation (3,16,17).
Importantly, high glucose can promote apoptosis of human umbilical
vein endothelial cells (HUVECs) (18). Hence, alleviation of high
glucose-induced oxidative stress and apoptosis of endothelial cells
may play an important role in the prevention and treatment of AS
and other diabetic macrovascular diseases. The PI3K/AKT signaling
functions critically in cellular death progression (19). It is well known that the survival
of endothelial cells could be maintained by AKT. Moreover, AKT
could activate the expression of endothelial nitric oxide synthase
(eNOS) (20), which is responsible
for the production of NO (21).
Therefore, in this study, we paid close attention to the AKT/eNOS
signaling for its protective role in endothelial cells.
As previously described (22), oleanolic acid (OA), which is a
pentacyclic triterpenoid compound, is widespread in plants. OA has
several pharmacological activities. The antitumor effect of OA has
been noted previously (23–26).
Studies have also reported that OA could exert anti-oxidative
effect (27,28). OA is believed to be able to treat
diabetes (29,30), and it is reported to be related to
arteriosclerosis (31). Thus, it
is of research significance to assess the effects of OA on
high-glucose induced oxidative stress in HUVECs.
In the present study, we sought to evaluate the
effects of OA in HUVECs cultivated in vitro via detecting
the oxidative response and apoptosis of HUVECs. The mechanism
action of OA was as well investigated. Our research provides
reference for developing candidate agent in the treatment of AS in
diabetics.
Materials and methods
Cell culture and treatment
HUVECs (ATCC, USA) were maintained in Dulbecco's
modified Eagle's medium (DMEM) (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) that contained 10% FBS (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and
penicillin/streptomycin (Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China) at 37°C in an incubator with 5%
CO2. The normal blood glucose level ranged from 3.9 and
5.5 mM (32) in non-diabetics.
Blood sugar levels increased more than normal range may be an
indicator of diabetes. According to some researches on
hyperglycemia injury model in vitro (33,34),
HUVECs were respectively treated with glucose at 5 and 33 mM for
the control and injury model for 48 h. The incubation concentration
of OA was determined in reference to a previous study (35). The research groups in this study
were classified as follow: Blank group (blank): no glucose
treatment; control group (Con): 5 mM glucose treatment; High
glucose model group (GC): 33 mM glucose (Sangon Biotech Co., Ltd.,
Shanghai, China) treatment; positive control group (OA-H/Con): 40
mM OA (purity >98%; Beijing Solarbio Science & Technology
Co., Ltd.) (dissolved in ethanol) for incubation for 24 h prior to
5 mM glucose treatment; low OA treatment group (OA-L/GC): Cells
were treated with 20 mM OA at 37°C for 24 h prior to 33 mM glucose
treatment; high OA treatment group (OA-H/GC): Cells were treated
with 40 mM OA at 37°C for 24 h prior to 33 mM glucose
treatment.
Cell viability assay
The cell survival rate was examined using Cell
Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology, Haimen,
China) assay, according to the manufacturer's protocols. To explain
further, 1×104/each well were seeded into a 96-well
plate and incubated at 37°C for 24 h. CCK-8 (10 µl/well) was added
into the 96-well plate, and then the cells were incubated at 37°C
for 4 h. Absorbance at 450 nm was detected using a microplate
reader (PerkinElmer, Inc., Waltham, MA, USA).
ROS measurement
Intracellular ROS level was detected using
fluorescent probe DCFH-DA probe (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Cells were collected and washed by PBS. Next,
the cells were maintained with 10 µM DCHF-DA in darkness at 37°C
for 30 min. Flow cytometry analysis was carried out to examine the
fluorescence signals corresponding to DCHF-DA on flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA) with Cell Quest software
version 5.1 (BD Biosciences). At least 10,000 events were analyzed
in each examination.
Detection for mitochondrial
transmembrane potential (MMP)
The Rho 123 accumulation was determined by flow
cytometry analysis as previously described (36). Following the treatment above, the
cells in 24-well plates (2×105 cells/well) were
incubated with 50 mM Rho 123 (Sigma-Aldrich; Merck KGaA) at 37°C
for 30 min. The reaction was terminated at 5 min of incubation on
ice. Subsequently, the fluorescent of Rho 123 was detected by FACS
Calibur flow cytometer (BD Biosciences). Cell Quest software
version 5.1 (BD Biosciences) was used to perform the analysis.
Detection of apoptosis
Annexin V-FITC (Sigma-Aldrich; Merck KGaA)/PI
(Sigma-Aldrich; Merck KGaA) double-staining for apoptosis analysis
was used to determine the aopotosis by flow cytometry analysis. To
be more specific, the cells in 6-well plate at a density of
2×106/well were treated as above. Subsequently, the
cells were incubated with 5 ul Annexin V-FITC for10 min at room
temperature and with 5 µl PI at room temperature for 15 min in the
darkness. FACS Calibur flow cytometer (BD Biosciences) and Cell
Quest software version 5.1 were used to count the apoptotic
cells.
Measurement of NO level
The generation of NO was examined with commercial
Nitrate/Nitrite Fluorometric Assay kit (Cayman Chemical Company,
Ann Arbor, MI, USA) through nitrate reductase method via its
breakdown products, Nitrate/Nitrite, After conducting treatment as
designed, the determination was performed following manufacturer's
protocols.
