Introduction
Osteosarcoma (OS) is one of the most common types of
primary malignant bone tumors, accounting for 60% of malignant bone
tumors in adolescents (1). The
management of OS remains challenging, particularly under conditions
of metastases, and is often complicated by local recurrence
following treatment. With continuing developments in adjuvant
chemotherapy (2) and improvements
in surgical techniques, the 5-year survival rates have risen from
<20% to ~60–80% (3); however,
distant metastases may occur at early stages of the condition,
while recurrence is often detected without distant metastases
(4). For such patients, treatment
outcomes remain poor (4)
Adriamycin (ADM) is an important chemotherapeutic
agent used in the treatment of OS. ADM affects the structure and
function of DNA via intercalation, prevents DNA replication and RNA
synthesis, and induces apoptosis in tumor cells (5). As ADM possesses a narrow therapeutic
index, severe cardiotoxicity and bone marrow suppression are common
side effects of treatment (6).
SM-164 is a novel non-peptide, symmetric alkyne small molecule. The
binding capacity of SM-164 to the baculovirus inhibitor of
apoptosis protein repeat (BIR) domain is ~300–7,000 times that of
monovalent Smac mimetics and of the AVPI peptide of wild Smac
(7). SM-164 notably induces
apoptosis in Smac-sensitive tumor cells without inflicting marked
toxicity to normal cells (8). Its
key mechanism of action involves the inhibition of the X-linked
inhibitor of apoptosis protein (XIAP) (9).
The family of inhibitor of apoptosis (IAP) proteins
contains some of the most important apoptosis inhibitors, including
XIAP, cellular inhibitor of apoptosis protein (cIAP)-1 and −2,
neuronal apoptosis inhibitor protein, survivin, livin/melanoma-IAP
and apollon, all of which mainly inhibit caspase activity and the
induction of the apoptotic pathway (10). Overexpression of IAPs has been
reported in a variety of tumor cells, and is an important cause of
tumor cell apoptosis and chemoresistance (11–13).
At present, XIAP is the most potent known member of IAPs, while
survivin has a strong inhibitory effect on apoptosis (14).
The present study investigated how combined SM-164
and ADM treatment, as well as XIAP silencing, affected U2-OS cells.
The results may provide insight for the development of novel
chemotherapeutic and genetic treatment strategies in the management
of OS.
Materials and methods
Cell cultures
U2-OS cells were purchased from The Cell Bank of
Type Culture Collection of Chinese Academy of Sciences (Beijing,
China) and cultured in RPMI-1640 (KGM31800S-500; Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China). The culture was supplemented
with 10% fetal bovine serum (FBS; cat. no. 04-007-1A; Biological
Industries USA, Inc., Cromwell, CT, USA) and 100 U/ml
penicillin-streptomycin (cat. no. P1400; Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China) in 5% CO2 at
37°C. Cells that were 70% confluent were used in experiments.
Cultured cells were divided into four groups:
Control, ADM, SM-164 (200 nM), and combined treatment (0.5 µg/ml
ADM + 200 nM SM-164). Following the addition of ADM into cell media
for 2 h at 37°C, cells were washed and then treated with SM-164
(200 nM) for 24 h at 37°C.
MTT assay
Cells (3×103/ml) were seeded in 96-well
plates. After the indicated treatments, an MTT assay was applied to
evaluate cell viability as previously described (15). The optical density (OD) was
determined via a microplate reader (Bio-Tek Instruments, Inc.,
Winooski, VT, USA) at 570 nm and represented cell viability.
Flow cytometry
Following the indicated treatments, U2-OS cells were
collected after trypsin digestion (0.25%, 2 min at 37°C) and
underwent centrifugation (780 × g for 3 min at room temperature); 5
µl of Annexin V-fluorescein isothiocyanate and 5 µl propidium
iodide were subsequently added to each tube for 5 min at room
temperature. After light mixing, apoptosis was detected within 1 h
by FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes, NJ,
USA) and analyzed by FlowJo version 10 (FlowJo LLC, Ashland, OR,
USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following the indicated treatments, U2-OS cells were
collected after trypsin digestion (0.25%, 2 min at 37°C). Total RNA
was extracted with a TRIzol (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Subsequently, RNA was
amplified using a one-step RT-PCR kit (cat. no. 00081405; CWBIO,
Taizhou, China). Primers were added into a 25-µl ULtraSYBR Mixture
(cat. no. 01170; CWBIO, Beijing, China). qPCR was conducted as
follows using an Applied Biosystems 7500 (Thermo Fisher Scientific,
Inc.) for 6 repeats: 95°C denaturation for 10 sec, 56°C annealing
for 30 sec, and 72°C extension for 30 sec (40 cycles). The primer
sequences were listed in Table I.
The quantification cycle value for each gene was detected and the
expression levels of target genes were calculated using the
2−ΔΔCq method (16,17).
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Genes | Primers (5′-3′) |
|---|
| cIAP-1-F |
CTCGTGGAGTGGAAGACA |
| cIAP-1-R |
GGAGTGATCGTGGTAAGG |
| XIAP-F |
TGTGGGAAACAGAAATCA |
| XIAP-R |
GGGCTTAAATGGGCATAG |
| GAPDH-F |
GAAGGTCGGAGTCAACGGAT |
| GAPDH-R |
CCTGGAAGATGGTGATGGG |
Western blot analysis
Protein was extracted from cells for western
blotting using a protein isolation kit according to the
manufacturer's protocols (cat. no. C1053; Applygen Technologies
Inc., Beijing, China) and concentrations were determined using a
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.). SDS-PAGE (12%) was used to separate 25 µg of protein from
each group. Samples were then transferred onto nitrocellulose
membranes for western blot analysis. After a 2-h blocking in 5%
bovine serum albumin (BSA) at room temperature, membranes were
incubated with the following primary antibodies: Anti-GAPDH
(1/2,000; TA-08; OriGene Technologies, Inc., Beijing, China);
anti-caspase 7 (1/2,000; ab25900; Abcam, Cambridge, UK);
anti-active caspase 9 (1/5,000; ab2324, Abcam); anti-active caspase
3 (1/10,000; ab2302; Abcam); anti-poly (ADP-ribose) polymerase
(PARP; 1/5,000; ab32138; Abcam); anti-XIAP (1/500; bs-1281R; BIOSS,
Beijing, China); anti-cIAP1 (1/5,000; ab108361; Abcam) and
anti-survivin (1/5,000; ab76424; Abcam). Following incubation with
primary antibodies at 4°C overnight, nitrocellulose membranes were
washed three times and incubated with a secondary antibody
(horseradish peroxidase-labeled goat anti-rabbit IgG; cat. no.
