Introduction
Age-related cataract (ARC) is one of the major
causes of visual impairment or visual loss. Despite various
in-depth investigations on ARC in previous years, surgical
replacement with an artificial lens remains the only cure for ARC
due to its complicated pathogenesis and unclear mechanism.
Therefore, it is of importance to further examine the pathogenic
mechanism of ARC, and to develop novel therapies for its prevention
and treatment.
Lens epithelial cells can differentiate into lens
fibers, which are important in the biological function of the lens.
Previous studies have found that, during the development of ARC,
several genes undergo structural or functional alterations
(1). This leads to the apoptosis
of lens epithelial cells and degeneration of lens proteins and
ultimately causes all types of cataract, with the exception of
congenital cataracts (2).
MicroRNAs (miRNAs) are a small type of RNA molecule that are
extensively present in animals and plants and are involved in
post-transcriptional gene regulation. They are also involved in a
variety of other biological activities, including cell
proliferation, differentiation and apoptosis (3). Previous studies have identified miRNA
(miR)-15a as an active contributor in the apoptosis of several cell
types (4,5). miR-15a is the natural antisense
interfering RNA of the proto-oncogene B-cell lymphoma 2 (BCL2). The
nine bases at the 5′end of miR-15a are complementary to bases
3,287-3,279 of BCL2 (4). Their
combination results in downregulation of the expression of BCL2 and
thus promotes cell apoptosis. As another member of the BCL2 family,
the expression of myeloid cell leukemia sequence 1 (MCL1) can also
be inhibited by miR-15a, which promotes apoptosis (6). Our previous study identified elevated
expression of miR-15a-3p in the lens epithelial cells of patients
with ARC (7). Is it possible that
the interaction between miR-15a-3p and anti-apoptotic proteins BCL2
or MCL1 leads to the apoptosis of lens epithelial cells and altered
proliferation and differentiation of lens epithelial cells,
ultimately causing ARC. To confirm this hypothesis, the present
study transformed miR-15a-3p mimic into the HLE-B3 human lens
epithelial cell line and examined its proliferation, migration and
apoptosis. The study also aimed to reveal the role of miR-15a-3p in
the pathogenesis of ARC.
Materials and methods
Ethics
The present study was approved by the Institutional
Review Board of Yantai Yuhuangding Hospital (Yantai, China).
Cell culture and transfection
The HLE-B3 cells were obtained from Shanghai
Genechem Co., Ltd. (Shanghai, China) and were maintained in DMEM
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% FBS (Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA), under a 37°C humidified, 5% CO2
environment. The cells were transfected with miR-15a-3p mimic,
miR-15a-3p mock or mimic negative control (NC) from GenePharma
(Shanghai, China) using Lipofectamine RNAiMax (Invitrogen; Thermo
Fisher Scientific, Inc.), following the manufacturer's protocol.
After 48 h, the cells were harvested for subsequent
experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The miRNA expression levels were determined by
RT-qPCR analysis. In brief, total RNA was extracted from the
cultured cells using an RNA extraction kit (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Then, 2
µg total RNA was reverse transcribed using ReverTra Ace (Toyobo
Life Science, Osaka, Japan). RT-qPCR analysis for miR-15a-3p was
conducted using the ABI7500 (Applied Biosystems; Thermo Fisher
Scientific, Inc.) in accordance with the manufacturer's protocol.
The PCR amplifications were performed using a 50 µl final reaction
mixture containing 25 µl SYBR Green PCR Master mix (Platinum SYBR
Green qPCR SuperMix-UDG kit, Life Technologies; Thermo Fisher
Scientific, Inc.), 1.5 µl primer mix (10 µM) and 1 µl template.
Cycling conditions were: 50°C for 2 min; 95°C for 5 min; followed
by 40 cycles of 95°C, 10 sec and 60°C, 45 sec. The melting curve
analysis and agarose gel electrophoresis were used to confirm the
specific PCR products. The expression of RNA U6 small nuclear 2
(RNU6B) was also measured as an internal control. The primer
sequences were as follows: miR-15a-3p RT,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTGAGGCA-3′; miR-15a-3p,
forward 5′-ACACTCCAGCTGGGCAGGCCATATTGTGCTGCCTC-3′ and reverse
5′-TGGTGTCGTGGAGTCG-3′; RNU6B, forward 5′-CTCGCTTCGGCAGCACA-3′ and
reverse 5′-AACGCTTCACGAATTTGCGT-3′. Product specificity was
confirmed by melting curve analysis. All experiments were performed
in triplicate with three separate samples. All results were
determined as the expression of miR-15a-3p relative to that of the
internal control in cell lines using the 2−ΔΔCq method
(8).
Cell proliferation assay
MTT assays were performed to assess the
proliferative ability of the transfected HLE-B3 cells. In each well
of a 96-well plate, 3×103 cells were seeded and, 24 h
later, were transfected with 5 pmol miR-15a-3p mimic, miR-15a-3p
mock or mimic NC (GenePharma). At 0, 24, 48, 72 and 96 h following
transfection, 20 µl of MTT (5 mg/ml, Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was added to the wells and incubated at 37°C
for 2 h. Subsequently, the culture medium was replaced with 150 µl
of dimethylsulfoxide (Sigma-Aldrich; Merck KGaA) and then agitated
at 0.04 × g for 15 min at room temperature. The optical density
(OD) value of each well was then evaluated using the ELISA
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
at a wavelength of 490 nm. The experiments were performed in
triplicate and repeated at least three times.