Measurement of SOD, CAT, and MDA
level
Cells (1×104/well) were seeded into a
96-well plate and incubated at 37°C for 24 h. Then, the cells were
centrifuged at 200 g for 5 min at 4°C. After being washed with PBS,
the cells were resuspended and collected for the detection of the
enzymes activities. The cellular SOD and CAT enzyme were determined
using corresponding kits (Beijing Solarbio Science & Technology
Co., Ltd.), according to the protocols. The MDA content was
detected with thiobarbituric acid substance provided by
Malondialdehyde Assay Kit (Beyotime, China).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was exacted using RNAiso Plus (Invitrogen;
Thermo Fisher Scientific, Inc.). M-MLV (Promega Corporation,
Madison, WI, USA) and oligo (dT) primers (Takara Bio, Inc., Otsu,
Japan) were used to synthesis cDNA from RNA (1 µg). The temperature
and incubation protocol was set at 25°C for 10 min, at 42°C for 50
min, at 70°C for 10 min. Amplification of the related genes was
conducted using LightCycler® 480 SYBR-Green I Master
(Roche Diagnostics, Indianapolis, IN, USA). The thermal cycle
conditions was as follows: At 95°C, 4 min; at 35 cycles of 95°C, 15
Sec; at 60°C, 45 Sec; at 72°C, 7 min. The expression of target
genes was normalized using the expression levels of GAPDH according
to 2−ΔΔCq method (37).
The primers used were as follows: Fas forward
5′-GTGCTTTGCTTAGGGTTCCC-3′, Fas reverse 5′-AACTTGCACTTCTGGCCATG-3′;
Fasl forward 5′-GTCCAACTCAAGGTCCATGC-3′, Fasl reverse
5′-TTGTTGCAAGATTGACCCCG-3′; Bax forward 5′-GTGCCGGAACTGATCAGAAC-3′,
Bax reverse 5′-CCAAAGTAGGAGAGGAGGCC-3′; Bcl-2 forward
5′-GCCTTCTTTGAGTTCGGTGG-3′, Bcl-2 reverse
5′-GAAATCAAACAGAGGCCGCA-3′; GAPDH forward:
5′-CACAGTCCATGCCATCACTG-3′; GAPDH reverse:
5′-ATCTCGCTCCTGGAAGATGG-3′;
Western blot analysis
Cells were lysed using 200 µl lysis buffer NP40
(Beyotime Institute of Biotechnology). The concentration of the
samples was determined by BCA protein quantitative kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Proteins were separated
using 10% SDS-PAGE gels and transferred onto a PVDF membrane (EMD
Millipore, Billerica, MA, USA). After being blocked with 5%
non-skimmed milk at room temperature for 2 h, the membrane was
