Introduction
Asthma is a heterogeneous, chronic respiratory
disease characterised by airway inflammation, increased airway
hyperresponsiveness and variable airflow obstruction (1,2).
Asthma has the highest rate of morbidity worldwide, and the
prevalence has increased significantly over the past decades
(3,4). However, the pathophysiology of asthma
remains unclear (5).
Interleukin (IL)-4 and IL-13, cytokines expressed by
T-helper type 2 (Th2) cells, play key roles in the pathogenesis of
atopy and atopic asthma (6,7).
Both IL-4 and IL-13 promote acute inflammatory processes of asthma
and underlying structural changes in the airways; the receptors are
expressed by various cell types (8,9). The
inflammatory mediator IL-4 induces eosinophil infiltration in the
airway and promotes inflammatory cell chemotaxis (10). IL-13 also induces eosinophil
inflammation, airway hyperreactivity (AHR) and mucus hypersecretion
(11,12).
MicroRNAs (miRNAs) are small, noncoding RNAs
involved in various physiological processes and diseases (13). miRNAs regulate gene expression
post-transcriptionally by binding to the 3′-untranslated regions
(UTRs) of targeted messenger RNAs (mRNAs) for degradation or
translational repression (14,15).
Emerging evidence supports a link between miRNAs and bronchial
asthma. The levels of miR-148, miR-26a, Let-7a and Let-7d have been
found to be altered in sera of asthmatic patients (16,17).
miR-410 has not previously been reported to be involved in asthma
but affects retinal neovascularisation during oxygen-induced
retinopathy in mice, exerting an anti-inflammatory action, although
the underlying mechanism has not been described (18). We speculated that miR-410 may also
be involved in the pathogenesis of airway inflammation. Our aim was
to explore whether intranasal administration of miR-410 attenuates
such inflammation in OVA-induced asthmatic mice, and the possible
therapeutic mechanism in play.
Materials and methods
Animals and reagents
Female BALB/c mice (6–8 weeks old, 20±2 g) were
obtained from the Laboratory Animal Centre of Qingdao University
Medical College. The mice were randomly divided into 4 groups, with
6 mice in each group: PBS group (A), OVA-induced asthma group (B),
OVA+miR-410 mimic group (C), OVA+miR-410 control group (D). All
procedures involving animals were approved by the Laboratory Animal
Centre of Qingdao University Medical College Animal Care and Use
Committee and conformed to guidelines for the Care and Use of
Laboratory Animals of the Ministry of Health, China.
Luciferase experiment
We used dual-luciferase reporter assay to explore
whether miR-410 targets IL-4 and/or IL-13 mRNA. The wild-type
murine IL-4 and IL-13 mRNA 3′-UTR segments were amplified and
cloned into the psiCHECK vector (Sangon Biotech Co., Shanghai,
China). 293T cells were transfected with 0.1 µg of
psiCHECK-IL-13-mutant, psiCHECK-IL-4-mutant, psiCHECK-IL-13 or
psiCHECK-IL-4 along with 40 nM miR-410 mimic or miR-410 control
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The reporter gene assay was performed 48 h
after transfection using the Dual luciferase assay kit (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
instructions.
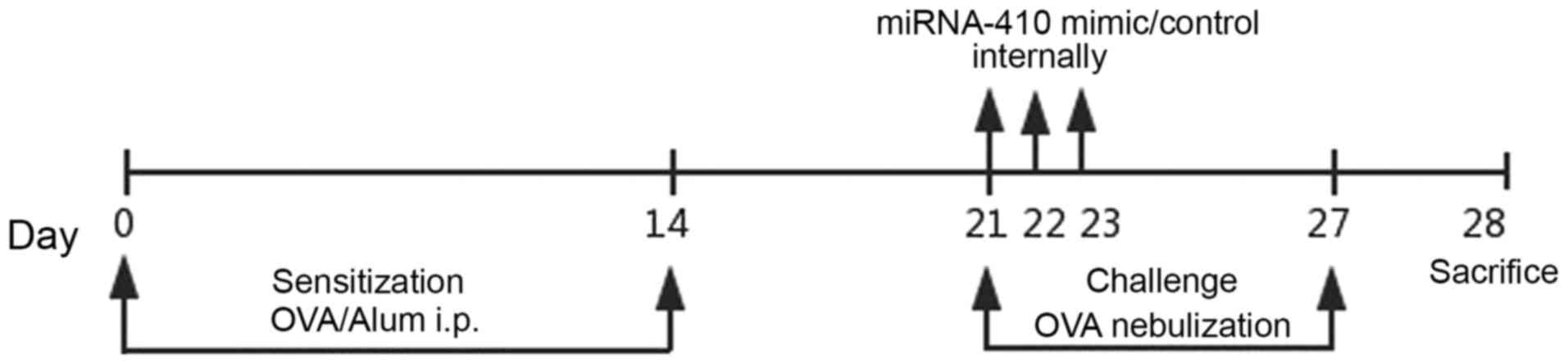
Establishing the OVA-induced asthmatic
murine model and intranasal administration of miR-410 in asthmatic
mice
Female BALB/c mice weighing ~20 g were randomly
divided into four groups (n=6). Mice were maintained in a
controlled environment and fed standard food pellets and water.