A16104SAMPLE; Thermo Fisher Scientific, Inc.) at 4°C for 2 h. Bands
were visualized using an enhanced chemiluminescence kit (Thermo
Fisher Scientific, Inc.). Blot densities were quantified using
Quantity One software (v4.62; Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
XIAP silencing
Short hairpin RNA (shRNA) was used to silence the
expression of XIAP; the sequences were: Forward
5′-CACCCATGCAGCTGTAGATAGATGGCAATCGAAATTGCCATCTATCTACA-3′ and
reverse, 5′-AAAACATGCAGCTGTAGATAGATGGCAATTTCGATTGCCATCTATCTACA-3′
(Sangon Biotech Co., Ltd., Shanghai, China). After annealing the
double chain DNA at 56°C for 30 sec, the sequences were integrated
into the lentiviral skeleton carrier PDS019_pL_shRNA_F (Shanghai
Novobio Co., Ltd., Shanghai, China). Recombinant lentiviral plasmid
liposomes (Invitrogen; Thermo Fisher Scientific, Inc.) were
transfected into 293T cells (American Type Culture Collection,
Manassas, VA, USA) to produce recombinant lentivirus
PDS019-PL-shRNA-GFP-homo- XIAP. Viral replication was detected by
fluorescence microscopy, while the titer was determined to be
1.8×1011 pfu/ml according to a 50% tissue culture
infective dose method (18). Cells
(1×105/ml) were seeded in 96-well plates. The lentivirus
was applied for transfection 24 h later. On the second day, 2 ml of
original medium (RPMI-1640) was replaced with medium containing 6
mg/ml Polybrene. An appropriate amount of viral suspension was
added. After 4 h, 2 ml of fresh medium was added to dilute the
Polybrene. Fresh medium was added 2 h later.
U2-OS cells were treated with 0.5 µg/ml ADM + 200 nM
SM-164 as aforementioned. Following the addition of ADM into
cellular media for 2 h, cells were washed with PBS and treated with
SM-164 (200 nM). Cells were further divided into three groups after
24 h: ADM + SM-164, ADM + SM-164 + vector (empty), and ADM + SM-164
+ shRNA-XIAP (sh-XIAP) groups. MTT, RT-qPCR and western blotting
were performed as aforementioned 24 h after transfection.
Statistical analysis
All numerical data from six repeated experiments
were expressed as the mean ± standard deviation. Statistical
analyses were performed using one-way analysis of variance followed
by a Bonferroni post-hoc test. Analysis was conducted using SPPS
software version 17 (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Combination of SM-164 with ADM reveals
enhanced inhibition of cell viability
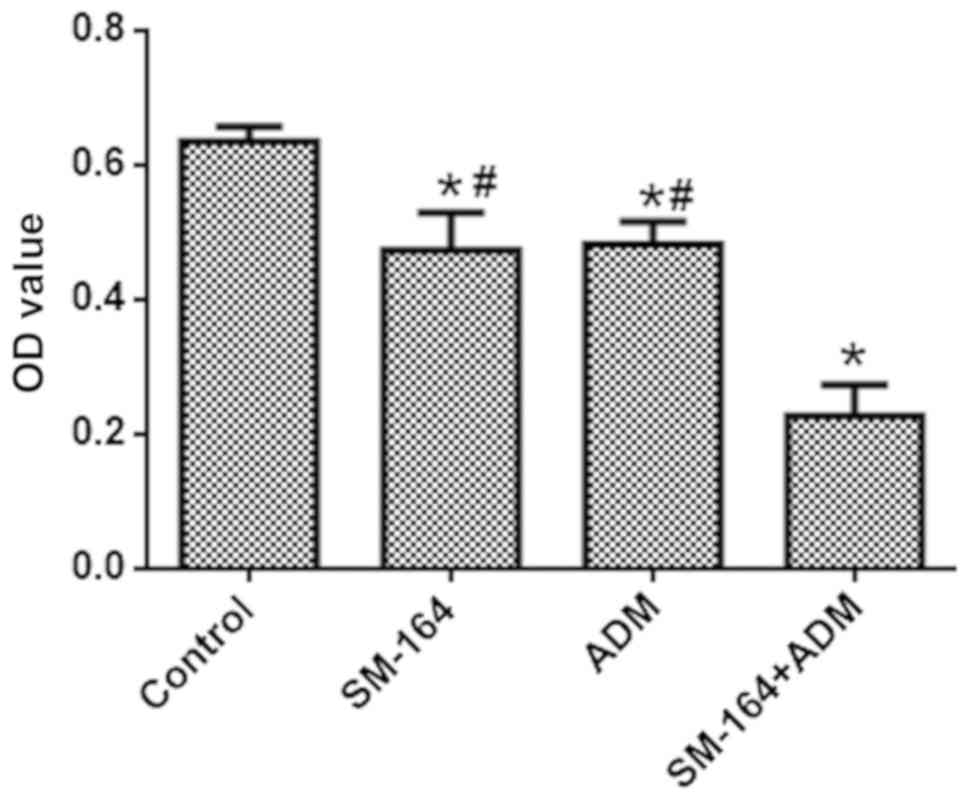
As presented in Fig.