incubated with primary antibodies as listed: cleaved caspase-3
(ab32042, 1:500; Abcam, Cambridge, UK), Fas (ab82419, 1:1,000),
Fasl (ab15285, 1 ug/ml), Bax (ab32503,1:2,000), Bcl-2 (ab32124,
1:1,000), p-AKT (ab812831:5,000), AKT 1/2 (ab182729, 1:5,000),
p-eNOS (ab184154, 1:1,000), eNOS (ab76198, 1:1,000), and GAPDH
(ab8245, 1:5,000), at 4°C overnight. Then, horseradish
peroxidase-conjugated secondary antibody (ab6721, 1:4,000) was
added and maintained at room temperature for 1 h. The blot bands
were developed using BeyoECL Star (Beyotime Institute of
Biotechnology). The gray density was calculated with Quantity One
software version 4.6 (Bio-Rad Laboratories, Inc.).
Statistical analysis
Statistical analysis was calculated using GraphPad
Prism software 6.0 (GraphPad Software, Inc., La Jolla, CA, USA).
One-way ANOVA following Tukey's post hoc test was used to compare
the difference between groups. The results were shown as means ±
standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
OA protected against
high-glucose-induced injury in HUVECs
We first determined the effect of OA on cell
viability of HUVECs. The CCK-8 test showed that the cell survival
rate of HUVECs decreased obviously in the GC group in comparison to
that in the control group, and that the pretreatment of OA improved
the cell survival rate in a dose-dependent manner (Fig. 1A). Moreover, the nitric oxide (NO)
generation was rescued by the pretreatment of OA (Fig. 1B). We also found that as an
indicator of lipid peroxidation, the production of MDA caused by
high glucose treatment was inhibited by the pretreatment of OA
(Fig. 1C). Similarly, the
activities of SOD and CAT were recovered in the pretreatment groups
(Fig. 1D and E).
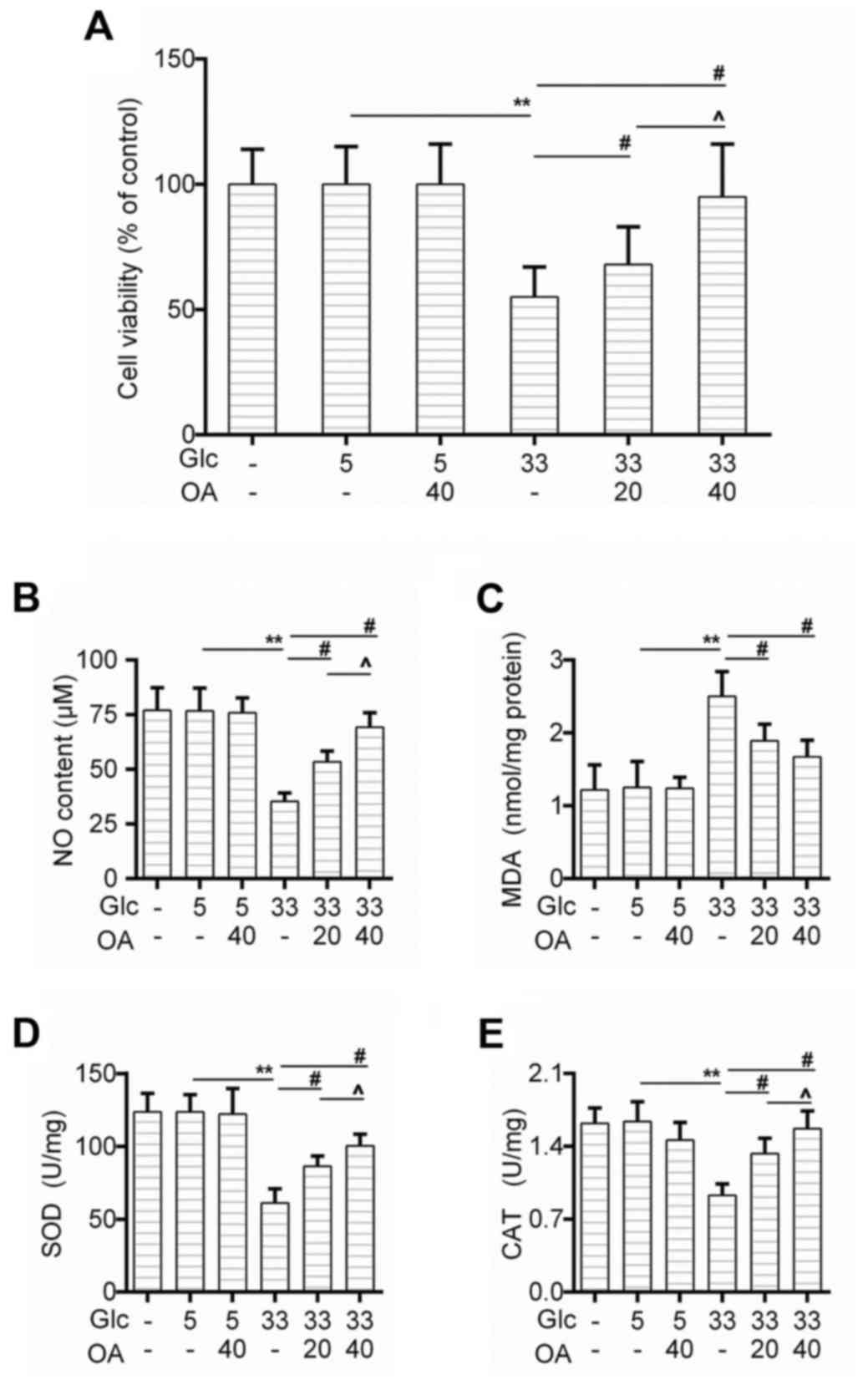 | Figure 1.OA protected against
high-glucose-induced injury in HUVECs. (A) Cell viability of HUVECs
was detected by CCK-8 assay when the cells were treated with high
glucose and/or OA. (B-E) Detection of (B) NO content, (C) MDA
level, (D) SOD activity and (E) CAT by available commercial kit
when the cells were treated with high glucose and/or OA. Blank
group (Blank), cuntreated cells; control group (5 mM)
