Introduction
Prostate cancer (PCa) is one of the most common
types of cancer affecting male health (1). In recent years, the incidence of PCa
in China has been increasing annually and the age of onset is
becoming younger. This phenomenon may be accounted for by
lifestyle, longevity, aging and diagnostic technology (2,3).
Local invasion or distant metastasis may occur once the lesion
develops to the later stages (4).
The survival rate of patients with localized PCa is ~100%, whereas,
the annual survival rate of patients with distant metastasis is
reduced to ~29% (5). Therefore, it
is necessary to identify a treatment to inhibit the growth and
metastasis of PCa.
For decades, isolated products from plants served a
pivotal role in the treatment of malignant tumors (6,7).
Zeylenone (Zey), a naturally occurring cyclohexene oxide, is
isolated from Uvaria grandiflora Roxb (8). Though Zey exhibits strong toxicity to
tumor cells, it is less toxic to normal cells (8,9).
Previous studies have demonstrated that Zey exhibits potent
activity against tumors, for example, cervical carcinoma and lung
cancer (8,10). In cervical carcinoma, a previous
study demonstrated that Zey was able to induce the apoptosis of
cervical carcinoma cells (8).
Another previous study on lung cancer tumor-bearing mice observed
that the Zey-loaded mice demonstrated an antitumor effect (10). A previous study has reported that
Zey had inhibitory effect on proliferation of PC-3 cells (11). However, the role of Zey in PCa and
its underlying mechanism requires further investigations.
The extracellular matrix (ECM) and
epithelial-mesenchymal transition (EMT) are closely pertinent to
tumor metastasis (12,13). The ECM is primarily composed of
interstitial collagen, laminin, elastin and fibronectin. In the
dynamic balance of metabolic renewal, degradation and remodeling,
it maintains the microenvironment of tumor cell growth and
regulates the gene expression of tumor cells in contact with it,
thereby affecting the metabolism, growth and metastasis of tumors
(12,14). The occurrence of EMT is a dynamic
and multi-step process, which includes the loss of intercellular
adhesion, destruction of the basement membrane and ECM, and
remodeling of cytoskeleton, ultimately resulting in enhanced
motility and migration of tumors (15).
The Wnt/β-catenin signaling pathway is highly
conserved in biological evolution and its members are highly
homologous from Drosophila melanogaster to higher mammals.
The pathway regulates the stability of transcription factor
β-catenin and is dependent on the expression of the β-catenin gene
(16,17). The Wnt/β-catenin signal
transduction pathway is associated with human cancer. Therefore,
previous studies of the pathway not only aided understanding of the
mechanism of cancer; however, additionally suggested a series of
novel targets for the treatment of cancer (18–20).
In the preset study, the effect of Zey on the
viability and metastasis of human PCa cell line DU145 was
investigated. The present study additionally aimed to examine
whether potential mechanisms are regulated by the ECM, EMT and
Wnt/β-catenin pathways. The present study demonstrated the role of
Zey on human PCa and its possible mechanism of action. The present
study may provide a novel candidate anti-tumor agent in the
treatment of PCa.
Materials and methods
Preparation of Zey
Zey was obtained from Yuanye Bio-technology Co.,
Ltd. (Shanghai, China) with a purity of 98%. Zey was prepared for
subsequent experiments as described previously (9) and was diluted to the desired
concentrations (2.5, 5, 10, 20 and 40 µmol/l).
Cell culture
The human PCa cell line DU145 (American Type Culture
Collection, Manassas, VA, USA) was cultured in Eagle's Minimum
Essential Medium (American Type Culture Collection) with 10% fetal
bovine serum (FBS; American Type Culture Collection) and 1%
penicillin/streptomycin (Thermo Fisher Scientific, Inc., Waltham,
MA, USA) in a 37°C incubator (Thermo Fisher Scientific, Inc.) with
95% humidity and 5% CO2.
Cell Counting Kit-8 (CCK-8) assay
Cells were inoculated in 96-well plates
(2.5×103 cells/well) and cultured in an incubator for 24
h. Subsequent to being cultured, cells were exposed to different
concentrations of Zey at 0, 2.5, 5, 10, 20 and 40 µmol/l for 12, 24
and 48 h. CCK-8 solution (Beyotime Institute of Biotechnology,
Haimen, China) was added to the cells and the plate was transferred
to the incubator for 4 h. Finally, absorbance was measured at 450
nm with a FilterMax F3/F5 microplate reader (Molecular Devices,
LLC, Sunnyvale, CA, USA).
Matrigel assay
Cells were inoculated in 6-well plates
(2.5×104 cells/well) and subsequently cultured in an
incubator for 24 h. Cells were treated with different
concentrations of Zey for 48 h at 10 (Zey1), 20 (Zey2) and 40
(Zey3) µmol/l. The untreated cells served as a control. BD matrigel
(Beijing Solarbio Science & Technology Co., Ltd., Shanghai,
China) was filled in the upper chamber of the transwell plates at
room temperature for 25 min. Subsequently, the transwell membrane
was placed in the culture plate. F-12K medium was added into the
upper chamber for 20 min and subsequently removed. F-12K medium
with 15% FBS was added into the lower chamber for attracting cells.
Subsequently, the cell suspension was cultured in the upper chamber
at 37°C for 24 h. Cells were stained with 4 g/l crystal violet
(Tianjin Zhongxin Chemtech, Co., Ltd., Tianjin, China) for 15 min
at room temperature and washed with PBS three times. Finally, cells
were observed and images were captured using a light microscope
(magnification, ×400; Nikon Corporation, Tokyo, Japan). The number
of invasive cells was quantified using GraphPad Prism software 6.0
(GraphPad Software, Inc., La Jolla, CA, USA).
Wound healing assay
Cells were inoculated in 6-well plates
(2.5×104 cells/well) and cultured in an incubator for 24
h. Cells were scratched with a 200-µl pipette tip and washed with
medium three times. The cells were treated with different
concentrations of Zey at 10 (Zey1), 20 (Zey2) and 40 (Zey3) µmol/l.
Untreated cells served as a control. The cells were observed and
imaged prior to and at 12 and 24 h post-wound assay under a light
microscope (magnification, ×400). Wound width was quantified using
GraphPad Prism software 6.0.