Plate colony formation
experiments
Cells (1×104) in logarithmic growth phase
were seeded onto 6-cm plates and cultured for 2–3 weeks. When
visible colonies had appeared, the cells were washed with PBS
twice, fixed in methanol for 15 min, stained with crystal violet
for 20 min, washed with water, and air dried. The number of visible
colonies was counted under an inverted microscope (IX51; Olympus
Corporation, Tokyo, Japan) using a transparent film with grids.
Colony formation rate=(number of colonies/number of inoculated
cells) ×100%.
Cell apoptotic rate assay
Annexin V-FITC/propidium iodide (PI) staining was
conducted to assess the apoptotic rate of cells using flow
cytometry (BD Biosciences, Franklin Lakes, NJ, USA). Briefly,
following transfection, the HLE-B3 cells (1×106) were
collected and stained with Annexin V-FITC/PI using an apoptosis
detection kit (BD Biosciences). Following 15 min of incubation at
room temperature in the dark, the apoptotic cell percentages were
measured through flow cytometry. The Annexin V immunofluorescence
was presented on the x-axis and the plasma membrane integrity was
presented on the y-axis. Each experiment was conducted in
triplicate.
Wound-healing assay
The cells were inoculated in six-well plates and
routinely cultured until they reached 70–80% confluence. Cross
lines were introduced using a 200-µl sterile pipette tip. The cells
were washed three times with sterile PBS to remove the cells from
the scratched region and were then continually cultured in
serum-free culture medium. Images of the cells were captured at 0
and 24 h post-wounding. Cell migration distance=distance at 0
h-distance at 24 h.
Transwell assay
Transwell chamber inserts (BD Biosciences) with (for
invasion) or without (for migration) Matrigel (BD Biosciences) were
used. The cells were transfected with miR-15a-3p mimics,
inhibitors, mimic NC or inhibitor NC. At 6 h following
transfection, 1×104 transfected cells in 200 µl
serum-free DMEM were seeded in the upper chamber of the insert. The
bottoms of the inserts were incubated in medium with 10% FBS.
Following 36 h (for migration) or 48 h (for invasion), cells on the
upper membrane were removed using a cotton swab. Those cells that
had migrated or invaded to the bottom of the inserts were stained
with crystal violet (1 mg/ml, Sigma-Aldrich; Merck KGaA) for 15 min
at room temperature and counted using a Leica DMIRB inverted
microscope (×200 magnification). The experiments were performed in
triplicate and repeated at least three times.
Western blot analysis
Western blot analysis was performed as previously
described (9). Total proteins were
extracted in a lysis buffer (20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 5
mM EDTA, pH 8.0, 5 mM NaPPi, 1 mM sodium orthovanadate
(Na3VO4), 1 mM PMSF, 1% NP-40, and 10 µg/ml
each aprotinin and leupeptin). Tissue lysates were centrifuged at
10,000 g for 10 min at 4°C. Sample concentration was detected by
BCA. Then 20 µg protein was subjected to 10% SDS-PAGE, transferred
onto PVDF membranes and followed by immunoblotting. Following
blocking with TBS containing 5% bovine serum albumin (BSA;
Sigma-Aldrich; Merck KGaA) and 0.1% Tween-20, the membrane was
incubated with appropriate primary antibodies at 4°C overnight,
followed by incubation with a peroxidase-conjugated goat anti-mouse
IgG (1:5,000; ZB-2305, ZSGB-BIO, Beijing, China) at room
temperature for 1 h. GAPDH was used as a loading control. The
primary antibodies used were as follows: Anti-BCL2 (1:1,000;
sc-509, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA),
anti-MCL1 (1:1,000; sc-53951, Santa Cruz Biotechnology) and
anti-GAPDH (1:1,000; sc-293335, Santa Cruz Biotechnology, Inc.) For
quantification, densitometric analysis was performed by analyzing
the grey level intensity of target bands derived from scanned
films, processed using ImageJ version 1.47 (National Institutes of
Health, Bethesda, MD, USA).
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay
The cells (100 µl of 2×104 cells) were
added into each well of a 96-well plate and incubated overnight,
followed by cell transfection. Subsequently, the cells were
cultured in conventional conditions for 48 h. The medium was
discarded and the cells were fixed with 4% paraformaldehyde for 30
min prior to washing with PBS. Following the addition of 0.3%
H2O2-methanol solution, the cells were
incubated at room temperature for another 30 min. Following PBS
washing, 0.1% Triton X-100 in 0.1% sodium citrate solution was
added into wells for 2 min incubation in an ice-bath to
permeabilize cells, followed by PBS washing. With the addition of
50 µl TUNEL reagent (In Situ Cell Death Detection kit,
Fluorescein; Roche Diagnostics, Indianapolis, IN, USA), the cells
were cultured in a humidified incubator at 37°C for 60 min.
Following three PBS washes, 50 µl of 4′,6-diamidino-2-phenylindole
was added to cells, followed by incubation at 37°C in the dark and
another three PBS washes. The cells were examined under a
fluorescence microscope and images were captured.