After 1 week of adaptation, mice were injected intraperitoneally on
days 0 and 14 with either 20 µg OVA (Grade V; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and 20 mg Al(OH)3 in 0.2 ml
phosphate-buffered saline (PBS), or PBS only (control). Following
sensitisation, mice were exposed to either aerosolised 1% OVA/PBS
or PBS only for 20 min once daily on days 21–27 (19). The miR-410 mimic and a control
oligonucleotide were chemically synthesised and specially modified
for transfection into animals by Biomics Biotechnologies Co., Ltd.
(Shanghai, China). For both the OVA+miR-410 mimic group and
OVA+miR-410 control group, miR-410 or its control oligonucleotide
was dissolved in endotoxin-free water. Each working dilution was
administered intranasally into asthmatic mice at a dose of 10 µg on
days 21, 22 and 23. Mice were sacrificed after intraperitoneal
administration of 1% pentobarbital sodium (50 mg/kg, P3761;
Sigma-Aldrich; Merck KGaA) on day 28 (Fig. 1). Mice did not become severely ill
or moribund at any point during the experiment. All mice were
raised in a specific pathogen-free facility and sacrificed by
cervical dislocation which was following the guidelines for the
Care and Use of Laboratory Animals of the Ministry of Health,
China.
Bronchoalveolar lavage fluid
analysis
BALF was centrifuged at 240 × g for 5 min. The
supernatant was collected and stored at −80°C prior to measurement
of cytokine levels. Precipitated cells were resuspended in 0.4 ml
PBS and 0.1 ml was taken to determine the total cell count. The
remaining cells were smeared onto a clean slide and a differential
count was performed following Wright-Giemsa staininging. For
Wright-Giemsa staininging, two or three drops of resuspension
solution of BALF were spread onto the microscope slides, heated,
stained with Wright-Giemsa staining for 10 min, and subsequently
washed under running tap-water for 2 min (20). IL-4 and IL-13 levels were measured
using enzyme-linked immunosorbent assays (ELISAs) (eBioscience;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions.
RNA extraction and quantitative
polymerase chain reaction (PCR)
