Introduction
Human hepatocellular carcinoma (HCC) is one of the
most common types of cancer and the third leading cause of
cancer-associated mortality worldwide (1). The incidence of HCC is highest in
sub-Saharan Africa and East Asia (2). HCC is a multistep process mainly
associated with alcohol, aflatoxin and persistent infection with
hepatitis B virus or hepatitis C virus (3). Curative treatments, including
locoregional ablation, surgical resection and liver
transplantation, are only appropriate for a small number of
patients diagnosed at an early stage; the majority of patients are
diagnosed with HCC at an advanced disease stage, resulting in a
high mortality rate of HCC (4). At
present, sorafenib is the unique drug that has been approved by the
U.S. Food and Drug Administration (FDA) for patients with advanced
HCC (5). However, the majority of
patients are unable to afford the high expense of sorafenib,
therefore, there is an urgent need for novel therapies.
Several dietary flavonoids, abundant in various
fruits and vegetables, are regarded as bioactive components with
particular benefit to the health of the body. Accumulating evidence
has suggested that flavonoids exert effective biological activity,
including anticancer, anti-inflammatory and antiviral effects
against infection, in addition to potential protective activity
against liver damage (6–8). Avicularin
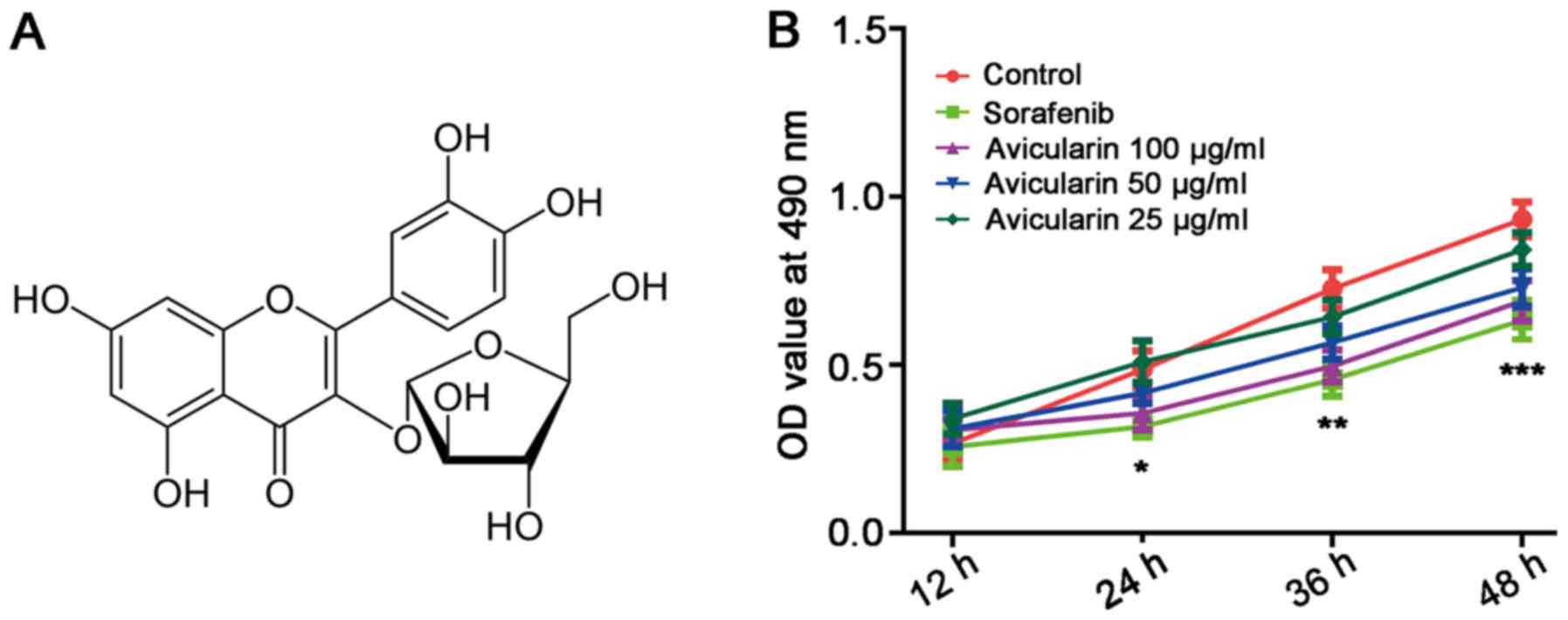
(quercetin-3-α-L-arabinofuranoside; Fig. 1A), a glycoside of quercetin, is a
type of flavonoid which has been reported to inhibit obesity,
inflammation and multidrug resistance (9–11).