(Con/Control), cells were treated with 5 mM glucose; high glucose
(33 mM) model group (GC), cells were treated with 33 mM glucose;
positive control group (OA-H/Con), cells were treated with 5 mM
glucose and 40 µM OA; low OA (20 µM) treatment group (OA-L/GC),
cells were treated with 33 mM glucose and 20 µM OA; high OA (40 µM)
treatment group (OA-H/GC), cells were treated with 33 mM glucose
and 40 µM OA. **P<0.01, #P<0.01 and
^P<0.05 as indicated. OA, oleanolic acid; HUVECs;
human umbilical vein endothelial cells; SOD, superoxide dismutase;
CAT, catalase. |
OA depressed the ROS production and
mitochondrial membrane potential (MMP) loss in HUVECs
The increased ROS level is one of the sources
causing mitochondria damage (38).
The loss of MMP is an early hallmark event of mitochondria damage
leading to apoptosis (39). Thus,
the ROS production and the MMP of HUVECs were measured by flow
cytometry analysis. Results showed that the ROS production was
triggered by high glucose treatment. Noticeably, the ROS level was
lower in the OA pretreatment groups than that in the GC group
(Fig. 2A and B). Meanwhile, the
loss of MMP caused by high glucose was recovered by the
pretreatment of OA (Fig. 2C and
D).
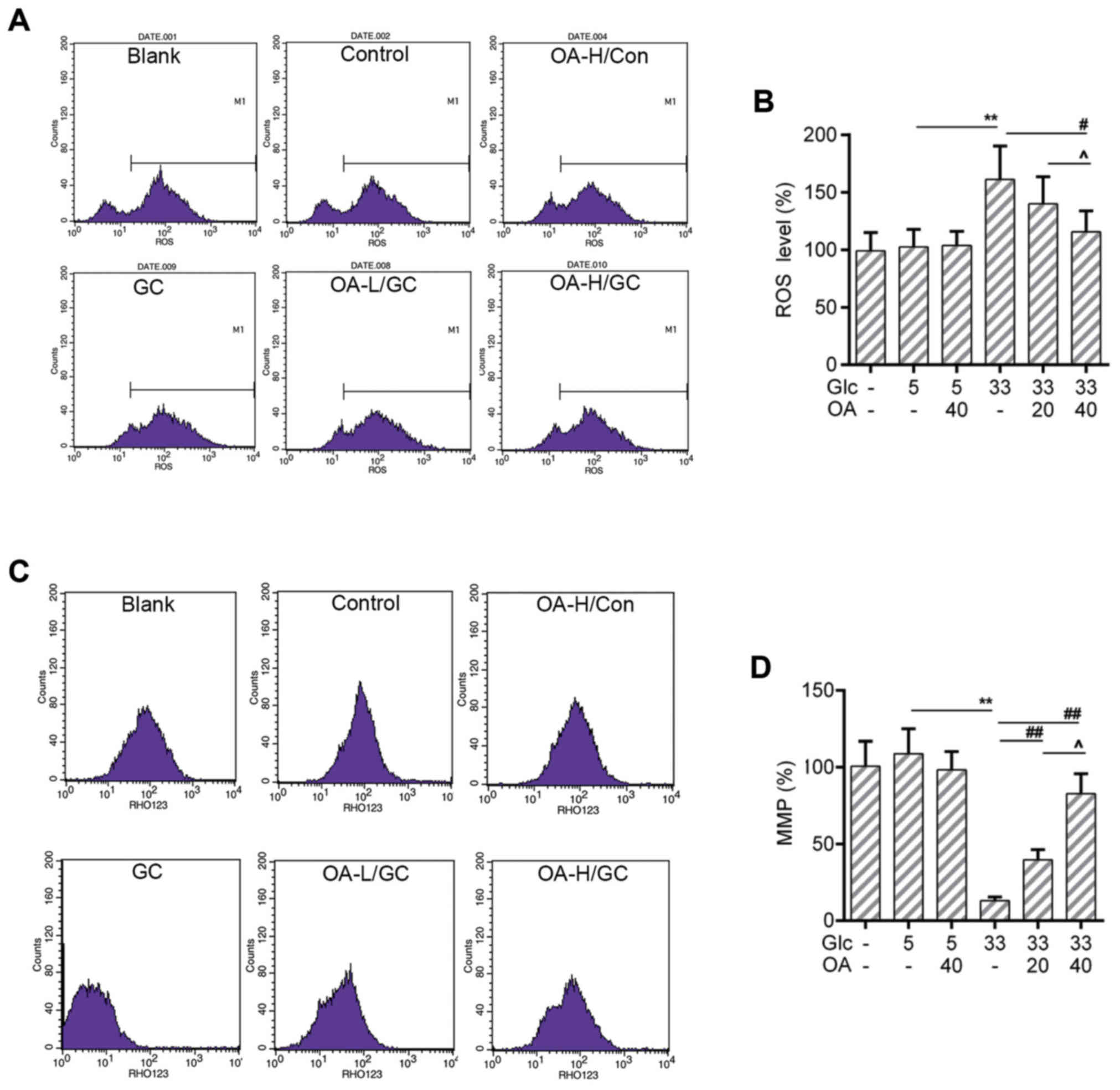 | Figure 2.OA prevented oxidative stress in
HUVECs. (A and B) Flow cytometry assay for detection of cellular
ROS analysis using DCFH-DA dye. (C and D) Flow cytometry analysis
for detection of MMP using Rho123. Blank group (Blank), cuntreated
cells; control group (5 mM) (Con/Control), cells were treated with
5 mM glucose; high glucose (33 mM) model group (GC), cells were
treated with 33 mM glucose; positive control group (OA-H/Con),
cells were treated with 5 mM glucose and 40 µM OA; low OA (20 µM)
treatment group (OA-L/GC), cells were treated with 33 mM glucose
and 20 µM OA; high OA (40 µM) treatment group (OA-H/GC), cells were
treated with 33 mM glucose and 40 µM OA. **P<0.01,
#P<0.05, ##P<0.01 and
^P<0.05 as indicated. OA, oleanolic acid; HUVECs;
human umbilical vein endothelial cells; ROS, reactive oxygen
species; MMP, mitochondrial transmembrane potential. |
OA inhibited apoptosis caused by
oxidative stress in HUVECs
