Introduction
Glioma is the most common type of malignant brain
tumour in adults and accounts for >70% brain tumours (1). Regarding the degree of malignancy,
glioma may be divided into four grades (grades I–IV) (2). Glioma is characterized by rapid
growth, cellular heterogeneity, angiogenesis, extensive invasion,
hypoxia and necrosis (3,4). At present, the predominant treatments
for gliomas include surgical resection followed by chemotherapy and
radiotherapy. Significant developments in the diagnosis and therapy
for gliomas have been made; however, the therapeutic outcomes of
patients with glioma remains unsatisfactory (5). The median duration of survival of
patients with glioma is ~15 months post-diagnosis (6). In addition, the therapeutic methods
available for the treatment of gliomas are restricted by poor
understanding of the formation and progression of this cancer
(7). Therefore, thorough
understanding of the molecular mechanisms underlying the
pathogenesis of glioma is essential to identify novel, effective
and targeted therapies for patients with this fatal disease.
MicroRNAs (miRNAs) comprise a series of endogenous,
short single-stranded non-coding RNAs (19–25 molecules) that
primarily serve as gene regulators (8). These molecules negatively regulate
gene expression by directly binding to partial complimentary sites
in the 3′-untranslated regions (3′-UTRs) of their target genes,
resulting in mRNA degradation or inhibition of translation
(9). In total, >1,000 miRNAs
have been identified in the human genome, and these miRNAs have
been estimated to regulate ~30% human protein-coding genes
(10). Numerous miRNAs have
recently been demonstrated to be aberrantly expressed in glioma,
including miR-613 (11), miR-219
(12), miR-936 (13) and miR-141-3p (14). Dysregulation of miRNA contributes
to tumorigenesis and tumour development by regulating numerous
diverse biological processes, including cell proliferation, the
cell cycle, apoptosis, differentiation, metastasis, chemoresistance
and radioresistance (15–17). Therefore, miRNAs may be developed
as diagnostic and therapeutic targets for the management of
malignancies.
miR-940 has been well studied in the pathogenesis of
a number of types of human cancer (18–22);
however, the expression pattern, roles and molecular mechanisms of
the regulatory actions of miR-940 in glioma remain unknown. The
present study aimed to further understand the roles of miR-940 by
studying its expression and mechanisms of action in glioma. In the
present study, significant upregulation of miR-940 in glioma
tissues and cell lines was observed. Additionally, inhibition of
miR-940 decreased cell proliferation and invasion in glioma;
Kruppel-like factor 9 (KLF9) was additionally proposed to be a
novel and direct target of miR-940 in glioma in the present study.
These results not only provide novel insight into the underlying
mechanisms associated with the occurrence and development of
glioma; however, may additionally contribute to the identification
of novel therapeutic targets for glioma.
Materials and methods
Human tissue specimens
A total of 23 pairs of glioma and adjacent normal
brain tissues were obtained from patients (14 males, 9 females; age
range, 47–72 years) diagnosed with glioma and treated via surgical
resection at The First Affiliated Hospital of Xi'an Medical
University (Xi'an, China) between October 2015 to March 2017. None
of the patients underwent radiotherapy or chemotherapy prior to
surgical resection. The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Xi'an Medical
University. Written informed consent was provided by all
participants.
Cell lines, culture conditions and
transfection
Normal human astrocytes (NHAs) were purchased from
ScienCell Research Laboratories, Inc. (San Diego, CA, USA) and
cultured in astrocyte medium (ScienCell Research Laboratories,
Inc.) according to the manufacturer's protocols. A total of four
glioma cell lines (U251, U87, T98G and LN229) were acquired from
the Cell Bank of Type Culture Collection of Chinese Academy of
Sciences (Shanghai, China), and cultured in Dulbecco's modified
Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) and
1% penicillin and 1% streptomycin (all obtained from HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) as described below. The
U87 cell line originated from the American Type Culture Collection
(Manassas, VA, USA), and is likely to be glioblastoma, but of
unknown origin. This condition did not affect the interpretation of
the results of the present study as glioblastoma multiforme is the
subtype of glioma. U251 and U87 cell lines exhibited relatively
higher miR-940 expression among the four glioma cell lines; thus,
these two cell lines were chosen for further functional assays. The
cell lines were maintained at 37°C in an incubator with 5%
CO2.
miR-940 inhibitor and the corresponding negative
control miRNA inhibitor (NC inhibitor) were synthesised by Shanghai
GenePharma Co., Ltd. (Shanghai, China). Small interfering RNA
(siRNA) against the KLF9 (KLF9 siRNA) and negative control siRNA
(NC siRNA) were chemically produced by Guangzhou RiboBio Co., Ltd.
(Guangzhou, China). The sequences were as follows: miR-940
inhibitor, 5′-GGGGAGCGGGGGUGCCUU-3′; NC inhibitor,
5′-CAGUACUUUUGUGUAGUACAA-3′; KLF9 siRNA, 5′-CAGUUCCGCUGUCCGCUGU-3′
and NC siRNA, 5′-UUCUCCGAACGUGUCACGUTT-3′. U251 and U87 cells were
seeded into 6-well plates with a density of 8×105 cells
per well, and transfected with miR-940 inhibitor (100 pmol) or NC
inhibitor (100 pmol), or KLF9 siRNA (100 pmol) or NC siRNA (100
pmol) using Lipofectamine 2000™ (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's protocol. Successful transfection was determined
through detecting the expression of miR-940 and KLF9 siRNA,
respectively. Reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and the cell invasion assay were conducted
following 48 h post-transfection. The MTT assay and western blot
analysis were performed at 24 and 72 h post-transfection,
respectively.
