Introduction
Gastric cancer (GC) is one of the most common types
of cancer worldwide and is one of the leading causes of
cancer-associated mortality, particularly in China (1). Recent advances in diagnosis and
treatment have improved the long-term survival rate of patients
with early-stage GC; however, the prognosis of patients with
late-stage GC remains poor, primarily due to cancer invasion and
metastasis (2–5). Tumour invasion and metastasis are
progressive, multifactorial and multistep processes that involve
cell adhesion and migration, local invasion into the adjacent
tissue, intravasation, survival in the circulatory system,
extravasation and migration into target organs, where tumour cells
may further proliferate (6). The
invasion of tumour cells into surrounding tissues is an important
early event in GC metastasis (7,8).
However, the molecular mechanism underlying GC invasion of
surrounding tissues remains unclear. Therefore, investigation of
this process is required to improve the understanding of tumour
metastasis. The present results may facilitate the identification
of novel molecular targets that may be used to develop novel
therapeutic strategies to inhibit GC metastasis.
Long noncoding RNAs (lncRNAs) are RNA molecules
>200 nucleotides in length. These RNAs have limited coding
capability; however, they are involved in various biological
processes, including epigenetic modifications (9,10),
and transcriptional (11) and
post-transcriptional regulation (12). Additionally, putative lncRNAs have
been identified to encode small peptides (13). Therefore, lncRNAs may serve various
roles in multiple diseases. Accumulating evidence has demonstrated
that lncRNAs are dysregulated in various types of cancer, serving
important roles in signalling pathways controlling tumourigenesis
and cancer progression (14).
Multiple lncRNAs, including HOX transcript antisense RNA (HOTAIR)
(15,16), gastric carcinoma proliferation
enhancing transcript 1 (GHET1) (17), hepatocellular carcinoma upregulated
long noncoding RNA (HULC) (18),
colon cancer associated transcript 1 (CCAT1) (19) and maternally expressed 3 (MEG3)
(20,21), were demonstrated to be associated
with GC progression, serving as oncogenes or tumour suppressor
genes.
A previous study by Iyer et al (22) identified various novel lncRNAs;
among these novel lncRNAs, stomach cancer-associated transcript 16
(STCAT16) exhibited a significant downregulation in GC tissues;
however, the association between STCAT16 and clinicopathological
characteristics of patients with GC remains unknown. In the present
study, the expression level of STCAT16 was identified to be
downregulated in GC tissues and cell lines. STCAT16 downregulation
was significantly associated with poor clinical features and
prognosis. Notably, STCAT16 overexpression inhibited proliferative
and invasive abilities of GC cells, and multivariate analysis
identified that high expression level of STCAT16 was an independent
predictor for improved overall survival (OS) rate in patients with
GC.
Materials and methods
Human samples
The present study was approved by The Institutional
Review Board of The Affiliated Hospital of Nantong University
(Nantong, China) and was performed in accordance with the
Declaration of Helsinki. Written informed consent was obtained
prior to the start of the study. GC tissues and matched normal
gastric tissues, located 5 cm from the tumour margin, were
collected from 59 patients with GC (age, 32–84 years; female to
male ratio, 24:35) from the Department of General Surgery
(Affiliated Hospital of Nantong University) between December 2014
and August 2015. Clinical data and follow-up information were
obtained from the medical records of the patients. GC stage was
assigned according to the tumour, node and metastasis (TNM)
classification and staging system recommended by the American Joint
Committee on Cancer (23). T and N
classification was based on non-invasive examinations and M
classification was determined from samples that were surgically
removed. OS was defined as the duration between the date of surgery
and the date of mortality or the last follow-up. All patients were
followed up until December 2017. The patients did not receive
radiation therapy or chemotherapy prior to surgery. Patients
included in the study did not exhibit long-term use of nonsteroidal
anti-inflammatory drugs or corticosteroids prior to the surgery.
Cancer classification and pathological types were assessed by two
experienced pathologists in a double-blind manner.
Cell culture and transfection
Human GC cell lines (SGC-7901, MKN-45, AGS, MGC-803
and BGC-823) and a human normal gastric mucosa cell line (GES-1)
were purchased from the Shanghai Institutes for Biological Sciences
of The Chinese Academy of Sciences (Shanghai, China). Cells were
cultured in RPMI-1640 medium (HyClone; GE Healthcare Life Sciences,
Logan, UT, USA) containing 10% foetal bovine serum (FBS; Zhejiang
Tianhang Biotechnology Co., Ltd., Huzhou, China), 100 µg/ml
streptomycin and 100 U/ml penicillin at 37°C with 5%
CO2. Cells in the logarithmic (log) phase were selected
and transfected in 24-well plates at 85% confluence using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The cells were dissociated
using a solution containing 0.25% trypsin-EDTA (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). Subsequent experiments were
performed 72 h post-transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The relative expression level of STCAT16 was
determined by RT-qPCR. Total RNA was extracted from 100 mg tissues
or cells using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.). Subsequently, RNA was reverse-transcribed into
cDNA with a Moloney murine leukaemia virus reverse transcriptase
kit (Thermo Fisher Scientific, Inc.). The temperature protocol was
as follows: 42°C for 60 min, 70°C for 5 min. According to the
manufacturer's instructions, qPCR was executed using SYBR-Green I
(Takara Biotechnology, Co., Ltd., Dalian, China) and an Applied
Biosystems 7500 Real-Time PCR System (Thermo Fisher Scientific,
Inc.). The qPCR reaction mixture contained 10 µl SYBR-Green/ROX
qPCR Master Mix (Takara Biotechnology, Co., Ltd.), 1 µl forward
primers (STCAT16, 5′-CATCAAGGCTTGTGGGATGT-3′; GAPDH,
5′-CTGGGCTACACTGAGCACC-3′), 1 µl reverse primers (STCAT16,
5′-AAGCCGAAAGGTCAACTGC-3′; GAPDH, 5′-AAGTGGTCGTTGAGGGCAATG-3′), 2
µl cDNA and 6 µl sterile double steamed water. GAPDH was used as a
reference gene. The thermocycling conditions were the following:
95°C for 10 min, followed by 40 cycles at 95°C for 15 sec and 60°C
for 60 sec. Primers of STCAT16 were synthesized by Shanghai Ruian
BioTechnologies Co., Ltd. (Shanghai, China). The data were
quantified using the 2−ΔΔCq method (24). Each experiment was performed in
triplicate.