ROS accumulation and the loss of MMP were both
considered to be closely associated with apoptosis (40). Therefore, we further detected the
apoptosis of HUVECs using flow cytometry analysis. The results
indicated that the apoptosis of HUVECs induced by high glucose was
apparently inhibited in the OA pretreatment groups (Fig. 3A and B). Moreover, the RT-qPCR and
western blot assays showed that the expressions of pro-apoptotic
genes including caspase-3, Fas, Fasl and Bax were lower in the OA
pretreatment groups than those in the model group. By contrast, the
expression of Bcl-2 was decreased in the model group but increased
by the pretreatment of OA (Fig.
4C-E).
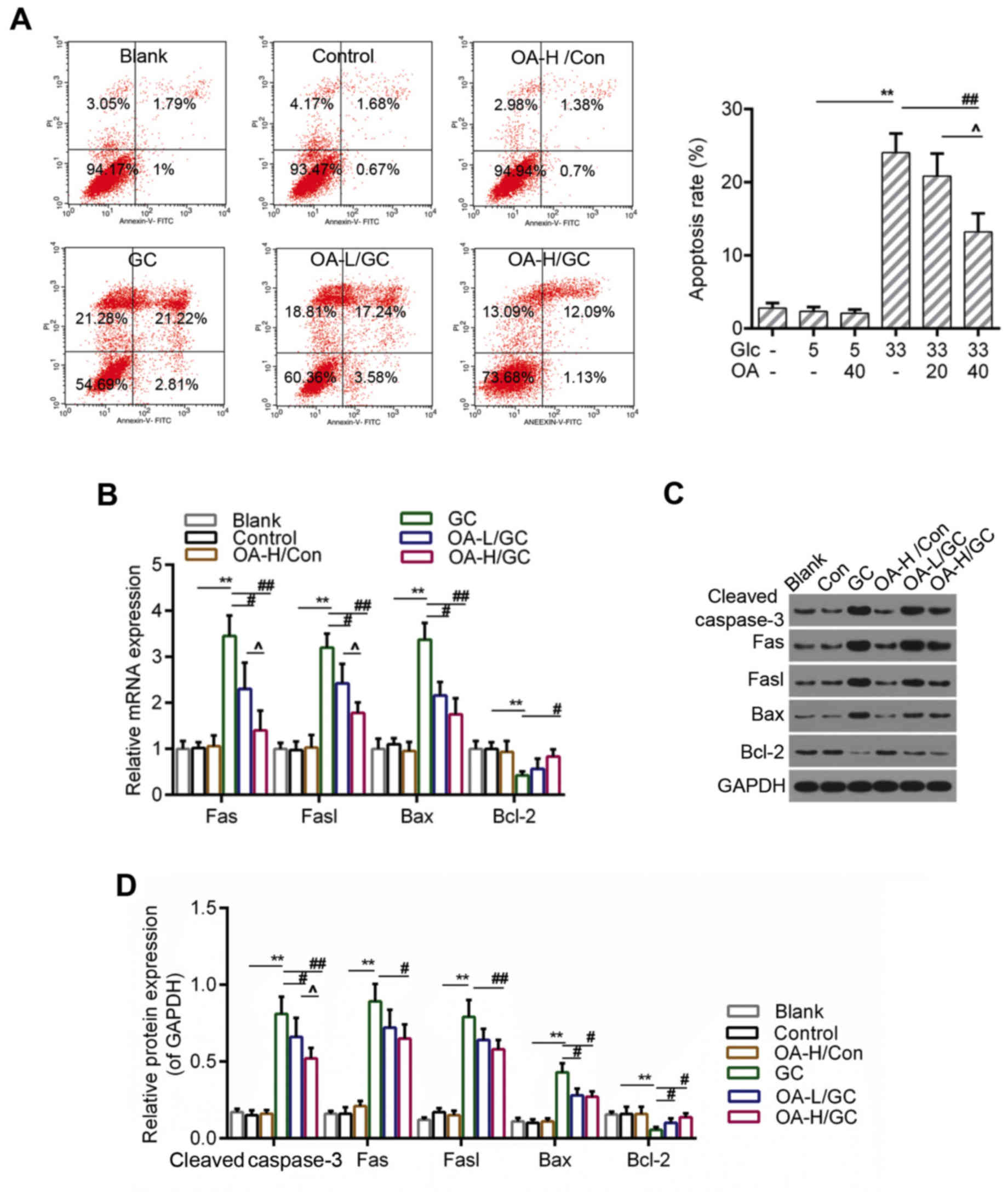 | Figure 3.OA decreased apoptosis of HUVECs. (A)
Flow cytometry analysis for detection of cell apoptosis using
Annexin V-FITC/PI staining. (B) Relative quantitative analysis of
caspase-3, Fas, Fasl, Bax, and Bcl-2 by RT-qPCR. (C and D) Western
blot analysis for caspase-3, Fas, Fasl, Bax, and Bcl-2. GAPDH was
assessed as sample loading control. Blank group (Blank), cuntreated
cells; control group (5 mM) (Con/Control), cells were treated with
5 mM glucose; high glucose (33 mM) model group (GC), cells were
treated with 33 mM glucose; positive control group (OA-H/Con),
cells were treated with 5 mM glucose and 40 µM OA; low OA (20 µM)
treatment group (OA-L/GC), cells were treated with 33 mM glucose
and 20 µM OA; high OA (40 µM) treatment group (OA-H/GC), cells were
treated with 33 mM glucose and 40 µM OA. *P<0.05, **P<0.01,
#P<0.05, ##P<0.01 and
^P<0.05 as indicated. OA, oleanolic acid; HUVECs;
human umbilical vein endothelial cells; FITC, fluorescein
isothiocyanate; PI, propidium iodide; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
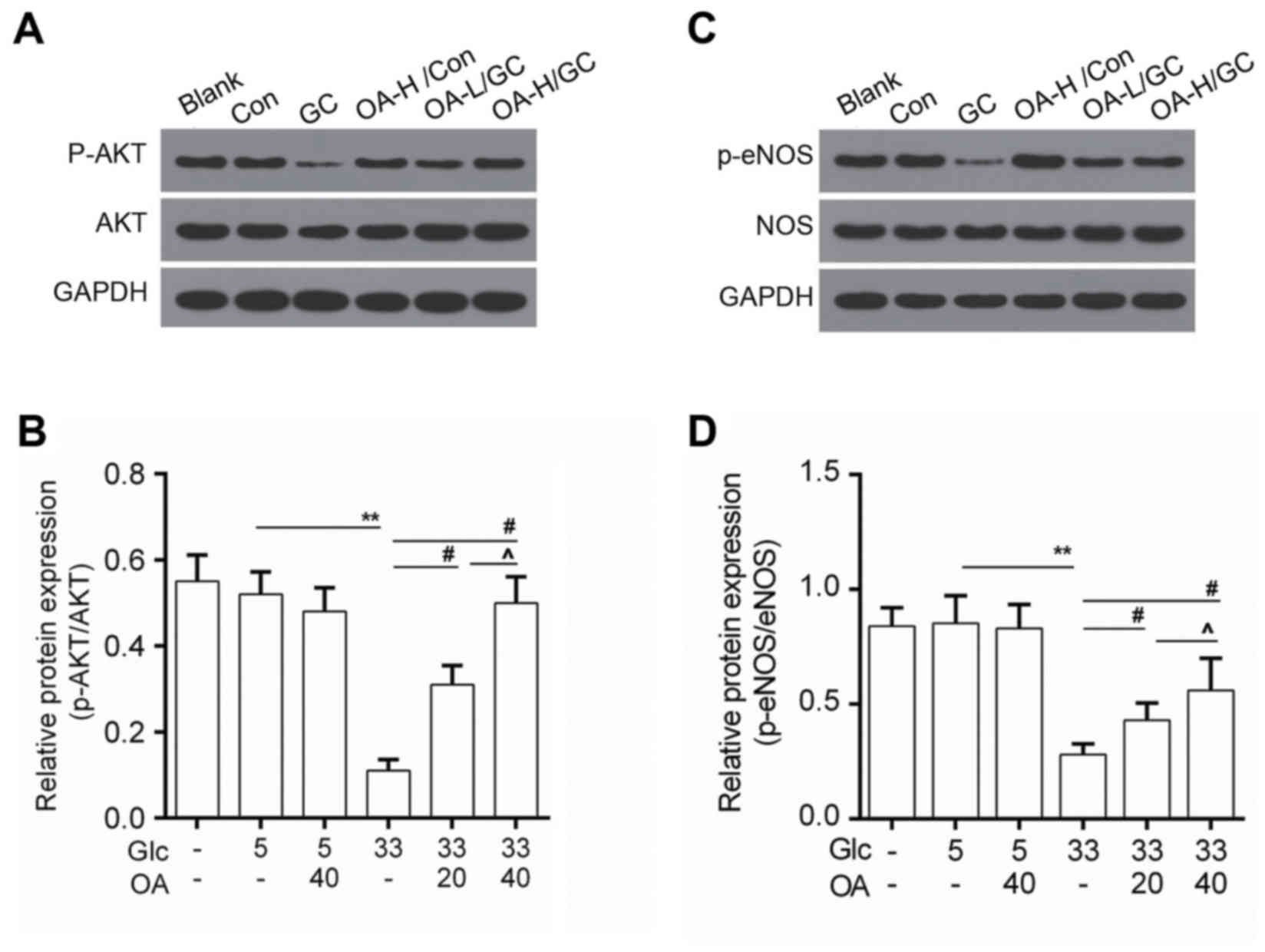
OA enhanced the activity of AKT/eNOS
signaling
To study the underlying mechanisms, the activity of
AKT/eNOS signals was determined by Western blot. Data showed that
protein level of phosphorylated AKT (p-AKT) was elevated in the OA
pretreatment groups (Fig. 4A and
B). Meanwhile, we noticed that the phosphorylation of eNOS
(p-eNOS) was higher in the OA pretreatment groups than that in the
model group (Fig. 4C and D).