In addition, previous studies have indicated that the quercetin
avicularin can induce cytotoxicity in cancer lines and tumor
tissues by activating the intrinsic pathway of apoptosis (12). However, the anticancer activity of
avicularin itself has been not fully elucidated.
The aim of the present study was to investigate the
effects of avicularin on HCC and its potential mechanism of action
in Huh7 cells. It was hypothesized that avicularin has a positive
influence on HCC, and that avicularin may be a potential effective
and safe therapy for HCC.
Materials and methods
Cell culture
The Huh7 HCC cells were obtained from the Cell Bank
of the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). The cells were cultured in Dulbecco's Modified
Eagle Medium supplemented with 10% fetal bovine serum (FBS; both
Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1% antibiotics
(100 U/ml penicillin and 100 mg/ml streptomycin) and 2 mM glutamine
at 37°C in a humidified atmosphere of 5% CO2. The cells
were treated with sorafenib (5 µmol/l; BioVision, Inc.) as the
positive control, and treated with different concentrations (100,
50 and 25 µg/ml) of avicularin (Aladdin Biochemical Technology,
Co., Ltd., Shanghai, China) as the treatment groups.
Cell viability assay
The cell viability assay was performed using a 3-(4,
5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT;
Sigma-Aldrich, Merck KGaA) assay. Briefly, the Huh7 cells were
seeded in 96-well plates at a density of 1×104
cells/well and incubated overnight at 37°C. Following incubation,
cells were treated with sorafenib/avicularin in 10%
FBS-supplemented medium for 12, 24, 36 and 48 h at 37°C,
respectively. The medium was replaced with MTT (0.5 mg/ml) for 4 h
at 37°C. The resulting formazan crystals were then dissolved in
DMSO. The absorbance was determined at 490 nm.
Cell migration assays
The cells were incubated at 5×105 per
dish with sorafenib or avicularin and grown to confluence for 48 h.
Subsequently, a pipette tip was used to generate a wound, vertical
to the parallel line (interval of 2–3 mm) marked on the bottom of
the dish. After 0 and 48 h, the migration distance was analyzed
using Image J software (version 1.46; National Institutes of
Health, Bethesda, MD, USA). The experiments were performed in
triplicate.
Transwell invasion assay
The invasion assay of cells was assessed using
Transwell cell culture chambers (8 µm pore size; EMD Millipore,
Billerica, MA, USA). Briefly, 600 µl medium containing 10% FBS was
added to the lower 24-well chamber. The treated cells were
resuspended in serum-free medium, and then cultured in the upper
chamber coated with 50 µl of 1 g/l Matrigel (BD Biosciences). After
24 h, the upper side of the membrane was gently wiped with a cotton
swab to remove cells whose upper surface did not penetrate, and the
membrane was stained with 1% crystal violet for 20 min at room
temperature, followed by washing with PBS. The invasive cells were
counted at ×200 magnification by an Olympus light microscope (BX43;
Olympus Corporation, Tokyo, Japan) in five randomly selected fields
per well. Each experiment was performed in triplicate.