1, the OD values of the groups respectively treated with ADM
and SM-164 were significantly decreased compared with the control
group; the OD value decreased further in the combined treatment
group. These data suggest that a combination of SM-164 and ADM
reveals enhanced inhibition of cell viability compared with single
application of SM-164 or ADM.
Combined SM-164 and ADM enhances
apoptosis
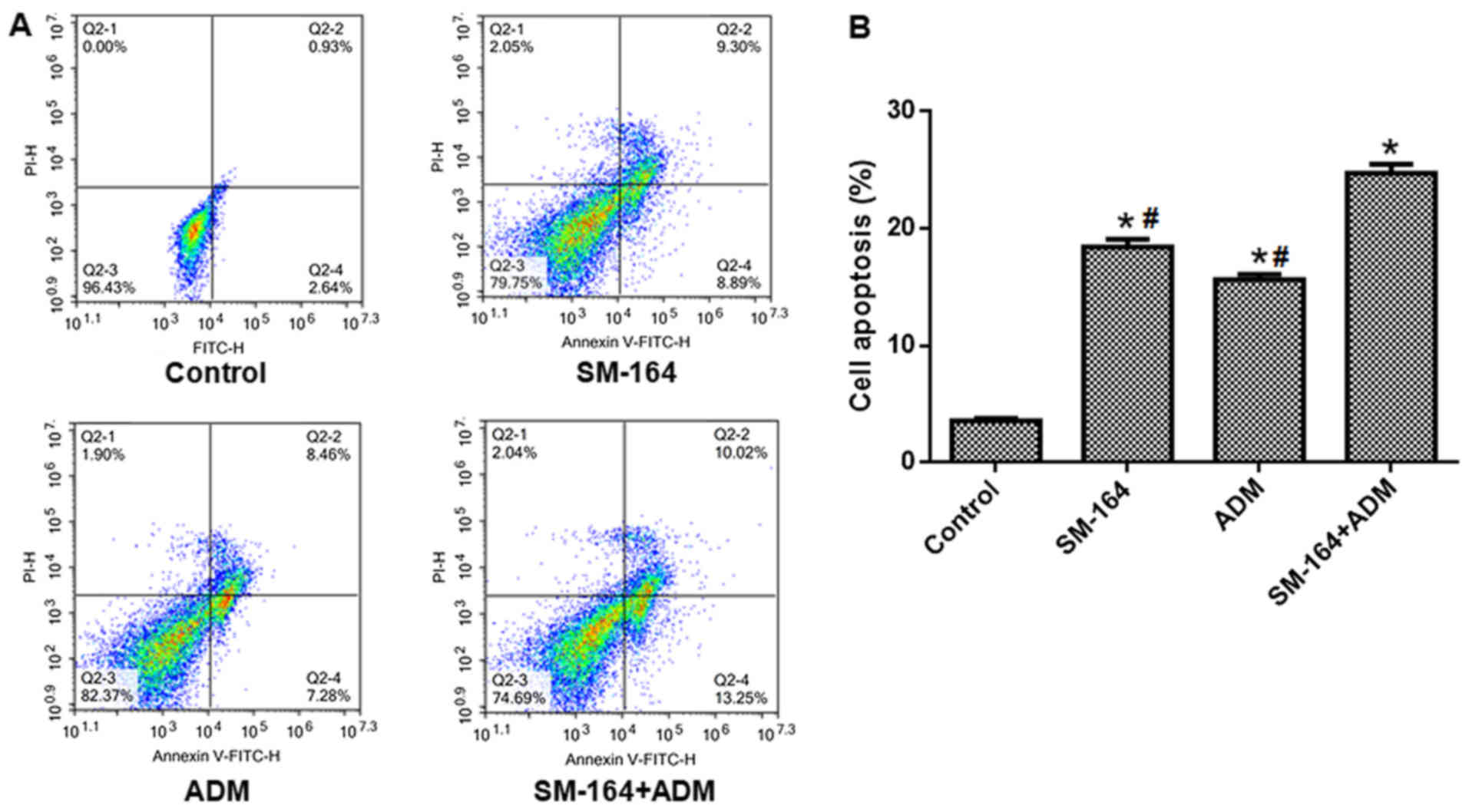
As presented in Fig.
2, the apoptosis of cells treated with ADM or SM-164 alone
increased significantly compared with the control group, but was
significantly reduced than under conditions of combined treatment.
These data suggest that combination of SM-164 with ADM reveals
enhanced apoptosis compared with single application of SM-164 or
ADM.