Total lung RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). Primers
amplifying miR-410, IL-4 and IL-13 were purchased from Sangon
Biotech Co. (Shanghai, China). mRNA levels were normalised to GAPDH
levels and quantitated using the 2−ΔΔCq method (21). PCR was performed on a 7500 Fast
Real-Time PCR System (Fermentas; Thermo Fisher Scientific,
Inc.).
Histopathology
Mouse lungs were removed 24 h after the final
challenge. Left lungs were immediately fixed in 10% (v/v) buffered
formalin and embedded in paraffin. Lung sections were stained with
hematoxylin and eosin (H&E) (Baso, Zhuhai, China) to detect
eosinophil infiltration, as described previously (20).
Statistical analysis
The results were analysed with GraphPad Prism
software (version 5.0; GraphPad Software, Inc., La Jolla, CA, USA)
and are expressed as means ± standard deviation. Correlations among
the four groups were calculated using one-way analysis of variance
followed by Tukey's multiple comparison post hoc test. A P-value
<0.05 was considered statistically significant.
Results
Prediction of miR-410 target
genes
To elucidate the potential relationship between
miR-410 and asthma, we first screened for potential target genes
that contain a highly conserved complementary 3′-UTR sequence using
a publicly available bioinformatics tool (http://www.mirna.org/) (22,23).
The 3′-UTRs of both IL-4 and IL-13 mRNAs were predicted to contain
a binding position for miR-410 (Fig.
2).
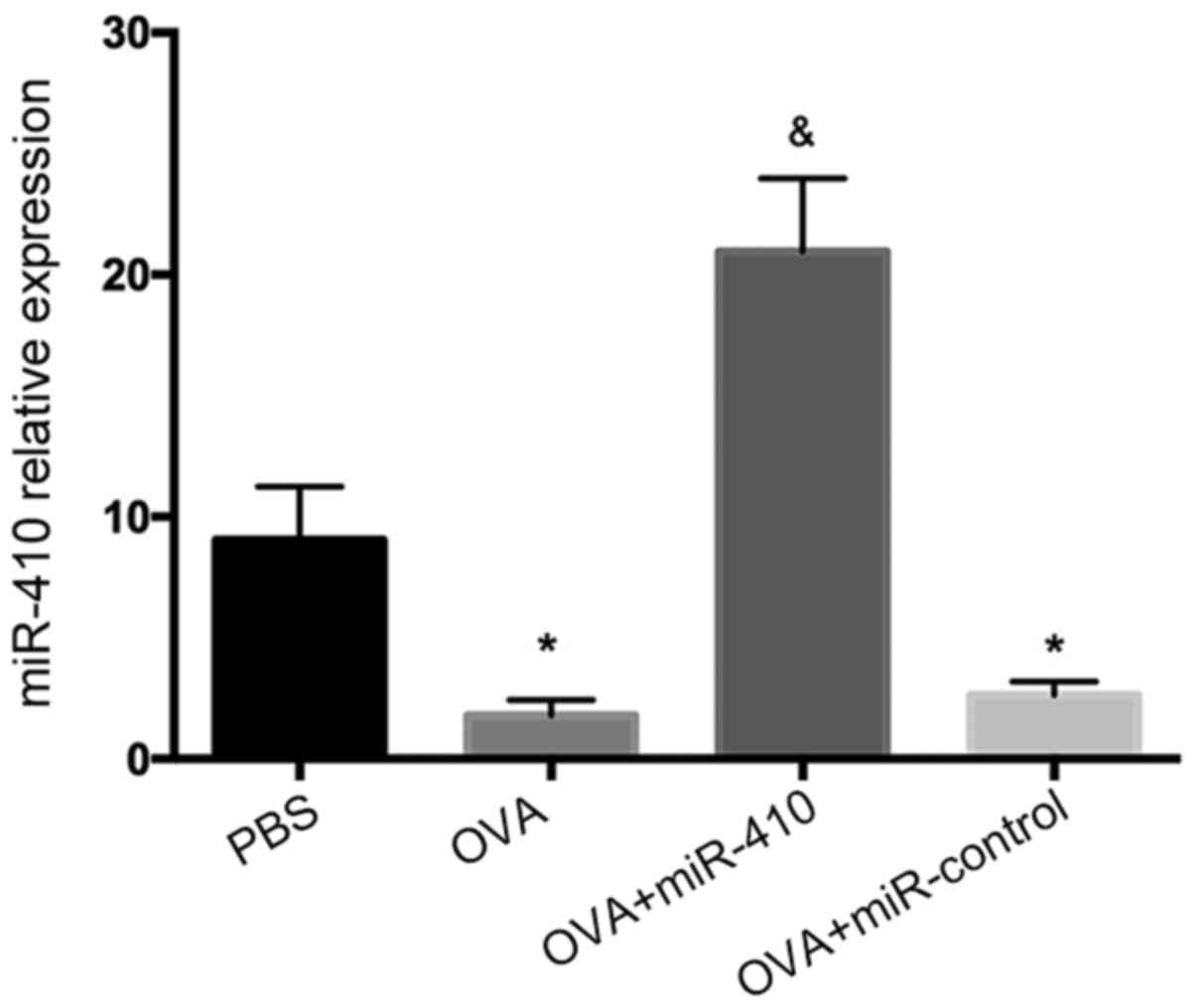
Reduced expression of miR-410 in lungs
of OVA-induced asthmatic mice
As shown in Fig. 3,
miR-410 expression was significantly decreased in the OVA-induced
asthmatic mice, suggesting that miR-410 may negatively regulate
OVA-induced asthma. To further evaluate the role played by miR-410,
we administered miR-410 intranasally, affording lung gene
transduction.
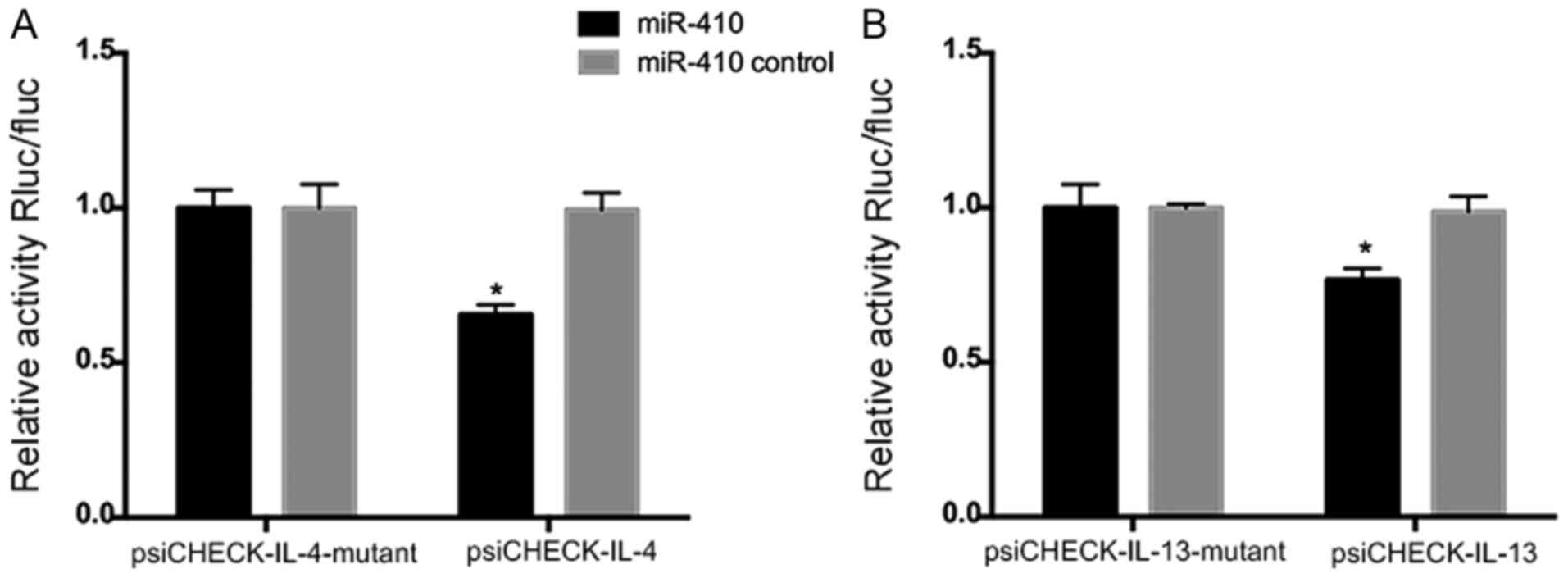
miR-410 targets IL-4 and IL-13
mRNAs
To explore whether miR-410 modulates IL-4 or IL-13
expression directly, we transfected miR-410 and the
luciferase-encoding plasmids psiCHECK-IL-4, psiCHECK-IL-13,
psiCHECK-IL-4-mutant, and psiCHECK-IL-13-mutant into 293T cells.