Reserve transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated with TRIzol (Invitrogen,
Thermo Fisher Scientific, Inc.) and complementary DNA was then
synthesized with PrimeScript™ RT reagent kit (Takara Bio, Inc.,
Otsu, Japan) using the following conditions: Initial incubation at
37°C for 15 min, followed by incubation at 85°C for 5 sec.
Subsequently, qPCR was performed by FastStart Universal SYBR-Green
Master (Roche Diagnostics, Mannheim, Germany) according to the
manufacturer's protocols. The qPCR thermocycling conditions were as
follows: Initial denaturation at 95°C for 10 min, followed by 40
cycles of 95°C for 10 sec and 60°C for 60 sec. The relative mRNA
expression was calculated using the 2−∆∆Cq method
(13), and normalized using GAPDH
as the reference gene. The following primers were used for the
experiment: Nuclear factor (NF)-κB (p65) forward,
5′-ACGATCTGTTTCCCCTCATCT-3′ and reverse,
5′-TGCTTCTCTCCCCAGGAATA-3′; peroxisome proliferator-activated
receptor γ (PPAR-γ) forward, 5′-AACTGCAGGGTGAAACTCTGGGAGATTCTCC-3′
and reverse, 5′-GGATTCAGCAACCATTGGGTCAGCTCT-3′; cyclooxygenase-2
(COX-2), forward, 5′-CTGAGGGGTTACCATTCCA-3′ and reverse,
5′-TGAGCAAGTCCGTGTTCAAG-3′; GAPDH forward,
5′-GTGAGGAGGGGAGATTCAG-3′ and reverse,
5′-GCATCCTGGGCTACACTG-3′.
Western blot analysis
The cells were lysed with RIPA buffer (50 mM
Tris-HCl, pH 7.4; 0.15 M NaCl; 1% Nonidet P-40, 0.1% sodium dodecyl
sulfate and 0.5% sodium deoxycholate) containing protease and
phosphatase inhibitors (Sigma-Aldrich, Merck KGaA), and the protein
concentration was measured using a BCA protein assay kit (Pierce,
Thermo Fisher Scientific, Inc.). The same amount (20 µg) of
proteins were separated on 10% SDS-PAGE gels and transferred onto
Immobilon polyvinylidene difluoride membranes (EMD Millipore), and
then blocked in 5% skim milk in TBST for 1 h at room temperature.
The primary antibodies against COX-2 (1:500; cat. no. ab23672),
PPAR-γ (1:500; cat. no. ab45036), NF-κB (p65) (1:2,000; cat. no.
ab86299) and GAPDH (1:2,500; cat. no. ab9485; Abcam, Cambridge, UK)
were added for overnight incubation at 4°C. Following washing three
times with 1X TBST buffer, a secondary antibody coupled to
horseradish peroxidase (1:2,000; cat. no. sc-2030) from Santa Cruz
Biotechnology, Inc. was incubated with the membranes for 1.5 h at
room temperature, and immunoactivity was detected using an ECL-Plus
kit (GE Healthcare Life Sciences, Pollards Wood, UK). Band
intensity on scanned films was quantified using ImageJ software
(version 1.46; National Institutes of Health, Bethesda, MD, USA)
and expressed as relative intensity compared with control. The
intensity of the protein bands was normalized to GAPDH and the
relative intensity was calculated.