Combined SM-164 and ADM treatment
exhibits enhanced effects on caspases-7, −9, −3, PARP, survivin and
cIAP-1 expression
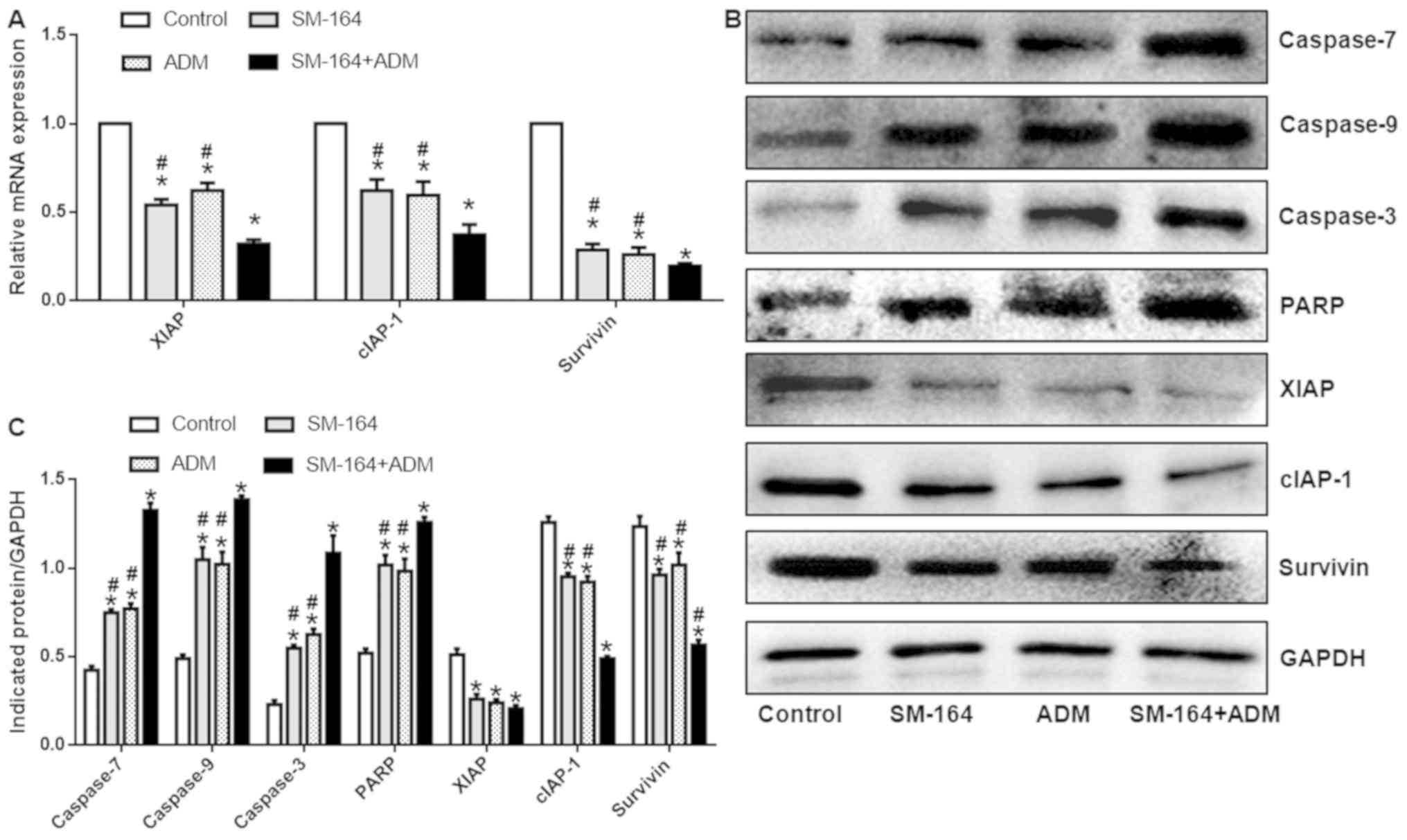
The mRNA expression levels of XIAP, cIAP-1 and
survivin were significantly decreased following ADM or SM-164
treatment alone; however, expression was significantly increased
compared with combined treatment (Fig.
3A). In addition, the protein expression levels of caspases-7,
−9, and-3, and PARP increased significantly, while that of XIAP,
survivin and cIAP-1 decreased significantly in groups treated with
ADM or SM-164, compared with the control. Of note, the expression
of these proteins was significantly upregulated following treatment
with ADM or SM-164 compared with combined treatment (Fig. 3B and C). These data suggest that
combination of SM-164 and ADM demonstrates increased effects on
apoptosis-related protein expression compared with single
application of SM-164 or ADM.
Silencing of XIAP further suppresses
viability and promotes apoptosis
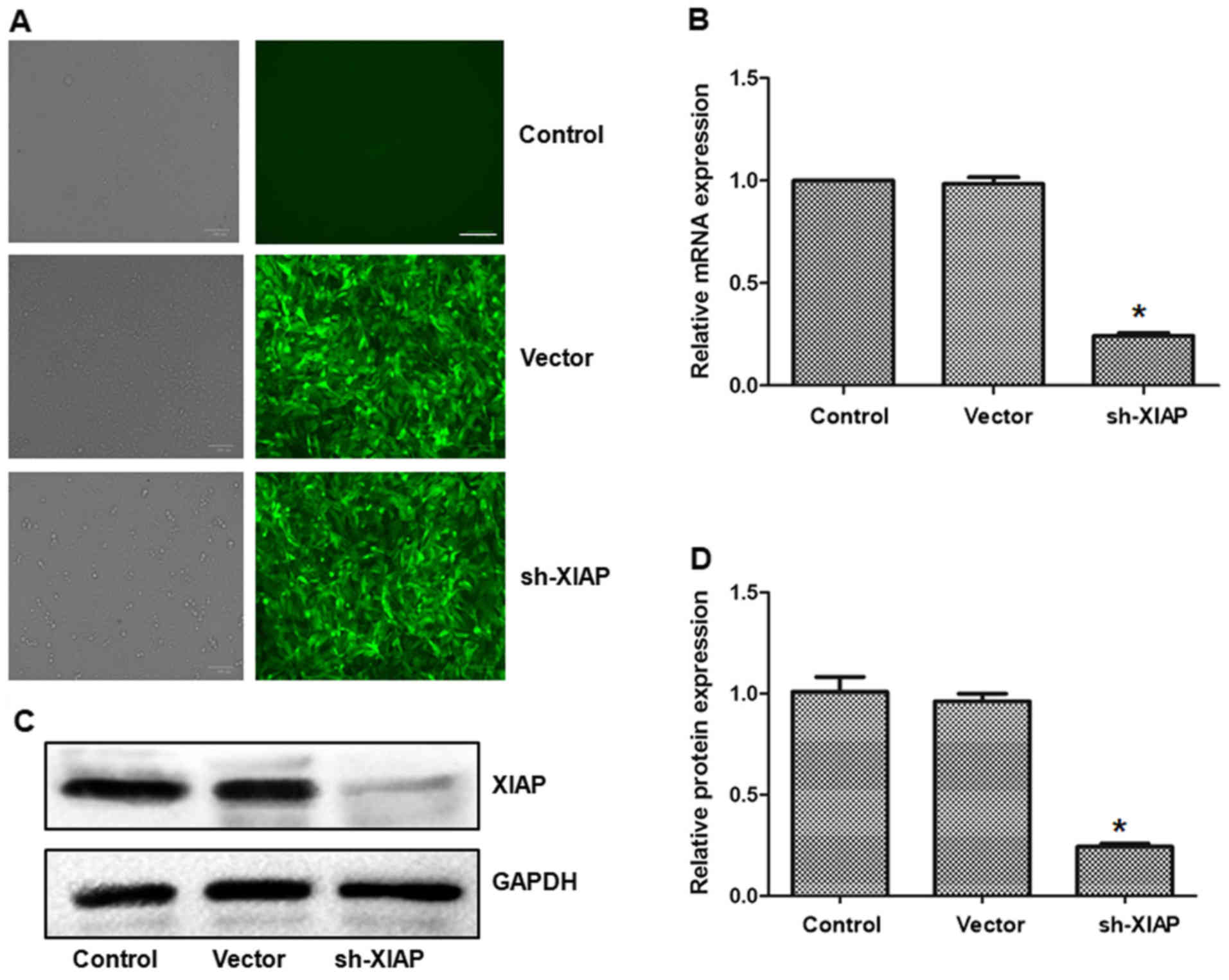
As presented in Fig.