ELISA
Cells were inoculated in 6-well plates
(2.5×104 cell/well) and cultured in an incubator for 24
h. Cells were treated with different concentrations of Zey for 48 h
at 10 (Zey1), 20 (Zey2) and 40 (Zey3) µmol/l. Untreated cells
served as a control. Subsequently, cells were digested with 0.25%
EDTA-trypsin (Beijing Solarbio Science & Technology Co., Ltd.)
and centrifuged at 800 × g for 5 min at 4°C. Following
centrifugation, cells were resuspended in F-12K medium. The
expression of MMP2/9 was detected using ELISA test kits (cat. nos.
MMP200 and DMP900, respectively; R&D Systems, Inc.,
Minneapolis, MN, USA). The standard curve was plotted with standard
samples. The assay diluent (50 µl) was added to each well. In
total, 50 µl sample in each group was added into each well. The
plates was covered with plate sealer and maintained for 2 h at room
temperature. Following washing, the conjugate reagent was added
into each well and the plate was placed on a shaker at room
temperature for 2 h. The substrate solution was added into each
well and maintained for 30 min at room temperature. Subsequently,
the stop solution was used to terminate the reaction. Finally, the
absorbance at 450 nm was read using a FilterMax F3/F5 microplate
reader.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) was applied to collect
total RNA. A total of 1 µg RNA was used to synthesize cDNA using an
RT Master Mix kit (Takara Biotechnology Co., Ltd., Dalian, China).
Reverse transcriptional reaction conditions were set at 85°C for 15
min. SYBR Premix Taq™ II kit (Takara Biotechnology Co.,
Ltd.) was used to amplify cDNA. The volumes of the amplified
reagent were as follows: 25 µl SYBR Green Mix; 1 µl forward/reverse
primers; 4 µl cDNA; and nuclease-free H2O to a total
volume of 50 µl. The temperature and duration of the amplification
were as follows: 92°C for 15 min, (at 92°C for 20 sec at 65°C for
45 sec) at 35 cycles, at 85°C for 20 sec, at 37°C for 2 min. All
primer sequences are listed in Table
I. β-actin was regarded as the internal control. The formula
2−ΔΔCq (21) was
performed to determine the gene expression levels.
 | Table I.Sequences of the primers. |
Table I.
Sequences of the primers.
| Primer name | Sequence,
5′-3′ | Product size,
bp |
|---|
| MMP-2-Forward |
CAGCCCTGCAAGTTTCCATT |
|
| MMP-2-Reverse |
GTTGCCCAGGAAAGTGAAGG | 210 |
| MMP-9-Forward |
GAGACTCTACACCCAGGACG |
|
| MMP-9-Reverse |
GAAAGTGAAGGGGAAGACGC | 238 |
| FN-1-Forward |
TGGCACTGATGAAGAACCCT |
|
| FN-1-Reverse |
GGGAAACTGTGTAGGGGTCA | 224 |
| TIMP-1-Forward |
AGACCACCTTATACCAGCGT |
|
| TIMP-1-Reverse |
GCCACAAAACTGCAGGTAGT | 217 |
|
Collagen-1-Forward |
CATGCCGTGACCTCAAGATG |
|
|
Collagen-1-Reverse |
TCCATCGGTCATGCTCTCTC | 227 |
|
Vimentin-Forward |
AATAAGATCCTGCTGGCCGA |
|
|
Vimentin-Reverse |
GGTGTTTTCGGCTTCCTCTC | 225 |
|
E-cadherin-Forward |
ACGCATTGCCACATACACTC |
|
|
E-cadherin-Reverse |
GGTGTTCACATCATCGTC | 217 |
|
β-actin-Forward |
GGGAAATCGTGCGTGACATT |
|
|
β-actin-Reverse |
AGGTAGTTTCGTGGATGCCA | 219 |
Western blotting assay
Cells were lysed with radioimmunoprecipitation assay
buffer (Beijing Solarbio Science & Technology Co., Ltd.). The
content of protein was evaluated by a bicinchoninic protein
quantification kit (Yeasen). Protein samples (25 µg per lane) were
separated with 10% SDS-PAGE and subsequently transferred to a
polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA,
USA). Subsequently, the membrane was blocked with 5% skimmed milk
powder at room temperature for 1.5 h. Following blocking, the
membrane was incubated with anti-matrix metalloproteinase (MMP)-2
(R&D Systems, Inc.; cat. no. IC903G-100UG; 1:1,000), anti-MMP-9
(Abcam, Cambridge, UK; cat. no. EP1254; 1:500), anti-tissue
inhibitor of metalloproteinases-1 (TIMP-1; Abcam; cat. no. ab61224;
1:700), anti-vimentin (R&D Systems, Inc.; cat. no. AF2105;
1:700), anti-epithelial (E)-cadherin (R&D Systems, Inc.; cat.
no. MAB1838; 1:1,000), anti-fibronectin 1 (FN1; Abcam; cat. no.
ab32419; 1:800), anti-collagen-1 (Abcam; cat. no. ab90395; 1:600),
anti-wnt5a (Abcam; cat. no. ab174963; 1:1,000), anti-β-catenin
(R&D Systems, Inc.; cat. no. AF1329; 1:1,000), anti-cyclin D1
(R&D Systems, Inc.; cat. no. MAB4314; 1:900) and anti-β-actin
(Abcam; cat. no. ab13772; 1:800) on the rocking table at 4°C for 24
h. The membrane was subsequently incubated in corresponding
horseradish peroxidase-conjugated secondary antibodies [rabbit
anti-mouse immunoglobulin (Ig)G; Cell Signaling Technology, Inc.,
Danvers, MA, USA; cat. no. 58802; 1:8,000; mouse anti-rabbit IgG;
Cell Signaling Technology, Inc.; cat. no. 5127; 1:7,000; goat
anti-mouse IgG; Abcam; cat. no. ab6785; 1:7,000] at 37°C for 1 h.