STCAT16 knockdown and overexpression
vectors construction
A total of four plasmids containing sequences for
short hairpin RNAs (shRNAs) targeting STCAT16 (STCAT16-shRNA-759,
STCAT16-shRNA-1161, STCAT16-shRNA-1356 and STCAT16-shRNA-1693), a
plasmid containing the negative control (NC) shRNA and a STCAT16
overexpression plasmid (STCAT16-pcDNA3.1) were constructed by
Shanghai GenePharma Co., Ltd (Shanghai, China). Cells were selected
in the logarithmic phase and transfected with 4.0 µg/well plasmid
DNA in 24-well plates (5×105 cells/well) at 85%
confluence using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Empty pcDNA3.1 plasmid was used as NC for the overexpression
experiments (mock-NC). The plasmid extraction kit was purchased
from Omega Bio-tek, Inc. (Norcross, GA, USA).
Cell proliferation assay
Cell proliferation was assessed using the Cell
Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology, Haimen,
China) according to the manufacturer's protocol. Cells growing in
the exponential phase were plated at a density of 5×104
cells/ml on 96-well plates and four plasmids (STCAT16-shRNA, sh-NC
and STCAT16-pcDNA3.1, mock-NC) were transfected with 4.0 µg/well
per plasmid DNA using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). At 0, 24, 48, 72 and 96 h, 10 µl
CCK-8 solution was added to each well followed by a 2-h incubation
at 37°C. The absorbance was recorded at 450 nm with a microplate
reader (Varioskan® Flash; Thermo Fisher Scientific,
Inc.). All experiments were performed in triplicate.
Colony formation assay
Colony formation assays were performed as previously
described (25). A total of 4.0
µg/well per plasmid DNA of four plasmids (STCAT16-shRNA, sh-NC and
STCAT16-pcDNA3.1, mock-NC) were transfected using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Cells
were seeded on 6-well plates at a concentration of 100 cells/well
and cultured at 37°C for 2–3 weeks. Surviving colonies were counted
following staining with Gentian Violet (Amresco, LLC, Solon, OH,
USA) for 5 min at room temperature. Colonies with >50 cells were
counted under an upright light microscope (magnification, ×100;
BX51 Microscope; Olympus Corporation, Tokyo, Japan). All
experiments were performed in triplicate.
Wound healing assay
Cells were plated on 6-well plates at a density of
5×105 cells/well and four plasmids (STCAT16-shRNA, sh-NC
and STCAT16-pcDNA3.1, mock-NC) were transfected with 4.0 µg/well
per plasmid DNA using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). The cell monolayers were scratched
in a straight line using a pipette tip at 24 h after transfection.
The debris was removed with three washes in PBS, and phase-contrast
images were acquired using an inverted fluorescence microscope
(IX71; Olympus Corporation, Tokyo, Japan; magnification, ×100).
Cells were cultured for an additional 24 h, and imaged at 6, 12 and
24 h in that incubation period. Subsequently, the number of
migrating cells was counted.
Cell invasion assay
STCAT16-shRNA, sh-NC and STCAT16- pcDNA3.1, mock-NC
plasmids (all 4.0 µg/well plasmid DNA) were transfected into GC
cells using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). The cell suspension containing
1×105 cells/ml transfected for 48 h was placed in an
upper chamber (Transwell®; Corning Inc., Corning, NY,
USA) precoated with Matrigel (BD Biosciences, San Jose, CA, USA). A
total of 600 µl RPMI-1640 supplemented with 20% FBS was plated in
the lower chamber. Invasive cells were able to pass through the
Matrigel layer. Subsequently, cells were incubated for 24 h and
stained with crystal violet (Beyotime Institute of Biotechnology)
at room temperature for 20 min. The field of view (n=15) of each
insert was randomly counted using an upright light microscope
(BX51; Olympus Corporation, Tokyo, Japan; magnification, ×100) and
the average value was calculated.
In vivo tumourigenesis assay
A total of 10 male BALB/c nude mice (age, 4 weeks;
weight, 18–20 g) were purchased from The Laboratory Animal Research
Centre of Nantong University (Nantong, China). Mice were maintained
at 24°C in a temperature-controlled environment with 40–60%
relative humidity, under a 12-h light/dark cycle using a dimmer,
with food and water ad libitum. Mice were randomly divided
into two groups (n=5 in each group) and injected with
STCAT16-pcDNA3.1-transfected AGS cells (STCAT16-pcDNA3.1 group) or
with empty pcDNA3.1-transfected AGS cells (mock-NC group). The
2×107 cells were subcutaneously injected into the back
of the mice. The mice with developing tumours were observed ≥3
times per week. The tumours were allowed to grow for 4 weeks, and
the tumour volume (V) was calculated using the following formula:
V=0.5 × length × width2. All animal experiments were
approved by The Animal Care and Use Committee of Nantong
University.
Immunohistochemistry
Tumour tissue sections were fixed in 10% buffered
formalin for 24 h at room temperature, embedded in paraffin and cut
into 4 µm sections. The sections were deparaffinized in xylene,
rehydrated in graded ethanol solutions and treated with 0.1%
hydrogen peroxide in methanol for 20 min. Antigen retrieval was
performed by microwaving the sections immersed in 10 mM citric acid
buffer for 10 min, followed incubation for 1 h at room temperature.
Sections were incubated with 5% bovine serum albumin (cat. no.
LLBB-1000-01, SurModics, Inc., Eden Prairie, USA) to block
non-specific protein binding for 10 min at 37°C, followed by 1-h
incubation at room temperature and overnight at 4°C with monoclonal
antibody against Ki-67 (1:50 dilution; cat. no. ab16667; Abcam,
Cambridge, UK). Sections were washed with PBS and incubated with a
horseradish peroxidase-conjugated polyclonal goat anti-rabbit
immunoglobulin G biotin secondary antibody (1:500 dilution; cat.
no. E030110-01; EarthOx, LLC, San Francisco, CA, USA) for 1 h at
room temperature, followed by 0.1% hydrogen peroxide and 0.6 mM
3,3′-Diaminobenzidine (DAB) in PBS for 8 min at room temperature.
All sections were stained using a Hematoxylin and Eosin Staining
kit (cat. no. C0105; Beyotime Institute of Biotechnology) as
previously described (26), and
examined using a light microscope (BX51; Olympus Corporation;
magnification, ×400). The nuclear was stained blue and cytoplasm
was stained red.