4A, cells in the vector and XIAP-silencing groups were
successfully transfected. The expression of XIAP in the sh-XIAP
group was significantly reduced at the mRNA and protein levels than
in the ADM + SM-164 group (P<0.05; (Fig. 4B-D).
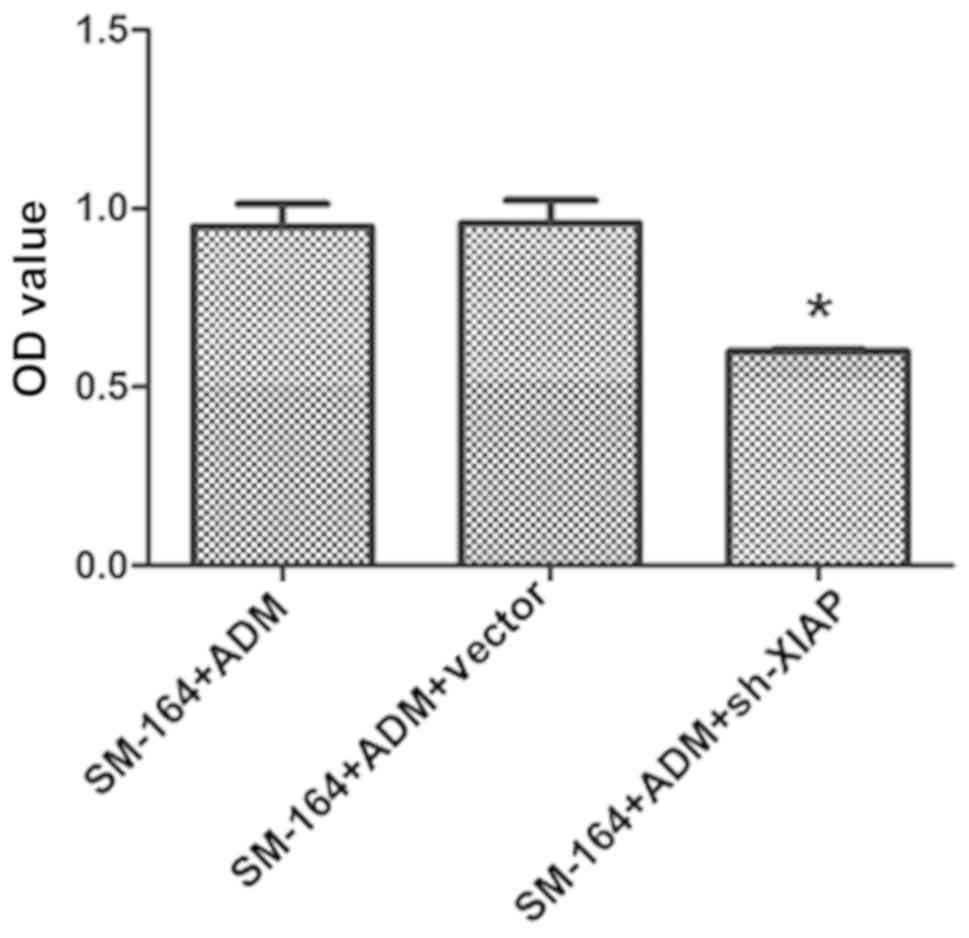
As presented in Fig.