The protein was visualized using an enhanced chemiluminescence
detection kit (Beyotime Institute of Biotechnology). β-actin was
used as an internal control. Densitometry was conducted using
Quantity One software 4.21 (Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
Statistical analysis
Data was analyzed using GraphPad Prism software 6.0
(GraphPad Software, Inc., La Jolla, CA, USA). The data are
presented as the mean ± standard deviation from at least three
independent experiments. The differences between groups were
assessed by one-way analysis of variance followed by Turkey's text.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Zey represses the viability of DU145
cells
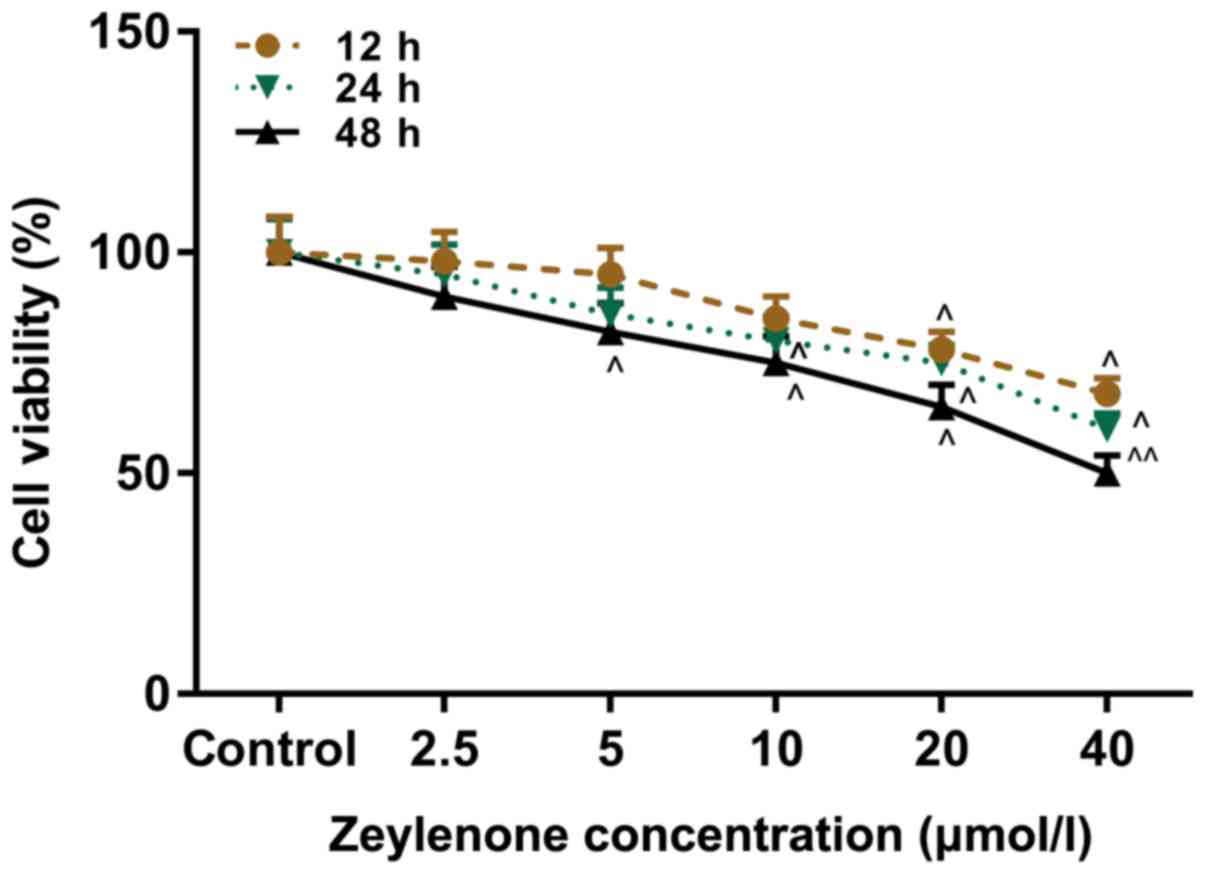
A CCK-8 assay was performed to investigate the
effect of Zey on the viability of DU145 cells. Cells were treated
with Zey at 0, 2.5, 5, 10, 20 and 40 µmol/l for 12, 24 and 48 h.
The data from the present study demonstrated that cell viability
decreased with increasing concentrations of Zey and incubation
time. The half maximal inhibitory concentration of Zey was 40
µmol/l at 48 h. The treatment with Zey at 10, 20 and 40 µmol/l for
48 h caused a significant decrease in the viability of the cells
(P<0.05; Fig. 1). Therefore,
these concentrations and time were selected as the conditions of
the subsequent experiments.