RNA isolation and RT-qPCR
A TRIzol® Reagent Kit (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract total RNA from tissue
samples or cell lines. To analyse miR-940 expression, RT was
conducted using a TaqMan MicroRNA RT Kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.); qPCR was performed using a TaqMan
MicroRNA assay (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The temperature protocol for RT-PCR was as follows: 16°C for
30 min, 42°C for 30 min and 85°C for 5 min. The thermocycling
conditions for qPCR were as follows: 50°C for 2 min, 95°C for 10
min, followed by 40 cycles of denaturation at 95°C for 15 sec and
annealing/extension at 60°C for 60 sec. To examine the mRNA
expression levels of KLF9, cDNA was synthesized from total RNA
using a SYBR Premix Ex Taq II Kit (Takara Biotechnology Co., Ltd.,
Dalian, China) followed by qPCR with a SYBR Premix Ex
Taq™ (Takara Biotechnology Co., Ltd.). The temperature
protocol for RT-PCR was as follows: 37°C for 15 min and 85°C for 5
sec. The thermocycling conditions for qPCR were as follows: 5 min
at 95°C, followed by 40 cycles of 95°C for 30 sec and 65°C for 45
sec. U6 and GAPDH were used as endogenous controls to normalize the
expression levels of miR-940 and KLF9 mRNA, respectively. The
primers were designed as follows: miR-940, forward
5′-CACACATCCGTCTGGGGCTAGG-3′, reverse, 5′-CTACAGAATGCCGCCGCTGCT-3′;
U6, forward 5′-GCTTCGGCAGCACATATACTAAAAT-3′, reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; KLF9, forward
5′-ACAGTGGCTGTGGGAAAGTC-3′, reverse, 5′-TCACAAAGCGTTGGCCAGCG-3′ and
GAPDH, forward 5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′. Relative gene expression was
calculated using the 2−ΔΔCq method (23).
MTT assay
An MTT assay was conducted to evaluate the effects
of miR-940 on glioma cell proliferation. Transfected cells were
seeded into 96-well plates at a density of 3,000 cells/well and
incubated at 37°C for various durations (0, 24, 48 and 72 h). At
each time point, 20 µl MTT solution (Beyotime Institute of
Biotechnology, Haimen, China) was added into each well, which was
further incubated at 37°C for 4 h. The supernatant was subsequently
discarded, and 200 µl dimethyl sulfoxide was added into each well
to dissolve the formazan crystals. Cell proliferation was
determined by detecting the optical density at a wavelength of 490
nm with an enzyme-linked immunosorbent assay reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Cell invasion assay
A cell invasion assay was performed to detect the
invasive abilities of gliomas cells. Transfected U251 and U87 cells
were collected and suspended in FBS-free DMEM. A total of
1×105 suspended cells were seeded into the upper
chambers (Corning, Inc., Corning, NY, USA) coated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). The lower chambers were
covered with 500 µl DMEM supplemented with 10% FBS. After 48 h
incubation, non-invasive cells that did not invade via the pores
were gently removed with cotton swabs. The invasive cells were
fixed with 4% polyoxymethylene at room temperature for 20 min,
stained with 0.5% crystal violet at room temperature for 20 min;
cells were analyzed under an inverted light microscope
(magnification, ×200; CKX41; Olympus Corporation, Tokyo, Japan).
The number of invasive cells was counted in at least five randomly
selected fields.
Bioinformatics analysis
Online miRNA target prediction software, including
TargetScan 7.1 (www.targetscan.org/) and miRanda (www.microrna.org) were used to predict the putative
target genes of miR-940.
Dual-luciferase reporter assay
The 3′-UTR of KLF9 containing putative wild-type
(Wt) or mutated (Mut) miR-940-binding sequences was cloned and
inserted into the pmirGLO Dual-Luciferase miRNA target expression
vector (Promega Corporation, Madison, WI, USA), and labelled as
pmirGLO-KLF9-3′-UTR Wt or pmirGLO-KLF9-3′-UTR Mut, respectively.
Cells were seeded into 24-well plates with a density of
1.0×105 cells per well one night prior to transfection.
Cells were co-transfected with miR-940 inhibitor or NC inhibitor
and pmirGLO-KLF9-3′-UTR Wt or pmirGLO-KLF9-3′-UTR Mut, using
Lipofectamine 2000™ (Thermo Fisher Scientific, Inc.).
Following transfection for 48 h, transfected cells were collected
and subjected to luciferase activity analysis using a
Dual-Luciferase reporter assay system (Promega Corporation)
according to the manufacturer's protocol. The firefly luciferase
activity was normalized to Renilla luciferase activity.
Western blot analysis
Total protein of tissues and transfected cells was
isolated using radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology), and was quantified using a
bicinchoninic acid protein assay Kit (Beyotime Institute of
Biotechnology). An equal amount of protein (30 µg) was separated by
10% SDS-PAGE, which were subsequently transferred to a
nitrocellulose membrane. Subsequently, the membrane was blocked at
room temperature with 5% non-fat milk for 1 h and incubated
overnight at 4°C with primary antibodies against KLF9 (1:1,000;
cat. no. ab227920; Abcam, Cambridge, UK) or GAPDH (1:1,000; cat.
no. ab128915; Abcam). The membrane was subsequently incubated with
goat anti-rabbit immunoglobulin G horseradish peroxidase-conjugated
secondary antibody (1:5,000; cat. no. ab205718; Abcam) at room
temperature for 2 h and visualised with an enhanced
chemiluminescence reagent (Bio-Rad Laboratories, Inc.). GAPDH
served as the loading control. Protein expression was quantified
using Quantity One software version 4.62 (Bio-Rad Laboratories,
Inc.).