Statistical analysis
All statistical analyses were performed with SPSS
for Windows version 18.0 (SPSS, Inc., Chicago, IL, USA). Data are
presented as the mean ± standard deviation (SD). Error bars
represent the SD of three independent experiments. Differences
between groups were compared using one-way analysis of variance
with a post hoc test (LSD). All statistical analyses were
two-sided, and P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression of miR-15a-3p in
transfected HLE-B3 cells causes morphological changes
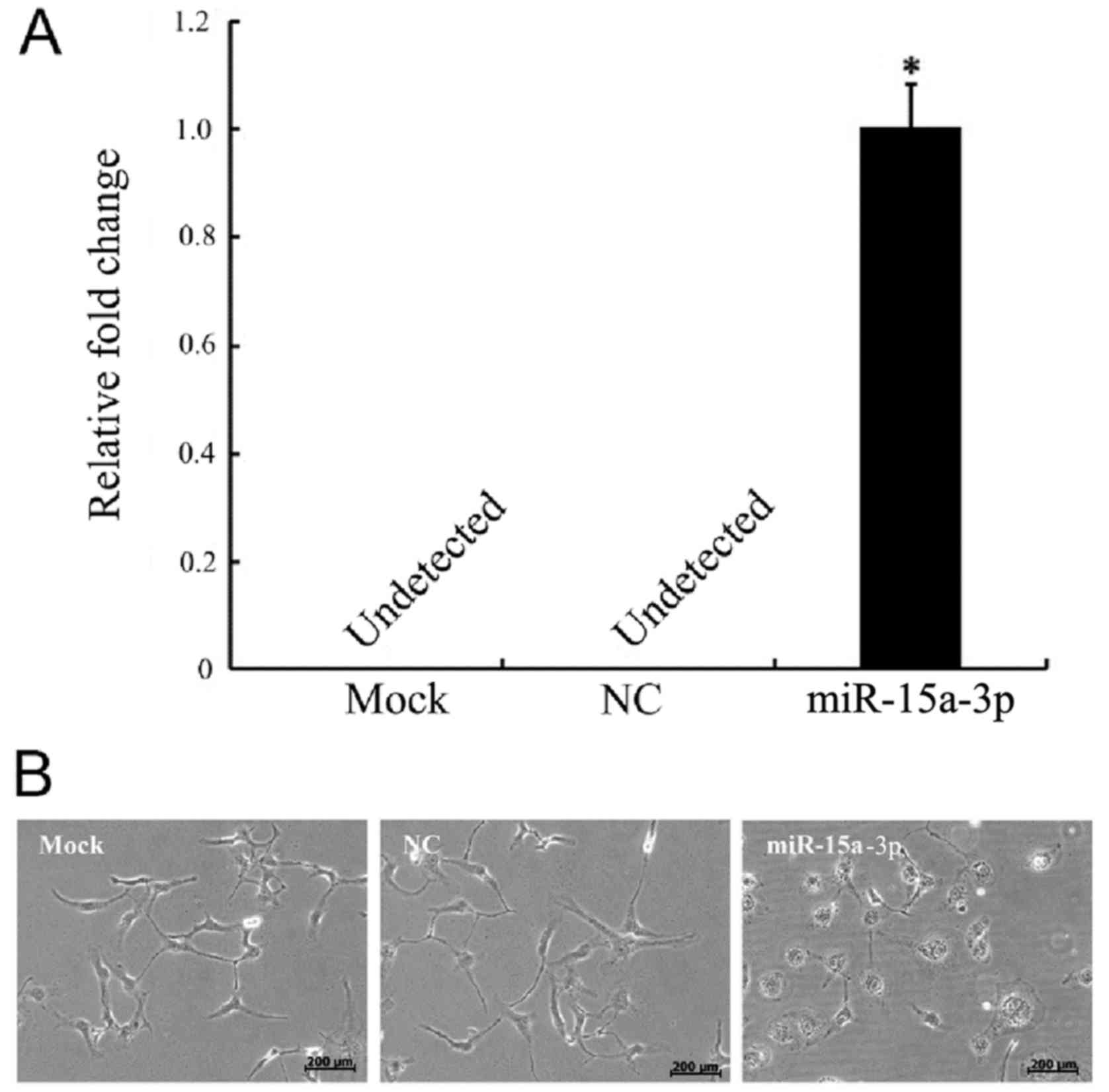
The HLE-B3 cells were transfected with miR-15a-3p
mimic (miR-15a), miR-15a-3p mock and mimic NC. No expression of
miR-15a-3p was present in the NC cells, as detected by RT-qPCR
analysis, whereas there was significantly elevated expression of
miR-15a-3p in the miR-15a-3p mimic-transfected cells (Fig. 1A).
Morphological changes occurred in the miR-15a-3p
cells. The normal HLE-B3 cells were irregular polygon-shaped with
interactions that formed network structures. The morphology of the
miR-15a-3p mimic-transfected cells began changing from 24 h
post-transfection. The cells became large and round, with unclear
boundaries and reduced networks (Fig.
1B).