5, the OD value of the ADM + SM-164 + XIAP silencing group
decreased significantly, and the difference was statistically
significant compared with the ADM + SM-164 group (P<0.05). As
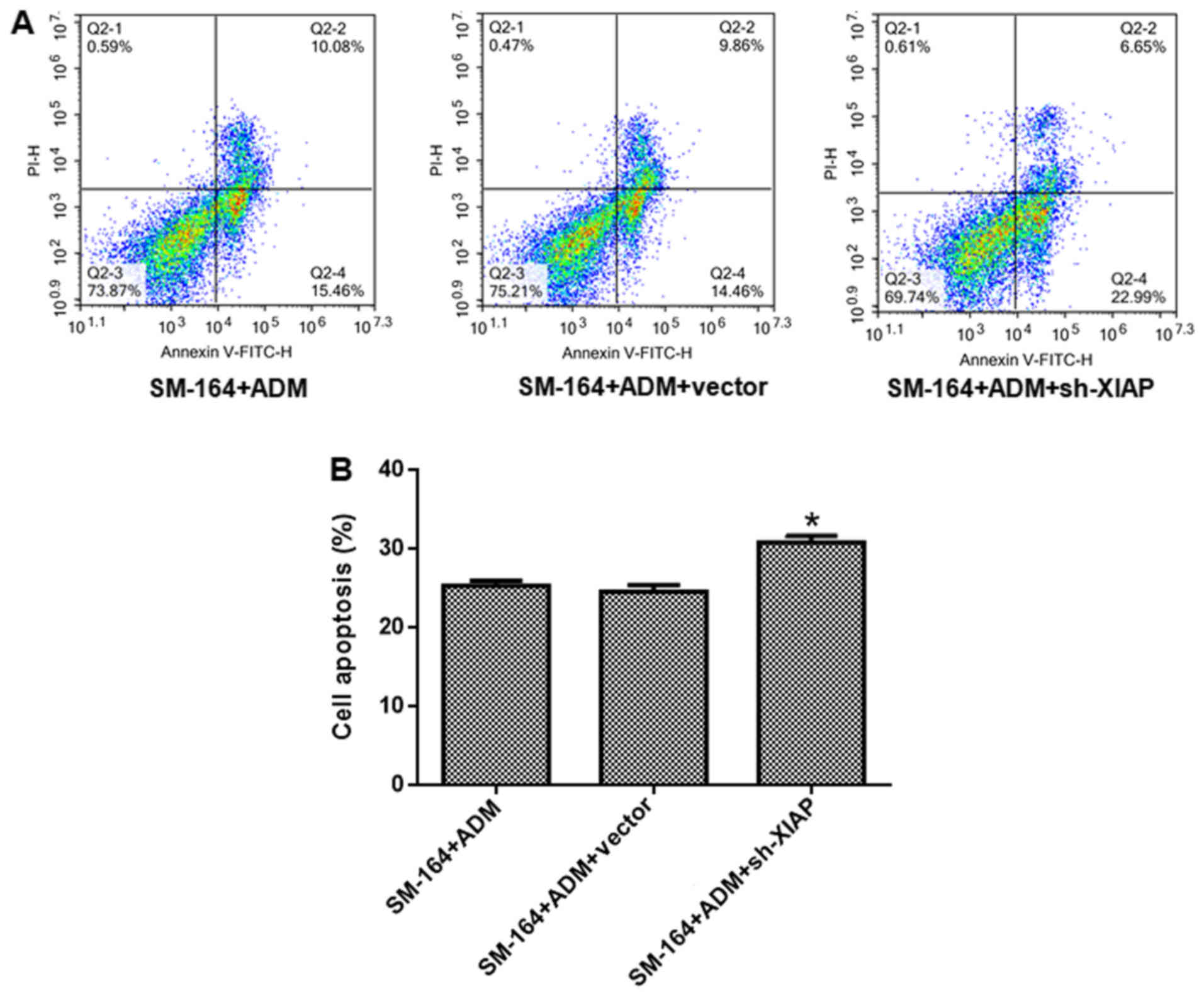
shown in Fig. 6, the number of
apoptotic cells in the ADM + SM-164 + XIAP silencing group
increased significantly; the difference was statistically
significant compared with the ADM + SM-164 group (P<0.05). These
data suggest that silencing of XIAP further suppresses viability
and promotes apoptosis compared with SM-164 treatment.
Effects XIAP silencing on caspases-7,
−9 and −3, PARP, survivin and cIAP-1 expression
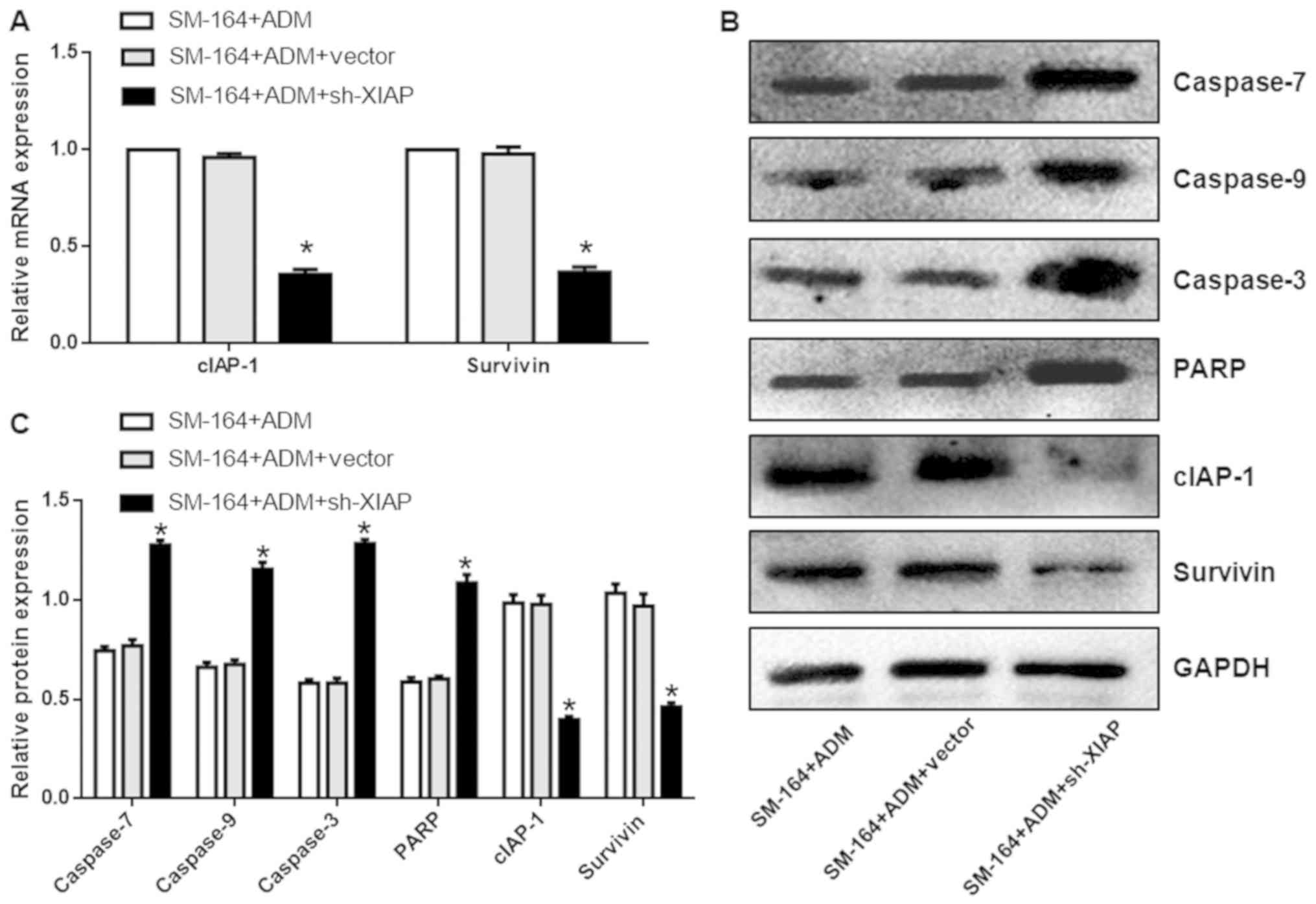
As shown in Fig.