Discussion
AS is an arteriosclerotic vascular disease in
endothelial cells. Diabetes is one of the major risk factors for
the formation of AS (41).
Recently, OA has attracted much attention in protecting against AS
(42,43).
In the presented study, we found that OA could
improve cell viability of HUVECs that was declined by the high
glucose treatment, suggesting a protective effect of OA on HUVECs.
To explore the role of OA in high-glucose mediated injury of
HUVECs, we detected the oxidative response of HUVECs after the
treatment with OA. The results showed that OA recovered the NO
generation and decreased MDA content. The activities of SOD and CAT
were higher in the OA pretreatment groups than those in the model
group, indicating that OA had a potential of protecting against the
hyperglycemia injury in diabetes via suppressing the oxidative
stress. The cell viability is largely dependent on the function of
mitochondria (44). Thus, we
detected the function of mitochondria by estimating the ROS
production and MMP. Interestingly, our results revealed that ROS
burst and MMP loss caused by high glucose were alleviated in the OA
pretreatment groups. Moreover, apoptosis was reported to be closely
related to oxidative stress (45).
Therefore, the apoptosis of HUVECs was detected by flow cytometry
assay. Results showed that apoptosis of HUVECs caused by high
glucose was depressed by OA. Additionally, it is well known that
caspase-3 (46), Fas and Fasl
(47) act as pro-apoptotic
signals. Bcl-2 (48) is considered
as an important anti-apoptotic factor. We found that the
expressions of caspase-3, Fas and Fasl decreased in the OA
pretreatment groups both in the transcriptional and translated
levels. Conversely, the expression of Bcl-2 increased in the OA
pretreatment groups in comparison to the model group, indicating
that OA depressed the cell apoptosis of HUVECs by modulating the
expression of apoptosis-related factors.
Furthermore, according to previous study, AKT/eNOS
contributes to the prevention of apoptosis in endothelial cells
(49). Another previous study
pointed out that the phosphorylation of AKT/eNOS was decreased by
high glucose in HUVECs (50). To
illustrate the action mechanism of OA underlying the high
glucose-induced HUVECs injury, we examined the activity of AKT/eNOS
signaling. The results showed that the pretreatment of OA increased
the levels of phosphorylated AKT (p-AKT) and phosphorylated eNOS
(p-eNOS) in comparison to model group. A previous study showed that
the endothelial injury was alleviated through enhancing the
activity of PI3K/Akt/eNOS signaling (51). Although the effects of OA in this
study still needed further validation, it provided a new molecular
insight for understanding the effects of OA on AS in diabetes.
Furthermore, OA could exert anti-tumor effect by inhibiting the
activation of Akt signaling (25,52).
This was contrary to the activation of Akt signaling by OA in this
study. These results seemed to be contradictory; however, they
demonstrated the protective effect of OA. The distinct results may
be attributed to different cell type and cell context. In addition,
due to the complexity of signal transduction, the signal modulation
net would be better understood with efforts from further
studies.
In summary, OA alleviated the cell injury mediated
by high glucose by improving the cell viability, increasing the NO
content, decreasing MDA level and rescuing the activities of SOD
and CAT. Moreover, the ROS burst and MMP loss caused by high
glucose could be mitigated by OA. The apoptosis induced by high
glucose was inhibited by OA via modulating the expressions of
caspase-3, Fas, Fasl and Bcl-2. Furthermore, the protective effects
of OA may be associated with the activation of AKT/eNOS signaling
pathway. Collectively, our results provided inspiration of new
therapeutic strategies for the treatment of AS on diabetics.
Acknowledgements
Not applicable.
Funding
This work was supported by Health and Family
Planning Commission of Sichuan Province Project (grant no.
2016-2017).
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
WZ and BC made substantial contributions to
conception and design of the study. JF, QL and XC contributed to
the data acquisition, analysis and interpretation. BC revised the
article for important intellectual content. QL provided final
approval of the version to be published. XC agreed to be
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of the work are appropriately
investigated and resolved. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Resnick HE and Howard BV: Diabetes and
cardiovascular disease. Annu Rev Med. 53:245–267. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee RT, Schoen FJ, Loree HM, Lark MW and
Libby P: Circumferential stress and matrix metalloproteinase 1 in
human coronary atherosclerosis. Implications for plaque rupture.