Statistical analysis
The results of all statistical analyses conducted in
the present study are presented as the mean ± standard deviation
from three independent experiments. Data were analysed with SPSS
17.0 software (SPSS, Inc., Chicago, IL, USA). Differences between
groups were analyzed with a two-tailed Student's t-test or one-way
analysis of variance followed by Tukey's or Bonferroni post hoc
test. Spearman's correlation analysis was adopted to investigate
the correlation between miR-940 and KLF9 mRNA expression in glioma
tissues. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-940 is upregulated in glioma
tissues and cell lines
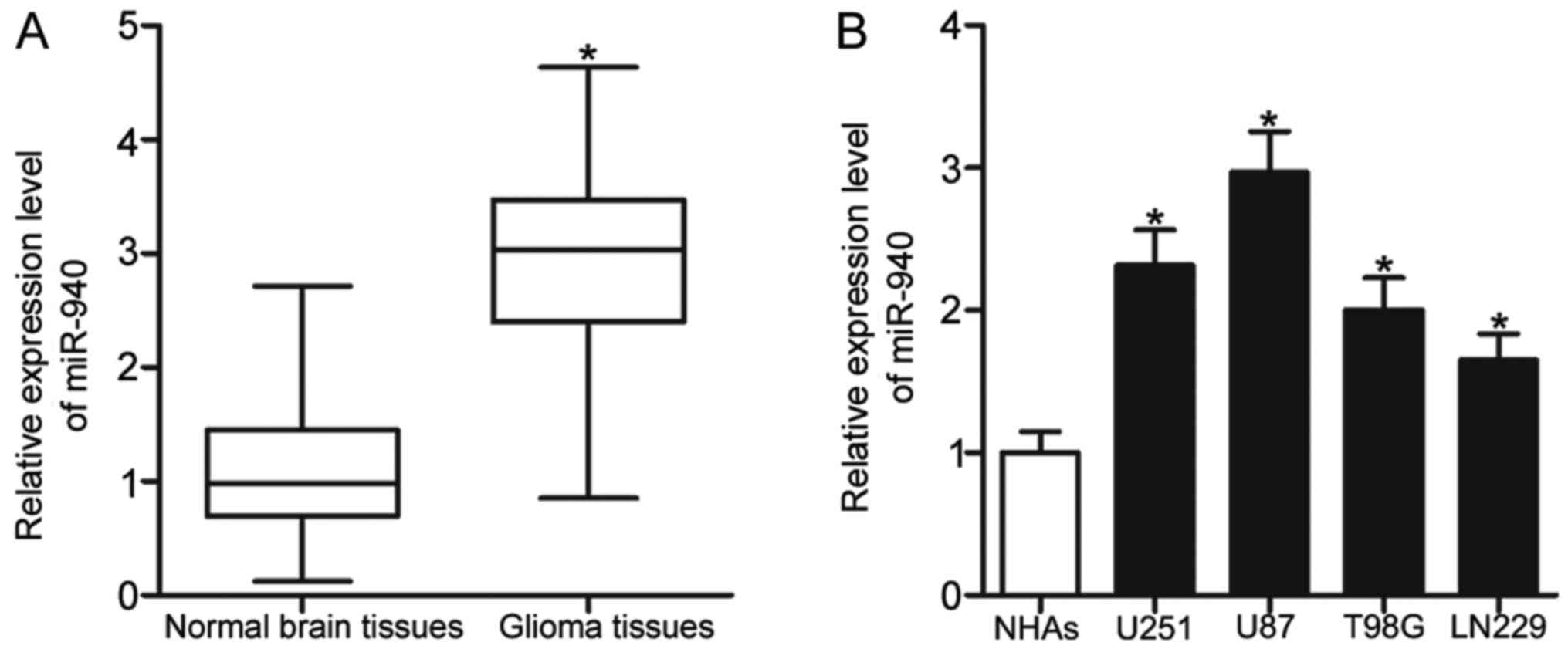
To determine the biological role of miR-940 in
glioma, its expression was analysed in 23 paired glioma and
adjacent normal brain tissues via RT-qPCR. The data demonstrated
significant upregulation of miR-940 expression in glioma tissues
compared with adjacent normal brain tissues (Fig. 1A; P<0.05). Expression levels of
miR-940 were additionally determined in four glioma cell lines and
NHAs. Compared with the NHAs, all glioma cell lines exhibited
significantly increased expression levels of miR-940 (Fig. 1B; P<0.05). These results
demonstrated that miR-940 is upregulated in glioma.
Inhibition of miR-940 inhibits glioma
cell proliferation and invasion
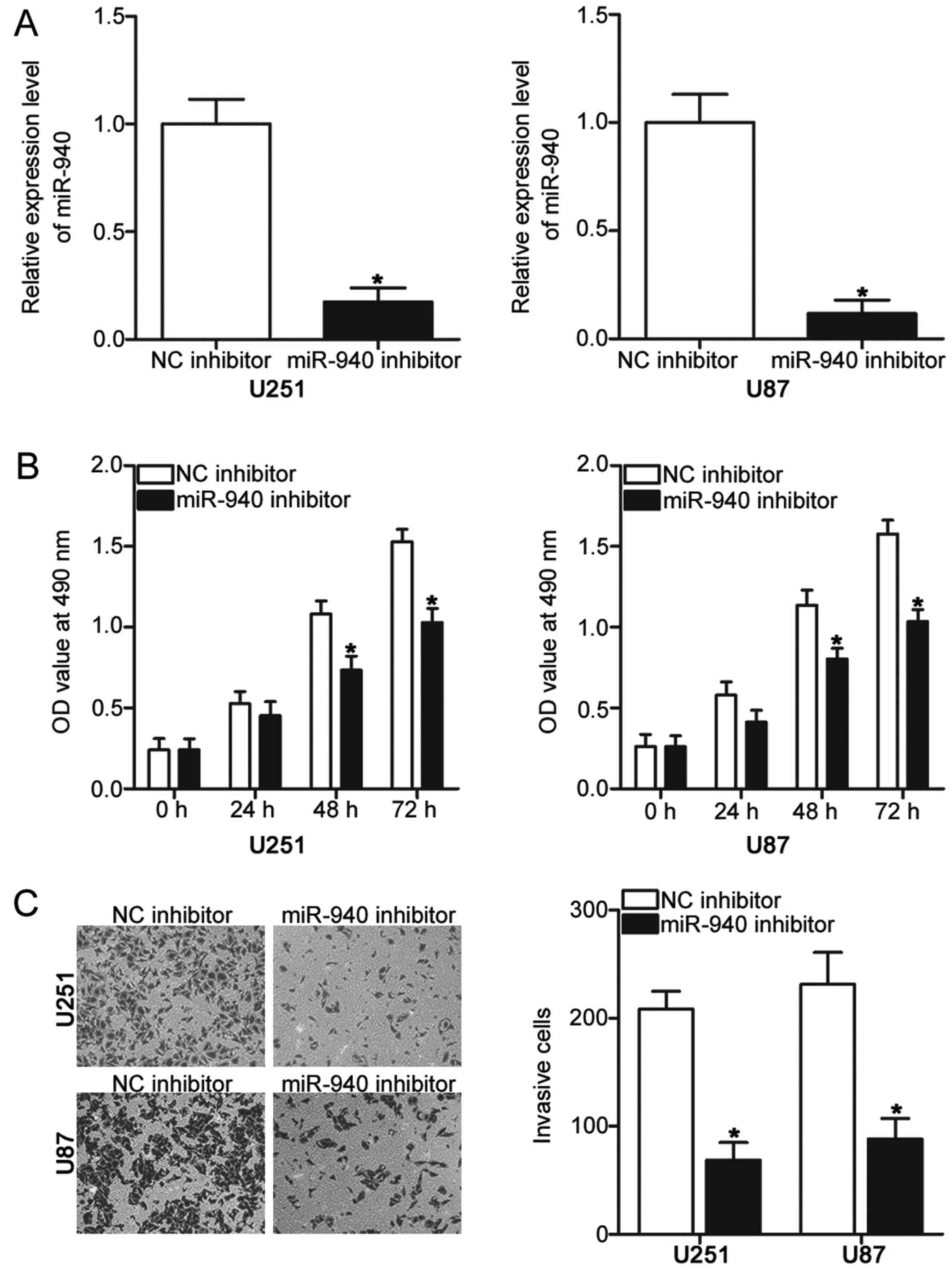
To examine the biological functions of miR-940 in
glioma, an miR-940 inhibitor was transfected into U251 and U87
cells to decrease endogenous expression levels. After 48 h
transfection, RT-qPCR analysis was used to assess transfection
efficiency. The results of the present study demonstrated a
significant downregulation of miR-940 in U251 and U87 cells
following transfection with miR-940 inhibitor compared with cells