Regarding assessment, Ki-67 staining intensity was
evaluated using a four rating-level-scheme, where scores ranging
from 0 to 3 indicated negative, weak, medium and strong staining,
respectively. For extent of staining, a five rating-level-scheme
was employed. Thus, based on the total amount of positive stained
areas in the whole carcinoma region, the extent of staining was
evaluated with scores ranging from 0 to 4 as follows: 0, 0; 1,
1–25; 2, 26–50; 3, 51–75; and 4, 76–100%. In each specimen, five
high-power fields (×400) were randomly selected, together with
examination of nuclear staining. The product of intensity scores
multiplied by degree scores was used as the final staining score
(0–12) for Ki-67 assessment. The staining was evaluated as
previously described (27).
Statistical analysis
All experiments were performed in triplicate. All
statistical analyses were performed using SPSS software (version
17.0; SPSS, Inc., Chicago, IL, USA). Graphs were generated using
GraphPad Prism software (version 5.0; GraphPad Software, Inc., La
Jolla, CA, USA). Data are presented as the mean ± standard
deviation. Differences between two groups and in more than two
groups were evaluated using a paired Student's t-test and one-way
analysis of variance, respectively. Student-Newman-Keuls post hoc
test was performed following ANOVA. The association between the
expression level of STCAT16 and clinical characteristics was
analysed using χ2 test or Fisher's exact probability.
Parameters associated with OS and time to recurrence (TTR) were
identified using univariate and multivariate Cox proportional
hazards regression models. Kaplan-Meier plots (log-rank test) were
used to examine OS and TTR. P<0.05 was considered to indicate a
statistically significant difference.
Results
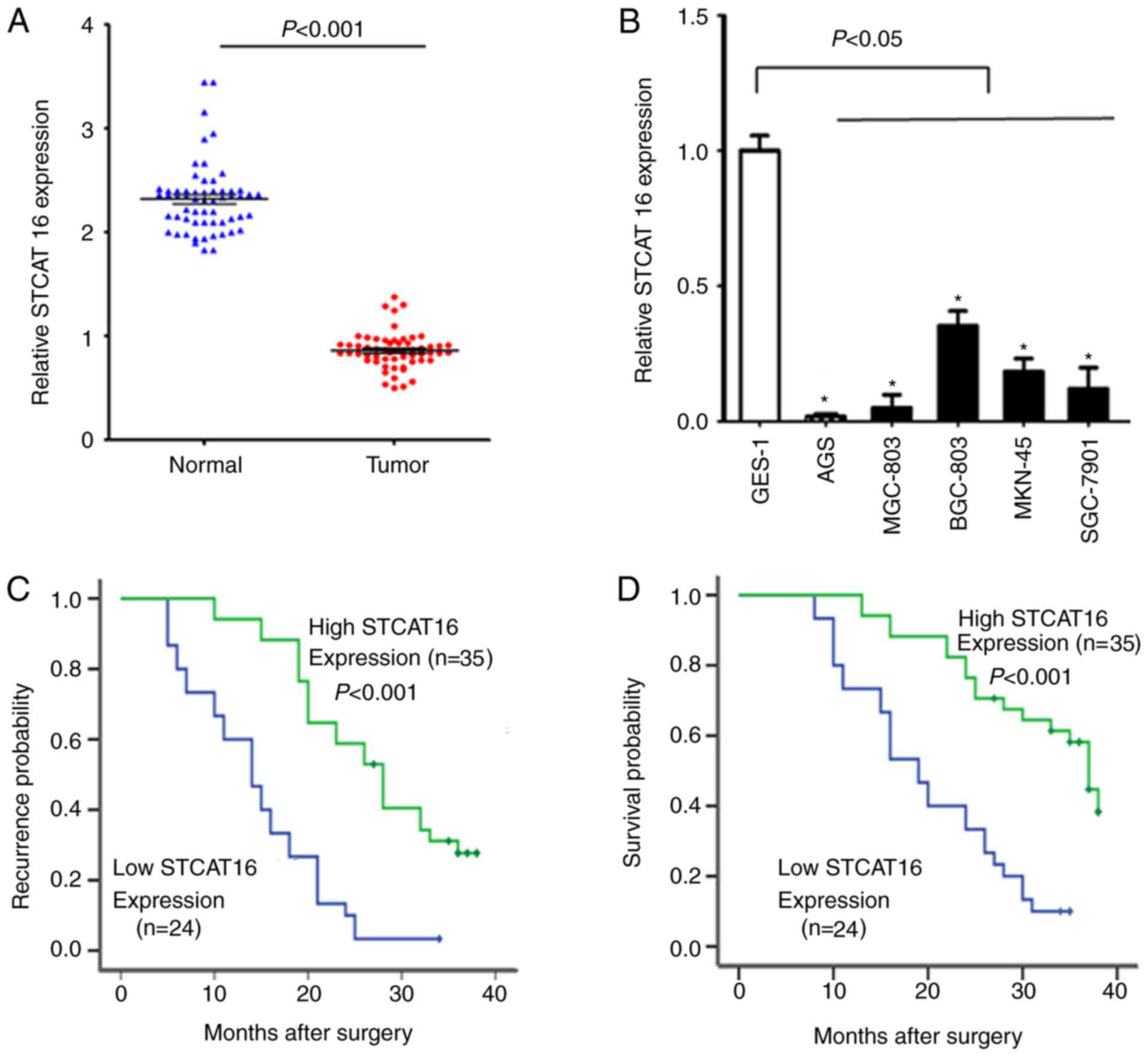
Expression level of STCAT16 is
downregulated in human gastric cancer and decreased expression
level of STCAT16 is associated with poor OS
The expression level of STCAT16 in GC tissues was
significantly decreased compared with adjacent normal gastric
tissues (Fig. 1A). The expression
level of STCAT16 varied among GC cell lines; however, all cell
lines exhibited a decreased expression level of STCAT16 compared
with GES-1 cells (Fig. 1B).
Compared with GES-1 cells, AGS and BGC-823 cells exhibited a
relative expression level of STCAT16 of 0.015±0.006 and
0.332±0.056, respectively.
The clinical features of the enrolled patients are
listed in Table I. If the
expression level of STCAT16 in the tumour tissue was decreased
>2-folds compared with the matched adjacent normal tissue, the
expression level of STCAT16 was considered as low. The expression
level of STCAT16 in GC tissues was identified to be associated with
TNM stage and lymph node metastasis. The TTR of patients with low
STCAT16 expression was significantly decreased compared with
patients exhibiting high STCAT16 expression (Fig. 1C). Additionally, OS was
significantly decreased in patients exhibiting low expression level
of STCAT16 (Fig. 1D).
 | Table I.Association between STCAT16
expression and clinical characteristics. |
Table I.
