Introduction
Sepsis is considered to be a harmful, non-resolving
inflammatory response to infection with the presence of organ
dysfunction (1,2). In spite of the progress made in the
development of medical technology, well-equipped intensive care
units and improved practice treatments, the morbidity and mortality
rates of sepsis remain high (3).
Severe sepsis may cause multiple organ dysfunction (MODS) and the
collapse of the circulatory system (septic shock), which mainly
results from the extensive activation of inflammatory and
coagulation pathways (3,4). Poor clinical outcomes are closely
associated with the development of organ dysfunction. The greater
the severity of the organ damage, the higher the risk of mortality
(5). The dysfunction of organs
including the lung, kidney, intestine, liver and brain, in addition
to the disorder of including hematopoietic and cardiovascular
systems, have been extensively studied (6). However, little attention has been
focused on the function of the pancreas in sepsis. Previously,
research had identified that the pancreas has a vulnerability to
inflammation and injury in patients with sepsis and animal models
of sepsis induced by cecal ligation and puncture (CLP) (7).
Carbon monoxide (CO) has long been known as a toxic
gaseous molecule due to its ability to combine with hemoglobin.
However, a small quantity of endogenous CO, a byproduct of
inducible heme oxygenase-1, is continuously generated in mammals
(8). Under stress condition, the
intracellular levels of CO substantially increase, which has an
effect on cyto-protective functions (9). Previously, transition metal carbonyls
have been identified as potential CO-releasing molecules (CORMs)
with the potential to facilitate the pharmaceutical use of CO by
liberating it to the affected tissues and organs (8). Lipid-soluble metal carbonyl complex
tricarbonyldichlororuthenium (II) dimer
([Ru(CO)3Cl2]2), known as CORM-2, is the
first compound to corroborate the feasibility of this technology,
in a controlled manner without substantially altering
carboxy-hemoglobin (CO-Hb) levels (10–12).
Studies have revealed that CORM-2 is able to modulate inflammation
and inhibit the lipopolysaccharide (LPS)-induced production of
cytokines in vivo and in vitro (13–15).
Previous studies have revealed that CORM-2 attenuates leukocyte
sequestration in organs including the lung, liver and small
intestine in burned and CLP-induced mouse models of sepsis through
interfering with nuclear factor-κB (NF-κB) activation and
inhibiting the expression of adhesion molecules (16–19).
Nevertheless, to the best of our knowledge, no previous studies
have determined the regulatory effects of exogenous CO on
pancreatic function in a CLP-induced mouse model of sepsis.
The present study used a CLP-induced septic mouse
model, which was designed as a prospective experiment, to
investigate the effects of exogenous CO on the regulation of
pancreatic function and to investigate the molecular mechanisms of
underlying the therapeutic effect of CO (20,21).
The present study will provide further theoretical foundations and
strategies for the treatment of sepsis.
Materials and methods
Ethics statement
All experiments were performed in accordance with
the Guide for the Care and Use of Laboratory Animals published by
the US National Institutes of Health (NIH publication no. 85–23,
revised 1996; http://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdf).
All experimental protocols were ethically authorized by the Council
on Animal Care at Jiangsu University (Jiangsu, China) on the
Protection and the Welfare of Animals and conducted in accordance
with the National Institutes of Health of China guidelines for the
care and use of experimental animals.
Materials
CORM-2, dimethyl sulfoxide (DMSO) and
radioimmunoprecipitation assay buffer were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). CORM-2 was
dissolved in DMSO to acquire a 40 mmol/l stock solution, as
previously described (22). An
inactive form of CORM-2 (iCORM-2, used as the negative control) was
prepared as followed: The stock of CORM-2 was incubated at 37°C in
a 5% CO2 humidified atmosphere for 24 h to liberate CO.
The iCORM-2 solution was finally bubbled with nitrogen to remove
the residual CO present in the solution. The primary antibodies of
NF-κB (sc-7386), phosphorylated inhibitor of κB (p-IκB-α;
sc-52943), ICAM-1 (sc-1511) and VCAM-1 (sc1504) were purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). The nuclear
protein extraction buffer kit was obtained from Vazyme (Piscataway,
NJ, USA). Other reagents and instruments included tumor necrosis
factor-α (TNF-α; JER-06) interleukin-6 (IL-6; JEM-04) and IL-1β
(JEM-01) enzyme-linked immunosorbent assay (ELISA) kits were all
purchased from Joyee Biotechnics Co., Ltd. (Shanghai, China). All
other chemicals were of reagent grade and obtained from
Sigma-Aldrich (Merck KGaA) unless otherwise stated.
Sepsis mouse model establishment
C57BL/6 mice (n=60; male, 6–8 weeks, weight, 20±2 g)
were obtained from the Experimental Animal Center of Jiangsu
University, Zhenjiang, Jiangsu, China. Mice were housed in standard
wire-topped cages and in temperature-controlled units (18–23°C with
40–60% humidity and 12-hour light/12–hour dark cycle). Food and
water were supplied ad libitum. CLP was executed to induce
polymicrobial sepsis as previously described (3,23).
The mice were anesthetized with 2% isoflurane in oxygen through a
face-mask. Under aseptic conditions, a midline incision (1-cm) was
created through the abdominal wall. The cecum was identified then
ligated with a 3-0 silk suture at a 1 cm position from the distal
to the base of the cecum. Care was taken not to cause intestinal
obstruction. Two punctures of the cecal wall were performed using a
22-gauge needle at the top and bottom. The cecum was lightly
squeezed to extrude about 1-mm3 droplet of fecal
material from the puncture sites to ensure a full-thickness
perforation. The cecum was returned to the abdominal cavity, and
the incision was closed with 5-0 surgical sutures in layers.
Subsequent to the operation, the mice were immediately injected
with 1 ml pre-warmed sterile saline subcutaneously to replace the
fluid lost. For the sham group animals, mice underwent the same
procedure, except with no CLP treatment.
Experimental protocol
A total of 3 time points were selected following a
sham operation or CLP treatment in the experiments: 6, 12 and 24 h.
At each point in time, a total of 60 mice were randomly divided
into 4 groups: Sham group (n=15); CLP group (n=15); CLP+CORM-2
group (n=15); and CLP+iCORM-2 group (n=15). Mice in the CLP+CORM-2
and CLP+iCORM-2 groups received an injection of CORM-2 [8 mg/k
intravenously (i.v.)] or iCORM-2 (8 mg/kg, i.v.) immediately
subsequent to CLP, respectively. The dosage of CORM-2 used in the
present study was based on the results of previous studies
(16,18). The negative control (iCORM-2) was
used to examine whether the effects observed were due to CO
liberated by CORM-2 or caused by other components of the molecules.
Sham animals received normal saline intravenously following the
sham operation in the same regimen.
Tissue collection
Mice were euthanized using excessive anesthesia
administration and were sacrificed at 6, 12 and 24 h after CLP or
sham surgery as previously described (23,24).
Blood samples were collected using cardiac puncture, and the
samples were stored in serum tubes and immediately centrifuged at
3,000 × g for 5 min at room temperature. The serum was isolated
from these samples for subsequent lipase and amylase level
determination. Pancreatic tissues were removed and immediately
frozen in liquid nitrogen or fixed at room temperature overnight in
10% formalin for further studies.
Biochemical measurement
The activities of serum amylase and lipase were
measured to evaluate the pancreatic injury by using a commercial
kit (SNM144-BOU, Biolebo, Beijing, China). Levels of amylase and
lipase were determined according to the manufacturers'
protocol.
Morphological examination
Samples of 10% formalin-fixed pancreas tissue were
embedded in paraffin and sectioned at a thickness of ~4 µm for
routine histology for each group. Fixed tissues were then stained
with hematoxylin and eosin (H&E) according to the
manufacturer's protocol, and examined by two experienced
morphologists who were blinded to the sample identity. For these
studies, 10 randomly selected microscopic fields (at a
magnification of 10×20) were examined for each tissue sample. Each
section was scored for the severity of pancreatic injury on a scale
of 0–4 (normal to severe) as previously described (25).
Immunohistochemical staining. Pancreatic tissues
were fixed in 10% formalin and 4-µm thick sections were prepared
from paraffin-embedded tissues. Following deparaffinization and
rehydration, and sections were subjected to heat-mediated antigen
retrieval in sodium citrate buffer (10 mM sodium citrate, pH 6.0)
prior to blocking in 10% normal goat serum for 2 h at room
temperature. The primary antibodies of ICAM-1 and VCAM-1 were
diluted to 1/200 and the samples were incubated at 4°C overnight.