The luciferase activities of psiCHECK-IL-4 and psiCHECK-IL-13 were
significantly decreased in the presence of miR-410 mimics (Fig. 4A and B). Hence, modulation of IL-4
and IL-13 expression by miR-410 was sequence-specific.
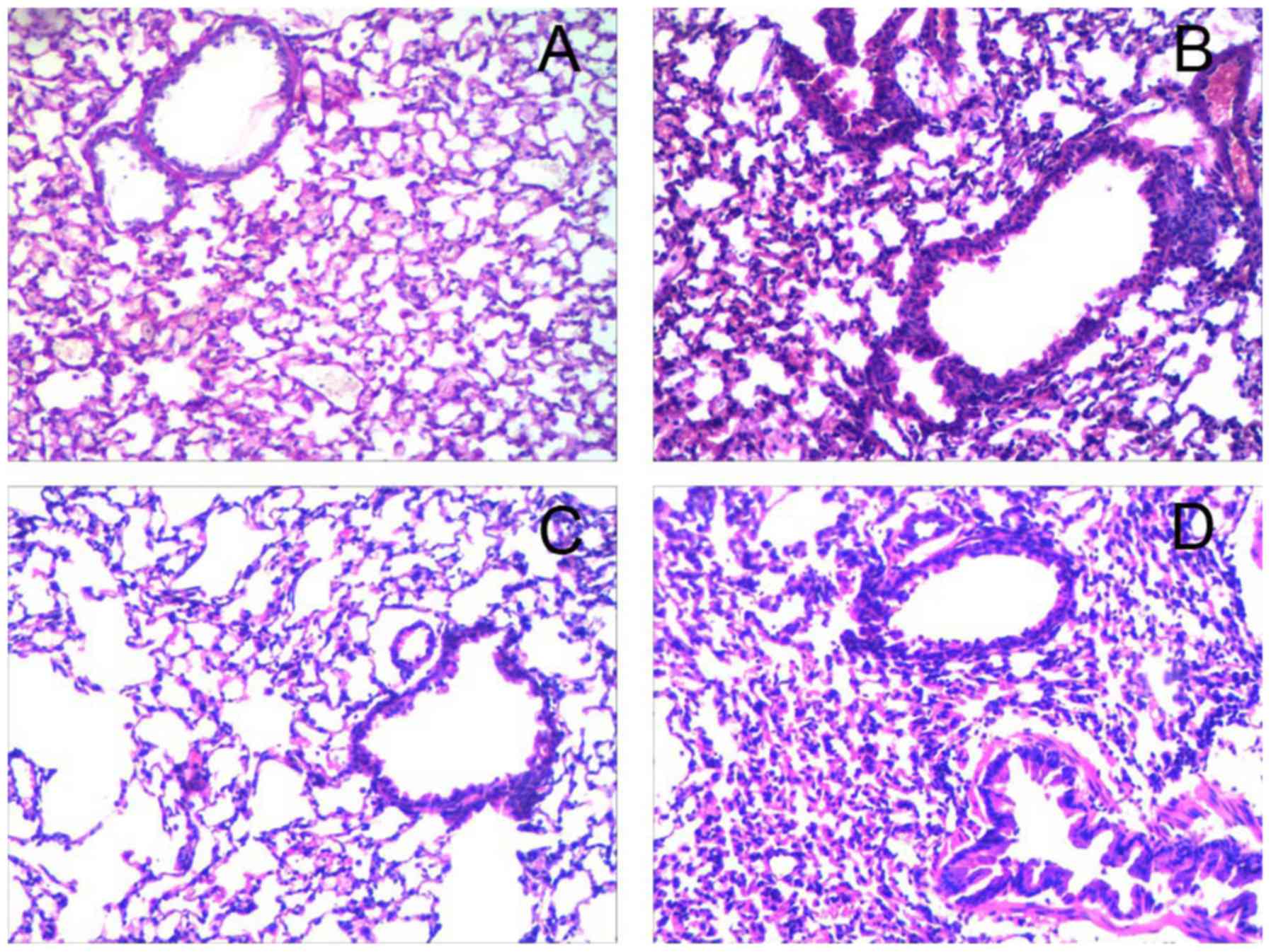
Alleviation of airway inflammation in
asthmatic mice via intranasal instillation of miR-410
Histological analysis revealed numerous inflammatory
cell infiltrations around the bronchiole and a thickened airway
epithelium in the OVA and OVA+miR-410 control groups compared with
the PBS group. However, OVA-induced asthmatic mice treated with the
miR-410 mimic exhibited a marked reduction in bronchiole
inflammatory cell infiltration (Fig.