Association between STCAT16
expression and clinical characteristics.
|
|
| STCAT16 |
|
|---|
|
|
|
|
|
|---|
| Clinical
characteristics | n | High (n=35) | Low (n=24) | P-value |
|---|
| Age (years) |
|
|
| 0.598 |
|
<60 | 27 | 15 | 12 |
|
|
≥60 | 32 | 20 | 12 |
|
| Sex |
|
|
| 0.504 |
|
Male | 35 | 22 | 13 |
|
|
Female | 24 | 13 | 11 |
|
| Tumour size
(cm) |
|
|
| 0.679 |
|
<5 | 40 | 23 | 17 |
|
| ≥5 | 19 | 12 | 7 |
|
| Tumour, node and
metastasis stage |
|
|
| 0.027a |
|
I/II | 20 | 16 | 4 |
|
|
III/IV | 39 | 19 | 20 |
|
| Differentiation
degree |
|
|
| 0.400 |
| High or
medium | 26 | 17 | 9 |
|
|
Low | 33 | 18 | 15 |
|
| Lymph node
metastasis |
|
|
| 0.025a |
|
Yes | 34 | 16 | 18 |
|
| No | 25 | 19 | 6 |
|
To further investigate the prognostic potential of
STCAT16, univariate and multivariate survival analyses were
performed examining clinicopathological characteristics of patients
with GC. The univariate analysis was performed using six prognostic
factors: Age, sex, differentiation degree, lymph node metastasis,
TNM stage and STCAT16 expression level. The multivariate analysis
identified that the TNM stage and the expression level of STCAT16
were independent predictors of OS (Table II).
| Table II.Univariate and multivariate analysis
of clinicopathological characteristics and overall survival in 59
patients with gastric cancer. |
Table II.
Univariate and multivariate analysis
of clinicopathological characteristics and overall survival in 59
patients with gastric cancer.
| A, Univariate
analysis |
|---|
|
|---|
| Clinical
characteristics | Hazard ratio | 95% confidence
interval | P-value |
|---|
| Age (≥60 and
<60) | 1.101 | 0.751–2.451 | 0.931 |
| Sex | 0.461 | 0.251–1.072 | 0.442 |
| Differentiation
degree (high/middle/low) | 0.908 | 0.742–2.411 | 0.658 |
| Tumour, node and
metastasis stage (I/II/III/IV) | 5.455 | 1.890–11.080 | 0.055 |
| Lymph node
metastasis (no/yes) | 0.723 | 0.818–3.524 | 0.620 |
| STCAT16 expression
level (high/low) | 6.560 | 2.331–17.232 | 0.018 |
|
| B, Multivariate
analysis |
|
| Clinical
characteristics | Hazard
ratio | 95% confidence
interval | P-value |
|
| Age (≥60 and
<60) | 1.224 | 1.331–2.221 | 0.828 |
| Sex | 1.008 | 0.411–1.722 | 0.345 |
| Differentiation
degree (high/middle/low) | 1.244 | 0.311–3.616 | 0.120 |
| Tumour, node and
metastasis stage (I/II/III/IV) | 6.753 | 1.321–12.381 | 0.039 |
| Lymph node
metastasis (no/yes) | 1.232 | 0.223–2.919 | 0.237 |
| STCAT16 expression
level (high/low) | 7.393 | 1.928–15.156 | 0.025 |
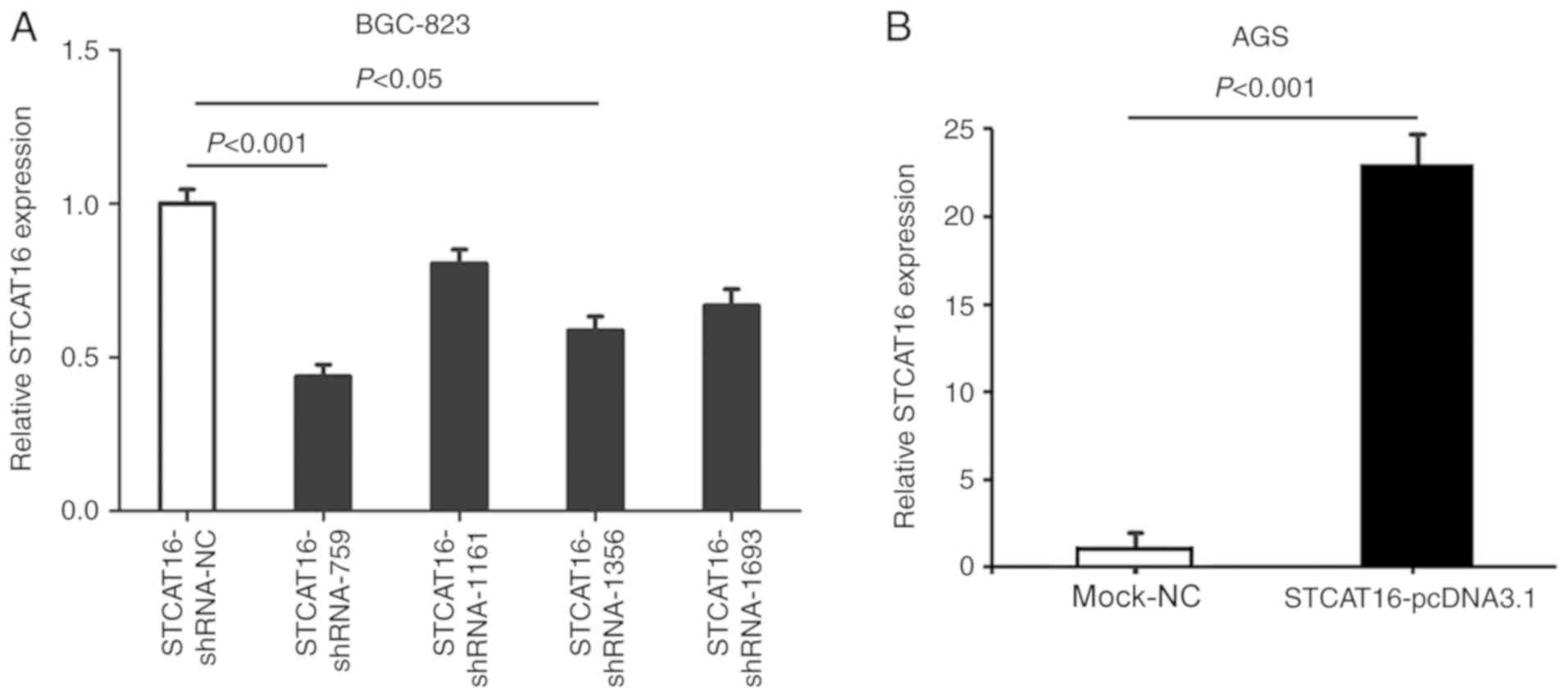
STCAT16 interference and
overexpression
To identify the most effective shRNA, four
STCAT16-shRNAs plasmids were transfected into BGC-823 cell line,
which exhibited an increased expression level of STCAT16. Among the
four shRNAs, STCAT16-shRNA-759 exhibited the most effective
knockdown compared with the negative control (STCAT16-sh-NC) group
(Fig. 2A). Therefore,
STCAT16-shRNA-759 was used in further experiments.