Following three washes with phosphate buffered saline, a
horseradish peroxidase (HRP)-conjugated goat anti-rabbit secondary
antibody (sc-2004, Santa Cruz Biotechnology, Inc.) was diluted
1:100 and incubated with the samples for 20 min at room
temperature. The slides were then washed again three times with
tris-buffered saline. Thereafter, the slides were filled with
freshly prepared 3,3′-Diaminobenzidine chromogen (brown) and
incubated at room temperature for 3 min, washed in distilled water
and counter-stained with hematoxylin for 10 sec at room
temperature. This was followed by dehydration with gradient ethanol
(100, 95 and 80%) and clearing using xylene. The slides were then
mounted with mounting medium, labelled and viewed under a light
microscope. For these studies, 10 randomly selected microscopic
fields (magnification, 10×20) were examined for each tissue sample.
Mean optical density for ICAM-1 and VCAM-1 were evaluated using
Image-Pro Plus version 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Myeloperoxidase (MPO) activity
MPO activity was determined in pancreatic tissues in
a procedure similar to that documented in other studies (26–28).
The tissue samples were homogenized in 50 mmol/l potassium
phosphate buffer (PB; pH 6.0) and centrifuged at 10,000 × g for 10
min at room temperature. The pellets were suspended in 50 mmol/l PB
including 0.5% hexadecyltrimethylammonium bromide. Subsequent to
sonication, the samples were centrifuged at 10,000 × g for 10 min
again at room temperature. Aliquots (0.3 ml) were added to 2.3 ml
of the reaction mixture containing o-dianisidine, 50 mmol/l PB and
20 mmol/l H2O2 solution. One unit of enzyme
activity was regarded to indicate the quantity of MPO present that
resulted in a change in the absorbance measured at 460 nm for 3
min. MPO activity was expressed as U/g tissue.
TNF-α, IL-6 and IL-1β level
detection
TNF-α, IL-6 and IL-1β levels in pancreas tissue
homogenates were measured using ELISA kits according to the
manufacturer's protocol of each kit.
Western blot analysis
Nucleic proteins were extracted using a nuclear
protein extraction buffer kit (Vazyme) according to the
manufacturer's protocol. A BCA protein assay kit (Pierce; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) was used to evaluate the
protein concentrations. Then, samples (10 µg protein) were
subjected to electrophoresis on 10% SDS-PAGE gels using a
discontinuous system and transferred onto polyvinylidene fluoride
membranes, which were then incubated with an anti-mouse
NF-κB-specific polyclonal antibody (1:1,000; cat no. Sc-514451;
Santa Cruz Biotechnology, Inc.), anti-mouse P-IκB-α-specific
polyclonal antibody (1:1,000; cat no. Sc-7977; Santa Cruz
Biotechnology, Inc.) or β-actin (1:1,000; cat no. Sc-81178; Santa
Cruz Biotechnology, Inc.) at 4°C overnight and a HRP-conjugated
goat anti-mouse immunoglobulin G antibody (1:10,000; cat no.
Sc-2031; Santa Cruz Biotechnology, Inc.; incubated at room
temperature for 1 h) was used as the secondary antibody.
Electrochemiluminescence reagent was used to visualize the bands
using FluorChem FC3 (ProteinSimple, San Jose, CA, USA), and
AlphaView version 3.4.0 software (ProteinSimple, Santa Clara, CA,
USA) was used for quantification analysis.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). Data were
presented as the mean ± the standard deviation. One-way factorial
analysis of variance followed by a Tukey's post-hoc test were
performed for comparisons between the groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of CORM-2 on the function of
the pancreas in septic mice
Sepsis results in MODS, which includes injury of the
pancreas (29,30). To demonstrate the protective
effects of CORM-2 on the function of pancreas in a CLP-induced
mouse sepsis model, the levels of serum amylase and lipase
(commonly used as bio-markers to detect pancreatic injury)
(30,31) were examined. The level of serum
amylase was significantly increased at 6, 12 and 24 h post-CLP
compared with the sham mice (P<0.05) (Fig. 1A). Administration of CORM-2 in
septic mice significantly decreased the level of serum amylase at
6, 12 and 24 h post-CLP compared with the CLP-alone group
(P<0.05; Fig. 1A). The septic
mice treated with iCORM-2 did not present altered serum amylase
activity when compared with the CLP group (Fig. 1A). Similar results were obtained
when assessing the serum lipase levels (Fig. 1B).
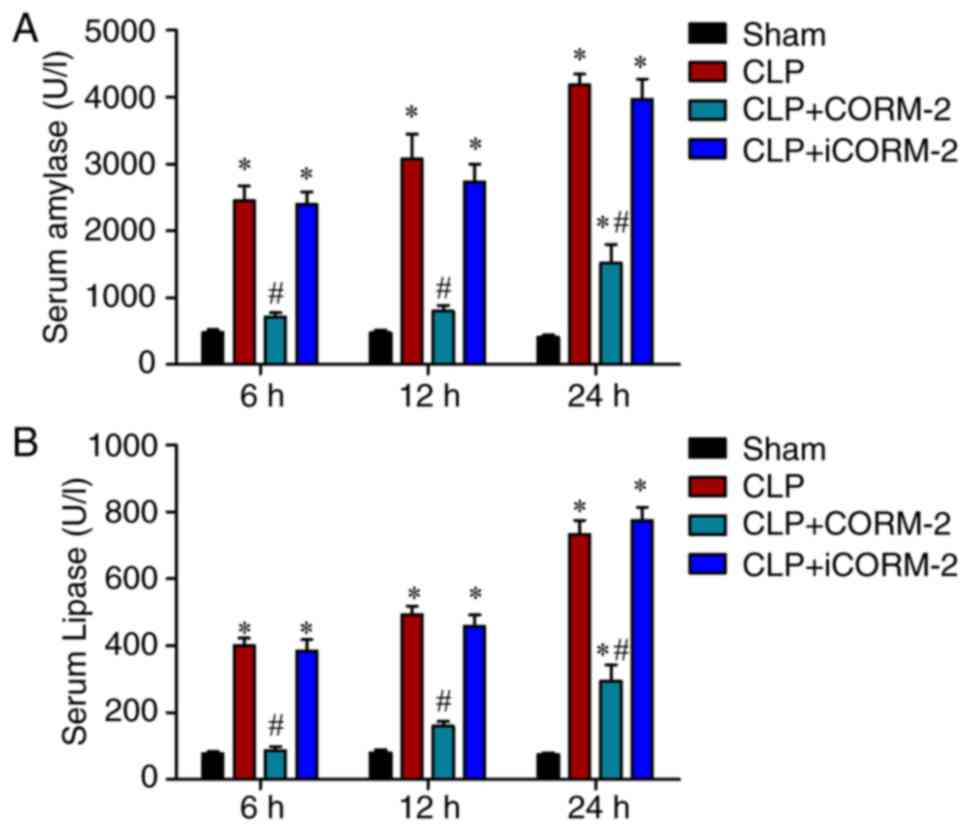 | Figure 1.Effect of CORM-2 on serum amylase and
lipase levels in CLP-induced septic mice. Serum (A) amylase and (B)
lipase levels were determined at 6, 12 and 24 h after the induction
of CLP treatment or a sham operation in four groups: A sham group;
CLP, mice subjected to CLP to induce polymicrobial sepsis;
CLP+CORM-2, mice subjected to CLP and treated with CORM-2 (8 mg/kg,
i.v.); and CLP+iCORM-2, mice subjected to CLP and treated with
iCORM-2 (8 mg/kg, i.v.). *P<0.05 vs. the sham group;
#P<0.05 vs. the CLP group. These data are expressed
as the mean ± standard deviation, n=5 for each group. CLP, cecal
ligation and puncture; CORM-2, carbon monoxide releasing
molecule-2; iCORM2, inactive CORM-2; i.v., intravenously. |
The damage to the pancreas was also examined by
histological evaluation. The pancreatic tissue from the sham group
mice exhibited normal architecture, while the pancreatic tissues in
the septic mice exhibited severe pathological injury at 24 h
post-CLP (Fig. 2A). The pancreatic
tissues in the septic mice exhibited characteristic edema,
inflammatory cell infiltration and necrosis of the acinar cells
(Fig. 2A). Treatment of the CLP
mice with CORM-2 for 24 h significantly reduced the extent and
severity of the histological signs of pancreatic injury compared
with the CLP-alone group, including edema, inflammatory cell
infiltration and necrosis (P<0.05; Fig. 2A and B), whereas alleviation in the
injury of pancreas was not observed in the iCORM-2 treatment group
compared with the CLP-alone group (Fig. 2A and B). The present study revealed
that CORM-2 treatment exerted protective effects in the CLP-induced
pancreatic damage.