transfected with NC inhibitor (Fig.
2A; P<0.05). MTT and cell invasion assays were conducted to
evaluate the effects of miR-940 downregulation on glioma cell
proliferation and invasion, respectively. The results demonstrated
that inhibition of miR-940 expression significantly decreased
proliferation (Fig. 2B; P<0.05)
at 48 and 72 h and invasion (Fig.
2C; P<0.05) of U251 and U87 cells compared with cells
transfected with NC inhibitor. These results suggested the possible
role of miR-940 in the progression of glioma.
KLF9 is a direct target gene of
miR-940 in glioma
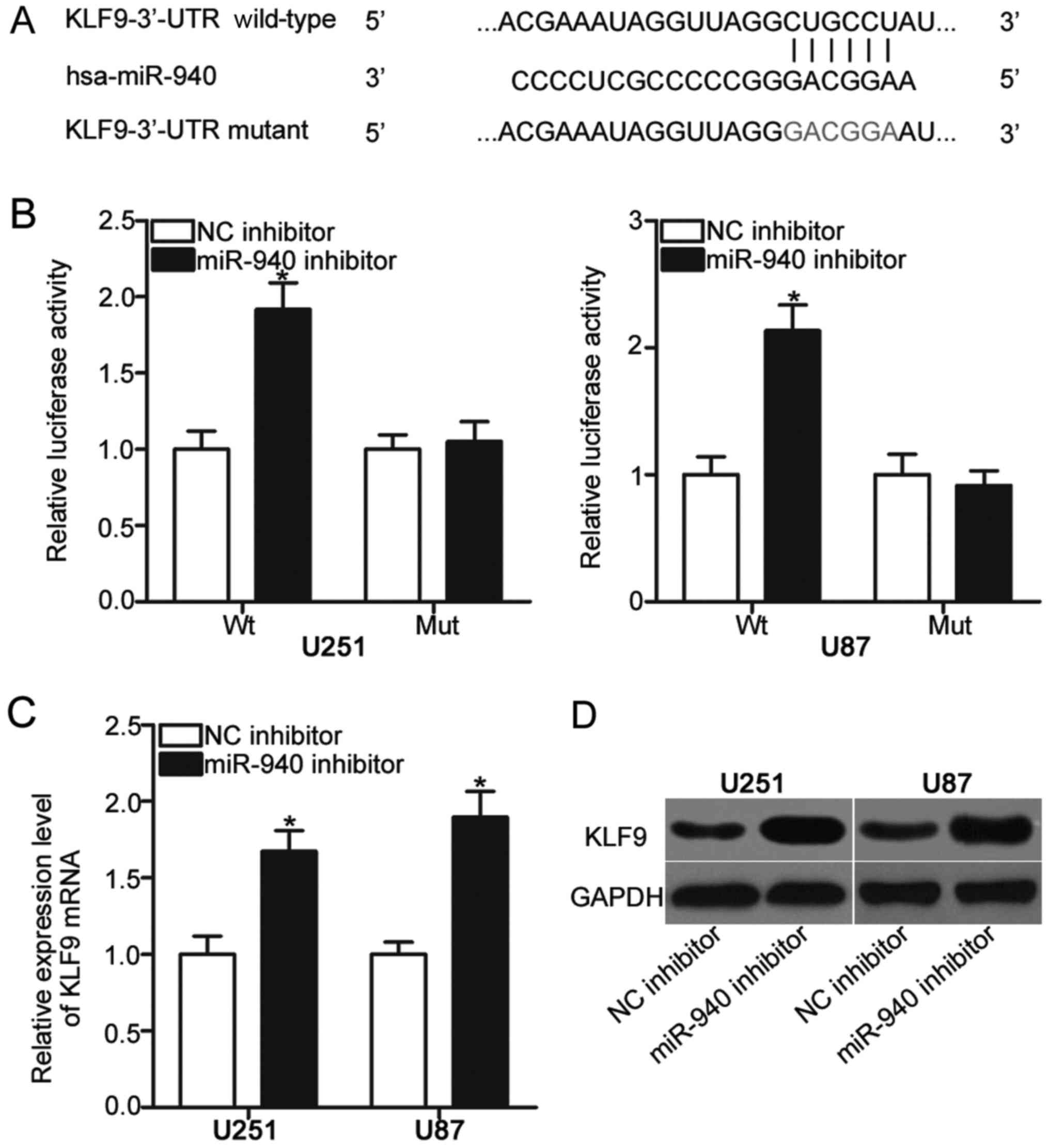
To investigate the underlying molecular mechanism of
miR-940 in glioma, the present study performed bioinformatics
analyses to identify the potential targets of miR-940. KLF9 was
predicted to be a target of miR-940 (Fig. 3A). To confirm whether miR-940
directly targets the 3′-UTR of KLF9, a dual-luciferase reporter
assay was conducted using U251 and U87 cells co-transfected with
miR-940 inhibitor or NC inhibitor and pmirGLO-KLF9-3′-UTR Wt or
pmirGLO-KLF9-3′-UTR Mut. As presented in Fig. 3B, the downregulation of miR-940
increased the luciferase activity of cells transfected with
pmirGLO-KLF9-3′-UTR Wt; however, luciferase activity of
pmirGLO-KLF9-3′-UTR Mut was notably unaffected in U251 and U87
cells. To further understand the regulation of KLF9 mediated by
miR-940, RT-qPCR and western blot analysis was conducted to measure
KLF9 expression levels in U251 and U87 cells transfected with
miR-940 inhibitor or NC inhibitor. As presented in Fig. 3C and D, KLF9 mRNA (P<0.05) and
protein (P<0.05) expression levels in miR-940
inhibitor-transfected U251 and U87 cells were significantly
upregulated compared with cells transfected with NC inhibitor.