Furthermore, STCAT16-pcDNA3.1 plasmid was
transfected into AGS cells, which exhibited a decreased expression
level of STCAT16 compared with other GC cell lines. The expression
level of STCAT16 was significantly increased in the
STCAT16-pcDNA3.1 group compared with the mock-NC group, which was
transfected with an empty vector.
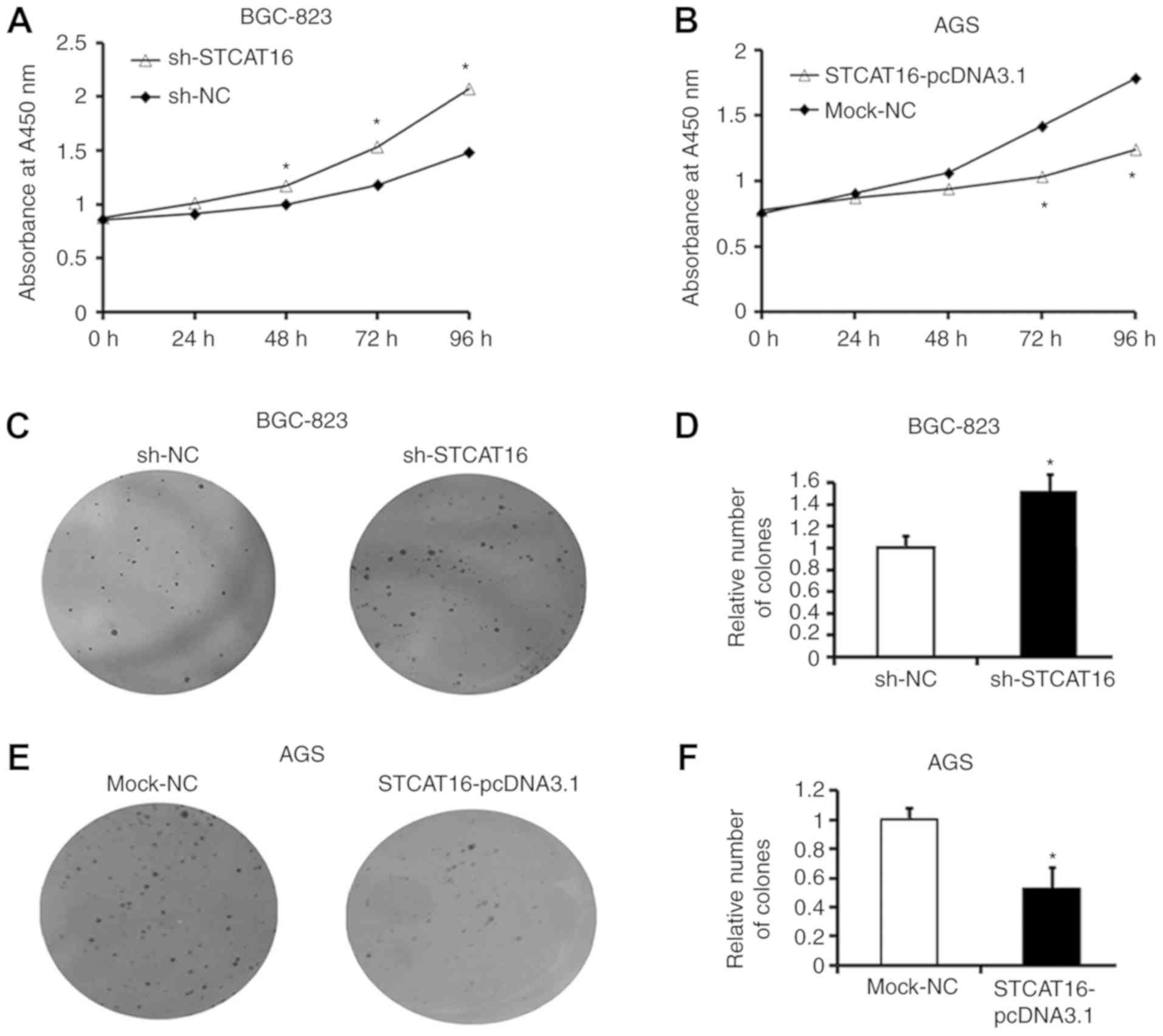
STCAT16 overexpression inhibits
gastric cancer cell proliferation and colony formation
To investigate the biological effects of STCAT16 on
GC cells, the expression level of STCAT16 was silenced by
transfecting sh-STCAT16 into BGC-823 cells and overexpressed by
transfecting STCAT16-pcDNA3.1 into AGS cells.
Following transfection with sh-STCAT16, the
proliferation of BGC-823 cells was significantly increased at 48,
72 and 96 h compared with the sh-NC group (Fig. 3A). In contrast, the proliferation
of AGS cells was significantly decreased at 48, 72 and 96 h
following transfection with STCAT16 overexpressing vector Fig. 3B).
A colony formation assay produced similar results.
Following sh-STCAT16 transfection, BGC-823 cells exhibited a
significantly increased number of colonies (Fig. 3C and D). Conversely, following
STCAT16-pcDNA3.1 transfection, AGS cells exhibited a significantly
decreased number of colonies compared with the mock-NC group
(Fig. 3E and F).
