Introduction
Glioblastoma (GBM) is one of the most common types
of malignant tumor and is a prevalent type of primary cancer of the
central nervous system in adults (1–4).
Developments in the treatments for GBM, including chemotherapy with
temozolomide, radiotherapy, and combination therapy with tumor
resection have been made; in patients with newly diagnosed
malignant glioma, improvements in the 2-year survival rate (from 11
to 27%), the 3-year survival rate (from 4 to 16%) and the 5-year
survival rate (from 2 to 10%) were observed in 2014 (5,6).
However, these survival rates remain relatively low; the underlying
molecular mechanisms of GBM-associated tumorigenesis, chemotherapy
resistance, progression and metastasis require further
investigation. Therefore, it is of clinical importance that novel
therapeutic targets and biomarkers associated with GBM are
identified.
MicroRNAs (miRNA/miRs) are a class of small
non-coding RNAs of 19–24 nucleotides in length that serve important
regulatory roles in post-transcriptional gene expression (7,8).
Additionally, downregulation of miR-454-3p has been identified in
certain types of cancer in humans, including pancreatic ductal
adenocarcinoma (9) and breast
cancer (10), while upregulation
of miR-454 has been observed in other types of cancer, including
hepatocellular carcinoma, non-small cell lung cancer (11) and colorectal cancer (12). In pancreatic ductal adenocarcinoma,
it has been reported that miR-454 is able to regulate stromal cell
proliferation, thereby controlling the growth of pancreatic ductal
adenocarcinoma (13).
Additionally, in osteosarcoma, miR-454 functions as a tumor
suppressor gene that suppresses cell proliferation and invasion by
directly targeting c-Met (14). In
hepatocellular carcinoma, miR-454 functions as an oncogene by
inhibiting chromodomain-helicase-DNA-binding protein 5, resulting
in the inhibition of tumor cell proliferation and invasion
(15). Additionally, in uveal
melanoma, miR-454 is upregulated and promotes cell proliferation,
colony formation, invasion and induction of the cell cycle
(16). Most relevant to the
present study, a recent investigation into GBM revealed that
miR-454 was able to inhibit cell proliferation by suppressing
pyruvate dehydrogenase kinase 1 expression (17); however, the role and underlying
mechanisms of miR-454 downregulation in GBM are largely
unknown.
Cytoplasmic polyadenylation element-binding protein
1 (CPEB1) is a sequence-specific RNA-binding protein that regulates
mRNA translation by dynamically controlling the length of the
poly(A) tail length of an mRNA transcript (18). The biological roles of
CPEB1-mediated regulation include the control of cell cycle
progression, cellular senescence and inflammation (19–21).
Additionally, CPEB1 has been considered to serve as a tumor
suppressor gene. For instance, a recent study reported that CPEB1
mediated epithelial-to-mesenchymal transition (EMT) and breast
cancer metastasis by regulating the apical localization of tight
junction protein ZO-1 mRNA (22).
In melanoma, miR-455-5p has been reported to promote melanoma
metastasis via inhibition of CPEB1 (23); in glioma, knockdown of CPEB1
reduces cell senescence by regulating the expression and
distribution of cellular tumor antigen p53, and CPEB1 was also
demonstrated to be directly regulated by the tumor suppressor
miR-101 (24).
The results of the present study revealed that the
overexpression of miR-454 in GBM cells in vitro inhibited
cell proliferation, migration and invasion. Importantly, the
present study also observed that miR-454-3p negatively regulated
the target gene CPEB1. Thus, the present study investigated whether
miR-454-3p and CPEB1 may be a novel therapeutic target for the
treatment of patients with GBM.
Patients and methods
Patient and tissue samples
A total of 30 human glioma and matched adjacent
tissues were collected between January 2015 and December 2016 at
Huai'an First People's Hospital (Huai'an, China). The median age of
the patients was 51 years, and the age ranged from 29–73 years and
there were 19 male and 11 female. All tissue samples were collected
for analysis upon obtaining informed consent from all patients. In
addition, patients did not receive chemotherapy, radiotherapy or
any other treatments prior to surgery. The present study was
approved by the Research Ethics Committee of Huai'an First People's
Hospital (Huai'an, China).
Cell culture
Human GBM cell lines (LN-229, A172 and GL15) and
normal human astrocyte cells (HA1800) were obtained from the
Shanghai Institutes for Biological Sciences (Chinese Academy of
Sciences, Shanghai, China). Adherent cultures of LN-229 and HA1800
cell lines were maintained in high-glucose Dulbecco's modified
Eagle's medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; Beijing
Solarbio Science and Technology Co., Ltd., Beijing, China). All
cells were cultured in a humidified incubator at 37°C in an
atmosphere of 5% CO2.
Cell transfection
The human GBM cells (LN-229) were seeded in 6-well
plates (2×105 cells/well) 1 day prior to transfection,
which was conducted when cells reached 60–70% confluence. The
LN-229 cells were untransfected or transfected with mimic control
or miR-454-3p mimic (Shanghai GenePharma Co., Ltd., Shanghai,
China) at a concentration of 40 nM, and the untreated cells served
as the control. The successful transfection was determined by
RT-qPCR. The sequences of miR-454-3p was:
5′-UAGUGCAAUAUUGCUUAUAGGGU-3′, miR-454-3p mimic was:
5′-UAGUGCAAUAUUGCUUAUAGGGU-3 and mimic control was:
5′-UCACAACCUCCUAGAAAGAGUAGA-3′. Then the cells were incubated for
24 h to continue the further analysis. Transfection with the miRNAs
and with the plasmids for the luciferase assay (described below)
was performed using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Human GBM cell lines (LN-229, A172 and GL15) and
normal human astrocyte cells (HA1800) were lysed in
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) for total RNA extraction; a total of 1 µg total RNA was used
for cDNA synthesis with an RNA PCR kit (Takara Biotechnology Co.,
Ltd., Dalian, China). For cDNA synthesis, samples were incubated at
42°C for 30 min, 99°C for 5 min and 5°C for 5 min. qPCR was
conducted using an ABI7500 Real-Time PCR Instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The qPCR conditions
were as follows: Pre-denaturation at 95°C for 5 min, followed by
initiation at 94°C for 30 sec, annealing at 60°C for 30 sec, and
elongation at 75°C for 1.5 min for 32 cycles, following which
samples were stored at 4°C. Hairpin-it™ of miR-454-3p
and U6 RT-qPCR Primer Set (Shanghai GenePharma Co., Ltd.) were used
to measure the relative quantity of miR-454-3p; the expression of
miR-454 was normalized to the endogenous expression of U6. The
primers used were as follows: miR-454-3p forward,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACCCTATA-3′ and reverse,
5′-ACACTCCAGCTGGGTAGTGCAATATTGCTTA-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
Additionally, metadherin, astrocyte elevated gene-1 (AEG-1) and
matrix metalloproteinase-9 (MMP-9) RT-qPCR Primer Sets (Shanghai
GenePharma Co., Ltd.) were used to measure the relative quantities
of metadherin, AEG-1 and MMP-9, with normalization of the mRNA
expression levels to the endogenous expression levels of GAPDH. The
primers used were as follows: Metadherin forward,
5′-AAATGGGCGGACTGTTGAAGT-3′ and reverse,
5′-CTGTTTTGCACTGCTTTAGCAT-3′; AEG-1 forward,
5′-AAATGGGCGGACTGTTGAAGT-3′ and reverse,
5′-CTGTTTTGCACTGCTTTAGCAT-3′; MMP-9 forward,
5′-GGGACGCAGACATCGTCATC-3′ and reverse, 5′-TCGTCATCGTCGAAATGGGC-3′;
and GAPDH forward, 5′TGTGGGCATCAATGGATTTGG3′ and reverse,
5′-ACACCATGTATTCCGGGTCAAT-3′. All reactions were performed in
triplicate. The relative expression of genes was normalized to
GAPDH. Data were analyzed using the 2−ΔΔCq method
(25).