7A, compared with the ADM + SM-164 group, the mRNA expression
levels of survivin and cIAP-1 were significantly lower. In
addition, the protein expression levels of caspases-7, −9, and −3,
and PARP in the ADM + SM-164 + XIAP silencing group were
significantly increased compared with the ADM + SM-164 group; the
expression of cIAP-1 and survivin were also downregulated
(P<0.05; Fig. 7B and C). These
data suggest that silencing of XIAP further promotes
apoptosis-related protein expression compared with SM-164
treatment.
Discussion
Neoadjuvant chemotherapy can induce primary tumor
necrosis, facilitate surgical resection and eliminate
micrometastases (19). The main
anticancer effect of ADM has been reported to involve the
inhibition of topoisomerase II and reduced DNA stability caused by
free radicals (20–22). SM-164 can combine with IAPs,
promote the rapid degradation of cIAP-1, and effectively interfere
with the inhibition caspases-9, −3 and −7 mediated by of XIAP.
SM-164 is an effective inducer of apoptosis in tumor cells and
xenotransplanted tissues (9). The
results of the present study revealed that SM-164 and ADM
suppressed the viability of U2-OS cells, with combined SM-164 and
ADM treatment exhibiting a synergistic pharmacological effect. In
addition, we reported SM-164 and ADM to promote the apoptosis of
U2-OS cells, and that combined use of the two drugs further
increased apoptosis.
IAPs mainly inhibit caspases, inactivate apoptosis
pathways, and interfere in cell apoptosis (11). IAPs can inhibit the
caspase-dependent apoptosis pathway by combining with caspases-3,
−7 and −9; IAPs have become ideal target proteins for altering drug
resistance of several key chemotherapeutic drugs (23). ADM can act in concert with survivin
(and other IAP family proteins) in most tumor cells via
downregulation of IAP anti-apoptotic factors; thus, the
mitochondrial pathway of tumor cell apoptosis may be induced
(11). SM-164 is a Smac protein
small molecule mimic. In a variety of tumor cells expressing IAPs,
caspases-9 and −3 are activated to inhibit the apoptosis of tumor
cells (24). SM-164 is able to
enhance the antitumor effects of ADM and reduce toxicity to normal
cells (8). SM-164 can stimulate
Smac protein to initiate apoptotic signaling pathways and further
alleviate the inhibitory effects of IAPs to its effectors
(caspases-3 and −7, and the apoptosis-initiating factor caspase-9),
which mediates a caspase cascade reaction to induce U2-OS cell
apoptosis (25).
Our results revealed that treatment with ADM and
SM-164 alone significantly increased the expression of
apoptosis-associated factors including caspases-3, −7 and −9. The
expression of anti-apoptotic factors, such as XIAP, cIAP and
survivin were also reported to be decreased; combined drug
treatment was observed to be the most effective. This is also
consistent with the results of a recent study (24). XIAP is the most potent caspase
inhibitor in the IAP family of proteins, possessing three BIR
domains at its N-terminal, which can regulate the death-receptor
pathway and mitochondrial pathway-dependent apoptosis (26). BIR domain 3 in XIAP can effectively
inhibit the activity of caspase-9 (27). The association between BIR1 and
BIR2 can selectively inhibit caspases-3 and −7 (28).
Furthermore, the results of the present study
demonstrated that XIAP silencing inhibited cell viability and
increased apoptosis. Our findings also revealed that silencing XIAP
suppressed the expression of cIAP and survivin, yet the expression
of caspases-3, −7 and −9 were upregulated. Therefore, XIAP may
serve a regulatory role in tumor cell apoptosis. ADM and SM-164
were reported to inhibit cell viability and promote apoptosis,
which could possibly occur via XIAP downregulation. These results
suggest the potential application of this combined treatment in
clinical settings; however, further investigation is required. Our
future studies aim to confirm these findings using a XIAP
overexpression vector. Overexpression of XIAP may provide insight
into the combined effects of ADM + SM-164 as a potential treatment
of OS.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81460406 and
81760488).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JC, XuC, XiC and HS performed the experiments and
analyzed the data. DY designed the study and wrote the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang LL: Biology of osteogenic sarcoma.
Cancer J. 11:294–305. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ryu K, Susa M, Choy E, Yang C, Hornicek
FJ, Mankin HJ and Duan Z: Oleanane triterpenoid CDDO-Me induces
apoptosis in multidrug resistant osteosarcoma cells through
inhibition of Stat3 pathway. BMC Cancer. 10:1872010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ramakrishnan V, Painuly U, Kimlinger T,
Haug J, Rajkumar SV and Kumar S: Inhibitor of apoptosis proteins as
therapeutic targets in multiple myeloma. Leukemia. 28:1519–1528.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Du C, Fang M, Li Y, Li L and Wang X: Smac,
a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yao C, Wu S, Li D, Ding H, Wang Z, Yang Y,
Yan S and Gu Z: Co-administration phenoxodiol with doxorubicin
synergistically inhibit the activity of sphingosine kinase-1
(SphK1), a potential oncogene of osteosarcoma, to suppress
osteosarcoma cell growth both in vivo and in vitro. Mol Oncol.