Expression of miR-15a-3p inhibits
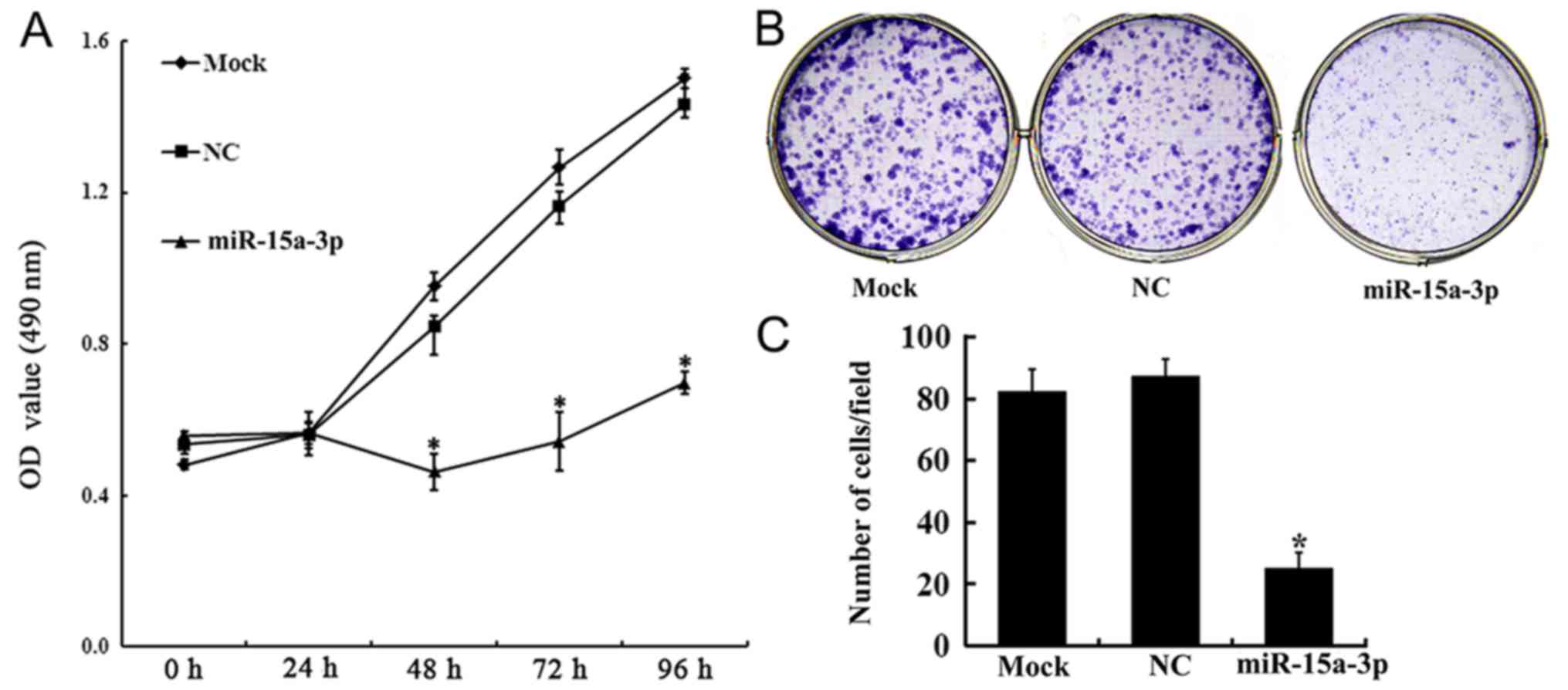
HLE-B3 cell proliferation
The effect of the expression of miR-15a-3p on HLE-B3
cell proliferation was investigated using an MTT assay. From 48 h
post-transfection, cell proliferation in the miR-15a group was
increasingly inhibited. On days 3, 4 and 5 post-transfection, the
proliferation rates of the miR-15a cells were significantly lower
than those of the Mock and NC cells (P<0.01) (Fig. 2A).
The proliferation of transfected HLE-B3 cells was
further confirmed using a plate colony formation assay. The results
showed that, compared with the NC and mock cells, the
colony-forming ability of the miR-15a cells was significantly
attenuated (Fig. 2B and C).