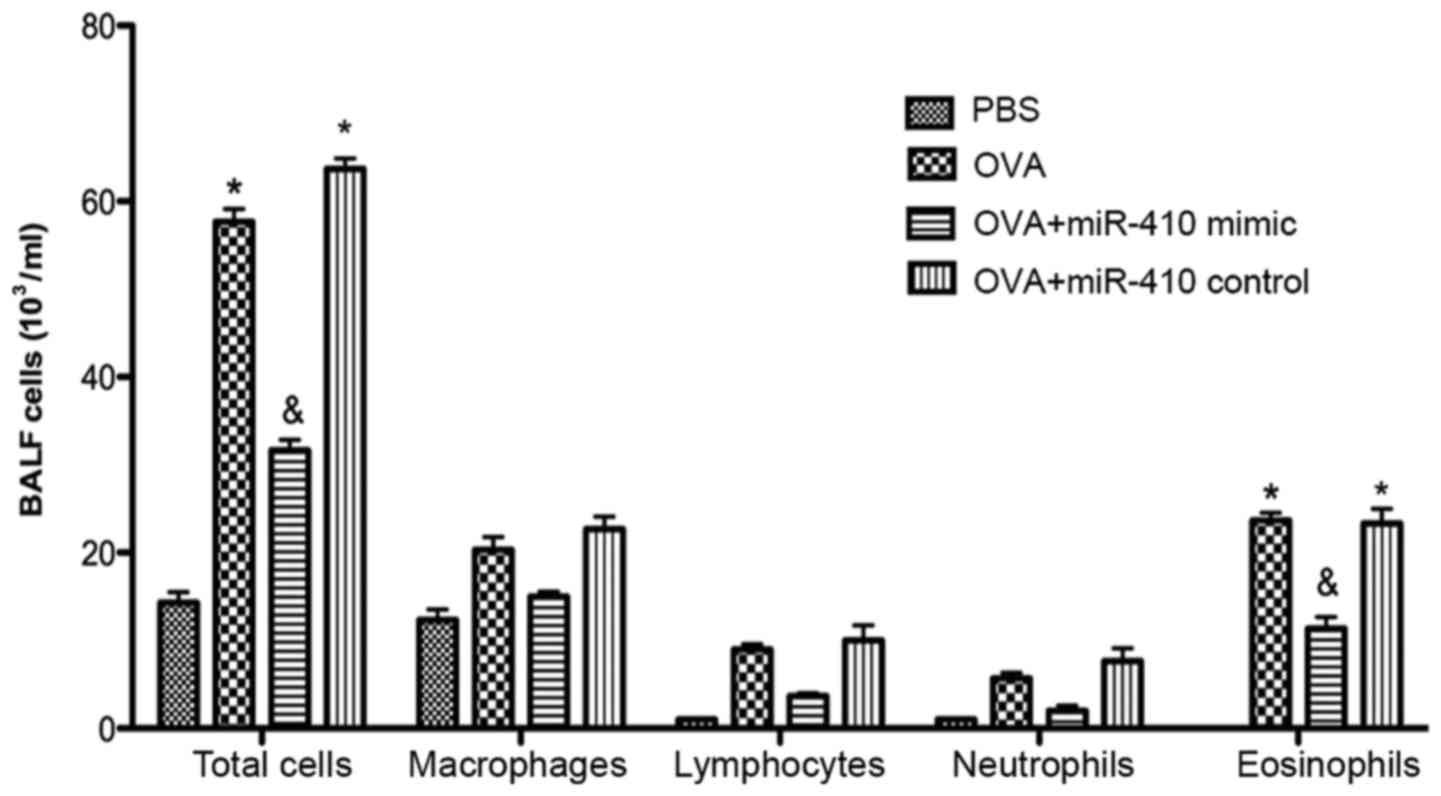
5). We examined whether miR-410 affects inflammatory cells in
BALF of OVA-induced asthmatic mice. The numbers of total cells and
eosinophils in BALF were significantly increased in the OVA-induced
mice after the last challenge, compared with the control mice.
These increases were not evident in the miR-410 mimic group
(Fig. 6).
IL-4 and IL-13 expression in lungs of
OVA-induced mice is suppressed by intranasal instillation of
miR-410
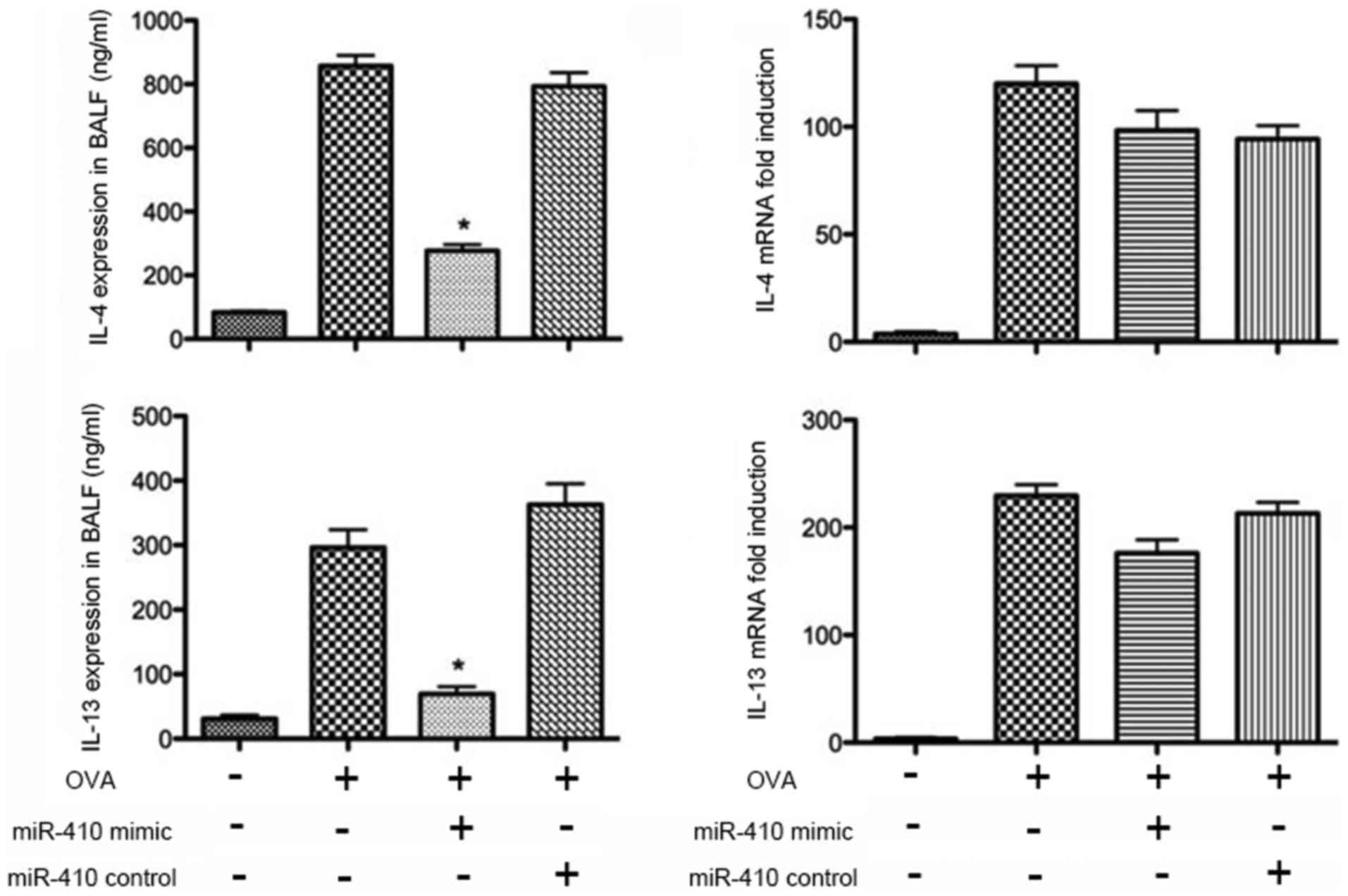
The protein levels of IL-4 and IL-13 in BALF were
determined by ELISAs. IL-4 and IL-13 were significantly increased
in the OVA group after the last challenge compared with the PBS
group. The increased levels of IL-4 and IL-13 were significantly
reduced following intranasal administration of the miR-410 mimic
(Fig. 7). We further assessed the
mRNA levels of IL-4 and IL-13 in the lung tissues; those of IL-4
and IL-13 did not differ significantly among the four groups
(Fig. 7).
Discussion
In the present study, the role of intranasal
instillation of miR-410 was evaluated in an OVA-induced murine
model of asthma. Administration of miR-410 intranasally attenuated
inflammatory cell infiltration in bronchioles of OVA-induced
asthmatic mice by directly targeting the 3′-UTRs of IL-4 and IL-13
mRNAs.
The comprehensive resource, microrna.org, was utilized for target site analysis;
this revealed that miR-410 had binding sites on the 3′-UTRs of both
IL-4 and IL −13 mRNAs. miRNA binds specifically to the 3′-UTR of
the target mRNA to suppress translation or induce degradation
(24–26). Accumulating evidence suggests that
miRNAs regulate a variety of biological processes, including
inflammation and allergic diseases (25). Abnormal expression of several
miRNAs has been observed in the airways, in BALF lymphocytes of