Flow cytometric analysis
Following treatment for 48 h, the cells were
harvested and washed in cold PBS. For cell cycle analysis, the
cells were fixed in 70% ethanol at −20°C overnight. Following
washing and centrifugation at 1,000 × g for 5 min at 4°C, the cells
were treated with RNase A and stained with propidium iodide (PI)
working solution for 30 min in the dark. Analysis was determined
using a FACS Caliber flow cytometer and CellQuest software (version
5.1; BD Biosciences). For the analysis of cell apoptosis, the
Annexin-V-FITC Apoptosis Detection kit I (BD Pharmingen) was used,
according to the manufacturer's protocol. Briefly, the cells were
resuspended with 1X binding buffer, following which the cells were
stained with 5 µl Annexin-V-FITC and 5 µl PI, and then incubated in
the dark for 15 min. Subsequently, the appropriate 1X binding
buffer was added and apoptosis was analyzed by the FACSCalibur flow
cytometer.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA). The results
are expressed as the mean ± standard deviation from at least three
independent experiments. Statistical significance was analyzed
using a two-tailed Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Avicularin inhibits the proliferation
of Huh7 cells
The physiological effect of avicularin on HCC was
investigated by observing the proliferation of Huh7 cells treated
with avicularin. As shown in Fig.
1B, sorafenib, an oral multikinase inhibitor approved for
advanced HCC (5), resulted in
marked inhibition of cell proliferation. Avicularin significantly
inhibited cell proliferation in a concentration-dependent manner.
With the growth of Huh7 cells cultured with avicularin, the