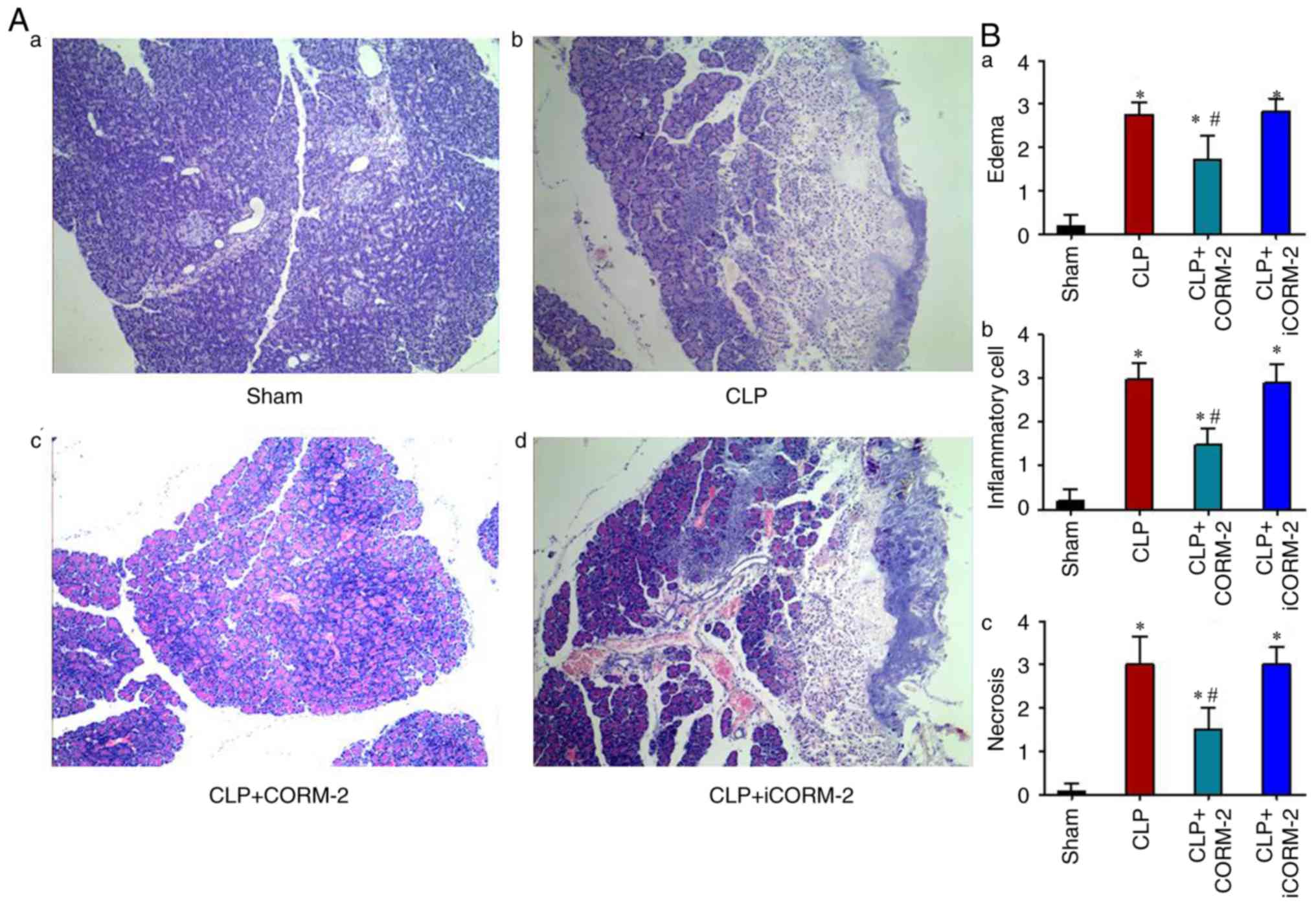 | Figure 2.Histological evaluation of the effect
of CLP and CORM-2 on the pancreas. (A) Effect of CORM-2 on
pancreatic injury at 24 h after the induction of CLP (H&E
staining, magnification ×200), in the pancreatic tissue obtained
from: (Aa) The sham group, (Ab) the CLP group (mice subjected to
CLP to induce polymicrobial sepsis), (Ac) the CLP+CORM-2 group
(mice subjected to CLP and treated with CORM-2, 8 mg/kg, i.v.) and
(Ad) the CLP+iCORM-2 group (mice subjected to CLP and treated with
iCORM-2, 8 mg/kg, i.v.). (B) Quantified effect of CORM-2 on
histopathological scores of pancreatic injury. H&E-stained
sections were evaluated for (Ba) edema, (Bb) inflammatory cell
infiltration and (Bc) acinar necrosis. These data are expressed as
the mean ± standard deviation, n=10 for each group. *P<0.05 vs.
the sham group; #P<0.05 vs. the CLP group. CLP, cecal
ligation and puncture; CORM-2, carbon monoxide releasing
molecule-2; iCORM2, inactive CORM-2; i.v., intravenously; H&E,
hematoxylin and eosin. |
Effect of CORM-2 on MPO activity in the pancreas of
septic mice. Inflammatory cell infiltration is a notable
pathogenesis of sepsis-induced acute pancreatic injury (32). To evaluate pancreatic neutrophil
infiltration, MPO activity, an index of neutrophil accumulation,
was measured in the pancreatic tissues obtained from the sham
group, the CLP group, the CLP+CORM-2 group and the CLP+iCORM-2
group post-operation at 6, 12 and 24 h. The results revealed that
MPO activity was significantly increased in septic mice at 6, 12
and 24 h post-CLP when compared with the sham group (P<0.05;
Fig. 3). Treatment with CORM-2
subsequent to CLP treatment significantly attenuated MPO activity
at 6, 12 and 24 h compared with the CLP-alone group (P<0.05;
Fig. 3), indicating that treatment
with CORM-2 decreased the infiltration of neutrophils in the
pancreatic tissues. No significant difference was identified
between the CLP group and the CLP+iCORM-2 group (Fig. 3).
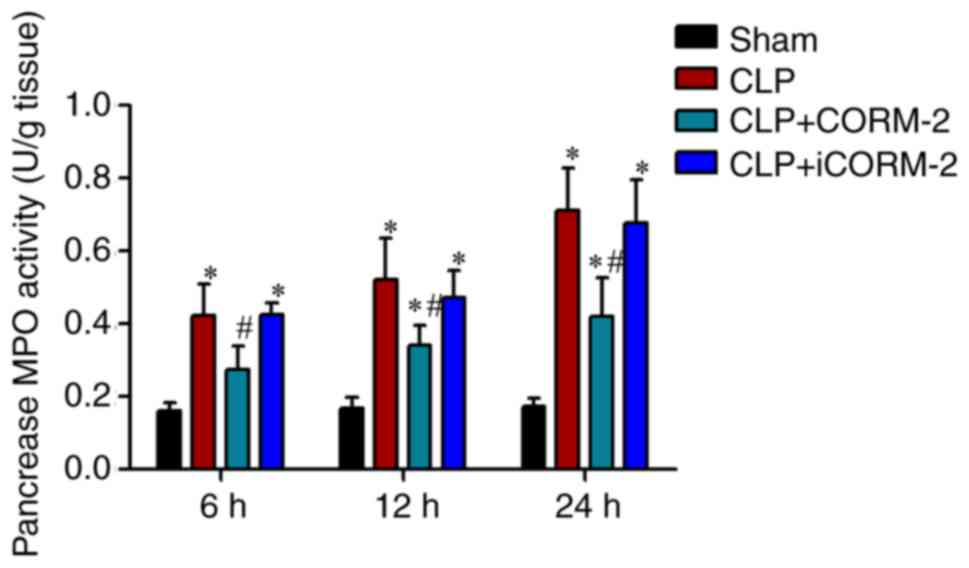 | Figure 3.Effect of CORM-2 on the MPO activity
in the pancreatic tissue of CLP-induced septic mice. MPO activity
in the pancreas was measured at 6, 12 and 24 h after the induction
of CLP in four groups: Sham, sham group; CLP, mice subjected to CLP
to induce polymicrobial sepsis; CLP+CORM-2, mice subjected to CLP
and treated with CORM-2 (8 mg/kg, i.v.); and CLP+iCORM-2, mice
subjected to CLP and treated with iCORM-2 (8 mg/kg, i.v.). These
data are expressed as the mean ± standard deviation, n=5 for each
group. *P<0.05 vs. the sham group; #P<0.05 vs. the
CLP group. CLP, cecal ligation and puncture; CORM-2, carbon
monoxide releasing molecule-2; iCORM2, inactive CORM-2; i.v.,
intravenously; MPO, myeloperoxidase. |
Effect of CORM-2 on the expression of ICAM-1 and
VCMP-1 in the pancreas of septic mice. The accumulation of
neutrophils in the tissue results from the increased adherence of
neutrophils to endothelial cells, which is mediated by adhesion
molecules (26). Infiltration of
neutrophils in the pancreas augments the injury of tissues.