Collectively, these results suggested that KLF9 may be a direct
target gene of miR-940 in glioma.
Inverse correlation between miR-940
and KLF9 expression levels in glioma tissues
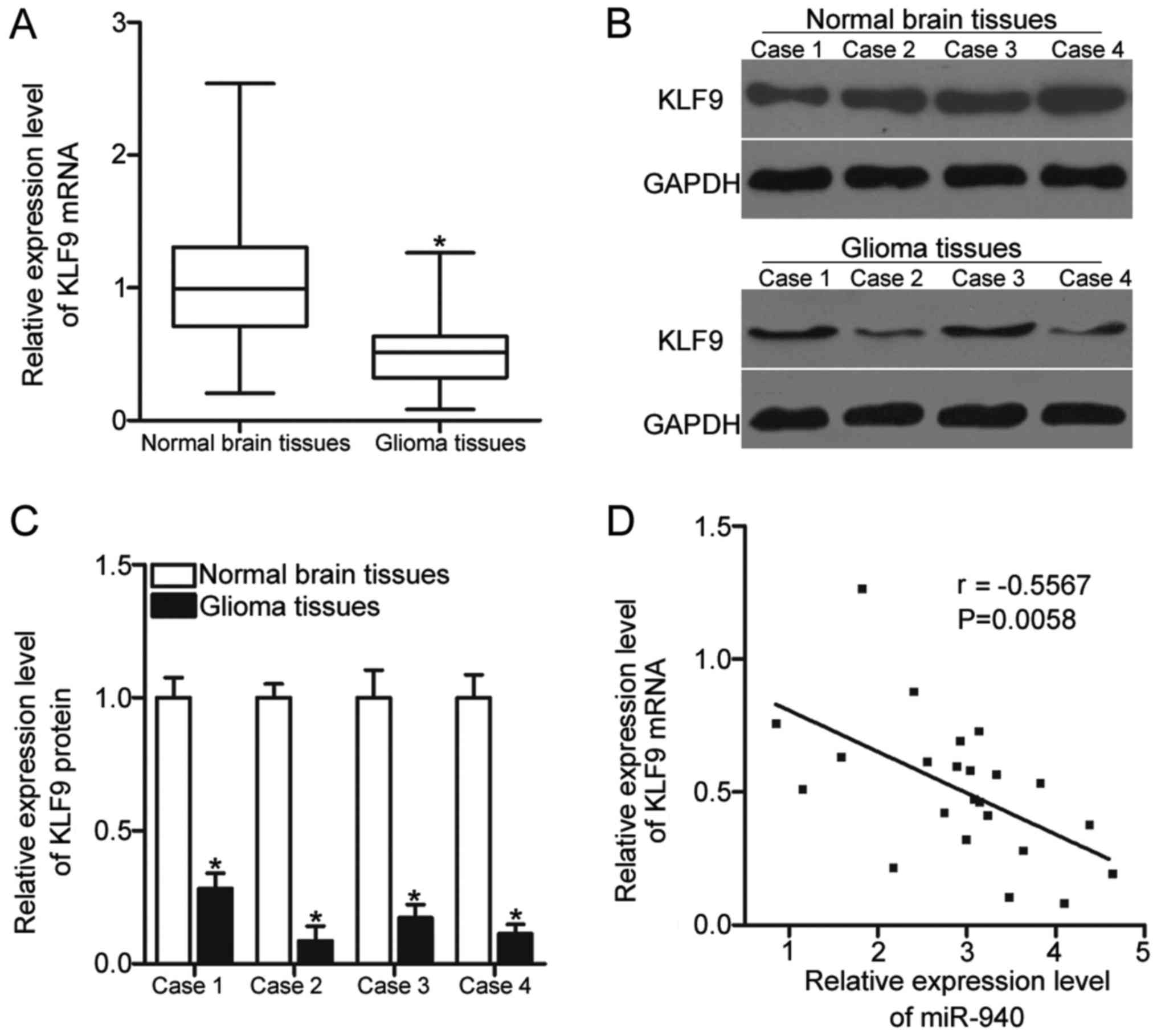
KLF9 expression was measured in 23 paired glioma
tissues and adjacent normal brain tissues to demonstrate the
association between miR-940 and KLF9. As presented in Fig. 4A, KLF9 mRNA expression levels in
glioma tissues were significantly decreased compared with adjacent
normal brain tissues (P<0.05). The protein expression levels of
KLF9 in a number of pairs of glioma and adjacent normal brain
tissues was determined using western blot analysis. The expression
levels of KLF9 protein were downregulated in glioma tissues
compared with adjacent normal brain tissues (Fig. 4B and C; P<0.05). Furthermore,
KLF9 mRNA expression levels in glioma tissues were inversely
correlated with miR-940 expression levels in glioma tissues
(Fig. 4D; R=−0.5567; P=0.0058).