Cell Counting kit-8 (CCK-8) assay
Cellular proliferation was measured with a CCK-8 kit
(Dojindo Molecular Technologies, Inc., Kumamoto, Japan), according
to the manufacturer's protocol. LN-229 cells were assigned with
untreated (control), mimic control and miR-454-3p mimic groups. At
24 h post-transfection, 100 µl cells suspension were seeded into
96-well plates at a density of 8×103 cells/well and
incubated at 37°C for 6 h. At the indicated time points (12, 24 and
48 h), 10 µl CCK-8 solution was added to each well. Following
incubation at 37°C for 1 h, the absorbance was measured with a
plate reader at 450 nm.
Transwell and scratch assays of LN-229
cell migration and invasion
Transwell culture inserts (8-mm pore size; Falcon;
BD Biosciences, Franklin Lakes, NJ, USA) were placed into the wells
of 24-well culture plates, separating the upper and lower chambers.
LN-229 cells were assigned with untreated (control), mimic control
and miR-454-3p mimic groups. For the cell invasion assay, Matrigel
(BD Biosciences) was used to pre-coat the upper side of the
membrane, which was subsequently incubated at 37°C for 1 h for gel
formation, and hydrated in FBS for 2 h prior to use. In the lower
chamber, 600 µl DMEM containing 10% FBS was added, and the three
groups cells were added to the upper chamber at a density of
1×105 cells/well respectively. Following incubation at
37°C for 24 h. The level of migration was observed under an optical
microscope (Leica DMI6000B; Leica Microsystems GmbH, Wetzlar,
Germany), and counted for 5 random (magnification, ×100) fields per
well. Cell counts are expressed as the mean number of cells per
field of view (26).
For the cell migration assay, the LN-229 cells in
the logarithmic phase were cultured in DMEM in 24-well plates at a
density of 104 cells/well to obtain a monolayer culture.
The monolayer was carefully scratched with a new 1-ml pipette tip
across the center of the well, with the long axis of the tip
remaining perpendicular to the bottom of the well, such that the
width of the scratch was equal to the outer diameter of the tip.
Following scratching, the cells were incubated at 37°C with 5%
CO2 for 24 h. Images of the migrated cells were captured
at 0 and 24 h and were captured from 5 random (magnification, ×5)
fields in each sample.
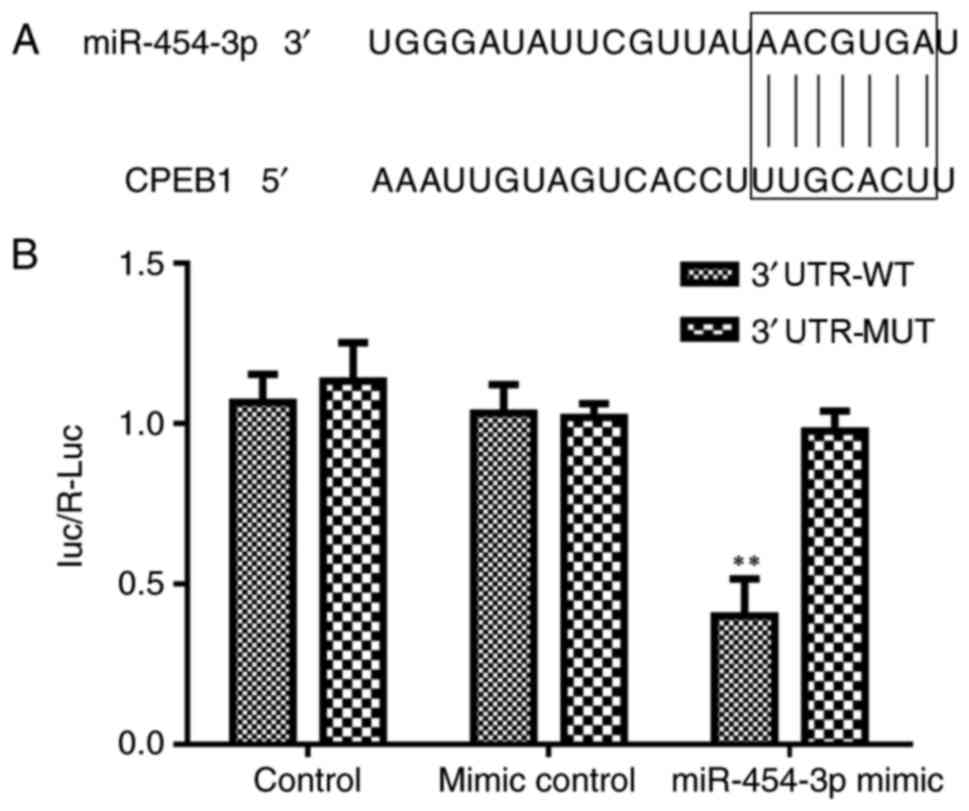
Target prediction
According to TargetScan 7.1 (http://www.targetscan.org/vert_71/), it was predicted
that CPEB1 may be targeted by miR-454-3p.