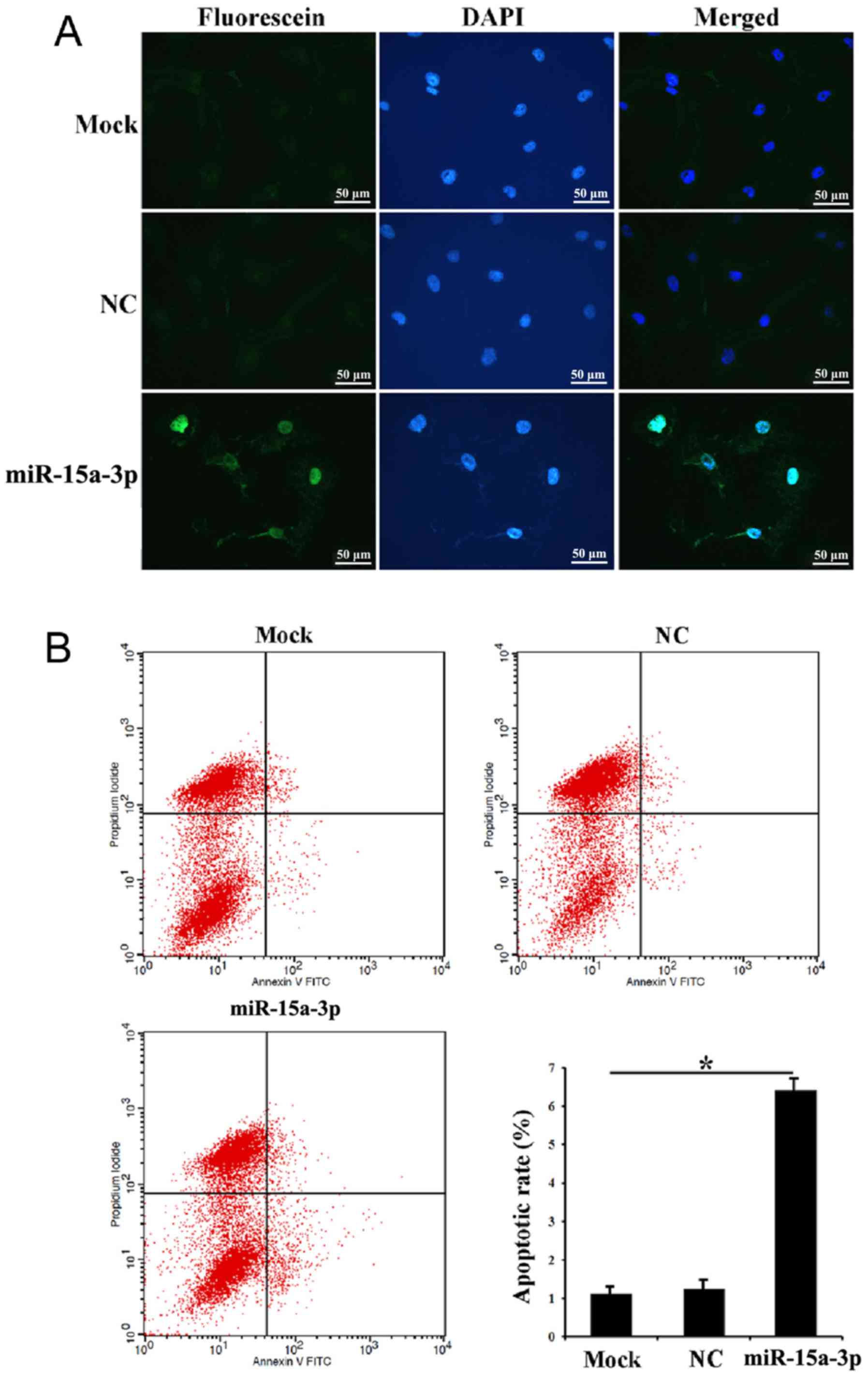
Expression of miR-15a-3p induces
HLE-B3 cell apoptosis
The in situ apoptosis of the transfected
HLE-B3 cells was investigated using a TUNEL assay. The results
showed that evident apoptotic signals were detected in the miR-15a
cells compared with the mock and NC cells, suggesting that
miR-15a-3p may induce HLE-B3 cell apoptosis (Fig. 3A). Following Annexin V-FITC/PI
double staining and flow cytometry, a significantly higher ratio of
early apoptotic cells was found in the miR-15a cells than in the
mock and NC cells. There was also more cell debris in the miR-15a
cells than the NC cells (Fig.
3B).
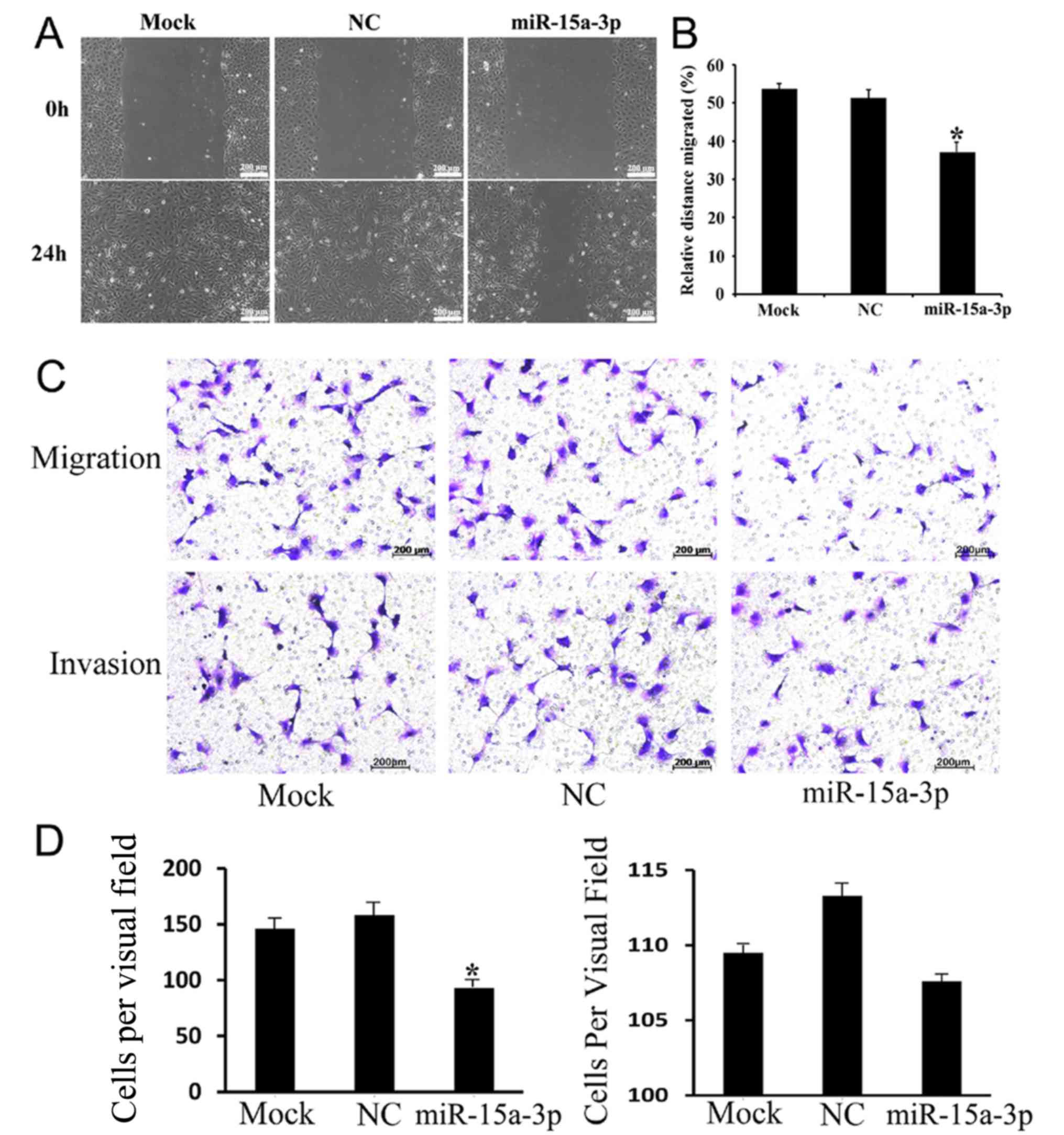
Expression of miR-15a-3p inhibits
HLE-B3 cell migration
To further investigate the effect of miR-15a-3p on
HEL-B3 cell mobility in vitro, wound-healing and Transwell
assays were performed. As the results of the wound-healing assay
showed, the miR-15a cells were less migratory than the mock and NC
cells (Fig. 4A and B). The
Transwell migration and invasion assays revealed that the
expression of miR-15a-3p inhibited the migration of HLE-B3 cells
but did not affect their invasiveness to a significant level
(Fig. 4C and D). These
observations suggested that miR-15a-3p is important in reducing the
migration of HLE-B3 cells in vitro.
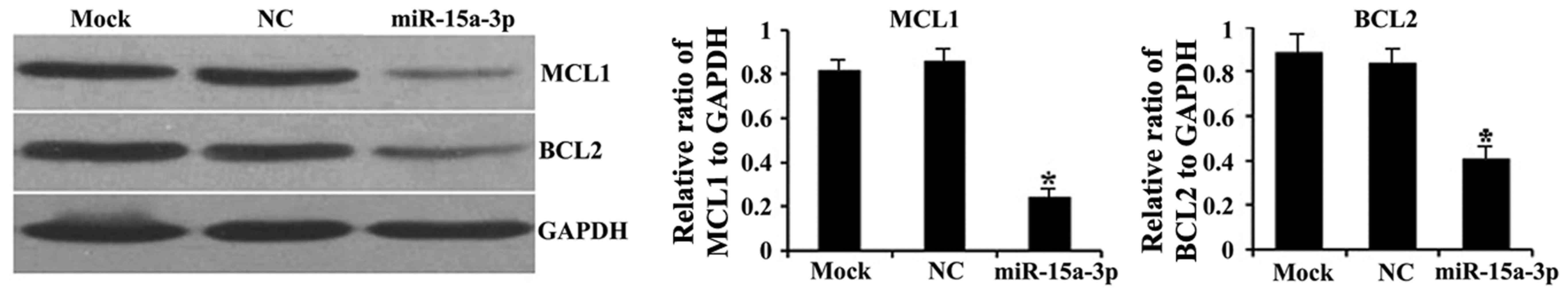
miR-15a-3p downregulates the
expression of BCL2 and MCL1
To determine whether miR-15a-3p affects the
intracellular expression of BCL2 and MCL1, western blot assays were