These results suggested that KLF9 may be a direct target of miR-940
in glioma tissues.
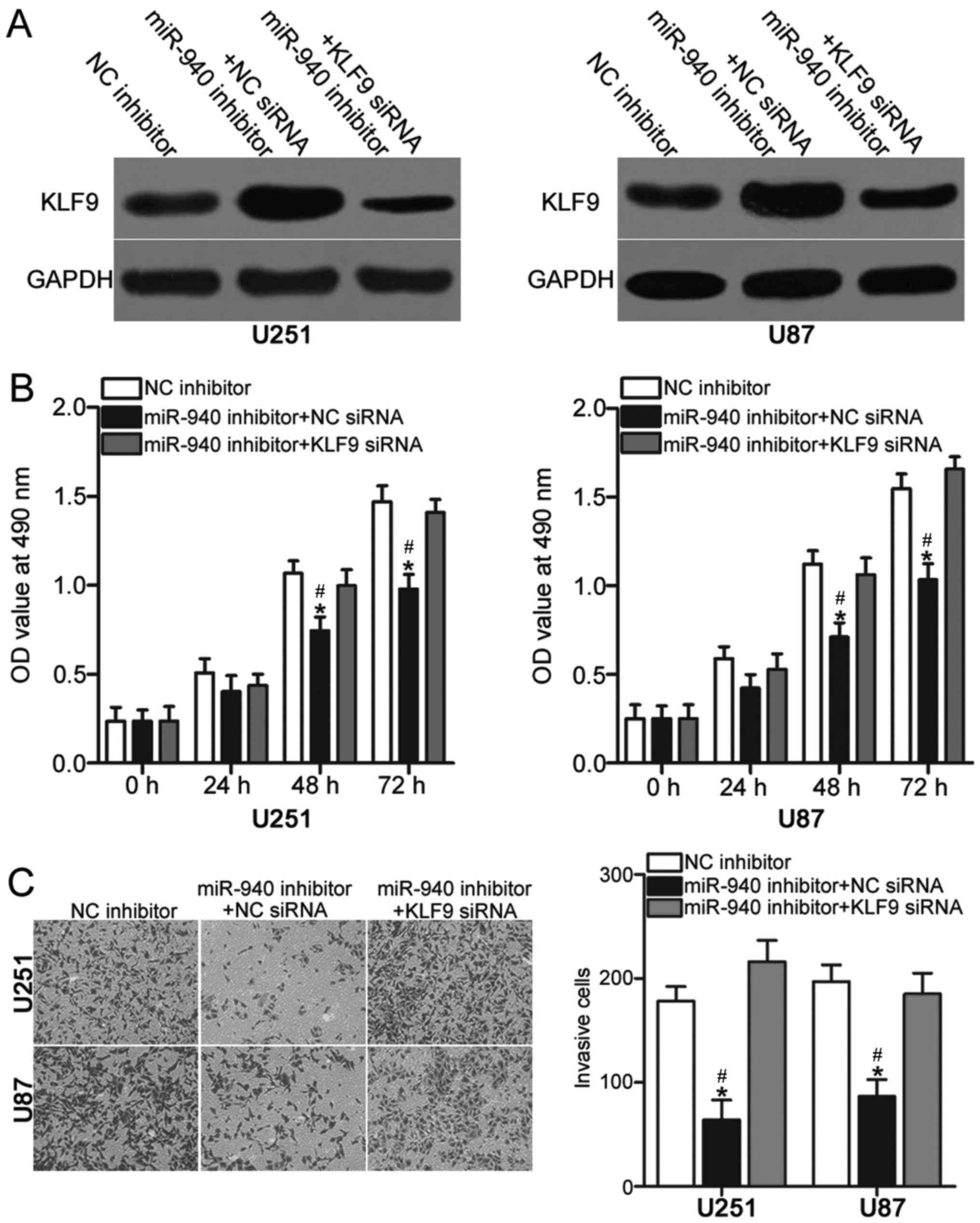
KLF9 knockdown reverses the effects of
miR-940 on glioma cell proliferation and invasion
As aforementioned, the downregulation of miR-940
suppressed cell proliferation and invasion in glioma and KLF9 was
identified as a direct target of miR-940. Therefore, the present
study aimed to determine whether alterations in cell proliferation
and invasion following miR-940 knockdown directly occur as a result
of KLF9 upregulation. U251 and U87 cells were transfected with
miR-940 inhibitor in the presence of KLF9 siRNA or NC siRNA.
Western blotting demonstrated that the increased expression levels
of KLF9 due to miR-940 inhibitor were suppressed via
co-transfection with KLF9 siRNA (Fig.
5A). The inhibition of proliferation (Fig. 5B; P<0.05) and invasion (Fig. 5C; P<0.05) associated with
miR-940 downregulation was reversed by KLF9 knockdown. The
oncogenic roles of miR-940 in glioma cell proliferation and
invasion may be, at least in part, attributable to the upregulation
of KLF9.
Discussion
In the present study, miR-940 expression levels were
upregulated in glioma tissues and cell lines. Downregulation of
miR-940 decreased the proliferation and invasion of glioma cells;
KLF9 was confirmed as a direct target gene of miR-940 in glioma. In
addition, the expression levels of KLF9 were downregulated in
glioma tissues and were negatively correlated with that of miR-940
in the present study. Furthermore, KLF9 knockdown partially
reversed the oncogenic effects of miR-940 on glioma cell
proliferation and invasion. The results of the present study
suggest that miR-940 may be further developed as a promising
therapeutic target for the treatment of NSCLC.
miR-940 was previously demonstrated to be aberrantly
expressed in a number of types of human cancer. miR-940 was
downregulated in hepatocellular carcinoma, and this downregulation
was significantly associated with Edmondson grade, tumor
microsatellite or multiple tumors, vascular invasion, recurrence
and metastasis (18,24). Patients with hepatocellular
carcinoma and low miR-940 expression levels exhibited poorer
prognosis compared with high expression levels (18). miR-940 was additionally
demonstrated to be downregulated in prostate (20), breast (21) and ovarian cancer (22); however, miR-940 expression was
observed to be upregulated in gastric cancer tissues, cell lines
and plasma (25). High miR-940
expression levels were strongly correlated with the advanced N
stage of gastric cancer (19).