asthmatic patients and in several asthmatic murine models (27,28).
To date, a~10 miRNAs have been found to play a role in asthma
pathogenesis; most have been validated in house dust mite- or
OVA-induced asthma mouse models (29,30).
Accordingly, our data suggest that miR-410 levels were
significantly decreased in the OVA-induced asthmatic mice and
miR-410 overexpression reduced the expression levels and
stabilities of IL-4 and IL-13 mRNAs. The luciferase reporter assay
showed that the miR-410 sequence specifically bound to the 3′-UTRs
of IL-4 and IL-13 mRNA. Therefore, we hypothesise that the
inhibitory effects of miR-410 on the production of IL-4 and IL-13
may be direct; however, further studies are needed to confirm this
using luciferase reporter plasmids containing the 3′-UTR of STAR-6,
which plays an important role in Th2 cytokine networking (31).
Asthma is a heterogeneous disease with patients
presenting with several distinct clinical phenotypes (2,32).
The OVA-induced allergic asthma model is characterised by chronic
airway inflammation with infiltration of eosinophils, lymphocytes,
macrophages and neutrophils into the bronchial lumen (3,33,34).
OVA-induced asthma is associated with pulmonary production of Th2
cytokines (IL-4, IL-5 and IL-13) in BALF (35). Similar changes, such as
histopathological alterations in lung tissues and inflammatory
cells in BALF, were observed in our present study. As Kumar et
al previously reported, intranasal delivery of miR-let-7 to
mice with ovalbumin (OVA)-induced allergic airway inflammation
triggered overexpression of miR-let-7; others found that intranasal
miRNA administration effectively triggered miRNA overexpression in
murine models (36,37). We successfully administered miR-410
or a control mimic intranasally to OVA-induced mice.