performed on the transfected HLE-B3 cells. Significantly lower
protein expression levels of BCL2 and MCL1 were observed in the
miR-15a cells than in the mock and NC cells (P<0.01; Fig. 5).
Discussion
ARC is one of the major causes of eye disease
leading to blindness (10,11). The apoptosis of lens epithelial
cells is the cellular basis for all types of cataracts, with the
exception of congenital cataracts (2). However, the mechanism of lens
epithelial cell apoptosis remains to be fully elucidated.
There have been reports suggesting that miRNAs are
involved in the apoptosis of lens epithelial cells (12,13).
Peng et al identified the expression of miR-let-7b in lens
epithelial cells from patients with ARC, and found that
miRNA-let-7b was closely associated with the occurrence and
development of ARC (14). Wu et
al found no expression of miRNA-923 or miRNA-let-7b in normal
human lens epithelial cells but significantly elevated expression
of both in lens epithelial cells from patients with ARC (15). Previous studies have shown that
miR-15a-3p is located at chromosome 13q14 and that the deletion or
downregulation of miR-15a-3p was associated with the origin and
development of multiple types of cancer, including chronic
lymphocytic lymphoma, prostate carcinoma and pituitary adenomas
(6,16,17).
In addition, miR-15a-3p is known to act as a tumor suppressor,
which can inhibit cell proliferation, promote the apoptosis of
cancer cells, and suppress tumorigenesis in vitro and in
vivo (18). However, the
function of miR-15a-3p in ARC has not been reported.