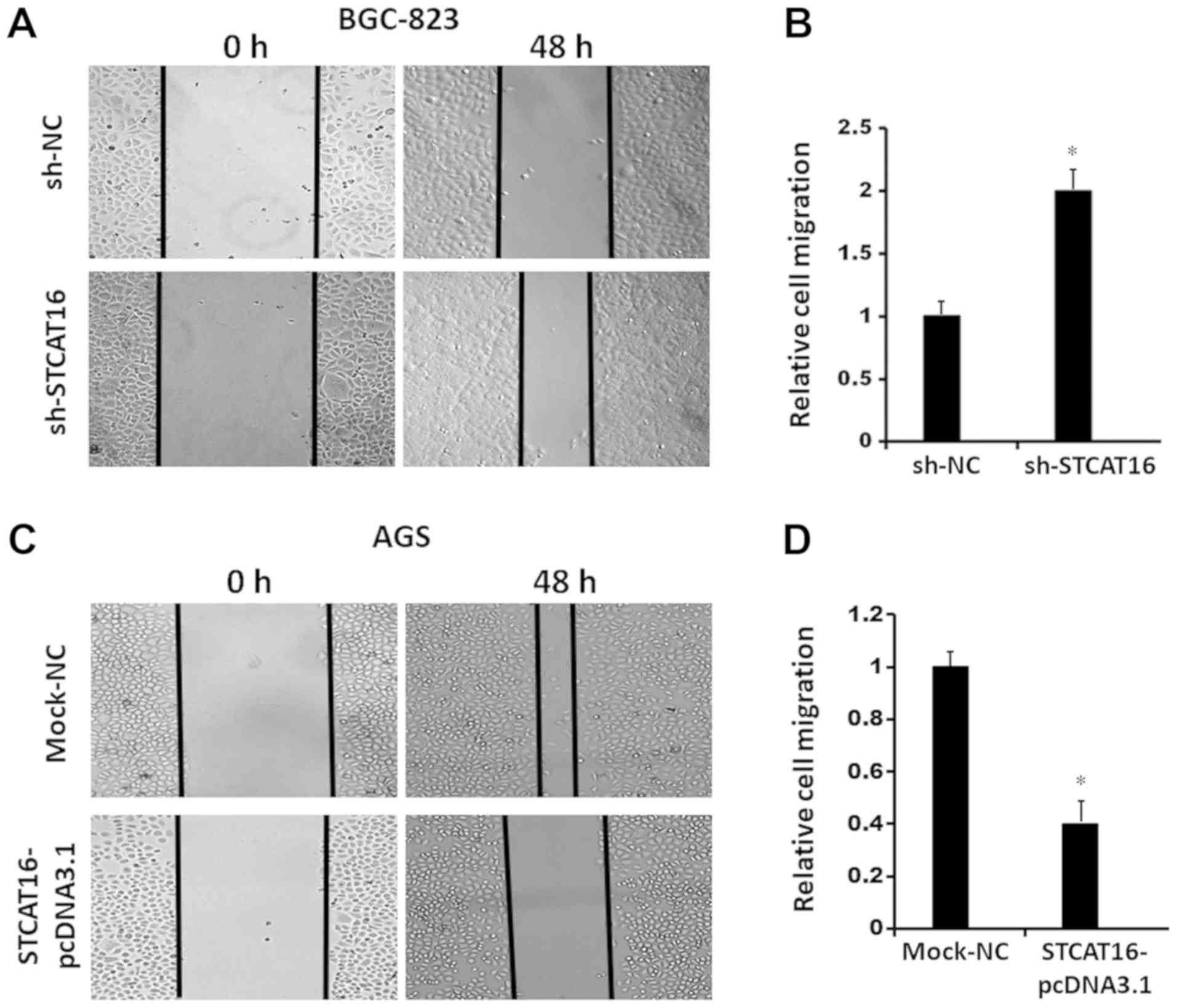
STCAT16 overexpression suppresses
gastric cancer cell migration and invasion in vitro
To investigate cell migration and invasion, wound
healing assays and a Matrigel assays were performed, respectively
(Fig. 4). The results obtained
from the wound-healing assay and the invasion assay suggested that
decreasing the expression level of STCAT16 promoted BGC-823 cells
migration (Fig. 4A and B) and
invasion (Fig. 4E and F).
Conversely, the migration (Fig. 4C and
D) and invasion (Fig. 4G and
H) of AGS cells were decreased following STCAT16
overexpression.
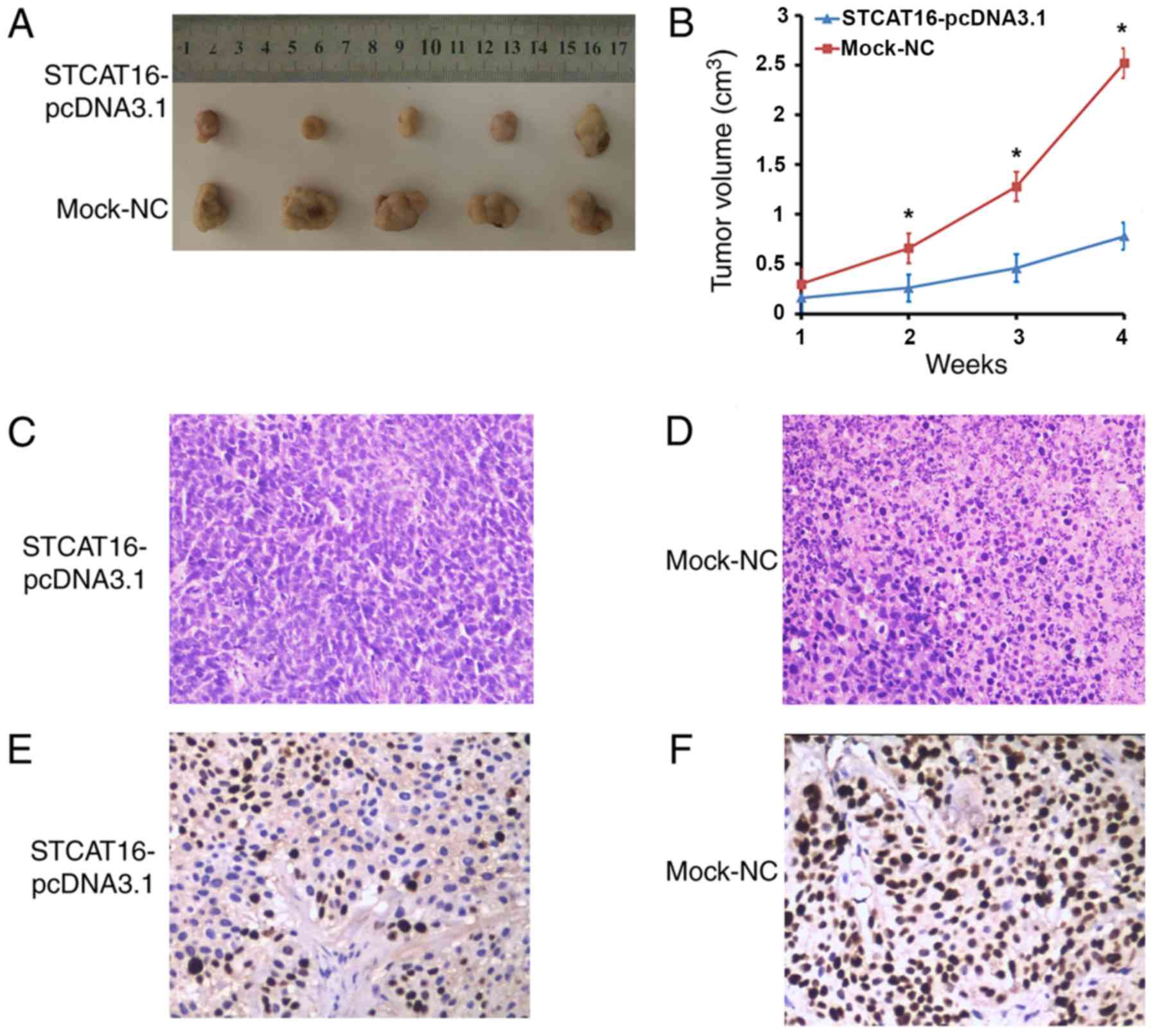
Overexpression of STCAT16 inhibits
tumour progression in BALB/c nude mice
To further investigate the tumour suppressive role
of STCAT16, AGS cells transfected with STCAT16-pcDNA3.1 vector or
empty vector were injected into BALB/c nude mice. Tumours were
detected in all animals injected with AGS cells.