Luciferase reporter assays
For the luciferase reporter assay, LN-229 cells were
seeded in 96-well plates at a density of 104 cells/well
and incubated at 37°C with 5% CO2 for 24 h.
Subsequently, pGL3-CPEB1-3′untranslated region (UTR)-wild type (WT)
or pGL3-CPEB1-3′UTR-mutant plasmids (constructed by Beyotime
Institute of Biotechnology, Haimen, China) were transfected into
the control, miR-454-3p mimics and negative control miR-454-3p
mimics groups using Lipofectamine 2000. The transfection efficiency
was normalised with an internal control Renilla luciferase
vector (pRL-CMV; Promega Corporation, Madison, WI, USA). Following
48 h incubation, a luciferase assay kit (cat. no. RG005; Beyotime
Institute of Biotechnology) was used to measure the reporter
activity according to the manufacturer's protocols.
Western blot analysis
After LN-229 cells were untransfected or transfected
with mimic control or miR-454-3p mimic cells were washed with cold
PBS and treated with a lysis buffer (cat. no. C3702; Beyotime
Institute of Biotechnology). The quality was detected using BCA
method and protein concentration was measured with a NanoDrop
instrument (NanoDrop; Thermo Fisher Scientific, Inc., Pittsburgh,
PA, USA). A total of 30 µg protein samples were separated by 15%
SDS-PAGE, and the proteins were transferred onto polyvinylidene
difluoride membranes. Subsequently, the membranes were blocked with
a buffer containing 10% non-fat milk in PBS with 0.05% Tween-20 for
2 h at room temperature, and then incubated with primary antibodies
against metadherin, AEG-1, MMP-9, CPEB1 and GAPDH for 1 h at room
temperature. Additionally, the membranes were incubated at room
temperature with horseradish peroxidase-labeled goat anti-rabbit
antibody (1:1,000; cat. no. A0208; Beyotime Institute of
Biotechnology, Haimen, China) conjugated to horseradish peroxidase
for 45 min. Protein bands were visualized by enhanced
chemiluminescence (Abbott Laboratories, Arlington Heights, IL,
USA). Evaluation of target protein expression was performed using
ImageJ version 1.38 (National Institutes of Health, Bethesda, MD,
USA).
The primary antibodies [anti-metadherin (1:500; cat.
no. MA515564), MMP-9 (1:1,000; cat. no. PA516851), CPEB1 (1:1,000;
cat. no. PA520561) and GAPDH (1:500; cat. no. PA519440)] were
purchased from Wuhan Khayal Bio-Technology Co., Ltd. (Wuhan,
China); anti-AEG-1 (1:1,000; cat. no. PL032231R) was purchased from
Otwo Biotech (Shenzhen), Inc. (Shenzhen, China).
Statistical analysis
All results were confirmed in at least three
independent experiments, and the qualitative data from single
representative experiments are presented. All quantitative data are
presented as the mean ± standard deviation. Statistical analysis
was performed using GraphPad Prism version 5.01 (GraphPad Software,
Inc., La Jolla, CA, USA). A Student's t-test was used for
comparisons between two groups. One-way analysis of variance
followed by a least significant difference post hoc test was used
for comparisons between multiple groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
miR-454-3p expression is significantly
downregulated in glioma tissues and GBM cell lines
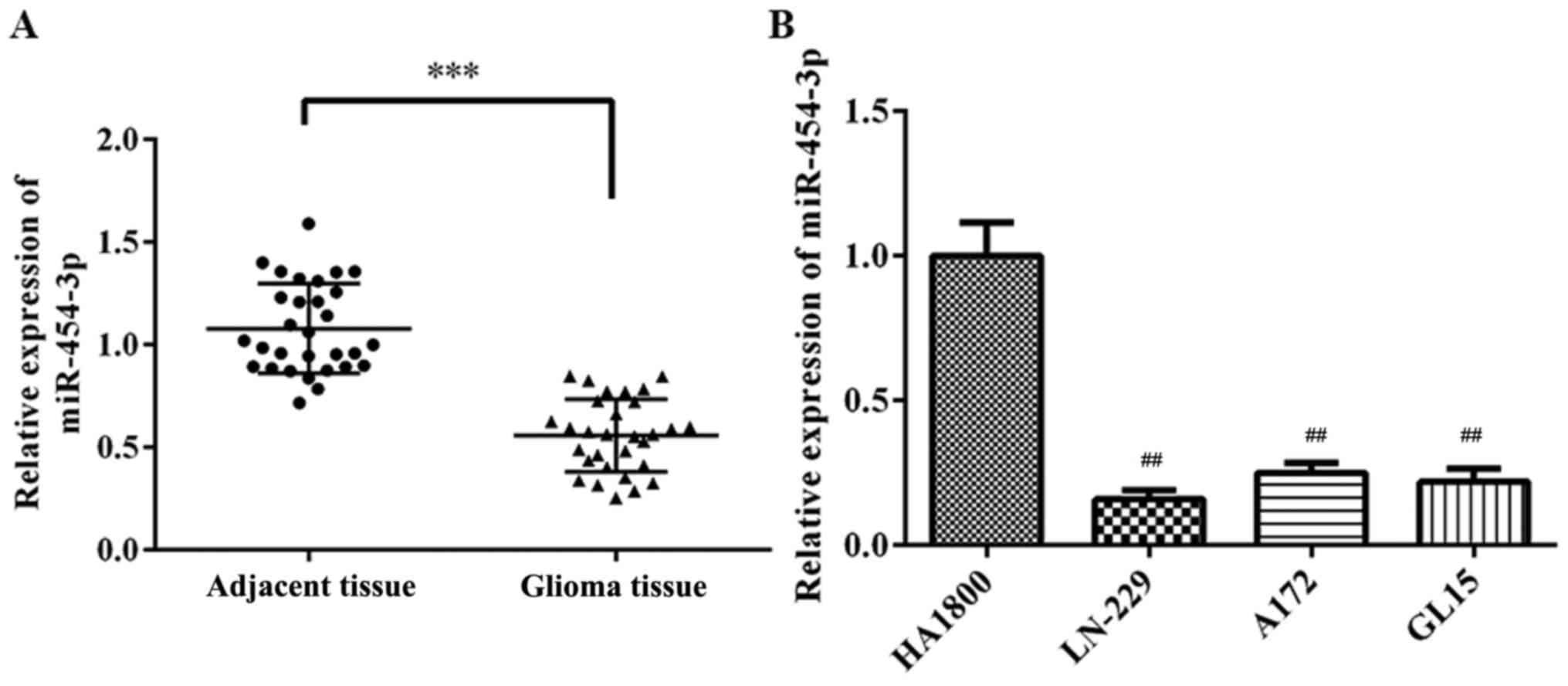
To investigate the role of miR-454-3p in GBM
development, the expression levels of miR-454-3p in tissue samples
and GBM cell lines were analyzed by RT-qPCR. As presented in
Fig. 1, it was observed that
miR-454-3p levels were significantly lowered in glioma tissues and
GBM cell lines (LN-229, A172 and GL15) compared with adjacent
tissues and normal human astrocyte cells (HA1800) (P<0.01),
respectively. From the results of Fig.