The aberrant Th2-type response to allergens during
asthma initiation and progression is characterised by excessive
production of IL-4 and IL-13, which in turn triggers inflammatory
airway infiltration by mast cells, eosinophils and basophils
(38). IL-4 triggers the type 2
immune response whereas IL-13 is an effector molecule (39). However, pharmacological experiments
showed that inhibition of IL-4 or IL-13 alone did not adequately
attenuate the allergic inflammation associated with asthma. Thus,
further research is needed. Intranasal miR-410 exerted a marked
anti-inflammatory effect on airway inflammation in the mouse,
attributable to significant reductions in the levels of IL-4, IL-13
and eosinophils in BALF. Through dual-luciferase reporter assay, we
found that miR-410 targets the 3′-UTRs of both IL-4 and IL-13
mRNAs. Therefore, miR-410 may directly target lung IL-4/IL-13
mRNAs, reducing lung pathology via a continued decrease in Th2
cytokine levels. Thus, intranasal miR-410 may alleviate asthma;
early studies evaluating IL-4- and IL-13-targeting therapies
yielded disappointing results (39).
In conclusion, intranasal administration of miR-410
significantly suppressed airway inflammation and exerted
therapeutic effects by directly targeting post-transcriptional
expression of IL-4 and IL-13. The mechanisms underlying these
outcomes are complex, involving both direct and indirect effects on
multiple regulatory processes. However, intranasal instillation of
miR-410 may protect against asthma.
Acknowledgements
Not applicable.
Funding
The present study was funded by NSFC grant no.
81701587.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
RL designed the study. RJ and RG carried out the
animal experiments. LL and SH carried out the ELISA analysis and
data analysis. XL carried out the histopathological analysis. RJ,
LL, XL and RG carried out preparation of the manuscript. All
authors contributed to the development of interim and final drafts,
and read and approved the final manuscript. All authors agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All procedures involving animals were approved by
the Laboratory Animal Centre of Qingdao University Medical College
Animal Care and Use Committee and conformed to guidelines for the
Care and Use of Laboratory Animals of the Ministry of Health,
China.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Hu M, Ou-Yang HF, Han XP, Ti XY and Wu CG:
KyoT2 downregulates airway remodeling in asthma. Int J Clin Exp
Pathol. 8:14171–14179. 2015.PubMed/NCBI
|
|
2
|
Robinson DS: Th-2 cytokines in allergic
disease. Br Med Bull. 56:956–968. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pedersen SE, Hurd SS, Lemanske RF Jr,
Becker A, Zar HJ, Sly PD, Soto-Quiroz M, Wong G and Bateman ED:
Global Initiative for Asthma: Global strategy for the diagnosis and
management of asthma in children 5 years and younger. Pediatr
Pulmonol. 46:1–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Izuhara K, Ohta S, Shiraishi H, Suzuki S,
Taniguchi K, Toda S, Tanabe T, Yasuo M, Kubo K, Hoshino T, et al:
The mechanism of mucus production in bronchial asthma. Curr Med
Chem. 16:2867–2875. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang HY and Chiang BL: siRNA as a therapy
for asthma. Curr Opin Mol Ther. 11:652–663. 2009.PubMed/NCBI
|
|
6
|
Liu W, Liu S, Verma M, Zafar I, Good JT,
Rollins D, Groshong S, Gorska MM, Martin RJ and Alam R: Mechanism
of TH2/TH17-predominant and neutrophilic
TH2/TH17-low subtypes of asthma. J Allergy
Clin Immunol. 139:1548–1558.e4. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lynch JP, Ferreira MA and Phipps S:
Th2/Th17 reciprocal regulation: Twists and turns in the complexity
of asthma phenotypes. Ann Transl Med. 4 Suppl 1:S592016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gour N and Wills-Karp M: IL-4 and IL-13
signaling in allergic airway disease. Cytokine. 75:68–78. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gandhi NA, Pirozzi G and Graham NMH:
Commonality of the IL-4/IL-13 pathway in atopic diseases. Expert
Rev Clin Immunol. 13:425–437. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tomkinson A, Tepper J, Morton M, Bowden A,
Stevens L, Harris P, Lindell D, Fitch N, Gundel R and Getz EB:
Inhaled vs subcutaneous effects of a dual IL-4/IL-13 antagonist in
a monkey model of asthma. Allergy. 65:69–77. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Moynihan BJ, Tolloczko B, El Bassam S,
Ferraro P, Michoud MC, Martin JG and Laberge S: IFN-gamma, IL-4 and
IL-13 modulate responsiveness of human airway smooth muscle cells
to IL-13. Respir Res. 9:842008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blease K: Therapeutics targeting IL-13 for
the treatment of pulmonary inflammation and airway remodeling. Curr
Opin Investig Drugs. 9:1180–1184. 2008.PubMed/NCBI
|
|
13
|
Zhou Z, Zhao Y, Gu L, Niu X and Lu S:
Inhibiting proliferation and migration of lung cancer using small
interfering RNA targeting on Aldo-keto reductase family 1 member
B10. Mol Med Rep. 17:2153–2160. 2018.PubMed/NCBI
|
|
14
|
Qin HB, Xu B, Mei JJ, Li D, Liu JJ, Zhao
DY and Liu F: Inhibition of miRNA-221 suppresses the airway
inflammation in asthma. Inflammation. 35:1595–1599. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li JJ, Tay HL, Maltby S, Xiang Y, Eyers F,
Hatchwell L, Zhou H, Toop HD, Morris JC, Nair P, et al: MicroRNA-9
regulates steroid-resistant airway hyperresponsiveness by reducing
protein phosphatase 2A activity. J Allergy Clin Immunol.