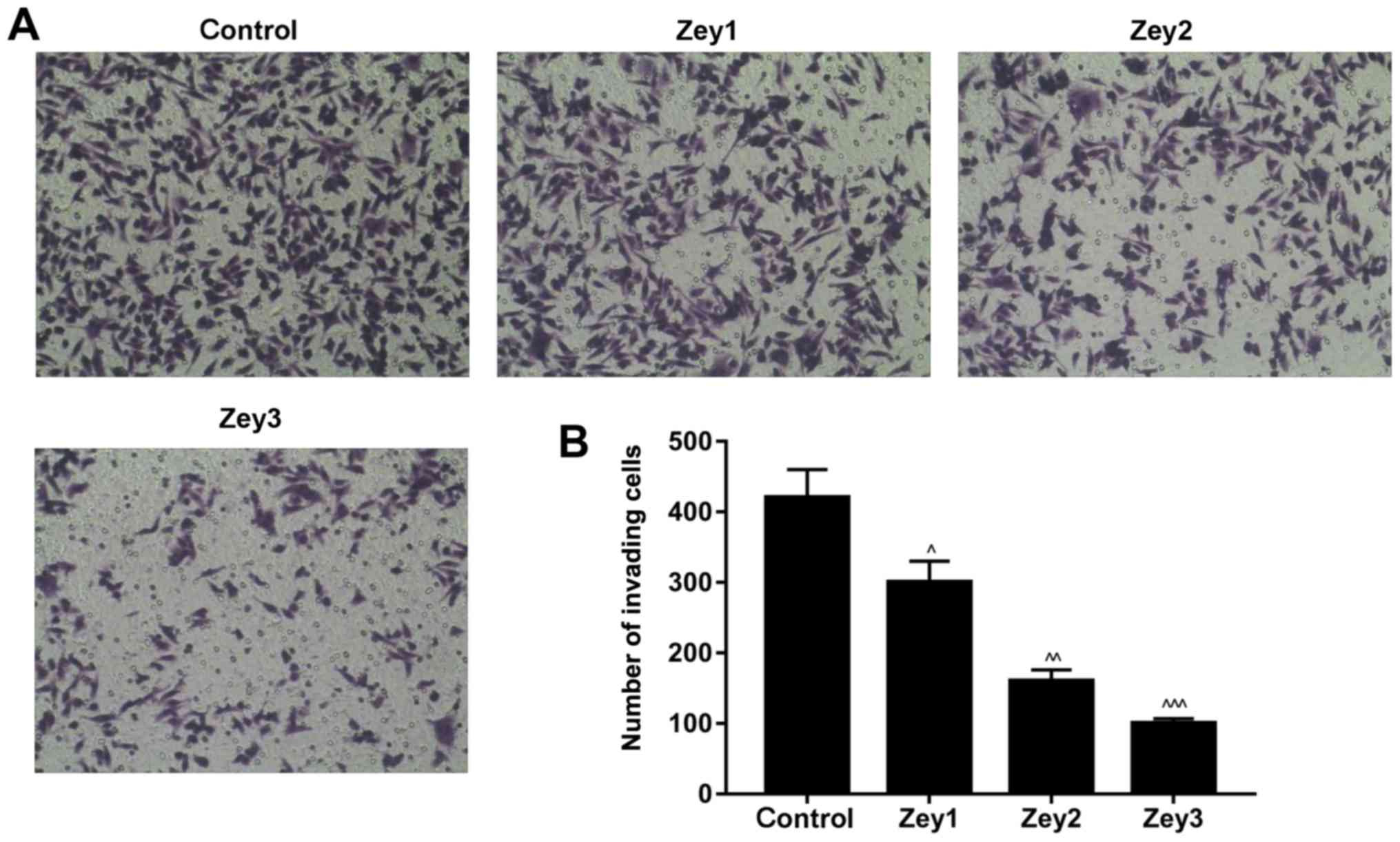
Zey inhibits the invasive ability of
DU145 cells
The Matrigel assay was conducted to study the
invasive ability of Zey-treated DU145 cells. Cells were treated
with Zey at 0 (control), 10 (Zey1), 20 (Zey2) and 40 (Zey3) µmol/l
for 48 h. The results demonstrated that treatment with Zey
decreased the number of invading cells in a dose-dependent manner,
compared with the control group. The proportion of invasive cells
in the Zey1 group was ~67%; whereas, in the Zey2 and Zey3 groups
the proportions were ~50 and ~30%, respectively (P<0.05;
Fig. 2).
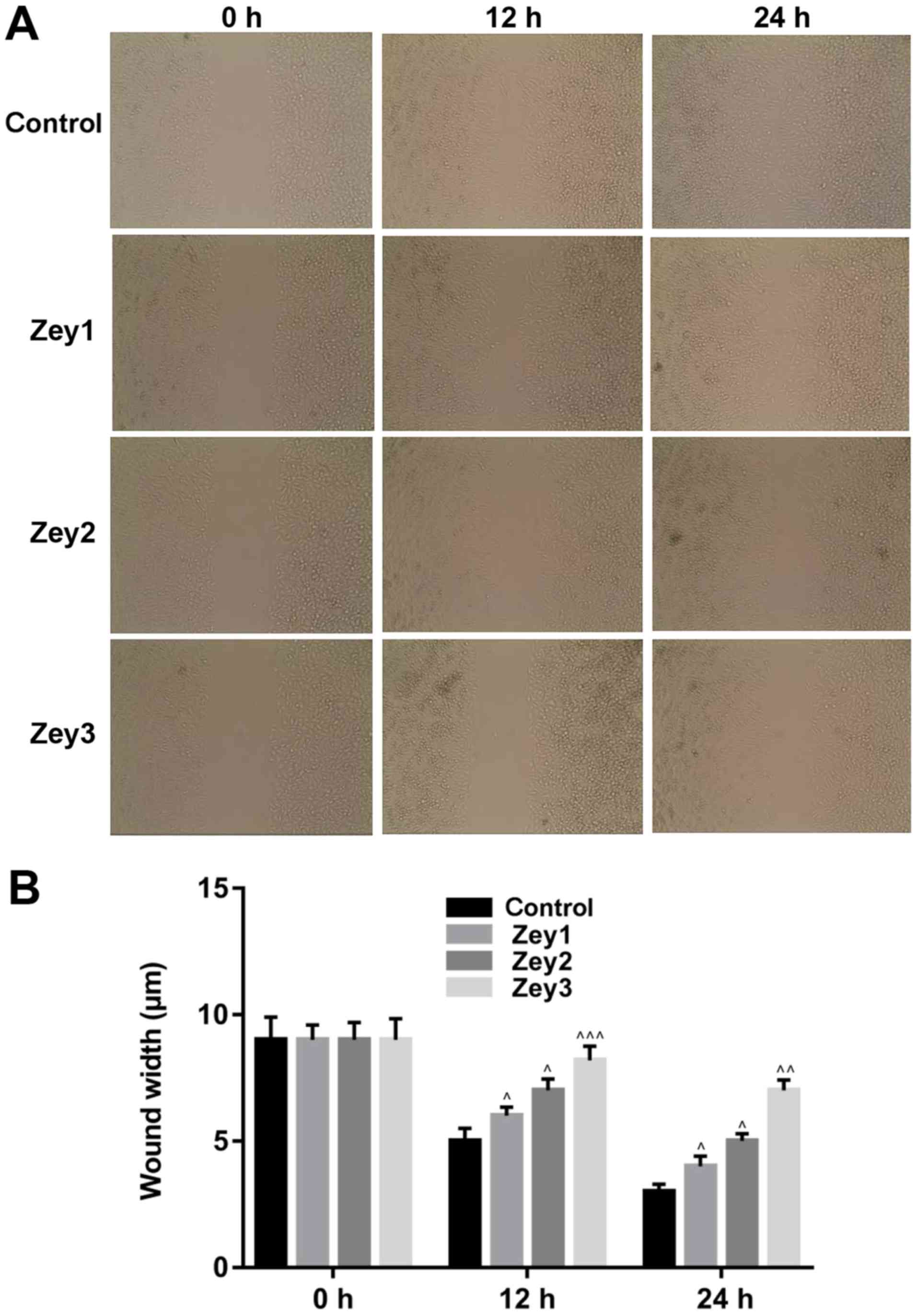
Zey decreases the migratory ability of
DU145 cells
A wound-healing assay was conducted to analyze the
effect of Zey on the migratory ability of DU145 cells. Cells were
treated with Zey at 0 (control), 10 (Zey1), 20 (Zey2) and 40 (Zey3)
µmol/l for 0, 12 and 24 h. No obvious migration of cells was
demonstrated when cells were exposed to 0, 10, 20 and 40 µmol/l Zey
for 0 h. However, the migratory ability of cells was significantly
attenuated when cells were treated with 10, 20 and 40 µmol/l Zey
for 12 and 24 h (P<0.05; Fig.