1, the miR-454-3p levels were significantly lower in LN-299
cell than in A172 and GL15 cells, so LN-229 cells were chosen for
further analysis.
Overexpression of miR-454-3p inhibits
the proliferation of LN-229 cells
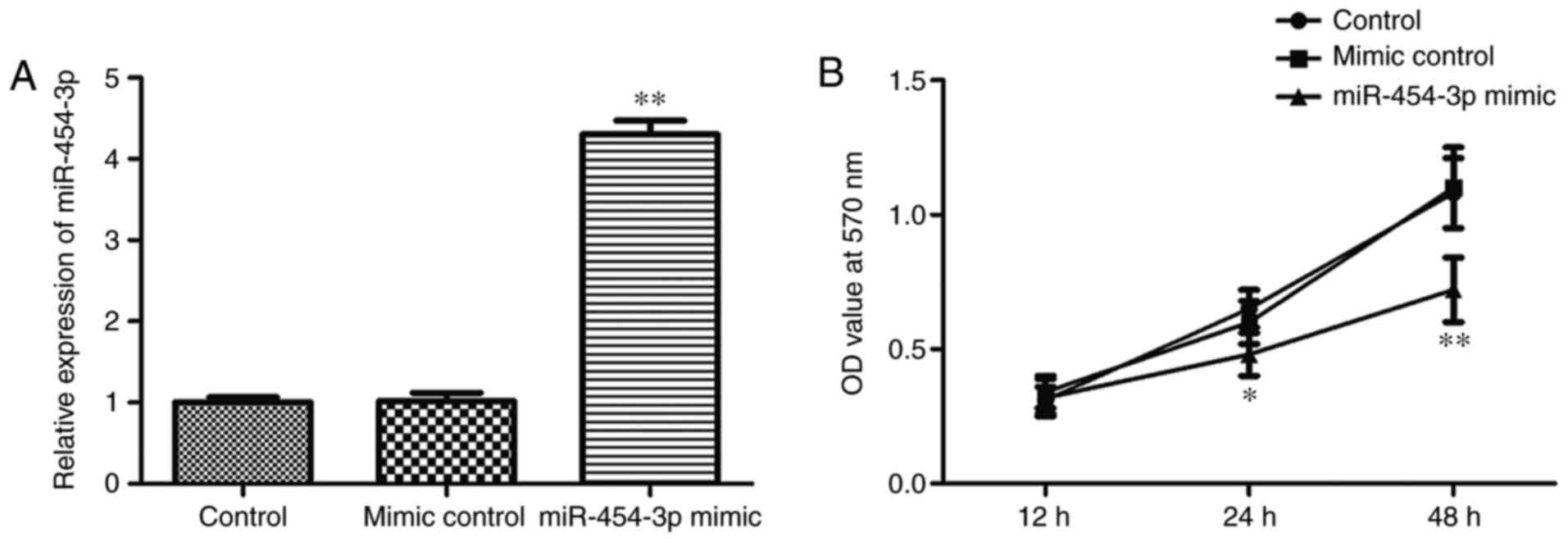
To investigate whether miR-454-3p serves a role as a
tumor suppressor in GBM, the effects of miR-454-3p overexpression
on the proliferation of GBM cells were analyzed. LN-229 cells were
transfected with control miR, miR-454-3p mimic or negative control
miR-454-3p mimic, and the relative expression of miR-454-3p was
verified by RT-qPCR (Fig. 2A). As
was demonstrated in Fig. 2A, the
miR-454-3p level was significantly increased following
overexpression of miR-454-3p compared with the negative control
miR-454-3p mimic (P<0.01). In addition, the effects of
miR-454-3p on cell proliferation was investigated via a CCK-8
assay. Analysis of the results indicated that the overexpression of
miR-454-3p significantly suppressed LN-229 cell proliferation
compared with the negative control group (Fig. 2B). Above date suggested that
miR-454-3p overexpression could inhibit the proliferation of LN-229
cells.