136:462–473. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dissanayake E and Inoue Y: MicroRNAs in
allergic disease. Curr Allergy Asthma Rep. 16:672016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feng MJ, Shi F, Qiu C and Peng WK:
MicroRNA-181a, −146a and −146b in spleen CD4+ T lymphocytes play
proinflammatory roles in a murine model of asthma. Int
Immunopharmacol. 13:347–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen N, Wang J, Hu Y, Cui B, Li W, Xu G,
Liu L and Liu S: MicroRNA-410 reduces the expression of vascular
endothelial growth factor and inhibits oxygen-induced retinal
neovascularization. PLoS One. 9:e956652014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jin R, Guo S, Wang MY, Li YH, Wu LX, Ma H,
Lowrie DB, Fan XY and Zhang JH: Administration of mycobacterial
Ag85A and IL-17A fusion protein attenuates airway inflammation in a
murine model of asthma. Int Immunopharmacol. 17:1067–1074. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu X, Lin R, Zhao B, Guan R, Li T and Jin
R: Correlation between oxidative stress and the NF-κB signaling
pathway in the pulmonary tissues of obese asthmatic mice. Mol Med
Rep. 13:1127–1134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Xue Y, Liu Y, Song G, Lv G, Wang
Y, Wang Y, Li X and Yang L: MicroRNA-146a expression inhibits the
proliferation and promotes the apoptosis of bronchial smooth muscle
cells in asthma by directly targeting the epidermal growth factor
receptor. Exp Ther Med. 12:854–858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Yang K, Shi H, Xu J, Zhang D, Wu Y,
Zhou S and Sun X: MiR-21 modulates human airway smooth muscle cell
proliferation and migration in asthma through regulation of PTEN
expression. Exp Lung Res. 41:535–545. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ariel D and Upadhyay D: The role and
regulation of microRNAs in asthma. Curr Opin Allergy Clin Immunol.
12:49–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Solberg OD, Ostrin EJ, Love MI, Peng JC,
Bhakta NR, Hou L, Nguyen C, Solon M, Nguyen C, Barczak AJ, et al:
Airway epithelial miRNA expression is altered in asthma. Am J
Respir Crit Care Med. 186:965–974. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou H, Li J, Gao P, Wang Q and Zhang J:
miR-155: A novel target in allergic asthma. Int J Mol Sci. 17(pii):
E17732016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Deshpande DA, Dileepan M, Walseth TF,
Subramanian S and Kannan MS: MicroRNA regulation of airway
inflammation and airway smooth muscle function: Relevance to
asthma. Drug Dev Res. 76:286–295. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Al-Quraishy S, Dkhil MA, Delic D,
Abdel-Baki AA and Wunderlich F: Organ-specific
testosterone-insensitive response of miRNA expression of C57BL/6
mice to Plasmodium chabaudi malaria. Parasitol Res.
111:1093–1101. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Y, Sun E, Li X, Zhang M, Tang Z, He
L and Lv K: miR-155 contributes to Df1-induced asthma by increasing
the proliferative response of Th cells via CTLA-4 downregulation.
Cell Immunol. 314:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sharma A, Kumar M, Ahmad T, Mabalirajan U,
Aich J, Agrawal A and Ghosh B: Antagonism of mmu-mir-106a
attenuates asthma features in allergic murine model. J Appl
Physiol. 113:459–464. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nelms K, Keegan AD, Zamorano J, Ryan JJ
and Paul WE: The IL-4 receptor: Signaling mechanisms and biologic
functions. Annu Rev Immunol. 17:701–738. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stirling RG and Chung KF: Future
treatments of allergic diseases and asthma. Br Med Bull.
56:1037–1053. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jia Y, Fang X, Zhu X, Bai C, Zhu L, Jin M,
Wang X, Hu M, Tang R and Chen Z: IL-13+ type 2 innate
lymphoid cells correlate with asthma control status and treatment
response. Am J Respir Cell Mol Biol. 55:675–683. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang L, Oh SY, Wu X, Oh MH, Wu F,
Schroeder JT, Takemoto CM, Zheng T and Zhu Z: SHP-1 deficient mast
cells are hyperresponsive to stimulation and critical in initiating
allergic inflammation in the lung. J Immunol. 184:1180–1190. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Veremeyko T, Siddiqui S, Sotnikov I, Yung
A and Ponomarev ED: IL-4/IL-13-dependent and independent expression
of miR-124 and its contribution to M2 phenotype of monocytic cells
in normal conditions and during allergic inflammation. PLoS One.
8:e817742013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kumar M, Ahmad T, Sharma A, Mabalirajan U,
Kulshreshtha A, Agrawal A and Ghosh B: Let-7 microRNA-mediated
regulation of IL-13 and allergic airway inflammation. J Allergy
Clin Immunol. 128:1077–1085, e1-e10. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Z, Chen X, Wu Q, Song J, Wang L and Li
G: miR-125b inhibits goblet cell differentiation in allergic airway
inflammation by targeting SPDEF. Eur J Pharmacol. 782:14–20. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cortes JR, Rivas MD, Molina-Infante J,
Gonzalez-Nuñez MA, Perez-G M, Masa JF, Sanchez JF and Zamorano J:
Omeprazole inhibits IL-4 and IL-13 signaling signal transducer and
activator of transcription 6 activation and reduces lung
inflammation in murine asthma. J Allergy Clin Immunol. 124:607–610,
610.e1. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Karo-Atar D, Bitton A, Benhar I and Munitz
A: Therapeutic targeting of the interleukin-4/interleukin-13
signaling pathway: In allergy and beyond. BioDrugs. May
7–2018.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|