Therefore, the present study evaluated the expression levels of
ICAM-1 and VCAM-1 adhesion molecules, which serve critical roles in
the firm attachment of neutrophils to the endothelium. No positive
staining for ICAM-1 was observed in the pancreatic tissue sections
obtained from the sham group mice (Fig. 4A). Sections obtained from
CLP-induced septic mice 24 h after CLP exhibited significant
positive staining for ICAM-1 and VCAM-1 compared with the sham
group (P<0.05; Fig. 4A-C). The
degree of pancreatic staining for ICAM-1 and VCAM-1 were
significantly reduced in tissue sections obtained from 24 h
post-CLP mice that had received CORM-2 treatment compared with the
CLP-alone group (P<0.05; Fig.
4A-C). No significant changes in staining were observed in
CLP-treated mice that had additionally been treated with iCORM-2
when compared with CLP-alone mice (Fig. 4A-C).
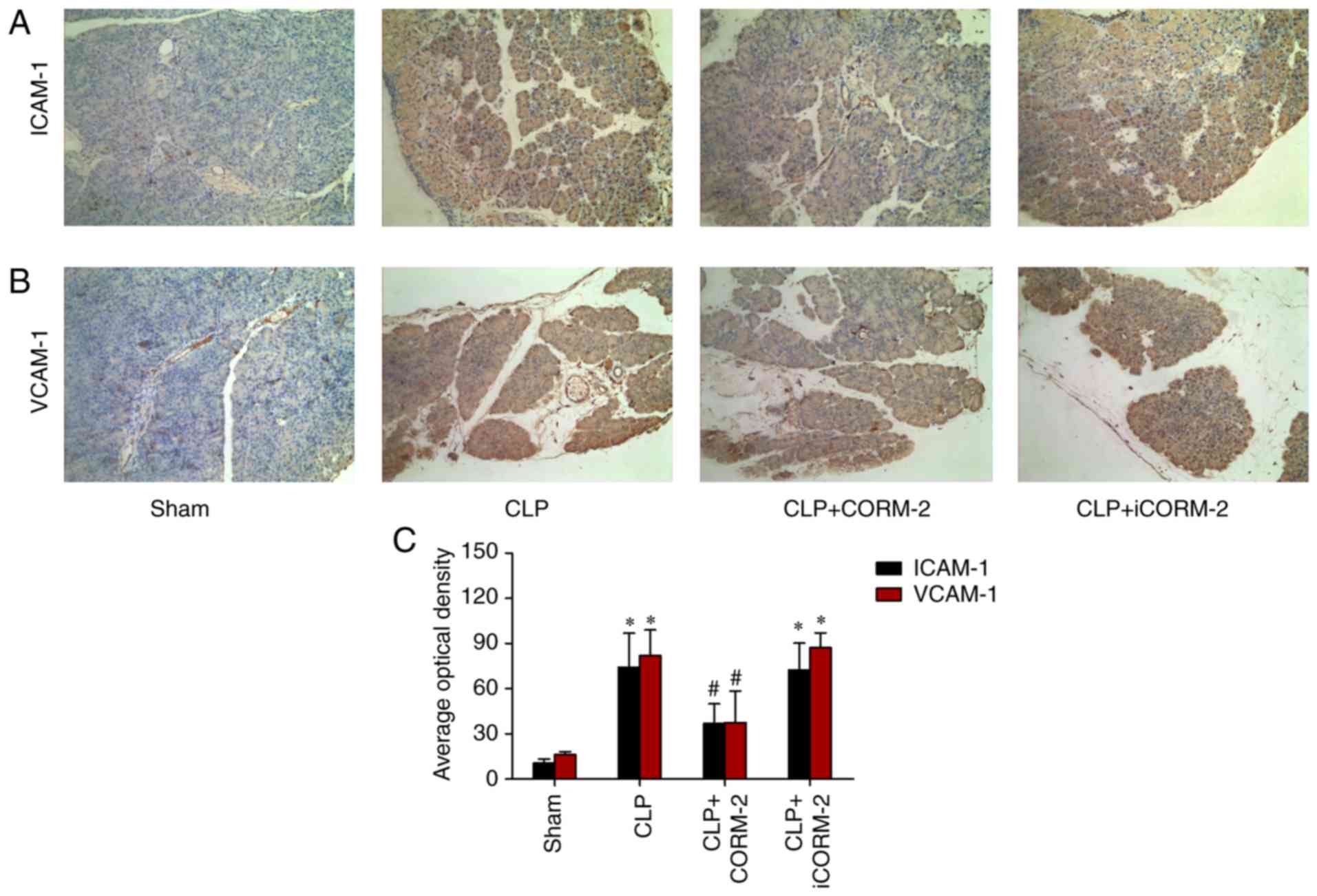 | Figure 4.Effect of CORM-2 on ICAM-1 and VCAM-1
levels in the pancreatic tissue of CLP-induced septic mice
(magnification, ×200). Expression of (A) ICAM-1 and (B) VCAM-1
levels in the pancreas were determined at 24 h post-CLP by
immunohistochemical staining, in the following four groups: Sham,
sham group; CLP, mice subjected to CLP to induce polymicrobial
sepsis; CLP+CORM-2, mice subjected to CLP and treated with CORM-2
(8 mg/kg, i.v.); and CLP+iCORM-2, mice subjected to CLP and treated
with iCORM-2 (8 mg/kg, i.v.). (C) Quantitative analysis of the mean
optical density for ICAM-1 and VCAM-1 in the pancreatic tissues.
These data are expressed as the mean ± standard deviation, n=10 for
each group. *P<0.05 vs. the sham group; #P<0.05
vs. the CLP group. CLP, cecal ligation and puncture; CORM-2, carbon
monoxide releasing molecule-2; iCORM2, inactive CORM-2; i.v.,
intravenously; ICAM-1, intercellular adhesion molecule 1; VCAM-1,
vascular cell adhesion molecule 1. |
Effect of CORM-2 on the production of
pro-inflammatory cytokines in the pancreas of septic mice
In order to investigate the regulation of
inflammation in the pancreas by CORM-2 treatment, the expression
levels of pro-inflammatory cytokines IL-6, IL-1β and TNF-α were
determined using ELISA. The levels of IL-6, IL-1β and TNF-α
significantly increased at 6, 12 and 24 h after CLP when compared
with the sham controls (P<0.05; Fig. 5A-C). Treatment with CORM-2
significantly decreased the CLP-induced increase in the expression
of IL-6, IL-1β and TNF-α compared with the CLP-alone group
(P<0.05; Fig. 5A-C).
Additionally, iCORM-2 treatment did not exert any effect in the
expression of these cytokines (Fig.
5A-C).