suppressing effect of avicularin on cell proliferation was more
marked.
Avicularin repress the migration and
invasion of Huh7 cells
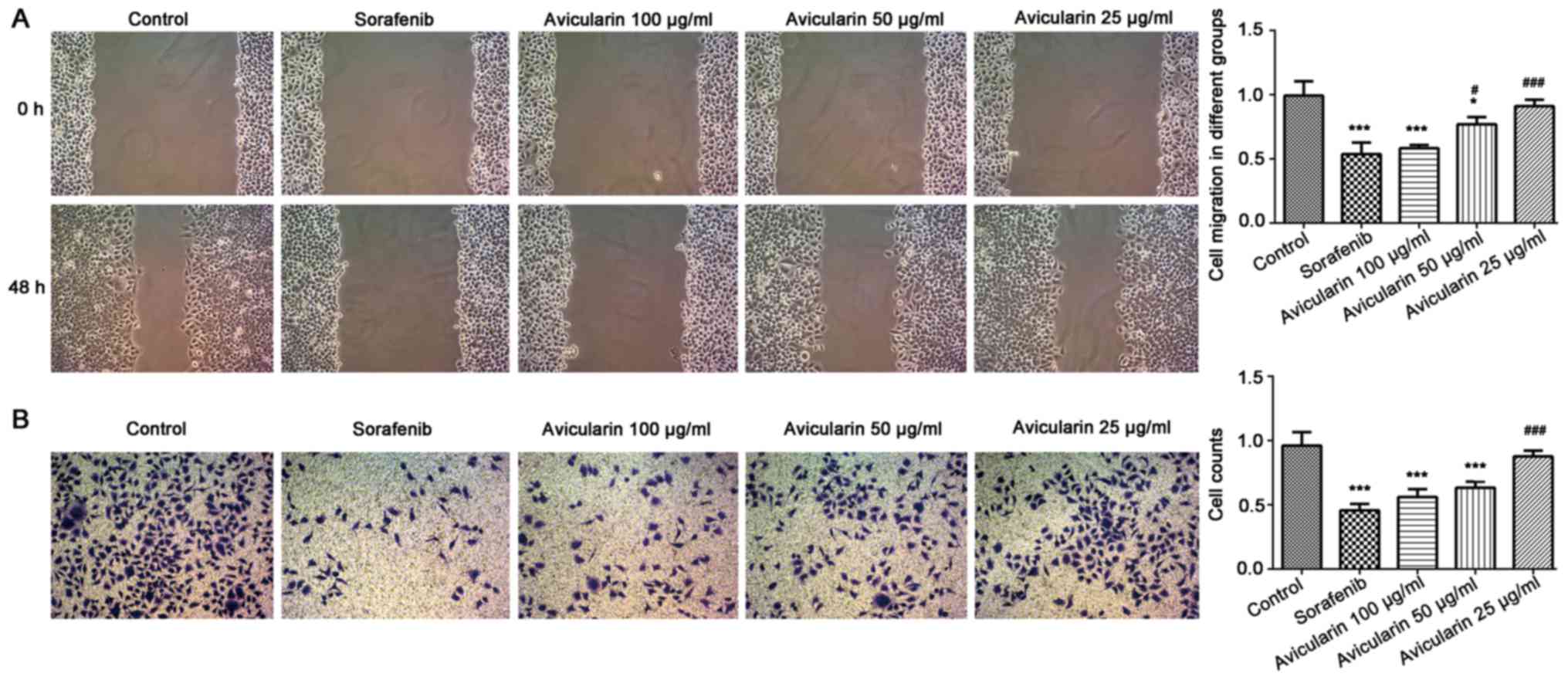
To confirm that avicularin functions as tumor
suppressor in HCC, the present study investigated the influence of
avicularin on the migration and invasion of Huh cells. The results
suggested that the wounding healing of Huh7 cells was markedly
retarded in the presence of sorafenib and avicularin (Fig. 2A). Therefore avicularin suppressed
cell migration following treatment at concentrations of 100 and 50
µg/ml, and was concentration-dependent. Additionally, the invasive
ability of Huh7 cells cultured under sorafenib/avicularin was
observed through a Transwell assay. As shown in Fig. 2B, compared with the control cells,
the cell counts were markedly decreased following sorafenib and
avicularin treatment. Avicularin resulted in a marked inhibition of
cell invasion at the concentrations of 100 and 50 µg/ml. As a
result, avicularin effectively suppressed the migration and
invasion of Huh7 cells in HCC.
Avicularin promotes cell apoptosis and
regulates cell cycle progression
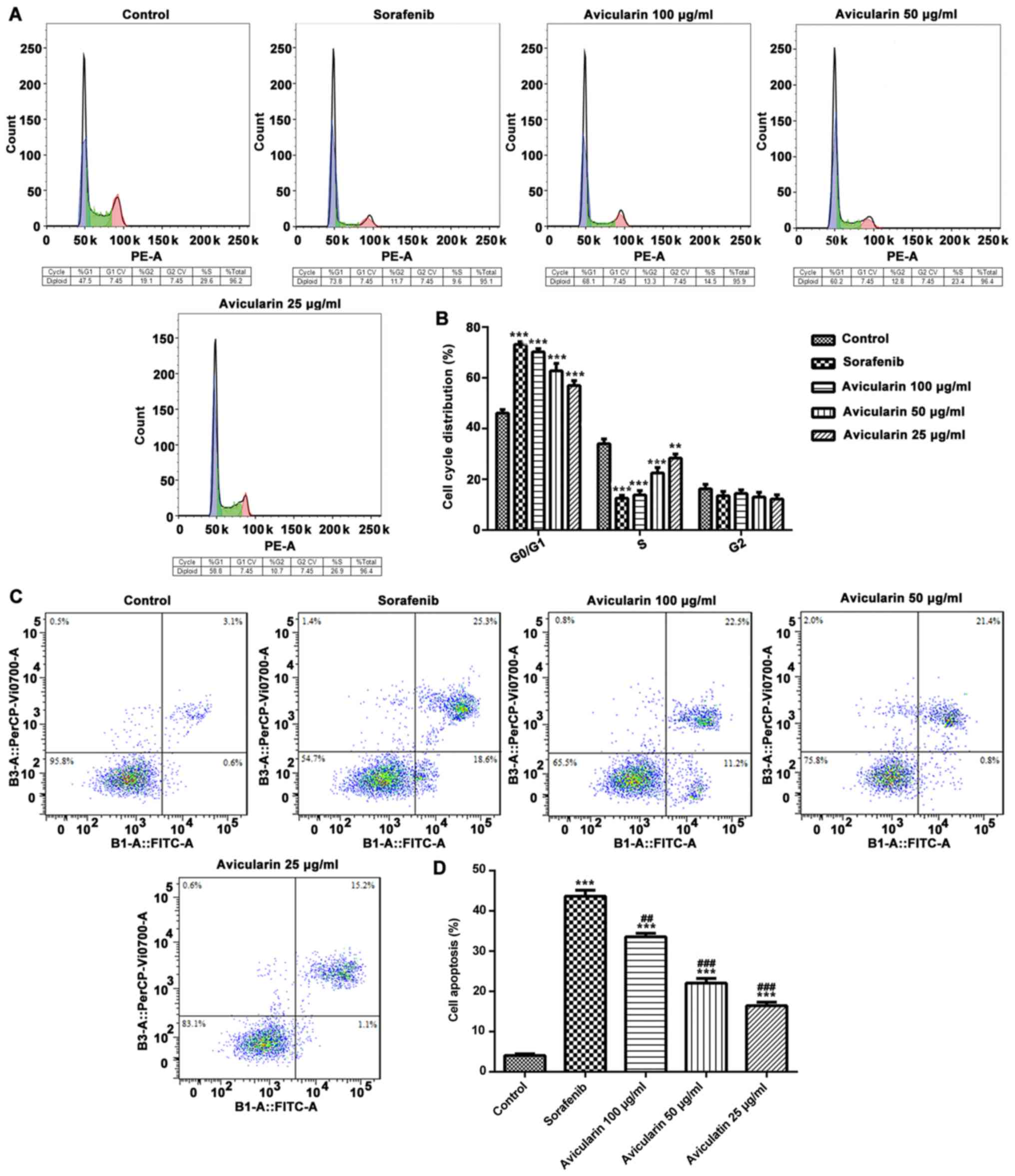
To determine the effects of avicularin on cell
cycle, the cells were treated with different concentrations (25, 50
and 100 µg/ml) of avicularin for 48 h, and the percentage of cells
in different cell cycle phases were analyzed. As shown in Fig. 3A and B, compared with the control
cells, sorafenib and avicularin induced notable G0/G1 arrest in the
Huh7 cells. Additionally, there was a marked decrease in the
accumulation of S-phase cells in the avicularin-treated group,
similar to sorafenib, particularly at the concentration of 100