3).
Zey regulates ECM-associated factors
in DU145 cells
To examine the expression levels of ECM-associated
factors, ELISA, RT-qPCR and western blotting assays were performed.
Cells were treated with Zey at 0 (control), 10 (Zey1), 20 (Zey2)
and 40 (Zey3) µmol/l for 48 h. The ELISA data identified that Zey
significantly decreased the expression levels of MMP-2 and MMP-9 in
a concentration-dependent manner (P<0.05; Fig. 4A). In addition, the results of
RT-qPCR demonstrated that when cells were exposed to Zey (10, 20
and 40 µmol/l), the mRNA expression levels of MMP-2, MMP-9 and FN-1
significantly decreased, whereas, the expression levels of TIMP-1
and collagen-1 significantly increased (P<0.05; Fig. 4B). As demonstrated by the western
blotting assays, the protein expressions were similar to that of
mRNA (P<0.05; Fig. 4C).
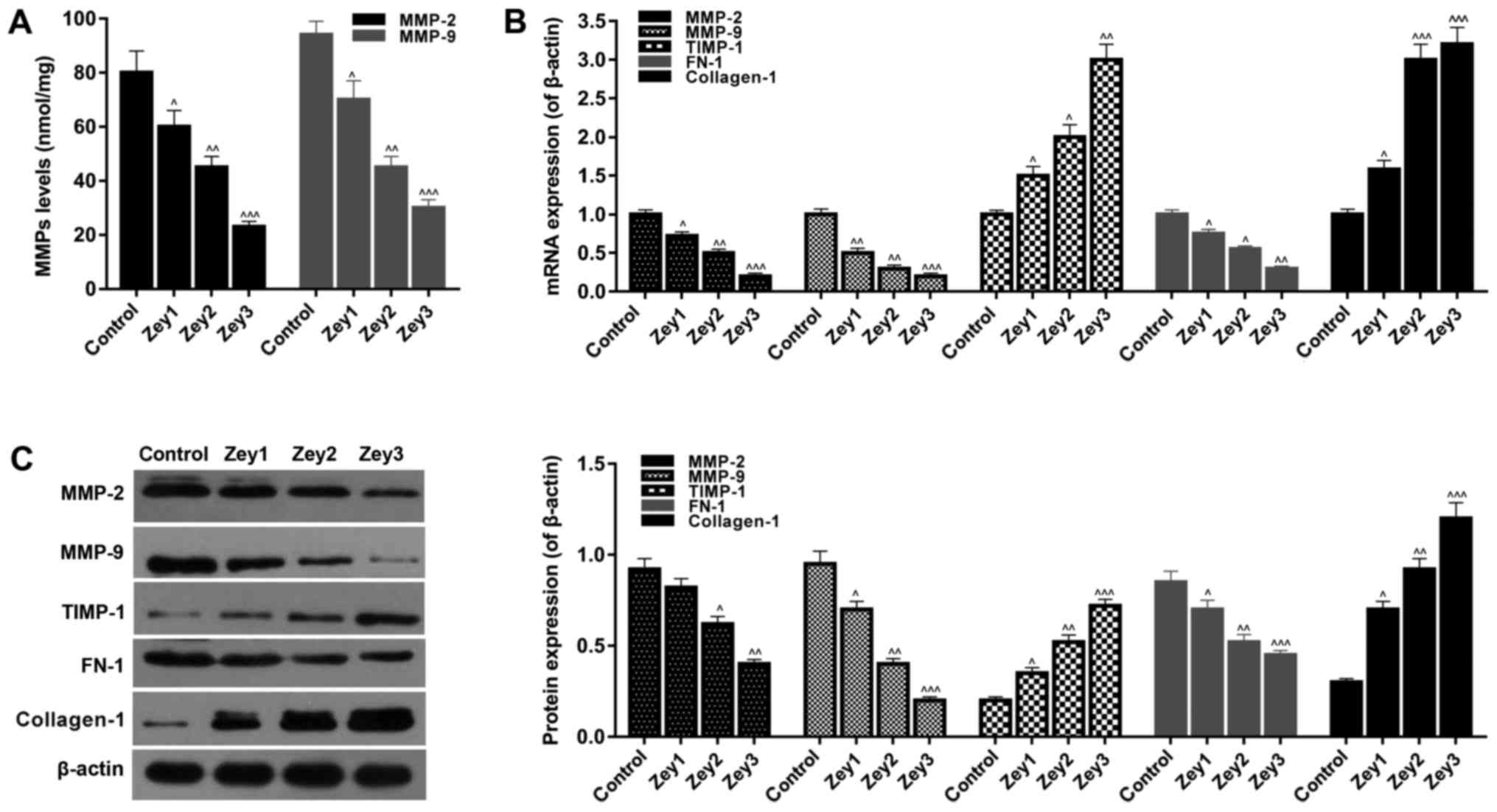 | Figure 4.Zey regulates extracellular
matrix-associated factors in DU145 cells. (A) DU145 cells exposed
to Zey at 0 (control), 10 (Zey1), 20 (Zey2) and 40 (Zey3) µmol/l
for 48 h. Expression levels of MMP-2 and MMP-9 were detected using
an ELISA. (B) mRNA expressions of MMP-2, MMP-9, TIMP-1, FN-1 and
collagen-1 were measured by a reverse transcription-quantitative
polymerase chain reaction assay. (C) Protein expression of MMP-2,
MMP-9, TIMP-1, FN-1 and collagen-1 were measured by western blot
analysis and normalized to β-actin expression. Gray value was
evaluated and quantified using Quantity One software.