Additionally, patients with hepatocellular carcinoma and high
miR-940 expression levels exhibited shorter median times of
recurrence compared with patients with low expression levels
(19). Upregulation of miR-940 was
additionally observed in pancreatic cancer tissues and cell lines
(26). These results suggested
that the expression profile of miR-940 exhibits tissue specificity
and may be developed as a biomarker for the detection and prognosis
of particular cancer types.
Dysregulation of miR-940 has been associated with
tumor initiation and the progression of numerous types of human
cancer (18–21). Upregulation of miR-940 suppressed
cell growth and promoted apoptosis of hepatocellular carcinoma
(18). Rajendiran et al
(20) observed that ectopic
expression of miR-940 decreased cell migration and invasion of
prostate cancer. Hou et al (21) demonstrated that miR-940
overexpression prohibited the proliferative and migratory abilities
of triple-negative breast cancer cells. Conversely, Wang et
al (22) identified that
restoration of miR-940 expression restricted cell proliferation and
induced apoptosis in ovarian cancer. However, miR-940 served
oncogenic roles in gastric cancer by promoting cell metastasis
in vitro and in vivo (19). Yang et al (26) proposed that upregulation of miR-940
increased cell proliferation and invasion of pancreatic cancer.
These results demonstrated the tissue specific biological roles of
miR-940 in various types of human cancer. In addition, miR-940 may
be an effective therapeutic target for the treatment of these human
malignancies.
Numerous targets of miR-940 have been identified,
including estrogen-related receptor g in hepatocellular carcinoma
(18), migration and invasion
enhancer 1 in prostate cancer (20), protein kinase C-δ in ovarian cancer
(22), zinc finger protein 24 in
gastric cancer (19), and glycogen
synthase kinase 3β (26) and
secreted frizzled related protein 1 (26) in pancreatic carcinoma. In the
present study, KLF9 was identified as a direct target gene of
miR-940 in glioma. Bioinformatics analysis predicted that KLF9 is a
potential target of miR-940. Additionally, luciferase reporter
assays demonstrated that downregulation of miR-940 increased the
luciferase activity of Wt; however, not the mutated plasmids,
suggesting that miR-940 may directly target the 3′-UTR of KLF9. In
addition, inhibition of miR-940 exhibited a significant
upregulation in the expression of KLF9 at the mRNA and protein
expression levels. Furthermore, KLF9 was downregulated in glioma
tissues and was negatively correlated with miR-940 expression
levels. Finally, KLF9 knockdown may counteract the effects of
miR-940 on glioma cell proliferation and invasion. Collectively,
miR-940 may serve oncogenic roles in glioma by directly targeting
KLF9.
KLF9, located on human chromosome 9q13, is a member
of the KLF family (27). The
expression of KLF9 was decreased in numerous types of human
malignant tumors, including pancreatic ductal adenocarcinoma
(28), esophageal squamous cell
carcinoma (29), hepatocellular
carcinoma (30) and prostate
cancer (31). Increasing evidence
suggests the significant role of KLF9 in carcinogenesis and cancer
progression; KLF9 serves as tumor suppressor and regulates a
variety of cellular processes, including cell proliferation,
apoptosis, migration and invasion (29,31–33).
Additionally, low levels of KLF9 expression have been detected in
glioma tissues and cell lines (34). Decreased KLF9 expression promotes
cell proliferation in vitro and tumor growth in vivo
(34). These results suggest that
KLF9 may be a promising target for patients with glioma.
In conclusion, miR-940 was overexpressed in glioma
tissues and cell lines in the present study. Decreased miR-940
expression levels may inhibit cellular proliferation and invasion
of glioma cells by directly targeting KLF9. The results of the
present study suggested a theoretical basis for the application of
miR-940/KLF9 in the treatment of patients with glioma.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SW and DZ designed the present study. DZ, PH and LJ
performed RT-qPCR, and the MTT and luciferase reporter assays. YW
and ZY conducted the cell invasion assay and western blot analysis.
All authors have read and approved the final draft.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Xi'an Medical
University (Xi'an, China). Written informed consent was provided by
all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Buckner JC, Brown PD, O'Neill BP, Meyer
FB, Wetmore CJ and Uhm JH: Central nervous system tumors. Mayo Clin
Proc. 82:1271–1286. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang R, Luo H, Wang S, Chen W, Chen Z,
Wang HW, Chen Y, Yang J, Zhang X, Wu W, et al: MicroRNA-377
inhibited proliferation and invasion of human glioblastoma cells by
directly targeting specificity protein 1. Neuro Oncol.
16:1510–1522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lukiw WJ, Cui JG, Li YY and Culicchia F:
Up-regulation of micro-RNA-221 (miRNA-221; chr Xp11.3) and
caspase-3 accompanies down-regulation of the survivin-1 homolog
BIRC1 (NAIP) in glioblastoma multiforme (GBM). J Neurooncol.
91:27–32. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Komotar RJ, Otten ML, Moise G and Connolly
ES Jr: Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma-a critical review. Clin Med Oncol. 2:421–422.
2008.PubMed/NCBI
|
|
6
|
Linz U: Commentary on effects of
radiotherapy with concomitant and adjuvant temozolomide versus
radiotherapy alone on survival in glioblastoma in a randomised
phase III study: 5-year analysis of the EORTC-NCIC trial (Lancet
Oncol. 2009;10:459-466). Cancer. 116:1844–1846. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Louis DN: Molecular pathology of malignant
gliomas. Annu Rev Pathol. 1:97–117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Q, Zhou L, Wang M, Wang N, Li C, Wang J
and Qi L: MicroRNA-613 impedes the proliferation and invasion of
glioma cells by targeting cyclin-dependent kinase 14. Biomed
Pharmacother. 98:636–642. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang B, Li M, Ji F and Nie Y:
MicroRNA-219 exerts a tumor suppressive role in glioma via
targeting Sal-like protein 4. Exp Ther Med. 14:6213–6221.