Overexpression of miR-454-3p inhibits
the invasion of LN-229 cells
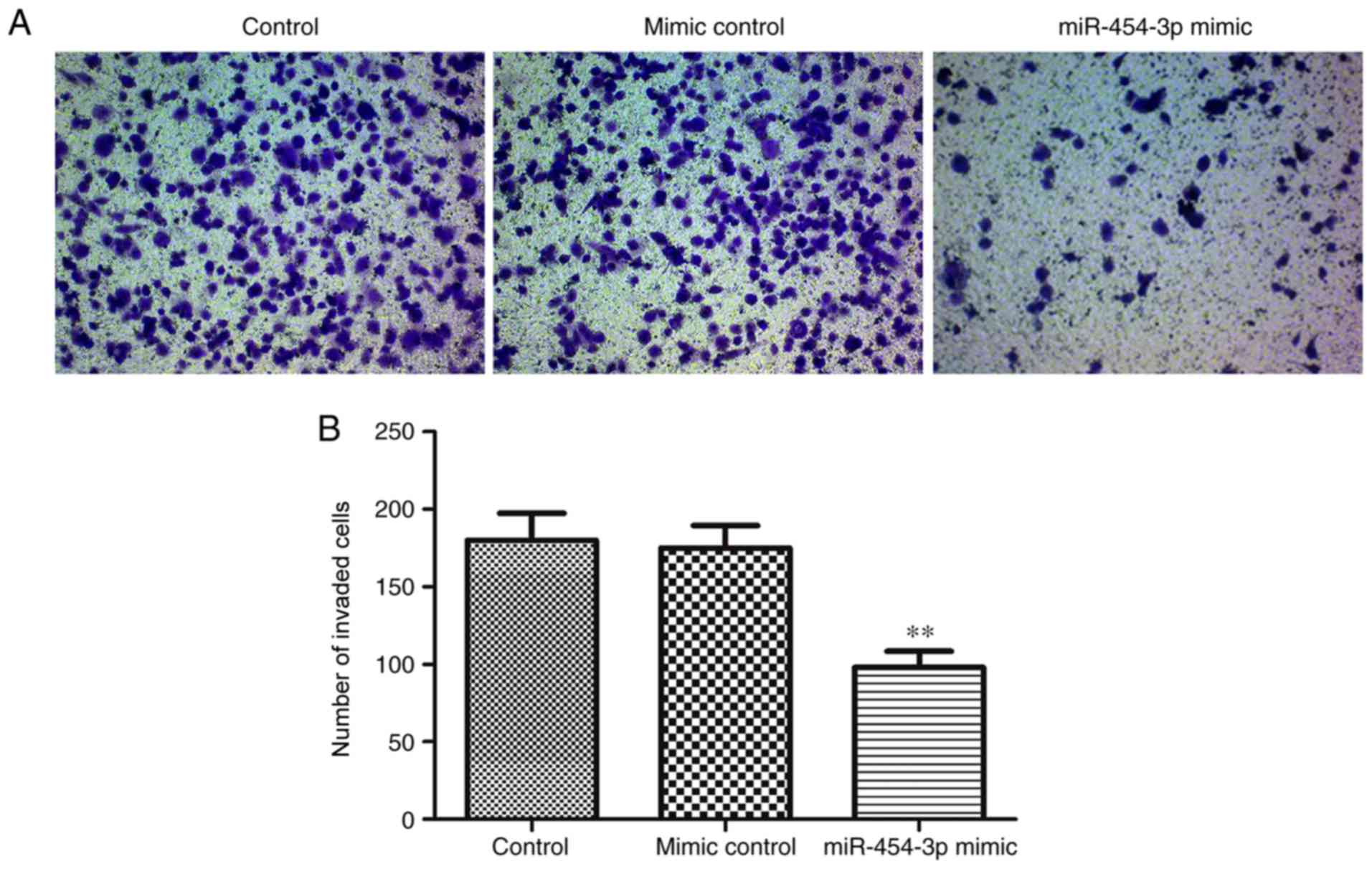
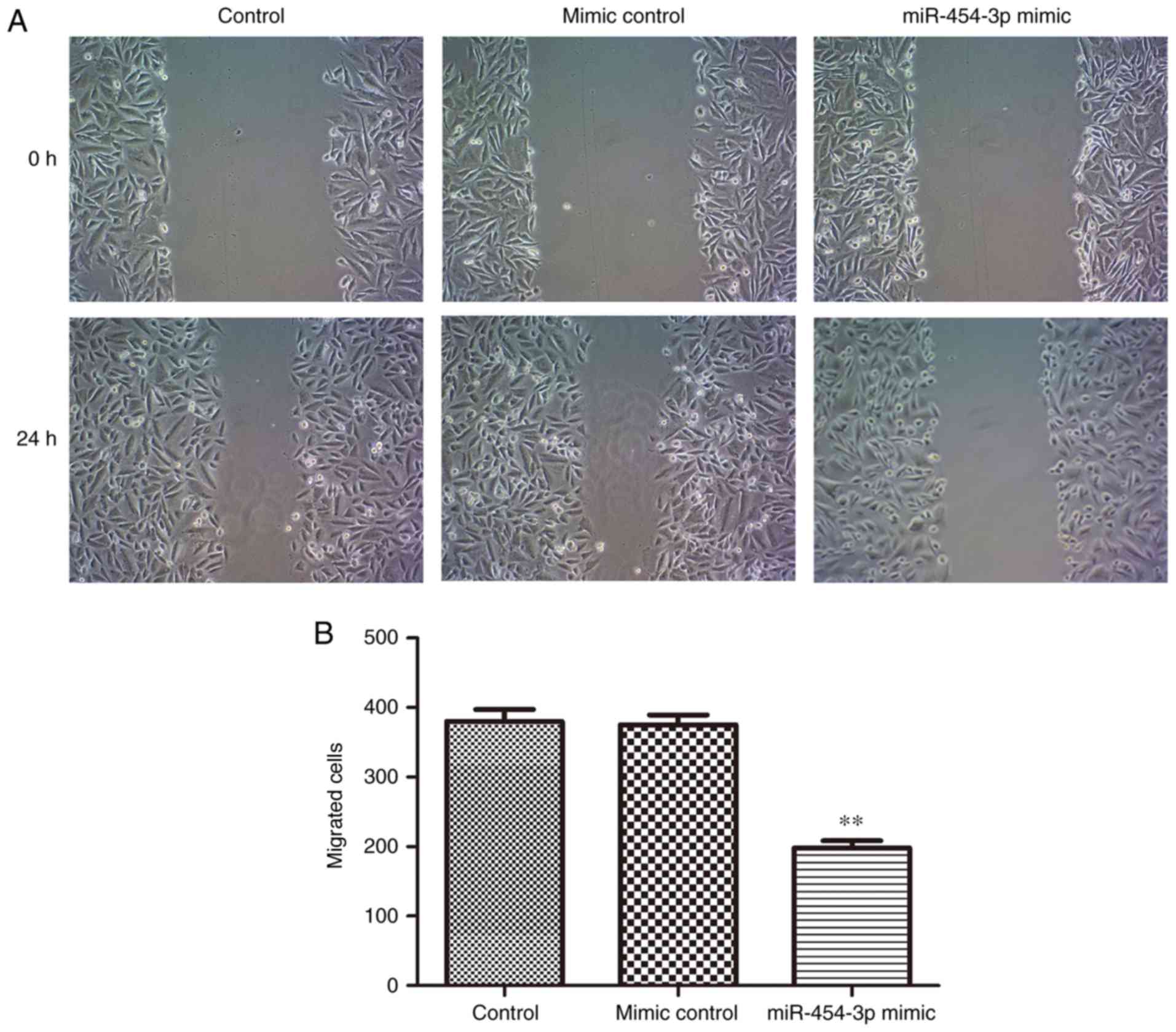
As migration and invasion are two key stages
associated with malignant progression and metastasis (27), the effects of miR-454-3p on the
migration and invasion of LN-229 cells was investigated in the
present study. The results of the Transwell and scratch assays
indicated that the overexpression of miR-454-3p significantly
inhibited the invasion and migration of LN-229 cells compared with
the negative control group (Figs.