^P<0.05, ^^P<0.01,
^^^P<0.001 vs. respective control. Zey, Zeylenone;
MMP, matrix metalloproteinase; TIMP, tissue inhibitor of
metalloproteinases; FN-1, fibronectin. |
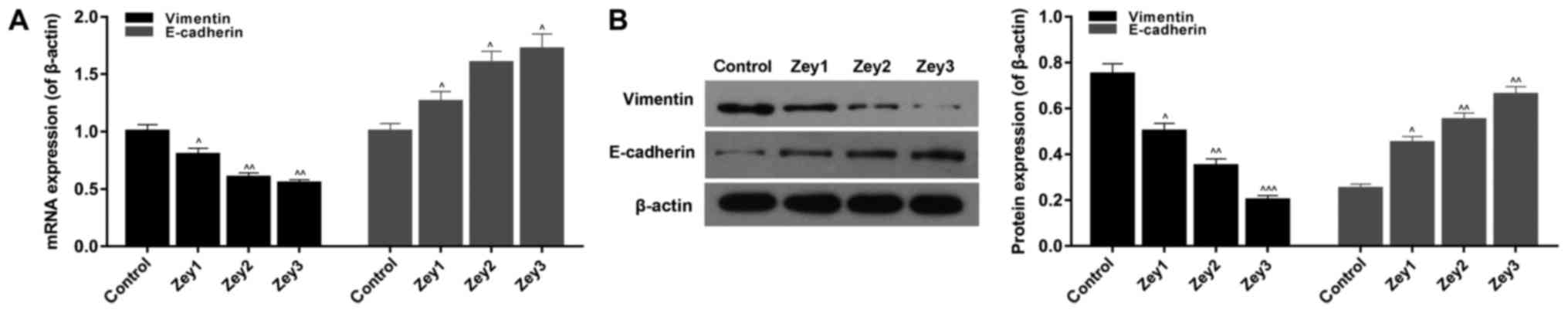
Zey regulates EMT-associated factors
in DU145 cells
To investigate the expression levels of
EMT-associated factors, RT-qPCR and western blot assays were
performed. Cells were treated with Zey at 0 (control), 10 (Zey1),
20 (Zey2) and 40 (Zey3) µmol/l for 48 h. The RT-qPCR results
demonstrated that Zey significantly decreased the mRNA expression
level of vimentin; however, increased the E-cadherin mRNA
expression level in a dose-dependent manner (P<0.05; Fig. 5A). In addition, the western
blotting data revealed that the protein expression of vimentin was
significantly downregulated; whereas, the protein expression level
of E-cadherin was upregulated in Zey-treated cells (P<0.05;
Fig. 5).
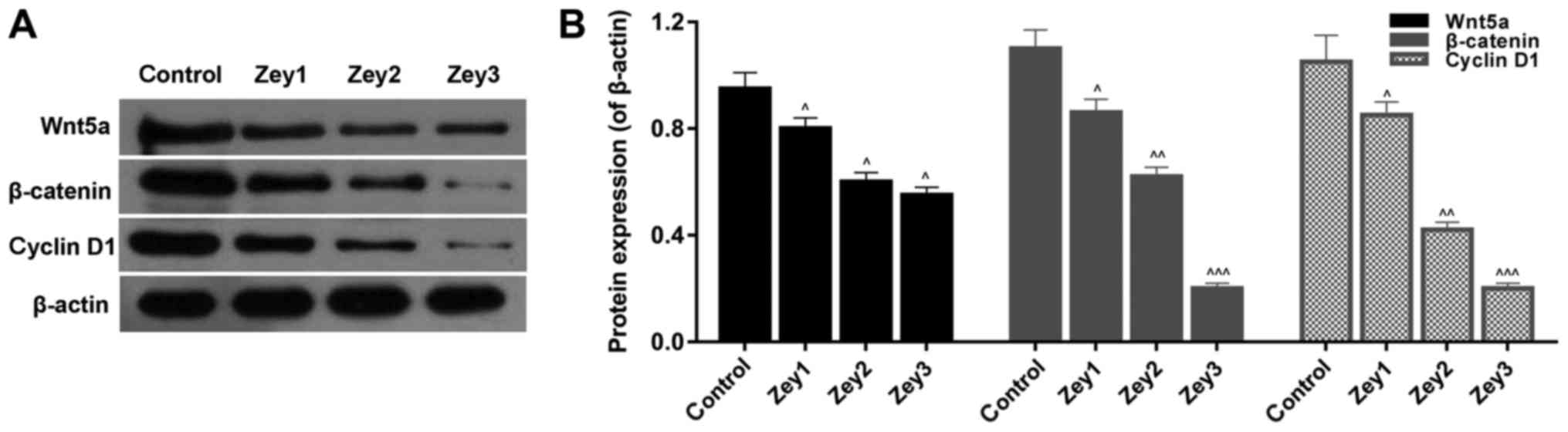
Zey suppresses the Wnt/β-catenin
signaling pathway in DU145 cells
To further analyze whether Zey suppressed growth and
metastasis by regulating the Wnt/β-catenin pathway, a western
blotting assay was used to determine the protein expression levels
of wnt5a, β-catenin and cyclin D1. Cells were subjected to Zey at 0
(control), 10 (Zey1), 20 (Zey2) and 40 (Zey3) µmol/l for 48 h. The
data demonstrated that the expressions of wnt5a, β-catenin and
cyclin D1 were significantly decreased in cells treated with Zey
(P<0.05; Fig. 6).
Discussion
PCa is one of the most common tumors of the urinary
system in men. The continuous metastasis and proliferation of PCa
cells are important clinical features and a principal cause of
mortality in the advanced stage of this cancer (22,23).
It was demonstrated that Zey demonstrates cytotoxicity to tumor
cells (10,24). Similar to previous studies
(8,9), the present study demonstrated that
Zey decreased the viability of DU145 cells. Previous studies
demonstrated that Zey markedly suppresses growth and promotes
apoptosis in cervical tumor cells (8,9).