Our preliminary RNA microarray identified apparently
upregulated expression of miR-15a-3p in lens epithelial cells from
patients with ARC (data not shown). In addition, 90 patients with
clinical ARC were surveyed and found to have significantly
upregulated expression levels of miR-15a-3p than normal individuals
without ARC (7). Therefore, it was
hypothesized that miR-15a-3p may be involved in the apoptosis of
lens epithelial cells; however, its possible mechanism remained to
be investigated.
miR-15a regulates the apoptosis and proliferation of
cells by functioning in the regulation of multiple intracellular
signaling pathways (19,20). It was previously reported that
miR-15a downregulated cyclin D1 and induced apoptosis and cell
cycle arrest in osteosarcoma (21). In addition, the overexpression of
miR-15a inhibited the growth and survival of multiple myeloma
cells, suppressed cell migration and arrested cells at the G1 phase
(22).
In normal lens epithelial cells, the expression of
miR-15a is low; therefore, in the present study, lens epithelial
cells were transfected with miR-15a-3p mimics in order to examine
its biological function in lens epithelial cells. The results
revealed increased apoptosis, and inhibition of proliferation and
migration in HLE-B3 cells expressing miR-15a-3p mimics. The in
vitro experiments suggested that miR-15a-3p may cause cataracts
through promoting the apoptosis of lens epithelial cells.
The present study also examined the protein
expression of two BCL2 family members. The BCL2 family is an
important anti-apoptotic gene family, which mainly includes BCL2
BCL2-like 1, MCL1, BCL2-associated death promoter, and
BCL2-associated X protein (23).
This family can suppress cell apoptosis either through regulating
cellular signal transduction by directly or indirectly affecting
the release of Ca2+ in endoplasmic reticulum, or through
suppressing lipid oxidation by oxygen free radicals (24). A previous study indicated that
miR-15a was an important regulator of BCL2 (25). The 3′UTR of the BCL2 transcript has
potential binding sites for miR-15a (25). Currently, investigations on BCL2
and MCL1 regulation by miR-15a are mainly focused on leukemia and
malignant tumors (4,26). Cimmino et al found that, in
chronic lymphocytic leukemia, the expression of miR-15a was
negatively correlated with BCL2, and that miR-15a negatively
regulated BCL2 at the post-transcription level (4). In the present study, overexpressed
miR-15a-3p mimics downregulated the expression level of BCL2 and
MCL1. Our preliminary study also found a negative correlation
between the level of miR-15a-3p and the expression of BCL2 and MCL1
in lens epithelial cells of patients with ARC (7). These results demonstrate that
miR-15a-3p may induce the apoptosis of lens cells by downregulating
the expression of BCL2 and MCL1.
One limitation of the present study was that the
function and mechanism of miR-15a were only investigated in a
single lens epithelial cell line. It remains unclear whether
miRNA-15a has the same role in other lens epithelial cell lines. It
would be beneficial to examine the involvement of miR-15a in the
pathogenesis of cataracts using multiple lens cell lines in the
future. In addition, the target gene of miR-15a was not verified,
therefore, future investigations aim to confirm that BCL2 is a
direct target of miR-15a. In addition, based on the results of our
previous study (7), the role of
hsa-miR-16-1-5p in human lens epithelial cells is currently under
investigation.
In conclusion, the present study demonstrated that
the overexpression of miR-15a-3p in vitro attenuated the
proliferation of lens epithelial cells, and downregulated
anti-apoptotic proteins BCL2 and MCL1, which may result in the
apoptosis of lens epithelial cells. This study revealed a potential
mechanism of miR-15a-3p regulating ARC and provide novel ideas for
the effective prevention of cataracts.
Acknowledgements
Not applicable.
Funding
This study was funded by the Shandong Natural
Science Foundation of China (grant no. ZR2015HL052), the Yantai
Technology Development Plan (grant nos. 2017WS109, 2017YD005,
2013WS204, and 2010148-24), the Natural Science Foundation of China
(grant no. 81701453) and the Youth Scientific Research Fund of
Yantai Yuhuangding Hospital (grant no. 201502).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YBL and SJL were responsible for the concept and
framework of the paper. SJL and WTW wrote the manuscript which was
revised by YBL and NL. SJL and NL planned and performed the
experiments. FLZ and YHY carried out RT-qPCR and western blotting.
WTW, HJY and YL carried out cell proliferation, apoptosis,
migration. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of Yantai Yuhuangding Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ARC
|
age-related cataract
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
RNU6B
|
RNA U6 small nuclear 2
|
|
OD
|
optical density
|
|
TUNEL
|
terminal deoxynucleotidyl transferase
dUTP nick end labeling
|
|
PI
|
propidium iodide
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
|
SD
|
standard deviation
|
References
|
1
|
Ruotolo R, Grassi F, Percudani R, Rivetti
C, Martorana D, Maraini G and Ottonello S: Gene expression
profiling in human age-related nuclear cataract. Mol Vis.