In the mouse xenograft models, the size and the
growth of tumours decreased in the STCAT16-pcDNA3.1 group compared
with the mock-NC group at 2, 3 and 4 weeks (Fig. 5A and B). Following 4 weeks, the
largest tumours exhibited a volume of 3,005.6 and 1,188.1
mm3 in the STCAT16-pcDNA3.1 group and in the mock-NC
group, respectively. The xenografts were fixed with formalin,
embedded in paraffin, stained with haematoxylin and eosin
(H&E), and incubated with MKI67 antibody. H&E staining
indicated that the tumor tissue in the STCAT16-pcDNA3.1 transfected
group had a smaller area of necrosis, higher differentiation, less
pleomorphism and fission, comparing with that in mock-NC group
(Fig. 5C and D). Furthermore,
tumours derived from cells transfected with STCAT16-pcDNA3.1
exhibited reduced growth compared with the mock group (4.21±0.90
and 9.80±1.23, respectively, P<0.05; Fig. 5E and F). The present result
suggested that overexpression of STCAT16 significantly inhibited
tumour growth in BALB/c nude mice.
Discussion
Accumulating evidence (28,29)
has demonstrated that the dysregulation of lncRNAs is involved in
the occurrence and development of GC and may promote tumour
invasion and metastasis. The overexpression of the lncRNA HOTAIR in
GC cells was identified to be associated with TNM stages and lymph
node metastasis and to promote colony formation and hepatic
metastasis (30). In contrast,
knockdown of HOTAIR was identified to reverse
epithelial-mesenchymal transition by regulating the expression
levels of matrix metalloproteinase (MMP)1 and 3 (31). The upregulation of the lncRNA GHET1
was identified to be associated with gastric tumour growth and
invasion and GHET1 overexpression increased the proliferation of GC
cells by promoting the interaction between the transcript of MYC
proto-oncogene, bHLH transcription factor (MYC) and insulin like
growth factor 2 mRNA binding protein 1 (17). HULC expression was identified in
hepatocellular cancer; however, it was also demonstrated to be
upregulated in GC (32).
Additionally, HULC promoted lymph node metastasis and distant
metastasis and was identified to be associated with advanced GC
(18). A previous study
demonstrated that the lncRNA CCAT1 promoted GC progression by
upregulating the expression level of MYC (19). The downregulation of the lncRNA
growth arrest specific 5 inhibited the proliferation and induced
the apoptosis of GC cells by regulating the activities of cyclin
dependent kinase inhibitor 1A and E2F transcription factor 1
(33). The lncRNA GATA6 antisense
RNA 1 (head to head) was identified to be downregulated in GC, and
its downregulation may lead to tumour-associated phenotypes and
metastasis by regulating the expression level of MMP9 (34). The downregulation of the lncRNA
MEG3 was demonstrated in GC tissues in vitro and in
vivo (20,35).
In the present study, the expression level of the
novel lncRNA STCAT16 was significantly downregulated in GC and was
identified to be associated with poor clinical features. STCAT16
was additionally downregulated in GC cell lines. The downregulation
of STCAT16 promoted cellular proliferation, migration and invasion,
and colony formation in vitro. Notably, these effects were
reversed following overexpression of STCAT16. In vivo, the
tumour sizes in the mice injected with cells overexpressing STCAT16
were decreased compared with tumours derived from cells transfected
with an empty vector. Furthermore, overexpressing STCAT16
significantly inhibited tumour cell proliferation in BALB/c nude
mice. Collectively, the present results suggested that STCAT16 may
serve as a tumour suppressor in GC.
Following the identification of dysregulated lncRNAs
in GC, it is required to investigate the mechanisms underlying the
function of these noncoding RNAs in tumour development and
progression. In previous studies, lncRNAs were identified to serve
multiple roles by altering gene expression at the transcriptional,
by interacting with mRNA molecules, and post-transcriptional
levels, by binding to promoter regions (36–38).
Additionally, lncRNAs may be involved in post-translational
regulation by directly binding to certain proteins involved in
tumour progression (36–38). STCAT16 may act as an oncosuppressor
via the discussed mechanisms. Further studies are required to
investigate the molecular mechanism underlying STCAT16 function in
GC.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Youth Foundation
of Affiliated Hospital of Nantong University (grant no. TDFY0332),
the Hospital Talent Training Foundation (grant no. 2015-68), the
Youth Foundation of Nantong Municipal Commission of Health and
Family Planning (grant no. WQ2016079) and the China Postdoctoral
Science Foundation (grant no. 2018M630592).
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
JFZ, WJ, ZBM and ZWW conceived and designed the
study. JFZ, WJ, QFZ and XLK performed the experiments. JFZ and WJ
wrote the manuscript. ZBM and ZWW reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Ethical approval was obtained from the independent
ethics committees at Affiliated Hospital of Nantong University. All
patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nie Y, Wu K, Yu J, Liang Q, Cai X, Shang
Y, Zhou J, Pan K, Sun L, Fang J, et al: A global burden of gastric
cancer: The major impact of China. Expert Rev Gastroenterol
Hepatol. 11:651–661. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li Q, Chen C, Kapadia A, Zhou Q, Harper
MK, Schaack J and LaBarbera DV: 3D models of epithelial-mesenchymal
transition in breast cancer metastasis: High-throughput screening
assay development, validation, and pilot screen. J Biomol Screen.