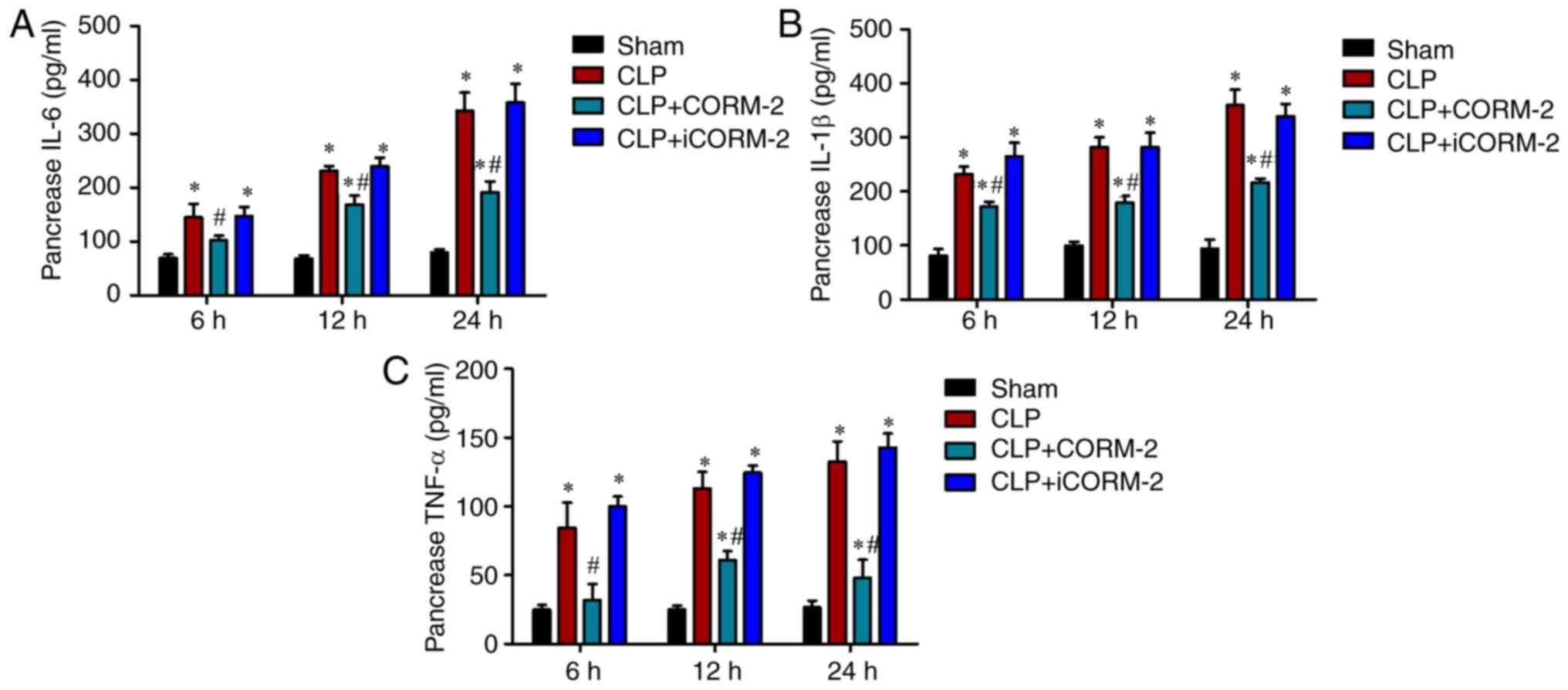 | Figure 5.Effects of CORM-2 on the levels of
pro-inflammatory cytokine in the pancreas of CLP-induced septic
mice. The expression levels of (A) IL-6, (B) IL-1β and (C) TNF-α in
the pancreas were assessed at 6, 12 and 24 h after the induction of
CLP or sham operation in the following four groups: Sham, sham
group; CLP, mice subjected to CLP to induce polymicrobial sepsis;
CLP+CORM-2, mice subjected to CLP and treated with CORM-2 (8 mg/kg,
i.v.); CLP+iCORM-2, mice subjected to CLP and treated with iCORM-2
(8 mg/kg, i.v.). These data are expressed as the mean ± standard
deviation, n=5 for each group. *P<0.05 vs. the sham group;
#P<0.05 vs. the CLP group. CLP, cecal ligation and
puncture; CORM-2, carbon monoxide releasing molecule-2; iCORM2,
inactive CORM-2; i.v., intravenously; IL-6, interleukin 6; IL-1β,
interleukin 1β; TNF-α, tumor necrosis factor α. |
Effect of CORM-2 on NF-κB activation
in the pancreas of septic mice
To determine whether NF-κB was involved in the
observed anti-inflammatory effects of CORM-2-derived CO, the levels
of NF-κB p65 in the pancreas were measured using western blot
analysis. The results demonstrated that 24 h post-CLP mice
exhibited a significantly increased protein level of NF-κB compared
with the sham group mice (P<0.05; Fig. 6A). This increase was significantly
inhibited by treatment with CORM-2 compared with the CLP-alone
group (P<0.05), whereas iCORM-2 treatment had no effect on the
levels of NF-κB (Fig. 6A).
Furthermore, p-IκB-α in the pancreas was also examined using
western blot analysis, which is required for the initiation of
NF-κB activation. There was significant increase of the levels of
p-IκB-α in the pancreas of septic mice 24 h after CLP compared with
the sham group (P<0.05), which was significantly suppressed by
the administration of CORM-2 compared with the CLP-alone group
(P<0.05; Fig. 6B). No
significant change in the p-IκB-α levels in the pancreas was
observed by treatment with iCORM-2 when compared with the CLP-alone
mice (Fig. 6B).
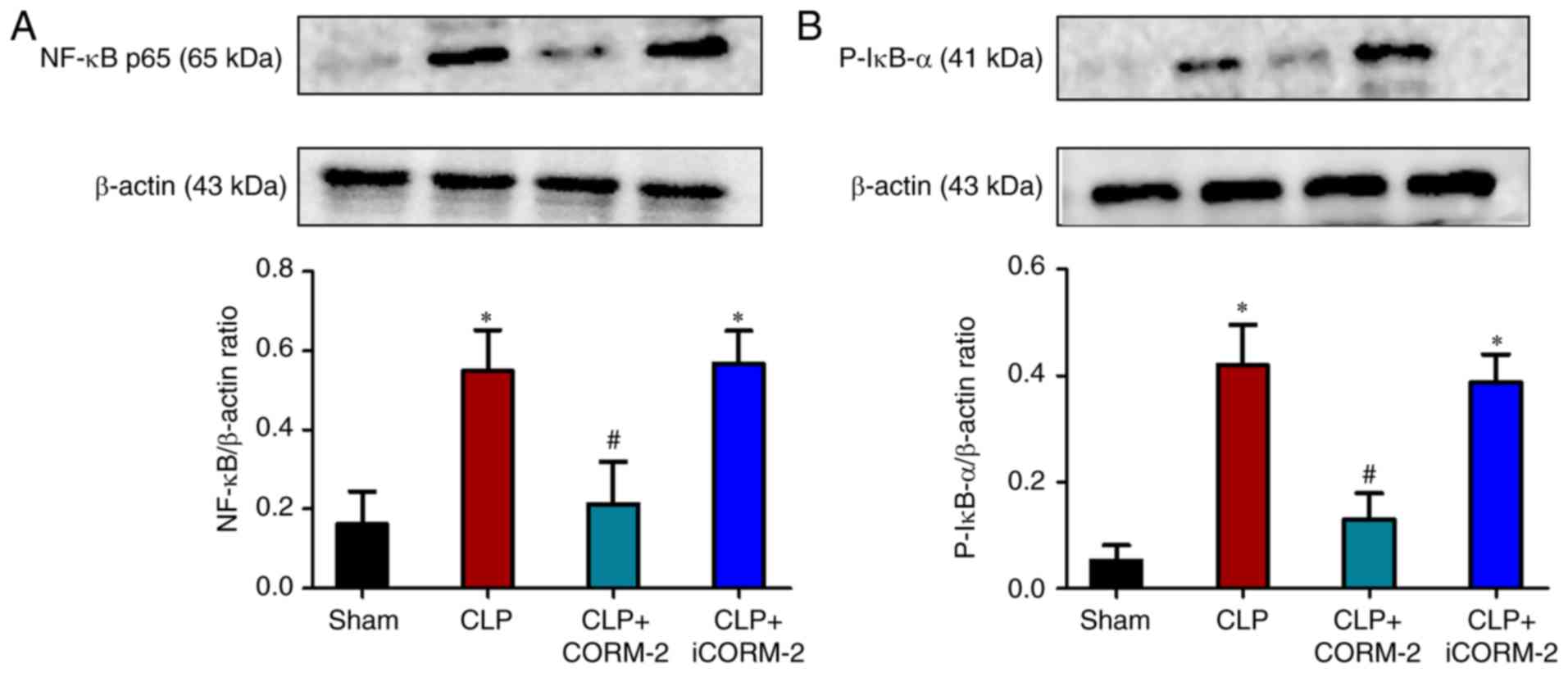 | Figure 6.Effect of CORM-2 on the levels of
NF-κB p65 and p-IκB-α in the pancreas of CLP-induced septic mice.