9:538–548. 2003.PubMed/NCBI
|
|
2
|
Li WC, Kuszak JR, Dunn K, Wang RR, Ma W,
Wang GM, Spector A, Leib M, Cotliar AM and Weiss M: Lens epithelial
cell apoptosis appears to be a common cellular basis for
non-congenital cataract development in humans and animals. J Cell
Biol. 130:169–181. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cimmino A, Calin GA, Fabbri M, Iorio MV,
Ferracin M, Shimizu M, Wojcik SE, Aqeilan RI, Zupo S, Dono M, et
al: miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl
Acad Sci USA. 102:13944–13949. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Calin GA, Cimmino A, Fabbri M, Ferracin M,
Wojcik SE, Shimizu M, Taccioli C, Zanesi N, Garzon R, Aqeilan RI,
et al: miR-15a and miR-16-1 cluster functions in human leukemia.
Proc Natl Acad Sci USA. 105:5166–5171. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aqeilan RI, Calin GA and Croce CM: miR-15a
and miR-16-1 in cancer: Discovery, function and future
perspectives. Cell Death Differ. 17:215–220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Y, Liu S, Zhang F, Jiang P, Wu X and
Liang Y: Expression of the microRNAs hsa-miR-15a and hsa-miR-16-1
in lens epithelial cells of patients with age-related cataract. Int
J Clin Exp Med. 8:2405–2410. 2015.PubMed/NCBI
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin C, Wang J, Wang Y, Zhu P, Liu X, Li N,
Liu J, Yu L and Wang W: GRP78 Participates in PCA3-regulated
prostate cancer progression. Anticancer Res. 37:4303–4310.
2017.PubMed/NCBI
|
|
10
|
West S: Epidemiology of cataract:
Accomplishments over 25 years and future directions. Ophthalmic
Epidemiol. 14:173–178. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Foster A and Resnikoff S: The impact of
Vision 2020 on global blindness. Eye (Lond). 19:1133–1135. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang F, Meng W and Tong B:
Down-regulation of MicroRNA-133b suppresses apoptosis of lens
epithelial cell by up-regulating BCL2L2 in age-related cataracts.
Med Sci Monit. 22:4139–4145. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong Y, Zheng Y, Xiao J, Zhu C and Zhao M:
MicroRNA let-7b induces lens epithelial cell apoptosis by targeting
leucine-rich repeat containing G protein-coupled receptor 4 (Lgr4)
in age-related cataract. Exp Eye Res. 147:98–104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng CH, Liu JH, Woung LC, Lin TJ, Chiou
SH, Tseng PC, Du WY, Cheng CK, Hu CC, Chien KH and Chen SJ:
MicroRNAs and cataracts: Correlation among let-7 expression, age
and the severity of lens opacity. Br J Ophthalmol. 96:747–751.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu C, Lin H, Wang Q, Chen W, Luo H, Chen W
and Zhang H: Discrepant expression of microRNAs in transparent and
cataractous human lenses. Invest Ophthalmol Vis Sci. 53:3906–3912.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Acunzo M and Croce CM: Downregulation of
miR-15a and miR-16-1 at 13q14 in Chronic Lymphocytic Leukemia. Clin
Chem. 62:655–656. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Renjie W and Haiqian L: miR-132, miR-15a
and miR-16 synergistically inhibit pituitary tumor cell
proliferation, invasion and migration by targeting Sox5. Cancer
Lett. 356:568–578. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang E, Liu R and Chu Y: miRNA-15a/16: As
tumor suppressors and more. Future Oncol. 11:2351–2363. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lan F, Yue X, Ren G, Li H, Ping L, Wang Y
and Xia T: miR-15a/16 enhances radiation sensitivity of non-small
cell lung cancer cells by targeting the TLR1/NF-κB signaling
pathway. Int J Radiat Oncol Biol Phys. 91:73–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ye EA, Liu L, Jiang Y, Jan J, Gaddipati S,
Suvas S and Steinle JJ: miR-15a/16 reduces retinal leukostasis
through decreased pro-inflammatory signaling. J Neuroinflammation.
13:3052016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cai CK, Zhao GY, Tian LY, Liu L, Yan K, Ma
YL, Ji ZW, Li XX, Han K, Gao J, et al: miR-15a and miR-16-1
downregulate CCND1 and induce apoptosis and cell cycle arrest in
osteosarcoma. Oncol Rep. 28:1764–1770. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hao M, Zhang L, An G, Sui W, Yu Z, Zou D,
Xu Y, Chang H and Qiu L: Suppressing miRNA-15a/-16 expression by
interleukin-6 enhances drug-resistance in myeloma cells. J Hematol
Oncol. 4:372011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vogler M, Walter HS and Dyer MJS:
Targeting anti-apoptotic BCL2 family proteins in haematological
malignancies-from pathogenesis to treatment. Br J Haematol.
178:364–379. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Frenzel A, Grespi F, Chmelewskij W and
Villunger A: Bcl2 family proteins in carcinogenesis and the
treatment of cancer. Apoptosis. 14:584–596. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang J, Wang Z, Chen L, Huang G and Hu X:
Gossypol acetate induced apoptosis of pituitary tumor cells by
targeting the BCL-2 via the upregulated microRNA miR-15a. Int J
Clin Exp Med. 8:9079–9085. 2015.PubMed/NCBI
|
|
26
|
Xia L, Zhang D, Du R, Pan Y, Zhao L, Sun
S, Hong L, Liu J and Fan D: miR-15b and miR-16 modulate multidrug
resistance by targeting BCL2 in human gastric cancer cells. Int J
Cancer. 123:372–379. 2008. View Article : Google Scholar : PubMed/NCBI
|