However, to the best of our knowledge, no study has investigated
the role of Zey in PCa. In the present study, the effect of Zey on
the metastasis of DU145 cells was analyzed. The results revealed
that Zey suppressed the invasive and migratory capabilities of
DU145 cells.
Subsequently, the molecular mechanism of Zey
inhibiting the proliferation and metastasis of PCa was further
examined. It is widely accepted that the ECM may regulate the rate
and direction of cell metastasis and provide ‘scaffolding’ for cell
metastasis. By affecting the balance of ECM degradation, MMPs
achieve a pathological and physiological significance (14). In addition, FN and collagen are the
primary components of the ECM. Their deregulated expression will
result in increased invasiveness of breast cancer cells (25). Furthermore, TIMP inhibits basement
membrane degradation, endothelial cell formation and angiogenesis
by depressing the function of MMPs, and in this way, TIMP affects
the invasion and migration progress of a tumor (26). The results of the present study
demonstrated that Zey decreased the expression levels of MMP-2,
MMP-9 and FN1, and promoted TIMP-1 and collagen-1 expression.
Therefore, the present study demonstrated that ECM components
contributed to Zey-inhibited metastasis in PCa cells.
Furthermore, EMT is dysregulated in various tumors,
which affects tumor proliferation and metastasis. E-cadherin
maintains intercellular connections through intracellular and
extracellular adhesion (27).
Therefore, reducing the expression of E-cadherin will induce
metastasis of cancer cells (28).
Additionally, vimentin is another key factor during the progression
of EMT. The increased expression of vimentin attenuates the cell
adhesion function, resulting in the occurrence of tumor metastasis
(29). Similar results were
observed in the present study; it was demonstrated that the
expression level of vimentin was downregulated; however, the
E-cadherin expression level was upregulated in DU145 cells exposed
to Zey, suggesting that EMT is additionally involved in
Zey-inhibited metastasis in PCa cells.
Previous studies demonstrated that the Wnt/β-catenin
signaling pathway is the key signal transduction pathway that
induces EMT in tumor cells (30–32).
A previous study suggested that Jatrophone depresses the
proliferation and metastasis of breast cancer via the inhibition of
the Wnt/β-catenin signaling pathway and EMT (30). The regulation of the Wnt/β-catenin
signaling pathway is caused by the phosphorylation/degradation of
β-catenin (33). Wnt5a, one of the
members of the Wnt protein family, may induce melanoma EMT and
metastasis through the protein kinase C signaling pathway (34). Therefore, it was hypothesized that
the molecular mechanisms of Zey on PCa growth and metastasis were
pertinent to the Wnt/β-catenin pathway. Consistent with the
mentioned previous studies, the results of the present study
demonstrated that Zey decreased the expression levels of Wnt5a,
β-catenin and cyclin D1 in DU145 cells, suggesting that Zey
attenuated the activity of the Wnt/β-catenin signaling pathway.
A limitation of the present study was that the
effects of Zey were only determined in one cell line of PCa, and
therefore future studies should investigate the role of Zey in
numerous PCa cell lines. In addition, future studies should
investigate the effect of Zey in vivo, which may elucidate a
candidate therapeutic agent for the prevention of PCa.
In conclusion, the present study demonstrated that
Zey repressed the viability, invasion and migration of PCa DU145
cells in a dose-dependent manner. The suppression of the
Wnt/β-catenin pathway was closely associated with the effect of
Zey. Therefore, Zey may be a novel therapeutic strategy for the
treatment of PCa.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analysed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SZ wrote the manuscript. SZ, BZ, JZ and WW performed
the experiments. SZ and CJ designed the study. SZ, BZ, JZ and WW
performed the data analysis. SZ and CJ revised the manuscript. All
authors reviewed the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pang C, Guan Y, Li H, Chen W and Zhu G:
Urologic cancer in China. Jpn J Clin Oncol. 46:497–501. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu Y, Yang XQ, Han CT, Dai B, Zhang HL,
Shi GH, Wang CF and Ye DW: Pathological features of localized
prostate cancer in China: A contemporary analysis of radical
prostatectomy specimens. PLoS One. 10:e01210762015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Acosta AM, Al Rasheed MRH, Rauscher GH,
Vormittag E, Mon KS, Sharif A, Kajdacsy-Balla A and Mohapatra G:
Tumor necrosis in radical prostatectomies with high-grade prostate
cancer is associated with multiple poor prognostic features and a
high prevalence of residual disease. Hum Pathol. 75:1–9. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang C, Tao W, Ni S, Chen Q, Zhao Z, Ma L,
Fu Y and Jiao Z: Tumor-suppressive microRNA-145 induces growth
arrest by targeting SENP1 in human prostate cancer cells. Cancer
Sci. 106:375–382. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ding C, Tian Q, Li J, Jiao M, Song S, Wang
Y, Miao Z and Zhang A: Structural modification of natural product
Tanshinone I leading to discovery of novel nitrogen-enriched
derivatives with enhanced anticancer profile and improved drug-like
properties. J Med Chem. 61:760–776. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han C, Li Z, Hou J, Wang Z, Xu D, Xue G
and Kong L: Bioactivity evaluation of natural product α-mangostin
as a novel xanthone-based lysine-specific demethylase 1 inhibitor
to against tumor metastasis. Bioorg Chem. 76:415–419. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang L, Huo X, Liao Y, Yang F, Gao L and
Cao L: Zeylenone, a naturally occurring cyclohexene oxide, inhibits
proliferation and induces apoptosis in cervical carcinoma cells via
PI3K/AKT/mTOR and MAPK/ERK pathways. Sci Rep. 7:16692017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang L, Jin J, Zhang L, Hu R, Gao L, Huo
X, Liu D, Ma X, Wang C, Han J, et al: Quantitative analysis of
differential protein expression in cervical carcinoma cells after
zeylenone treatment by stable isotope labeling with amino acids in
cell culture. J Proteomics. 126:279–287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu X, Han R, Quan LH, Liu CY and Liao YH:
Stabilization and sustained release of zeylenone, a soft cytotoxic
drug, within polymeric micelles for local antitumor drug delivery.