The levels of (A) NF-κB and (B) p-IκB-α were analyzed using western
blot analysis at 24 h after the induction of CLP or sham operation
in the following four groups: Sham, sham group; CLP, mice subjected
to CLP to induce polymicrobial sepsis; CLP+CORM-2, mice subjected
to CLP and treated with CORM-2 (8 mg/kg, i.v.); and CLP+iCORM-2,
mice subjected to CLP and treated with iCORM-2 (8 mg/kg, i.v.).
These data are expressed as the mean ± standard deviation, n=5 for
each group. *P<0.05 vs. the sham group; #P<0.05
vs. the CLP group. CLP, cecal ligation and puncture; CORM-2, carbon
monoxide releasing molecule-2; iCORM2, inactive CORM-2; i.v.,
intravenously; NF-κβ, nuclear factor-κβ; p-IκB-α, phosphorylated
inhibitor of κB. |
Discussion
Sepsis is a common phenomenon amongst critically ill
patients, and is associated with the progressive failure of
multiple organs (1). Although the
most common dysfunctions of organs in sepsis are in the lung and
kidney, the pancreas is also susceptible to inflammation and injury
in patients with sepsis (3,6).
Research suggests that sepsis has a notable impact on pancreatic
secretory function, which augments the severity of sepsis and is
worse in septic shock compared with sepsis without shock (6). Impaired exocrine function in the
pancreas is associated with Acute Physiology and Chronic Health
Evaluation III and Sequential Organ Failure Assessment scores
(6).
Previous studies have demonstrated the specific and
independent function of exogenous CO in the modulation of
inflammation (14,19,33–36).
As novel metal carbonyl-based compounds, CORMs have the ability to
deliver CO in biological systems in a controlled manner (11). CORM-derived CO has been
demonstrated to have various benefits, including vasoactive,
antihypertensive, cardio-protective and anti-rejection effects
(37–40). CORM-2, one of the novel group of
CORMs, has been determined to possess anti-inflammatory effects and
to ameliorate the function of the pancreas in acute pancreatitis
(20,21). In previous studies, the
administration of CORM-2 improved the function of numerous organs
in CLP and thermal injury animal models of sepsis (11,18).
However, it remains unclear whether CORM-2 serves a protective
effect on the function of the pancreas.
To determine the therapeutic function of CO, the
present study initially established CLP septic mice and sham mice.
The animals were separated into four groups: Sham group; CLP group;
CLP+CORM-2 group; and CLP+iCORM-2 group. Each group was divided
again at postoperative 6, 12 and 24 h time points. CORM-2 or
iCORM-2 were injected immediately following CLP. The levels of
serum amylase and lipase (which are used to detect pancreatic
injury) were examined in the present study. The results indicated
that treatment with CORM-2 in CLP mice suppressed the levels of
serum amylase and lipase compared with the sham group mice at all
three time points. The septic mice treated with iCORM-2 did not
exhibit altered serum amylase and lipase activity when compared
with the CLP group mice at all three postoperative time points.
Furthermore, the damage to the pancreas was also examined by
histological evaluation. At all three time points, CORM-2 exerted a
protective effect on the function of pancreas, and 24 h post-CLP
exhibited the most substantial effect. Therefore, the present study
selected the representative H&E stained pancreatic sections in
the four groups at the 24 h time point (Fig. 1). The pancreas from the mice of the
sham group exhibited normal architecture, while the pancreatic
tissues in the septic mice exhibited characteristic edema,
inflammatory cell infiltration and necrosis of the acinar cells.
Treatment with CORM-2 substantially reduced the extent and severity
of the histological signs of pancreatic injury compared with the
CLP-alone group, whereas alleviation in the injury of the pancreas
was not observed in the iCORM-2 treatment group. The results were
consistent with the experimental data of serum amylase and lipase
levels. As the specific mechanisms for the therapeutic effects of
CORM-2 in CLP are unknown, the aim of the present study was to
investigate the potential mechanisms and identify CORM-2 mediated
cellular targets.
MPO is an enzyme that is present predominantly in
the azurophilic granules of polymorphonuclears (PMNs), and is
frequently used to estimate tissue PMN accumulation (26,27).
Neutrophil infiltration that aggravates tissue inflammation and
damage is considered a principal contributor to sepsis mortality
(41,42). In the present study, it was
revealed that MPO activity in the pancreas was substantially
elevated at 6, 12 and 24 h after CLP compared with the sham group.
These effects were notably attenuated following the administration
of CORM-2 in vivo, but not iCORM-2, suggesting that CORM-2
effectively inhibited neutrophil chemotaxis and infiltration in the
pancreas subsequent to CLP, eventually decreased the production of
oxidants and reduced oxidant-mediated injury and the inflammatory
response in the pancreas.
The direct cause of neutrophil infiltration into
tissues following CLP is considered to be the overexpression of
adhesion molecules (for example, ICAM-1 and VCAM-1) (36). Adhesion molecules activate
neutrophils and endothelial cells, which in turn accelerate the
release of various inflammatory mediators (43,44).
In the present study, immunohistochemical staining was used to
detect the expression of ICAM-1 and VCAM-1 in pancreas tissue. The
present results demonstrated that the expression levels of ICAM-1
and VCAM-1 in the pancreas were increased at 24 h post-CLP compared
with the sham group. Administration of CORM-2 inhibited the
upregulation of ICAM-1 and VCAM-induced by CLP. No significant
change in the expression levels were observed in iCORM-2 treated
CLP mice when compared with the CLP-alone treated mice.
Research has revealed that sepsis induces a
widespread inflammatory response with a substantial increase of
pro-inflammatory cytokines in the serum, which are important
mediators in the pathogenesis of acute pancreatic injury (45). TNF-α, IL-1β and IL-6 are induced in
exaggerated production in the early stages of infectious diseases
(46,47). Studies have demonstrated that
CORM-2 treatment abolishes the elevation in the levels of TNF-α,
IL-1β and IL-6 in the serum in CLP-induced peritonitis sepsis and
LPS-induced sterile sepsis (3,20).
Therefore, the levels of TNF-α, IL-1β and IL-6 in the pancreas were
assessed in the present study. Similarly, the levels of TNF-α,
IL-1β and IL-6 increased at 6, 12 and 24 h post-CLP compared with
the sham group, which were attenuated by CORM-2 treatment. These
data indicated that the cytoprotective effects of CORM-2 were, at
least partly, due to the inhibition of the production and secretion
of TNF-α, IL-1β and IL-6 during the early period of sepsis.
The NF-κB family members are ubiquitous, which
rapidly trigger transcription factors that mediate immune and
inflammatory reactions by regulating the expression of certain
chemokines and cytokines (48,49).
Previously, it was reported that CORM-2 inhibited the activity of
NF-κB in the tissues of the liver and lung in thermal injury mice
models and E coli-induced murine sepsis models (11,26).
Therefore, the present study evaluated the involvement of NF-κB in
the observed anti-inflammatory effects of CORM-2-derived CO on the
pancreatic damage induced by CLP. For this purpose, the nuclear
translocation and DNA binding of p65, the key component of NF-κB
(50–52), was measured in the pancreas using
western blot analysis. Additionally, p-IκB-α in the pancreas was
also examined using western blot analysis, which is required for
the initiation of NF-κB activation. The results indicated that
NF-κB p65 and p-IκB-α increased 24 h after CLP treatment compared
with the sham group, and this increase was suppressed by treatment
with CORM-2. The levels of NF-κB p65 and p-IκB-α at 24 h post-CLP
were evaluated as CORM-2 exerted a notable protective effect on the
pancreatic function of septic mice 24 h post-CLP (as presented in
Fig. 1). In fact, at
post-operative 6 and 12 h time points, similar results of NF-κB p65
and p-IκB-α were observed (data not shown). Inhibition of CORM-2 on
the activation of NF-κB p65 and p-IκB-α may explain the decrease in
the expression of inflammatory factors including TNF-α, IL-1β and
IL-6 observed in the pancreas. However, despite p-IκB-α being
required for the initiation of NF-κB activation, and numerous other
studies have only assessed the expression of p-IκB-α to demonstrate
the activation of NF-κB (53–55),
in order for the experimental results to be more convincing, total
IκB-α also needs to be tested. This is a limitation of the present
study.