3 and 4). Collectively, these
results suggested that miR-454-3p overexpression exerted tumor
suppressive effects by suppressing the proliferation, migration and
invasion of the LN-229 GBM cell line.
Metadherin has been reported to regulate mesenchymal
marker protein expression in numerous types of tumor and to promote
cancer metastasis (28–30). Additionally, AEG-1 has been
reported to serve a pivotal oncogenic role in tumorigenesis
(31,32), and MMPs, a family of zinc-binding
proteins, including MMP-9, have been demonstrated to serve
important roles in tumor cell invasion and metastasis due to their
ability to degrade the extracellular matrix (33,34).
In the present study, western blotting and RT-qPCR revealed that
miR-454-3p significantly decreased the protein and mRNA expression
levels of CPEB1, metadherin, AEG-1 and MMP-9 compared with in the
control group (Fig. 5).
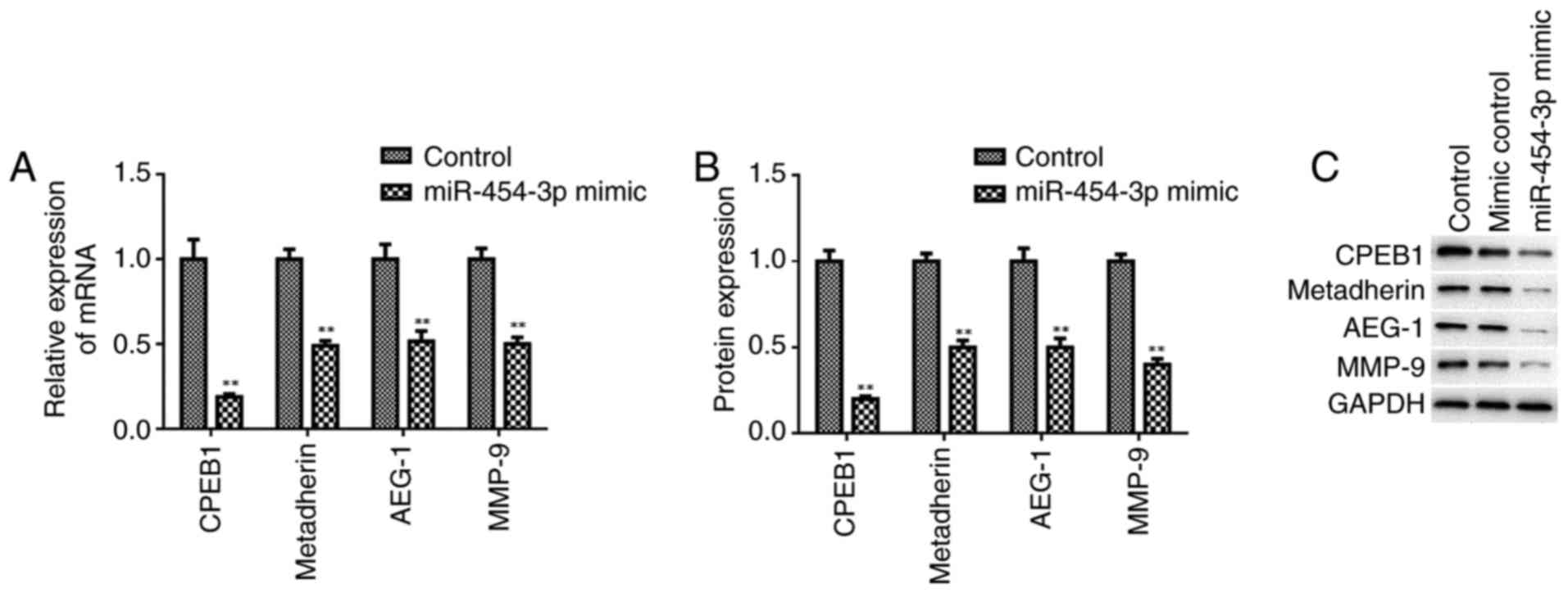 | Figure 5.Overexpression of miR-454-3p inhibits
the expression of tumor metastasis-associated proteins. The
relative expression levels of mRNA were quantified by reverse
transcription-quantitative polymerase chain reaction and protein
expression levels were determined by western blotting. (A) Relative
mRNA expression levels of CPEB1, metadherin, AEG-1 and MMP-9 in
LN-229 cells. (B) Protein expression profiles of CPEB1, metadherin,
AEG-1 and MMP-9 in LN-229 cells. (C) Western blotting image of the
protein bands for CPEB1, metadherin, AEG-1, MMP-9 and internal
reference GAPDH in LN-229 cells. **P<0.01 vs. respective
control. AEG-1, astrocyte elevated gene-1; CPEB1, cytoplasmic
polyadenylation element-binding protein 1; miR, microRNA; MMP-9,
matrix metalloproteinase-9. |
CPEB1 is a target gene of
miR-454-3p
The TargetScan database was used to identify the
potential target mRNA of miR-454-3p in the present study. The
results demonstrated that CPEB1 was a putative target gene of
miR-454-3p (Fig. 6A).
Subsequently, a dual-luciferase activity assay revealed that
miR-454-3p significantly suppressed the luciferase activity of the
WT 3′-UTR of CPEB1 compared with that of the mutant type (Fig. 6B). Furthermore, the overexpression
of miR-454-3p was observed to suppress the protein expression of
CPEB1 in the LN-229 cells by western blot analysis (Fig. 5B). These data indicated that CPEB1
is a direct target of miR-454-3p in GBM LN-229 cells.
Discussion
GBM is among the most lethal and aggressive forms of
brain cancer, and accounts for 15% of brain malignancies (35). Despite technological improvements
in the diagnosis and treatment of GBM, the rates of treatment
failure and recurrence remain an issue. The poor prognosis of
patients with GBM is primarily due to late diagnoses and poor
responses to chemotherapy (36).
Therefore, further investigation is urgently required to identify
and develop novel therapeutic approaches for the prevention and
treatment of GBM.
Recent studies have revealed that miRNAs contribute
to the progression and development of GBM (37,38).
For example, decreased expression levels of miR-146a in GBM have
been associated with the regulation of cell proliferation and
apoptosis via the targeting of notch 1 (39). In addition, upregulated miR-622 has
been observed to inhibit cell proliferation, motility and invasion
by repressing KRAS proto-oncogene GTPase in GBM (40). Additionally, miR-10b in GBM may
mediate TGF-β1-regulated cell proliferation, migration and EMT
(41); however, to the best of our
knowledge, the roles of miR-454-3p, which is located on chromosome
17q22, in the progression of GBM, particularly its effects on cell
proliferation and migration and target gene regulation, have not
yet been studied.