6:392–404. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chatterjee K, Zhang J, Honbo N and
Karliner JS: Doxorubicin cardiomyopathy. Cardiology. 115:155–162.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun H, Nikolovska-Coleska Z, Lu J, Meagher
JL, Yang CY, Qiu S, Tomita Y, Ueda Y, Jiang S, Krajewski K, et al:
Design, synthesis, and characterization of a potent, nonpeptide,
cell-permeable, bivalent Smac mimetic that concurrently targets
both the BIR2 and BIR3 domains in XIAP. J Am Chem Soc.
129:15279–15294. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bai L, McEachern D, Yang CY, Lu J, Sun H
and Wang S: LRIG1 modulates cancer cell sensitivity to Smac
mimetics by regulating TNFα expression and receptor tyrosine kinase
signaling. Cancer Res. 72:1229–1238. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu J, Bai L, Sun H, Nikolovska-Coleska Z,
McEachern D, Qiu S, Miller RS, Yi H, Shangary S, Sun Y, et al:
SM-164: A novel, bivalent Smac mimetic that induces apoptosis and
tumor regression by concurrent removal of the blockade of cIAP-1/2
and XIAP. Cancer Res. 68:9384–9393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Silke J and Meier P: Inhibitor of
apoptosis (IAP) proteins-modulators of cell death and inflammation.
Cold Spring Harb Perspect Biol. 5:2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gowda Saralamma VV, Lee HJ, Raha S, Lee
WS, Kim EH, Lee SJ, Heo JD, Won C, Kang CK and Kim GS: Inhibition
of IAP's and activation of p53 leads to caspase-dependent apoptosis
in gastric cancer cells treated with Scutellarein. Oncotarget.
9:5993–6006. 2017.PubMed/NCBI
|
|
12
|
Werner TA, Nolten I, Dizdar L, Riemer JC,
Schütte SC, Verde PE, Raba K, Schott M, Knoefel WT and Krieg A:
IAPs cause resistance to TRAIL-dependent apoptosis in follicular
thyroid cancer. Endocr Relat Cancer. 25:295–308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tamanini E, Buck IM, Chessari G,
Chiarparin E, Day JEH, Frederickson M, Griffiths-Jones CM, Hearn K,
Heightman TD, Iqbal A, et al: Discovery of a potent
nonpeptidomimetic, small-molecule antagonist of cellular inhibitor
of apoptosis protein 1 (cIAP1) and X-linked inhibitor of apoptosis
protein (XIAP). J Med Chem. 60:4611–4625. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dizdar L, Oesterwind KA, Riemer JC, Werner
TA, Mersch S, Möhlendick B, Schütte SC, Verde PE, Raba K, Topp SA,
et al: Preclinical assessment of survivin and XIAP as prognostic
biomarkers and therapeutic targets in gastroenteropancreatic
neuroendocrine neoplasia. Oncotarget. 8:8369–8382. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu G, Wang X, Wu S and Li Q: Involvement
of activation of PI3K/Akt pathway in the protective effects of
puerarin against MPP+-induced human neuroblastoma SH-SY5Y cell
death. Neurochem Int. 60:400–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu G, Li J, He L, Wang X and Hong X:
MPTP-induced changes in hippocampal synaptic plasticity and memory
are prevented by memantine through the BDNF-TrkB pathway. Br J
Pharmacol. 172:2354–2368. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
LaBarre DD and Lowy RJ: Improvements in
methods for calculating virus titer estimates from TCID50 and
plaque assays. J Virol Methods. 96:107–126. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wittig JC, Bickels J, Priebat D, Jelinek
J, Kellar-Graney K, Shmookler B and Malawer MM: Osteosarcoma: A
multidisciplinary approach to diagnosis and treatment. Am Fam
Physician. 65:1123–1132. 2002.PubMed/NCBI
|
|
20
|
Gewirtz DA: A critical evaluation of the
mechanisms of action proposed for the antitumor effects of the
anthracycline antibiotics adriamycin and daunorubicin. Biochem
Pharmacol. 57:727–741. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jung K and Reszka R: Mitochondria as
subcellular targets for clinically useful anthracyclines. Adv Drug
Deliv Rev. 49:87–105. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hurley LH: DNA and its associated
processes as targets for cancer therapy. Nat Rev Cancer. 2:188–200.
2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pilling AB, Hwang O, Boudreault A, Laurent
A and Hwang C: IAP antagonists enhance apoptotic response to
enzalutamide in castration-resistant prostate cancer cells via
autocrine TNF-α signaling. Prostate. 77:866–877. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang D, Zhao Y, Li AY, Wang S, Wang G and
Sun Y: Smac-mimetic compound SM-164 induces radiosensitization in
breast cancer cells through activation of caspases and induction of
apoptosis. Breast Cancer Res Treat. 133:189–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang J, Yang Z, Fan C, Sun H and Yang D:
SMAC mimetic SM-164 enhanced adriamycin induced apoptosis and cell
cycle arrest in osteosarcoma cell line HOS. Int J Clin Exp Med.
10:2818–2825. 2017.
|
|
26
|
Schwerd T, Pandey S, Yang HT, Bagola K,
Jameson E, Jung J, Lachmann RH, Shah N, Patel SY, Booth C, et al:
Impaired antibacterial autophagy links granulomatous intestinal
inflammation in Niemann-Pick disease type C1 and XIAP deficiency
with NOD2 variants in Crohn's disease. Gut. 66:1060–1073. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shiozaki EN, Chai J, Rigotti DJ, Riedl SJ,
Li P, Srinivasula SM, Alnemri ES, Fairman R and Shi Y: Mechanism of
XIAP-mediated inhibition of caspase-9. Mol Cell. 11:519–527. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin YF, Lai TC, Chang CK, Chen CL, Huang
MS, Yang CJ, Liu HG, Dong JJ, Chou YA, Teng KH, et al: Targeting
the XIAP/caspase-7 complex selectively kills caspase-3-deficient
malignancies. J Clin Invest. 123:3861–3875. 2013. View Article : Google Scholar : PubMed/NCBI
|