Int J Pharm. 450:331–337. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang LL, Li RY, Zhang LJ, Chen D, Huo XW,
Li LY and Cao L: Inhibitory effect of Zeylenone on proliferation
and apoptosis of human PC-3 cells. Chine Pharmacol Bull.
29:809–813. 2013.
|
|
12
|
Airola K, Karonen T, Vaalamo M, Lehti K,
Lohi J, Kariniemi AL, Keski-Oja J and Saarialho-Kere UK: Expression
of collagenases-1 and −3 and their inhibitors TIMP-1 and −3
correlates with the level of invasion in malignant melanomas. Br J
Cancer. 80:733–743. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barriere G, Tartary M and Rigaud M:
Metformin: A rising star to fight the epithelial mesenchymal
transition in oncology. Anticancer Agents Med Chem. 13:333–340.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Elkin M, Ariel I, Miao HQ, Nagler A, Pines
M, de-Groot N, Hochberg A and Vlodavsky I: Inhibition of bladder
carcinoma angiogenesis, stromal support, and tumor growth by
halofuginone. Cancer Res. 59:4111–4118. 1999.PubMed/NCBI
|
|
15
|
Kahlert UD, Nikkhah G and Maciaczyk J:
Epithelial-to-mesenchymal(-like) transition as a relevant molecular
event in malignant gliomas. Cancer Lett. 331:131–138. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gillers BS, Chiplunkar A, Aly H, Valenta
T, Basler K, Christoffels VM, Efimov IR, Boukens BJ and Rentschler
S: Canonical wnt signaling regulates atrioventricular junction
programming and electrophysiological properties. Cir Res.
116:398–406. 2015. View Article : Google Scholar
|
|
17
|
Monga SP: β-catenin signaling and roles in
liver homeostasis, injury, and tumorigenesis. Gastroenterology.
148:1294–1310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Yang S, Su N, Wang Y, Yu J, Qiu H
and He X: Overexpression of long non-coding RNA HOTAIR leads to
chemoresistance by activating the Wnt/β-catenin pathway in human
ovarian cancer. Tumour Biol. 37:2057–2065. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng Y, Zhang X, Ma Q, Yan R, Qin Y, Zhao
Y, Cheng Y, Yang M, Wang Q, Feng X, et al: MiRNA-194 activates the
Wnt/β-catenin signaling pathway in gastric cancer by targeting the
negative Wnt regulator, SUFU. Cancer Lett. 385:117–127. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vilchez V, Turcios L, Marti F and Gedaly
R: Targeting Wnt/β-catenin pathway in hepatocellular carcinoma
treatment. World J Gastroenterol. 22:823–832. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Method. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Huang K, Tang Y, He L and Dai Y:
MicroRNA-340 inhibits prostate cancer cell proliferation and
metastasis by targeting the MDM2-p53 pathway. Oncol Rep.
35:887–895. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Luo H, Xiao N, Duan J, Wang Z and
Wang S: Long noncoding RNA SChLAP1 accelerates the proliferation
and metastasis of prostate cancer via targeting miR-198 and
promoting the MAPK1 pathway. Oncol Res. 26:131–143. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei XL, Han R, Hu X, Quan LH, Liu CY,
Chang Q and Liao YH: Stabilization of zeylenone in rat plasma by
the presence of esterase inhibitors and its LC-MS/MS assay for
pharmacokinetic study. Biomed Chromatogr. 27:636–640. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang K, Wu F, Seo BR, Fischbach C, Chen W,
Hsu L and Gourdon D: Breast cancer cells alter the dynamics of
stromal fibronectin-collagen interactions. Matrix Biol.
60–61:86–95. 2017. View Article : Google Scholar
|
|
26
|
Zhou XM, Hou G, Gu DX, Wang QY and Zhao L:
Peroxisome proliferator-activated receptor-γ in induced sputum is
correlated with MMP-9/TIMP-1 imbalance and formation of emphysema
in COPD patients. J Thorac Dis. 9:3703–3710. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hatmaker AR, Gi YJ, Schmidt CR, Beauchamp
RD, Coffey RJ and Pearson AS: Targeted loss of E-cadherin is
sufficient to induce dedifferentiation, loss of intercellular
junctions, and increased invasive potential of colorectal cancer
cells. Cancer Res. 66:5882006.PubMed/NCBI
|
|
28
|
Vu T and Datta PK: Regulation of EMT in
colorectal cancer: A culprit in metastasis. Cancers (Basel). 9:pii:
E171. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chouat E, Zehani A, Chelly I, Njima M,
Maghrebi H, Bani MA, Njim L, Zakhama A, Haouet S and Kchir N: Tumor
budding is a prognostic factor linked to epithelial mesenchymal
transition in pancreatic ductal adenocarcinoma. Study report and
literature review. Pancreatology. 18:79–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fatima I, El-Ayachi I, Taotao L, Lillo MA,
Krutilina R, Seagroves TN, Radaszkiewicz TW, Hutnan M, Bryja V,
Krum SA, et al: The natural compound Jatrophone interferes with
Wnt/β-catenin signaling and inhibits proliferation and EMT in human
triple-negative breast cancer. PLoS One. 12:e01898642017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jiang Y, Ren W, Wang W, Xia J, Gou L, Liu
M, Wan Q, Zhou L, Weng Y, He T and Zhang Y: Inhibitor of β-catenin
and TCF (ICAT) promotes cervical cancer growth and metastasis by
disrupting E-cadherin/β-catenin complex. Oncol Rep. 38:2597–2606.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie SL, Fan S, Zhang SY, Chen WX, Li QX,
Pan GK, Zhang HQ, Wang WW, Weng B, Zhang Z, et al: SOX8 regulates
cancer stem-like properties and cisplatin-induced EMT in tongue
squamous cell carcinoma by acting on the Wnt/β-catenin pathway. Int
J Cancer. 142:1252–1265. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dissanayake SK, Wade M, Johnson CE,
O'Connell MP, Leotlela PD, French AD, Shah KV, Hewitt KJ, Rosenthal
DT, Indig FE, et al: The Wnt5A/protein kinase C pathway mediates
motility in melanoma cells via the inhibition of metastasis
suppressors and initiation of an epithelial to mesenchymal
transition. J Biol Chem. 282:17259–17271. 2007. View Article : Google Scholar : PubMed/NCBI
|