In the present study, downregulation of miR-454-3p
in glioma tissues and GBM cell lines was observed. Functional
experiments further revealed that the overexpression of miR-454-3p
may inhibit the proliferation, migration and invasion of the LN-229
GBM cell line. Recent studies have reported that miR-454 may act as
an oncogene or tumor suppressor in cancer (17,42).
For instance, miR-454-3p was reported to be upregulated and to
function as an oncogene in hepatocellular carcinoma, non-small cell
lung cancer (11) and colorectal
cancer (12); however, the present
study revealed that miR-454-3p served a tumor suppressing role in
GBM LN-229 cells. Thus, miR-454-3p may serve a variety of roles in
different types of tumor. To the best of our knowledge, there have
been no reports regarding the association between miR-454-3p and
CPEB1. The results of the present study indicated that miR-454-3p
negatively regulated CPEB1, and identified CPEB1 as a direct target
of miR-454-3p. Therefore, the findings of the present study
collectively indicated that miR-454-3p may be an important miRNA
that is potentially associated with the proliferation, migration
and invasion of GBM cells.
Metastasis is one of the principal factors that
contributes to the mortality of patients with GBM (43). The present study reported that the
overexpression of miR-454-3p inhibited the migration and invasion
of GBM cells in vitro. In epithelial cancers, EMT is
regarded as one of the major mechanisms that promote cell
migration, invasion and metastasis. A recent study demonstrated
that CPEB1 mediates EMT and metastasis in breast cancer (22). In the present study, miR-454-3p was
suggested to inhibit the migration and invasion of GBM cells by
negatively regulating CPEB1. In addition, MMPs are proteolytic
enzymes that serve a pivotal role in the transformation and
progression of tumors at all stages, particularly invasion and
metastasis (44). The present
study revealed that within GBM cells, miR-454-3p significantly
inhibited the expression of MMP-9. Collectively, these results
indicated that the upregulation of miR-454-3p in GBM cells
suppressed the migration and invasion of the tumor cells by
inhibiting CPEB1 and MMP-9 expression.
In conclusion, the present study demonstrated that
miR-454-3p was significantly downregulated in GBM cell lines, and
that miR-454-3p overexpression suppressed cell proliferation,
migration and invasion, potentially by targeting CPEB1 in the GBM
cells. These findings indicated a novel tumor suppressive role of
miR-454 in the development of GBM. Furthermore, miR-454-3p and
CPEB1 may be potential therapeutic targets in the treatment of
GBM.
Acknowledgements
The authors of the present study would like to thank
the Chinese Academy of Sciences (Shanghai, China) for providing the
cell lines and Shanghai GenePharma Co., Ltd. (Shanghai, China) for
their support with cell transfection.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH and SZ searched the literature and designed the
study. XH and YW performed the experiments and interpreted the
data. YW contributed to the materials. XH wrote and revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Huai'an First People's Hospital. All tissue
samples were collected for used after obtaining informed consent
from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics,2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Furnari FB, Fenton T, Bachoo RM, Mukasa A,
Stommel JM, Stegh A, Hahn WC, Ligon KL, Louis DN, Brennan C, et al:
Malignant astrocytic glioma: Genetics, biology, and paths to
treatment. Genes Dev. 21:2683–2710. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meyer MA: Malignant gliomas in adults. N
Engl J Med. 359:18502008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fisher JL, Schwartzbaum JA, Wrensch M and
Wiemels JL: Epidemiology of brain tumors. Neurol Clin. 25:867–890.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yao C, Lv S, Han M, Zhang J, Zhang Y,
Zhang L, Yi R, Zhuang D and Wu J: The association of Crk-like
adapter protein with poor prognosis in glioma patients. Tumor Biol.
35:5695–5700. 2014. View Article : Google Scholar
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Irwandi RA and Vacharaksa A: The role of
microRNA in periodontal tissue: A review of the literature. Arch
Oral Biol. 72:66–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fan Y, Shi C, Li T and Kuang T:
microRNA-454 shows anti-angiogenic and anti-metastatic activity in
pancreatic ductal adenocarcinoma by targeting LRP6. Am J Cancer
Res. 7:139–147. 2017.PubMed/NCBI
|
|
10
|
Cao ZG, Li JJ, Yao L, Huang YN, Liu YR, Hu
X, Song CG and Shao ZM: High expression of microRNA-454 is
associated with poor prognosis in triple-negative breast cancer.
Oncotarget. 7:64900–64909. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu DY, Li XN, Qi Y, Liu DL, Yang Y, Zhao
J, Zhang CY, Wu K and Zhao S: MiR-454 promotes the progression of
human non-small cell lung cancer and directly targets PTEN. Biomed
Pharmacother. 81:79–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liang HL, Hu AP, Li SL, Xie JP, Ma QZ and
Liu JY: miR-454 prompts cell proliferation of human colorectal
cancer cells by repressing CYLD expression. Asian Pac J Cancer
Prev. 16:2397–2402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fan Y, Xu LL, Shi CY, Wei W, Wang DS and
Cai DF: MicroRNA-454 regulates stromal cell derived factor-1 in the
control of the growth of pancreatic ductal adenocarcinoma. Sci Rep.
6:227932016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Niu G, Li B, Sun J and Sun L: miR-454 is
down-regulated in osteosarcomas and suppresses cell proliferation
and invasion by directly targeting c-Met. Cell Prolif. 48:348–355.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu L, Gong X, Sun L, Yao H, Lu B and Zhu
L: miR-454 functions as an oncogene by inhibiting CHD5 in
hepatocellular carcinoma. Oncotarget. 6:39225–39234. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun L, Wang Q, Gao X, Shi D, Mi S and Han
Q: MicroRNA-454 functions as an oncogene by regulating PTEN in
uveal melanoma. FEBS Lett. 589:2791–2796. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fang B, Zhu J, Wang Y, Geng F and Li G:
MiR-454 inhibited cell proliferation of human glioblastoma cells by
suppressing PDK1 expression. Biomed Pharmacother. 75:148–152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Richter JD: CPEB: A life in translation.
Trends Biochem Sci. 32:279–285. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Groisman I, Jung MY, Sarkissian M, Cao Q
and Richter JD: Translational control of the embryonic cell cycle.