Despite the therapeutic and anti-inflammatory
effects of exogenous CO in various disease and injury models, there
has been little progress with regard to its use in clinical
illness. The main problem is the method of administration of CO.
Although low dose inhalational CO has been revealed to have
anti-inflammatory effects against ventilatory-induced lung injury
and has been associated with a decreased level of TNF-α (21,56),
it is difficult to ensure a certain dose range of therapeutic CO
without increasing the level of CO-hemoglobin (57). CORM-2 or iCORM-2, at the
concentrations used in the present study, have previously been
demonstrated not to be toxic to mammalian cells in vitro and
to mice in vivo (21,58).
However, the safety of intravenous injection with CORM-2 in
vivo has not been demonstrated satisfactorily. Therefore, a
novel method of administration of CO requires further
investigation.
In summary, the present study demonstrated that the
application of CORM-2 attenuated the severity of sepsis and
improved the function of the pancreas. The mechanism by which
CORM-2-derived CO inhibited pro-inflammatory cytokines of the
pancreas may be via inhibiting NF-κB activation. However, this was
a preliminary study on the function of CORM-2 in pancreatic
function in sepsis model mice and its mechanism of action. There
remain numerous issues that require further investigation, for
example, whether the effect of CORM-2 on NF-κB activation is direct
or indirect; what will happen if NF-κB and/or p-IκB-α are blocked;
and if NF-κB and/or P-IκB-α are blocked, will any reverse effect
occur or not. Future studies will conduct in-depth research on
these issues.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81272148 and
81471903) and the Jiangsu Natural Science Foundation (grant no.
BK2012703).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
BS designed the study. YL was responsible for data
access and analysis and manuscript preparation. XW, XX and WQ
collaborated to perform data analysis and interpret the
results.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Affiliated Hospital of Jiangsu University (Jiangsu,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Abou Dagher G, Harmouche E, Jabbour E,
Bachir R, Zebian D and Bou Chebl R: Sepsis in hemodialysis
patients. BMC Emerg Med. 15:302015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Aikawa N: Revised Surviving Sepsis
Campaign Guidelines and therapy for severe sepsis. Jpn J Antibiot.
64:37–44. 2011.(In Japanese). PubMed/NCBI
|
|
3
|
Liu Z, Shi Q, Liu J, Abdel-Razek O, Xu Y,
Cooney RN and Wang G: Innate immune molecule surfactant protein D
attenuates sepsis-induced acute pancreatic injury through
modulating apoptosis and NF-κB-mediated inflammation. Sci Rep.
5:177982015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levy MM, Fink MP, Marshall JC, Abraham E,
Angus D, Cook D, Cohen J, Opal SM, Vincent JL and Ramsay G;
SCCM/ESICM/ACCP/ATS/SIS, . 2001 SCCM/ESICM/ACCP/ATS/SIS
International Sepsis Definitions Conference. Crit Care Med.
31:1250–1256. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marshall JC, Cook DJ, Christou NV, Bernard
GR, Sprung CL and Sibbald WJ: Multiple organ dysfunction score: A
reliable descriptor of a complex clinical outcome. Crit Care Med.
23:1638–1652. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tribl B, Sibbald WJ, Vogelsang H,
Spitzauer S, Gangl A and Madl C: Exocrine pancreatic dysfunction in
sepsis. Eur J Clin Invest. 33:239–243. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tribl B, Madl C, Mazal PR, Schneider B,
Spitzauer S, Vogelsang H and Gangl A: Exocrine pancreatic function
in critically ill patients: Septic shock versus non-septic
patients. Crit Care Med. 28:1393–1398. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun BW and Chen X: Carbon monoxide
releasing molecules: New insights for anticoagulation strategy in
sepsis. Cell Mol Life Sci. 66:365–369. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Soni H, Pandya G, Patel P, Acharya A, Jain
M and Mehta AA: Beneficial effects of carbon monoxide-releasing
molecule-2 (CORM-2) on acute doxorubicin cardiotoxicity in mice:
Role of oxidative stress and apoptosis. Toxicol Appl Pharmacol.
253:70–80. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chung SW, Liu X, Macias AA, Baron RM and
Perrella MA: Heme oxygenase-1-derived carbon monoxide enhances the
host defense response to microbial sepsis in mice. J Clin Invest.
118:239–247. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun B, Sun Z, Jin Q and Chen X:
CO-releasing molecules (CORM-2)-liberated CO attenuates leukocytes
infiltration in the renal tissue of thermally injured mice. Int J
Biol Sci. 4:176–183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee S, Lee SJ, Coronata AA, Fredenburgh
LE, Chung SW, Perrella MA, Nakahira K, Ryter SW and Choi AM: Carbon
monoxide confers protection in sepsis by enhancing beclin
1-dependent autophagy and phagocytosis. Antioxid Redox Signal.
20:432–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Coburn RF: The measurement of endogenous
carbon monoxide production. J Appl Physiol (1985). 112:1949–1955.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ryter SW and Choi AM: Carbon monoxide:
Present and future indications for a medical gas. Korean J Intern
Med. 28:123–140. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ozaki KS, Yoshida J, Ueki S, Pettigrew GL,
Ghonem N, Sico RM, Lee LY, Shapiro R, Lakkis FG, Pacheco-Silva A
and Murase N: Carbon monoxide inhibits apoptosis during cold
storage and protects kidney grafts donated after cardiac death.
Transpl Int. 25:107–117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu DM, Sun BW, Sun ZW, Jin Q, Sun Y and
Chen X: Suppression of inflammatory cytokine production and
oxidative stress by CO-releasing molecules-liberated CO in the
small intestine of thermally-injured mice. Acta Pharmacol Sin.
29:838–846. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Katada K, Bihari A, Mizuguchi S, Yoshida
N, Yoshikawa T, Fraser DD, Potter RF and Cepinskas G: Carbon
monoxide liberated from CO-releasing molecule (CORM-2) attenuates
ischemia/reperfusion (I/R)-induced inflammation in the small
intestine. Inflammation. 33:92–100. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun BW, Jin Q, Sun Y, Sun ZW, Chen X, Chen
ZY and Cepinskas G: Carbon liberated from CO-releasing molecules
attenuates leukocyte infiltration in the small intestine of
thermally injured mice. World J Gastroenterol. 13:6183–6190. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun B, Sun H, Liu C, Shen J, Chen Z and
Chen X: Role of CO-releasing molecules liberated CO in attenuating
leukocytes sequestration and inflammatory responses in the lung of
thermally injured mice. J Surg Res. 139:128–135. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen P, Sun B, Chen H, Wang G, Pan S, Kong
R, Bai X and Wang S: Effects of carbon monoxide releasing
molecule-liberated CO on severe acute pancreatitis in rats.
Cytokine. 49:15–23. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xue J and Habtezion A: Carbon
monoxide-based therapy ameliorates acute pancreatitis via TLR4
inhibition. J Clin Invest. 124:437–447. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu D, Liang F, Wang X, Cao J, Qin W and
Sun B: Suppressive effect of CORM-2 on LPS-induced platelet
activation by glycoprotein mediated HS1 phosphorylation
interference. PLoS One. 8:e831122013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu J, Abdel-Razek O, Liu Z, Hu F, Zhou Q,
Cooney RN and Wang G: Role of surfactant proteins A and D in
sepsis-induced acute kidney injury. Shock. 43:31–38. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang Z, Ni L, Javidiparsijani S, Hu F,
Gatto LA, Cooney R and Wang G: Enhanced liver autophagic activity
improves survival of septic mice lacking surfactant proteins A and
D. Tohoku J Exp Med. 231:127–138. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deng W, Hui Y, Yu J, Wang W, Xu S, Chen C
and Xiong X: A new pathological scoring method for adrenal injury
in rats with severe acute pancreatitis. Pathol Res Pract.