Arterioscler Thromb Vasc Biol. 16:1070–1073. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Salmi M, Stolen C, Jousilahti P, Yegutkin
GG, Tapanainen P, Janatuinen T, Knip M, Jalkanen S and Salomaa V:
Insulin-regulated increase of soluble vascular adhesion protein-1
in diabetes. Am J Pathol. 161:2255–2262. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nishikawa T, Edelstein D, Du XL, Yamagishi
S, Matsumura T, Kaneda Y, Yorek MA, Beebe D, Oates PJ, Hammes HP,
et al: Normalizing mitochondrial superoxide production blocks three
pathways of hyperglycaemic damage. Nature. 404:787–790. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Onat D, Brillon D, Colombo PC and Schmidt
AM: Human vascular endothelial cells: A model system for studying
vascular inflammation in diabetes and atherosclerosis. Curr Diab
Rep. 11:193–202. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Echtay KS, Murphy MP, Smith RA, Talbot DA
and Brand MD: Superoxide activates mitochondrial uncoupling protein
2 from the matrix side. Studies using targeted antioxidants. J Biol
Chem. 277:47129–47135. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Storz G and Imlay JA: Oxidative stress.
Curr Opin Microbiol. 2:188–194. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bullon P, Newman HN and Battino M:
Obesity, diabetes mellitus, atherosclerosis and chronic
periodontitis: A shared pathology via oxidative stress and
mitochondrial dysfunction? Periodontol. 64:139–153. 2014.
View Article : Google Scholar
|
|
9
|
Vornoli A, Pozzo L, Della Croce CM,
Gervasi PG and Longo V: Drug metabolism enzymes in a steatotic
model of rat treated with a high fat diet and a low dose of
streptozotocin. Food Chem Toxicol. 70:54–60. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang SJ, Je Lee W, Kim EA, Dal Nam K, Hahn
HG, Young Choi S and Cho SW: Effects of
N-adamantyl-4-methylthiazol-2-amine on hyperglycemia,
hyperlipidemia and oxidative stress in streptozotocin-induced
diabetic rats. Eur J Pharmacol. 736:26–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakata K, Kondo T, Mizuno N, Shoji M,
Yasui H, Yamamori T, Inanami O, Yokoo H, Yoshimura N and Hattori Y:
Roles of ROS and PKC-βII in ionizing radiation-induced eNOS
activation in human vascular endothelial cells. Vascul Pharmacol.
70:55–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou H and Wang C: Cytoprotective Effects
and Mechanisms of Δ-17 Fatty Acid Desaturase in Injured Human
Umbilical Vein Endothelial Cells (HUVECs). Med Sci Monit.
23:1627–1635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rubanyi GM: The role of endothelium in
cardiovascular homeostasis and diseases. J Cardiovasc Pharmacol. 22
Suppl 4:S1–S14. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mezzetti A: Pharmacological modulation of
plaque instability. Lupus. 14:769–772. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Haluzik M, Nedvídková J and Skrha J: The
influence of NO synthase inhibitor and free oxygen radicals
scavenger-methylene blue-on streptozotocin-induced diabetes in
rats. Physiol Res. 47:337–341. 1998.PubMed/NCBI
|
|
16
|
Szuster-Ciesielska A, Stachura A,
Słotwińska M, Kamińska T, Sniezko R, Paduch R, Abramczyk D, Filar J
and Kandefer-Szerszen M: The inhibitory effect of zinc on
cadmium-induced cell apoptosis and reactive oxygen species (ROS)
production in cell cultures. Toxicology. 145:159–171. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Antherieu S, Ledirac N, Luzy AP, Lenormand
P, Caron JC and Rahmani R: Endosulfan decreases cell growth and
apoptosis in human HaCaT keratinocytes: partial ROS-dependent
ERK1/2 mechanism. J Cell Physiol. 213:177–186. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baumgartner-Parzer SM, Wagner L,
Pettermann M, Grillari J, Gessl A and Waldhäusl W:
High-glucose-triggered apoptosis in cultured endothelial cells.
Diabetes. 44:1323–1327. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fulda S: Synthetic lethality by
co-targeting mitochondrial apoptosis and PI3K/Akt/mTOR signaling.
Mitochondrion. 19:85–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Michell BJ, Griffiths JE, Mitchelhill KI,
Rodriguez-Crespo I, Tiganis T, Bozinovski S, de Montellano PRO,
Kemp BE and Pearson RB: The Akt kinase signals directly to
endothelial nitric oxide synthase. Curr Biol. 9:845–848. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wiest R, Das S, Cadelina G, Garcia-Tsao G,
Milstien S and Groszmann RJ: Bacterial translocation in cirrhotic
rats stimulates eNOS-derived NO production and impairs mesenteric
vascular contractility. J Clin Invest. 104:1223–1233. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pollier J and Goossens A: Oleanolic acid.
Phytochemistry. 77:10–15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei J, Liu H, Liu M, Wu N, Zhao J, Xiao L,
Han L, Chu E and Lin X: Oleanolic acid potentiates the antitumor
activity of 5-fluorouracil in pancreatic cancer cells. Oncol Rep.
28:1339–1345. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Q, Liu H, Zhang L, Guo T, Wang P, Geng
M and Li Y: Synthesis and antitumor activities of naturally
occurring oleanolic acid triterpenoid saponins and their
derivatives. Eur J Med Chem. 64:1–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mu DW, Guo HQ, Zhou GB, Li JY and Su B:
Oleanolic acid suppresses the proliferation of human bladder cancer
by Akt/mTOR/S6K and ERK1/2 signaling. Int J Clin Exp Pathol.