Cell. 109:473–483. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ivshina M, Lasko P and Richter JD:
Cytoplasmic polyadenylation element binding proteins in
development, health, and disease. Annu Rev Cell Dev Biol.
30:393–415. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Udagawa T, Farny NG, Jakovcevski M,
Kaphzan H, Alarcon JM, Anilkumar S, Ivshina M, Hurt JA, Nagaoka K,
Nalavadi VC, et al: Genetic and acute CPEB1 depletion ameliorate
fragile X pathophysiology. Nat Med. 19:1473–1477. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagaoka K, Fujii K, Zhang H, Usuda K,
Watanabe G, Ivshina M and Richter JD: CPEB1 mediates
epithelial-to-mesenchyme transition and breast cancer metastasis.
Oncogene. 35:2893–2901. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shoshan E, Mobley AK, Braeuer RR, Kamiya
T, Huang L, Vasquez ME, Salameh A, Lee HJ, Kim SJ, Ivan C, et al:
Reduced adenosine-to-inosine miR-455-5p editing promotes melanoma
growth and metastasis. Nat Cell Biol. 17:311–321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiaoping L, Zhibin Y, Wenjuan L, Zeyou W,
Gang X, Zhaohui L, Ying Z, Minghua W and Guiyuan L: CPEB1, a
histone-modified hypomethylated gene, is regulated by miR-101 and
involved in cell senescence in glioma. Cell Death Dis. 4:e6752013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C((T) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang L, Dong Y, Zhu N, Tsoi H, Zhao Z, Wu
CW, Wang K, Zheng S, Ng SS, Chan FK, et al: microRNA-139-5p exerts
tumor suppressor function by targeting NOTCH1 in colorectal cancer.
Mol Cancer. 13:1242014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sttg PS: New insights into the tumor
metastatic process revealed by gene expression profiling. Am J
Pathol. 166:1291–1294. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Du Y, Jiang B, Song S, Pei G, Ni X, Wu J,
Wang S, Wang Z and Yu J: Metadherin regulates actin cytoskeletal
remodeling and enhances human gastric cancer metastasis via
epithelial-mesenchymal transition. Int J Oncol. 51:63–74. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tong L, Chu M, Yan B, Zhao W, Liu S, Wei
W, Lou H, Zhang S, Ma S, Xu J and Wei L: MTDH promotes glioma
invasion through regulating miR-130b-ceRNAs. Oncotarget.
8:17738–17749. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hou Y, Yu L, Mi Y, Zhang J, Wang K and Hu
L: Association of MTDH immunohistochemical expression with
metastasis and prognosis in female reproduction malignancies: A
systematic review and meta-analysis. Sci Rep. 6:383652016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang J, Chen X and Tong M: Knockdown of
astrocyte elevated gene-1 inhibited cell growth and induced
apoptosis and suppressed invasion in ovarian cancer cells. Gene.
616:8–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu X, Lv Z, Zou J, Liu X, Ma J, Sun C, Sa
N and Xu W: Elevated AEG-1 expression in macrophages promotes
hypopharyngeal cancer invasion through the STAT3-MMP-9 signaling
pathway. Oncotarget. 7:77244–77256. 2016.PubMed/NCBI
|
|
33
|
Skerenova M, Mikulova V, Capoun O, Zima T
and Tesarova P: Circulating tumor cells and serum levels of MMP-2,
MMP-9 and VEGF as markers of the metastatic process in patients
with high risk of metastatic progression. Biomed Pap Med Fac Univ
Palacky Olomouc Czech Repub. 161:272–280. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shin SS, Song JH, Hwang B, Noh DH, Park
SL, Kim WT, Park SS, Kim WJ and Moon SK: HSPA6 augments garlic
extract-induced inhibition of proliferation, migration, and
invasion of bladder cancer EJ cells; Implication for cell cycle
dysregulation, signaling pathway alteration, and transcription
factor-associated MMP-9 regulation. PLoS One. 12:e01718602017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Young RM, Jamshidi A, Davis G and Sherman
JH: Current trends in the surgical management and treatment of
adult glioblastoma. Ann Transl Med. 3:1212015.PubMed/NCBI
|
|
36
|
Krishnatry R, Zhukova N, Guerreiro
Stucklin AS, Pole JD, Mistry M, Fried I, Ramaswamy V, Bartels U,
Huang A, Laperriere N, et al: Clinical and treatment factors
determining long-term outcomes for adult survivors of childhood
low-grade glioma: A population-based study. Cancer. 122:1261–1269.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang X, Zhang X, Hu S, Zheng M, Zhang J,
Zhao J, Zhang X, Yan B, Jia L, Zhao J, et al: Identification of
miRNA-7 by genome-wide analysis as a critical sensitizer for
TRAIL-induced apoptosis in glioblastoma cells. Nucleic Acids Res.
45:5930–5944. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xiuju C, Zhen W and Yanchao S: SOX7
inhibits tumor progression of glioblastoma and is regulated by
miRNA-24. Open Med (Wars). 11:133–137. 2016.PubMed/NCBI
|
|
39
|
Hu HQ, Sun LG and Guo WJ: Decreased
miRNA-146a in glioblastoma multiforme and regulation of cell
proliferation and apoptosis by target notch1. Int J Biol Markers.
31:e270–275. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang X, Xin Z, Xu Y and Ma J: Upregulated
miRNA-622 inhibited cell proliferation, motility, and invasion via
repressing kirsten rat sarcoma in glioblastoma. Tumour Biol.
37:5963–5970. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ma C, Wei F, Xia H, Liu H, Dong X, Zhang
Y, Luo Q, Liu Y and Li Y: MicroRNA-10b mediates TGF-β1-regulated
glioblastoma proliferation, migration and epithelial-mesenchymal
transition. Int J Oncol. 50:1739–1748. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou L, Qu YM, Zhao XM and Yue ZD:
Involvement of miR-454 overexpression in the poor prognosis of
hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 20:825–829.
2016.PubMed/NCBI
|
|
43
|
Fidler IJ: The pathogenesis of cancer
metastasis the: Seed and soil' hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tabouret E, Boudouresque F, Farina P,
Barrié M, Bequet C, Sanson M and Chinot O: MMP2 and MMP9 as
candidate biomarkers to monitor bevacizumab therapy in highgrade
glioma. Neuro Oncol. 17:1174–1176. 2015. View Article : Google Scholar : PubMed/NCBI
|