16:141–154. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Klein CA: Cancer. The metastasis cascade.
Science. 321:1785–1787. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang C, Wang L, Ding Y, Lu X, Zhang G,
Yang J, Zheng H, Wang H, Jiang Y and Xu L: LncRNA structural
characteristics in epigenetic regulation. Int J Mol Sci. 18(pii):
E26592017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vance KW and Ponting CP: Transcriptional
regulatory functions of nuclear long noncoding RNAs. Trends Genet.
30:348–355. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoon JH, Abdelmohsen K and Gorospe M:
Posttranscriptional gene regulation by long noncoding RNA. J Mol
Biol. 425:3723–3730. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang JZ, Chen M, Chen, Gao XC, Zhu S,
Huang H, Hu M, Zhu H and Yan GR: A peptide encoded by a putative
lncRNA HOXB-AS3 suppresses colon cancer growth. Mol Cell.
68:171–184.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huarte M and Rinn JL: Large non-coding
RNAs: Missing links in cancer. Hum Mol Genet. 19:R152–R161. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pan W, Liu L, Wei J, Ge Y, Zhang J, Chen
H, Zhou L, Yuan Q, Zhou C and Yang M: A functional lncRNA HOTAIR
genetic variant contributes to gastric cancer susceptibility. Mol
Carcinog. 55:90–96. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang F, Xue X, Zheng L, Bi J, Zhou Y, Zhi
K, Gu Y and Fang G: Long non-coding RNA GHET1 promotes gastric
carcinoma cell proliferation by increasing c-Myc mRNA stability.
FEBS J. 281:802–813. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao Y, Guo Q, Chen J, Hu J, Wang S and
Sun Y: Role of long non-coding RNA HULC in cell proliferation,
apoptosis and tumor metastasis of gastric cancer: A clinical and
in vitro investigation. Oncol Rep. 31:358–364. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang F, Xue X, Bi J, Zheng L, Zhi K, Gu Y
and Fang G: Long noncoding RNA CCAT1, which could be activated by
c-Myc, promotes the progression of gastric carcinoma. J Cancer Res
Clin Oncol. 139:437–445. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peng W, Si S, Zhang Q, Li C, Zhao F, Wang
F, Yu J and Ma R: Long non-coding RNA MEG3 functions as a competing
endogenous RNA to regulate gastric cancer progression. J Exp Clin
Cancer Res. 34:792015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu G, Meng L, Yuan D, Li K, Zhang Y, Dang
C and Zhu K: MEG3/miR-21 axis affects cell mobility by suppressing
epithelial-mesenchymal transition in gastric cancer. Oncol Rep.
40:39–48. 2018.PubMed/NCBI
|
|
22
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nat Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
In H, Solsky I, Palis B, Langdon-Embry M,
Ajani J and Sano T: Validation of the 8th edition of the AJCC TNM
staging system for gastric cancer using the national cancer
database. Ann Surg Oncol. 24:3683–3691. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang JF, Qu LS, Qian XF, Xia BL, Mao ZB
and Chen WC: Nuclear transcription factor CDX2 inhibits gastric
cancer-cell growth and reverses epithelial-to-mesenchymal
transition in vitro and in vivo. Mol Med Rep. 12:5231–5238.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo P, Wang J, Gao W, Liu X, Wu S, Wan B,
Xu L and Li Y: Salvianolic acid B reverses multidrug resistance in
nude mice bearing human colon cancer stem cells. Mol Med Rep.
18:1323–1334. 2018.PubMed/NCBI
|
|
27
|
Zhang J, Ding W, Kuai X, Ji Y, Zhu Z, Mao
Z and Wang Z: Dermcidin as a novel binding protein of lncRNA STCAT3
and its effect on prognosis in gastric cancer. Oncol Rep.
40:2854–2863. 2018.PubMed/NCBI
|
|
28
|
Abbastabar M, Sarfi M, Golestani A and
Khalili E: lncRNA involvement in hepatocellular carcinoma
metastasis and prognosis. EXCLI J. 17:900–913. 2018.PubMed/NCBI
|
|
29
|
Gong P, Qiao F, Wu H, Cui H, Li Y, Zheng
Y, Zhou M and Fan H: LncRNA UCA1 promotes tumor metastasis by
inducing miR-203/ZEB2 axis in gastric cancer. Cell Death Dis.
9:11582018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shao Y, Chen H, Jiang X, Chen S, Li P, Ye
M, Li Q, Sun W and Guo J: Low expression of lncRNA-HMlincRNA717 in
human gastric cancer and its clinical significances. Tumour Biol.
35:9591–9595. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu ZY, Yu QM, Du YA, Yang LT, Dong RZ,
Huang L, Yu PF and Cheng XD: Knockdown of long non-coding RNA
HOTAIR suppresses tumor invasion and reverses
epithelial-mesenchymal transition in gastric cancer. Int J Biol
Sci. 9:587–597. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Klec C, Gutschner T, Panzitt K and Pichler
M: Involvement of long non-coding RNA HULC (highly up-regulated in
liver cancer) in pathogenesis and implications for therapeutic
intervention. Expert Opin Ther Targets. 23:177–186. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu L, Yan B, Yang Z, Zhang X, Gu Q and
Yue X: ncRuPAR inhibits gastric cancer progression by
down-regulating protease-activated receptor-1. Tumour Biol.
35:7821–7829. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Park SM, Park SJ, Kim HJ, Kwon OH, Kang
TW, Sohn HA, Kim SK, Moo Noh S, Song KS, Jang SJ, et al: A known
expressed sequence tag, BM742401, is a potent lincRNA inhibiting
cancer metastasis. Exp Mol Med. 45:e312013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dan J, Wang J, Wang Y, Zhu M, Yang X, Peng
Z, Jiang H and Chen L: LncRNA-MEG3 inhibits proliferation and
metastasis by regulating miRNA-21 in gastric cancer. Biomed
Pharmacother. 99:931–938. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moran VA, Perera RJ and Khalil AM:
Emerging functional and mechanistic paradigms of mammalian long
non-coding RNAs. Nucleic Acids Res. 40:6391–6400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu J and Hann SS: Functions and Roles of
long-non-coding RNAs in human nasopharyngeal carcinoma. Cell
Physiol Biochem. 45:1191–1204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013. View Article : Google Scholar : PubMed/NCBI
|