8:13864–13870. 2015.PubMed/NCBI
|
|
26
|
Li L, Wei L, Shen A, Chu J, Lin J and Peng
J: Oleanolic acid modulates multiple intracellular targets to
inhibit colorectal cancer growth. Int J Oncol. 47:2247–2254. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martin-Aragón S, de las Heras B,
Sanchez-Reus MI and Benedi J: Pharmacological modification of
endogenous antioxidant enzymes by ursolic acid on
tetrachloride-induced liver damage in rats and primary cultures of
rat hepatocytes. Exp Toxicol Pathol. 53:199–206. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xin W, Rui L, Wei Z, Xiaodi Z, Nai L, Zhao
W, Wenli L, Xujun Q and Chunxu H: Oleanolic acid improves hepatic
insulin resistance via antioxidant, hypolipidemic and
anti-inflammatory effects. Mol Cell Endocrinol. 376:70–80. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang X, Chen Y, Abdelkader D, Hassan W,
Sun H and Liu J: Combination therapy with oleanolic acid and
metformin as a synergistic treatment for diabetes. J Diabetes Res.
2015:9732872015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mukundwa A, Langa SO, Mukaratirwa S and
Masola B: In vivo effects of diabetes, insulin and oleanolic acid
on enzymes of glycogen metabolism in the skin of
streptozotocin-induced diabetic male Sprague-Dawley rats. Biochem
Biophys Res Commun. 471:315–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han D, Zhang X, Zhang J, Guo X, Zheng Y,
Sui S and Zheng J: Oleanolic acid suppresses vascular smooth muscle
cell proliferation by increasing lincRNA-p21 expression. Oncol
Lett. 12:3519–3522. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Engelgau MM, Narayan KM and Herman WH:
Screening for type 2 diabetes. Diabetes Care. 23:1563–1580. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu J, Deng W, Fan L, Tian L, Jin L, Jin
Z, Guo Q, Xu Y and Li N: The role of radix hedysari polysaccharide
on the human umbilical vein endothelial cells (HUVECs) induced by
high glucose. Eur J Intern Med. 23:287–292. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhong ZY and Tang Y: Upregulation of
periostin prevents high glucose-induced mitochondrial apoptosis in
human umbilical vein endothelial cells via activation of Nrf2/HO-1
signaling. Cell Physiol Biochem. 39:71–80. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tsai SJ and Yin MC: Antioxidative and
anti-inflammatory protection of oleanolic acid and ursolic acid in
PC12 cells. J Food Sci. 73:H174–H178. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ludescher C, Thaler J, Drach D, Drach J,
Spitaler M, Gattringer C, Huber H and Hofmann J: Detection of
activity of P-glycoprotein in human tumour samples using rhodamine
123. Br J Haematol. 82:161–168. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Arocho A, Chen B, Ladanyi M and Pan Q:
Validation of the 2-DeltaDeltaCt calculation as an alternate method
of data analysis for quantitative PCR of BCR-ABL P210 transcripts.
Diagn Mol Pathol. 15:56–61. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Indo HP, Davidson M, Yen HC, Suenaga S,
Tomita K, Nishii T, Higuchi M, Koga Y, Ozawa T and Majima HJ:
Evidence of ROS generation by mitochondria in cells with impaired
electron transport chain and mitochondrial DNA damage.
Mitochondrion. 7:106–118. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Singh AP, Sarkar S, Tripathi M and
Rajender S: Mucuna pruriens and its major constituent L-DOPA
recover spermatogenic loss by combating ROS, loss of mitochondrial
membrane potential and apoptosis. PLoS One. 8:e546552013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Haffner SM, Lehto S, Rönnemaa T, Pyörälä K
and Laakso M: Mortality from coronary heart disease in subjects
with type 2 diabetes and in nondiabetic subjects with and without
prior myocardial infarction. N Engl J Med. 339:229–234. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pan Y, Zhou F, Song Z, Huang H, Chen Y,
Shen Y, Jia Y and Chen J: Oleanolic acid protects against
pathogenesis of atherosclerosis, possibly via FXR-mediated
angiotensin (Ang)-(1–7) upregulation. Biomed Pharmacother.
97:1694–1700. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang Q, Hao R, Wang W, Gao H and Wang C:
SIRT1/Atg5/autophagy are involved in the antiatherosclerosis
effects of ursolic acid. Mol Cell Biochem. 420:171–184. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Benard G and Karbowski M: Mitochondrial
fusion and division: Regulation and role in cell viability. Semin
Cell Dev Biol. 20:365–374. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Buttke TM and Sandstrom PA: Oxidative
stress as a mediator of apoptosis. Immunol Today. 15:7–10. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Salvesen GS: Caspases: Opening the boxes
and interpreting the arrows. Cell Death Differ. 9:3–5. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wajant H: The Fas signaling pathway: More
than a paradigm. Science. 296:1635–1636. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vaux DL, Cory S and Adams JM: Bcl-2 gene
promotes haemopoietic cell survival and cooperates with c-myc to
immortalize pre-B cells. Nature. 335:440–442. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park CW, Kim HW, Lim JH, Yoo KD, Chung S,
Shin SJ, Chung HW, Lee SJ, Chae CB, Kim YS and Chang YS: Vascular
endothelial growth factor inhibition by dRK6 causes endothelial
apoptosis, fibrosis, and inflammation in the heart via the Akt/eNOS
axis in db/db mice. Diabetes. 58:2666–2676. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wu J, Lei MX, Xie XY, Liu L, She YM, Mo J
and Wang S: Rosiglitazone inhibits high glucose-induced apoptosis
in human umbilical vein endothelial cells through the PI3K/Akt/eNOS
pathway. Can J Physiol Pharmacol. 87:549–555. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ou HC, Lee WJ, Lee SD, Huang CY, Chiu TH,
Tsai KL, Hsu WC and Sheu WH: Ellagic acid protects endothelial
cells from oxidized low-density lipoprotein-induced apoptosis by
modulating the PI3K/Akt/eNOS pathway. Toxicol Appl Pharmacol.
248:134–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li X, Song Y, Peng Z, Zhu H, Chen L, Xiao
Y and Xing Y: Oleanolic acid inhibits cell survival and
proliferation of prostate cancer cells in vitro and in vivo through
the PI3K/Akt pathway. Tumour Biol. 37:7599–7613. 2016. View Article : Google Scholar : PubMed/NCBI
|