210:1011–1017. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen WC, Wang X, Qin WT, Qiu XF and Sun
BW: Exogenous carbon monoxide suppresses Escherichia coli vitality
and improves survival in an Escherichia coli-induced murine sepsis
model. Acta Pharmacol Sin. 35:1566–1576. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hillegass LM, Griswold DE, Brickson B and
Albrightson-Winslow C: Assessment of myeloperoxidase activity in
whole rat kidney. J Pharmacol Methods. 24:285–295. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou X, Liu Z, Jang F, Xiang C, Li Y and
He Y: Autocrine sonic hedgehog attenuates inflammation in
cerulein-induced acute pancreatitis in mice via upregulation of
IL-10. PLoS One. 7:e441212012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tribl B, Bateman RM, Milkovich S, Sibbald
WJ and Ellis CG: Effect of nitric oxide on capillary hemodynamics
and cell injury in the pancreas during Pseudomonas
pneumonia-induced sepsis. Am J Physiol Heart Circ Physiol.
286:H340–H345. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pieracci FM and Barie PS: Management of
severe sepsis of abdominal origin. Scand J Surg. 96:184–196. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kong XY, Du YQ, Li L, Liu JQ, Wang GK, Zhu
JQ, Man XH, Gong YF, Xiao LN, Zheng YZ, et al: Plasma miR-216a as a
potential marker of pancreatic injury in a rat model of acute
pancreatitis. World J Gastroenterol. 16:4599–4604. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Choudhury S, Kandasamy K, Maruti BS,
Addison MP, Kasa JK, Darzi SA, Singh TU, Parida S, Dash JR, Singh V
and Mishra SK: Atorvastatin along with imipenem attenuates acute
lung injury in sepsis through decrease in inflammatory mediators
and bacterial load. Eur J Pharmacol. 765:447–456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Queiroga CS, Vercelli A and Vieira HL:
Carbon monoxide and the CNS: Challenges and achievements. Br J
Pharmacol. 172:1533–1545. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Song M, Wang X, Qin W, Zhuang M, Xu X,
Zhang Y and Sun B: Effects of exogenous carbon monoxide-releasing
molecule 2 intervention in vitro on formation of human neutrophil
extracellular traps stimulated by endotoxin/lipopolysaccharide and
its mechanism. Zhonghua Shao Shang Za Zhi. 32:82–88. 2016.(In
Chinese). PubMed/NCBI
|
|
35
|
Liu D, Wang X, Qin W, Chen J, Wang Y,
Zhuang M and Sun B: Suppressive effect of exogenous carbon monoxide
on endotoxin-stimulated platelet over-activation via the
glycoprotein-mediated PI3K-Akt-GSK3β pathway. Sci Rep. 6:236532016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qin W, Zhang J, Lv W, Wang X and Sun B:
Effect of carbon monoxide-releasing molecules II-liberated CO on
suppressing inflammatory response in sepsis by interfering with
nuclear factor kappa B activation. PLoS One. 8:e758402013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Motterlini R, Clark JE, Foresti R,
Sarathchandra P, Mann BE and Green CJ: Carbon monoxide-releasing
molecules: Characterization of biochemical and vascular activities.
Circ Res. 90:E17–E24. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Motterlini R, Sawle P, Hammad J, Bains S,
Alberto R, Foresti R and Green CJ: CORM-A1: A new pharmacologically
active carbon monoxide-releasing molecule. FASEB J. 19:284–286.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Johnson TR, Mann BE, Clark JE, Foresti R,
Green CJ and Motterlini R: Metal carbonyls: A new class of
pharmaceuticals? Angew Chem Int Ed Engl. 42:3722–3729. 2005.
View Article : Google Scholar
|
|
40
|
Clark JE, Naughton P, Shurey S, Green CJ,
Johnson TR, Mann BE, Foresti R and Motterlini R: Cardioprotective
actions by a water-soluble carbon monoxide-releasing molecule. Circ
Res. 93:e2–e8. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cioffi WG, Burleson DG and Pruitt BA Jr:
Leukocyte responses to injury. Arch Surg. 128:1260–1267. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bradley PP, Priebat DA, Christensen RD and
Rothstein G: Measurement of cutaneous inflammation: Estimation of
neutrophil content with an enzyme marker. J Invest Dermatol.
78:206–209. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Christofidou-Solomidou M, Nakada MT,
Williams J, Muller WA and DeLisser HM: Neutrophil platelet
endothelial cell adhesion molecule-1 participates in neutrophil
recruitment at inflammatory sites and is down-regulated after
leukocyte extravasation. J Immunol. 158:4872–4878. 1997.PubMed/NCBI
|
|
44
|
Imamoto E, Yoshida N, Uchiyama K, Kuroda
M, Kokura S, Ichikawa H, Naito Y, Tanigawa T and Yoshikawa T:
Inhibitory effect of pioglitazone on expression of adhesion
molecules on neutrophils and endothelial cells. Biofactors.
20:37–47. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pereda J, Sabater L, Aparisi L, Escobar J,
Sandoval J, Viña J, López-Rodas G and Sastre J: Interaction between
cytokines and oxidative stress in acute pancreatitis. Curr Med
Chem. 13:2775–2787. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim GY, Roh SI, Park SK, Ahn SC, Oh YH,
Lee JD and Park YM: Alleviation of experimental septic shock in
mice by acidic polysaccharide isolated from the medicinal mushroom
Phellinus linteus. Biol Pharm Bull. 26:1418–1423. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiang K, Cheng L, Luo Z, Ren J, Tian F,
Tang L, Chen T and Dai R: Glycyrrhizin suppresses the expressions
of HMGB1 and relieves the severity of traumatic pancreatitis in
rats. PLoS One. 9:e1159822014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Baeuerle PA and Henkel T: Function and
activation of NF-kappa B in the immune system. Annu Rev Immunol.
12:141–179. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yin MJ, Yamamoto Y and Gaynor RB: The
anti-inflammatory agents aspirin and salicylate inhibit the
activity of I(kappa)B kinase-beta. Nature. 396:77–80. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Han B, Ji B and Logsdon CD: CCK
independently activates intracellular trypsinogen and NF-kappaB in
rat pancreatic acinar cells. Am J Physiol Cell Physiol.
280:C465–C472. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kulms D and Schwarz T: NF-kappaB and
cytokines. Vitam Horm. 74:283–300. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Song YS, Lee YS, Narasimhan P and Chan PH:
Reduced oxidative stress promotes NF-kappaB-mediated
neuroprotective gene expression after transient focal cerebral
ischemia: Lymphocytotrophic cytokines and antiapoptotic factors. J
Cereb Blood Flow Metab. 27:764–775. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dai B, Lei C, Lin R, Tao L, Bin Y, Peng H
and Lei B: Activation of liver X receptor α protects amyloid
β1-40 induced inflammatory and senescent responses in
human retinal pigment epithelial cells. Inflamm Res. 66:523–534.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tao L, Qiu Y, Fu X, Lin R, Lei C, Wang J
and Lei B: Angiotensin-converting enzyme 2 activator diminazene
aceturate prevents lipopolysaccharide-induced inflammation by
inhibiting MAPK and NF-κB pathways in human retinal pigment
epithelium. J Neuroinflammation. 13:352016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lu Y, Li F, Xu T and Sun J: Tetrandrine
prevents multidrug resistance in the osteosarcoma cell line, U-2OS,
by preventing Pgp overexpression through the inhibition of NF-κB
signaling. Int J Mol Med. 39:993–1000. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dolinay T, Szilasi M, Liu M and Choi AM:
Inhaled carbon monoxide confers antiinflammatory effects against
ventilator-induced lung injury. Am J Respir Crit Care Med.
170:613–620. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
El-Mousleh T, Casalis PA, Wollenberg I,
Zenclussen ML, Volk HD, Langwisch S, Jensen F and Zenclussen AC:
Exploring the potential of low doses carbon monoxide as therapy in
pregnancy complications. Med Gas Res. 2:42012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Desmard M, Foresti R, Morin D, Dagouassat
M, Berdeaux A, Denamur E, Crook SH, Mann BE, Scapens D, Montravers
P, et al: Differential antibacterial activity against Pseudomonas
aeruginosa by carbon monoxide-releasing molecules. Antioxid Redox
Signal. 16:153–163. 2012. View Article : Google Scholar : PubMed/NCBI
|