2017.PubMed/NCBI
|
|
13
|
Wang D, Zhi T, Xu X, Bao Z, Fan L, Li Z,
Ji J and Liu N: MicroRNA-936 induces cell cycle arrest and inhibits
glioma cell proliferation by targeting CKS1. Am J Cancer Res.
7:2131–2143. 2017.PubMed/NCBI
|
|
14
|
Zhou X, Wu W, Zeng A, Nie E, Jin X, Yu T,
Zhi T, Jiang K, Wang Y, Zhang J, et al: MicroRNA-141-3p promotes
glioma cell growth and temozolomide resistance by directly
targeting p53. Oncotarget. 8:71080–71094. 2017.PubMed/NCBI
|
|
15
|
Mao K, Lei D, Zhang H and You C:
MicroRNA-485 inhibits malignant biological behaviour of
glioblastoma cells by directly targeting PAK4. Int J Oncol.
51:1521–1532. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang J, Zhang J, Zhang J, Qiu W, Xu S, Yu
Q, Liu C, Wang Y, Lu A, Zhang J, et al: MicroRNA-625 inhibits the
proliferation and increases the chemosensitivity of glioma by
directly targeting AKT2. Am J Cancer Res. 7:1835–1849.
2017.PubMed/NCBI
|
|
17
|
Chen L, Wang W, Zhu S, Jin X, Wang J, Zhu
J and Zhou Y: MicroRNA-590-3p enhances the radioresistance in
glioblastoma cells by targeting LRIG1. Exp Ther Med. 14:1818–1824.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan B, Liang Y, Wang D and Luo F: MiR-940
inhibits hepatocellular carcinoma growth and correlates with
prognosis of hepatocellular carcinoma patients. Cancer Sci.
106:819–824. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu X, Ge X, Zhang Z, Zhang X, Chang J, Wu
Z, Tang W, Gan L, Sun M and Li J: MicroRNA-940 promotes tumor cell
invasion and metastasis by downregulating ZNF24 in gastric cancer.
Oncotarget. 6:25418–25428. 2015.PubMed/NCBI
|
|
20
|
Rajendiran S, Parwani AV, Hare RJ,
Dasgupta S, Roby RK and Vishwanatha JK: MicroRNA-940 suppresses
prostate cancer migration and invasion by regulating MIEN1. Mol
Cancer. 13:2502014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hou L, Chen M, Yang H, Xing T, Li J, Li G,
Zhang L, Deng S, Hu J, Zhao X, et al: MiR-940 inhibited cell growth
and migration in triple-negative breast cancer. Med Sci Monit.
22:3666–3672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang F, Wang Z, Gu X and Cui J: miR-940
upregulation suppresses cell proliferation and induces apoptosis by
targeting PKC-δ in ovarian cancer OVCAR3 cells. Oncol Res.
25:107–114. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ding D, Zhang Y, Yang R, Wang X, Ji G, Huo
L, Shao Z and Li X: miR-940 suppresses tumor cell invasion and
migration via regulation of CXCR2 in hepatocellular carcinoma.
Biomed Res Int. 2016:76183422016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu X, Kwong A, Sihoe A and Chu KM: Plasma
miR-940 may serve as a novel biomarker for gastric cancer. Tumour
Biol. 37:3589–3597. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang HW, Liu GH, Liu YQ, Zhao HC, Yang Z,
Zhao CL, Zhang XF and Ye H: Over-expression of microRNA-940
promotes cell proliferation by targeting GSK3β and sFRP1 in human
pancreatic carcinoma. Biomed Pharmacother. 83:593–601. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McConnell BB and Yang VW: Mammalian
Krüppel-like factors in health and diseases. Physiol Rev.
90:1337–1381. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mao Z, Fan X, Zhang J, Wang X, Ma X,
Michalski CW and Zhang Y: KLF9 is a prognostic indicator in human
pancreatic ductal adenocarcinoma. Anticancer Res. 37:3795–3799.
2017.PubMed/NCBI
|
|
29
|
Qiao F, Yao F, Chen L, Lu C, Ni Y, Fang W
and Jin H: Krüppel-like factor 9 was down-regulated in esophageal
squamous cell carcinoma and negatively regulated beta-catenin/TCF
signaling. Mol Carcinog. 55:280–291. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun J, Wang B and Liu Y, Zhang L, Ma A,
Yang Z, Ji Y and Liu Y: Transcription factor KLF9 suppresses the
growth of hepatocellular carcinoma cells in vivo and positively
regulates p53 expression. Cancer Lett. 355:25–33. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen P, Sun J, Xu G, Zhang L, Yang Z, Xia
S, Wang Y, Liu Y and Shi G: KLF9, a transcription factor induced in
flutamide-caused cell apoptosis, inhibits AKT activation and
suppresses tumor growth of prostate cancer cells. Prostate.
74:946–958. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bai XY, Li S, Wang M, Li X, Yang Y, Xu Z,
Li B, Li Y, Xia K, Chen H, et al: Krüppel-like factor 9
down-regulates matrix metalloproteinase 9 transcription and
suppresses human breast cancer invasion. Cancer Lett. 412:224–235.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mannava S, Zhuang D, Nair JR, Bansal R,
Wawrzyniak JA, Zucker SN, Fink EE, Moparthy KC, Hu Q, Liu S, et al:
KLF9 is a novel transcriptional regulator of bortezomib- and
LBH589-induced apoptosis in multiple myeloma cells. Blood.
119:1450–1458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang S, Wang C, Yi Y, Sun X, Luo M, Zhou
Z, Li J, Cai Y, Jiang X and Ke Y: Krüppel-like factor 9 inhibits
glioma cell proliferation and tumorigenicity via downregulation of
miR-21. Cancer Lett. 356:547–555. 2015. View Article : Google Scholar : PubMed/NCBI
|