µg/ml. These data suggested that the inhibition of cell
proliferation by avicularin was connected with an increase in the
G0/G1 phase and a decrease in the S phase of the cell cycle.
In order to assess whether avicularin inhibited cell
proliferation through an apoptotic mechanism in HCC, flow
cytometric analysis was performed in Huh7 cells. As shown in
Fig. 3C and D, the percentage of
cell apoptosis was increased in the sorafenib-treated cells. It was
confirmed that avicularin enhanced cell apoptosis, which occurred
in a concentration-dependent manner. The above results demonstrated
that avicularin regulates cell cycle progression and promotes cell
apoptosis, which may lead to the inhibition of cell proliferation
in HCC.
Avicularin serves an antineoplastic
role through the regulation of NF-κB (p65), COX-2 and PPAR-γ
Subsequently, to gain further insight into the
mechanism underlying the antiproliferative activity of avicularin
in HCC, the present study aimed to determine whether
avicularin-induced cell apoptosis and cell cycle arrest involves
NF-κB (p65), COX-2 and PPAR-γ, which have been known to be involved
in cell cycle progression and proliferation. As shown in Fig. 4A and B, sorafenib resulted in
significant inhibition of NF-κB (p65) and COX-2, not only at the
mRNA level, but also at the protein level. The mRNA level and
protein expression levels of PPAR-γ were increased. Similarly,
avicularin resulted in the same effects on NF-κB (p65), COX-2 and
PPAR-γ in a concentration-dependent manner. Taken together, these
results suggest that avicularin regulates cell apoptosis and cell
cycle via the downregulation of NF-κB (p65) and COX-2 and the
upregulation of PPAR-γ.
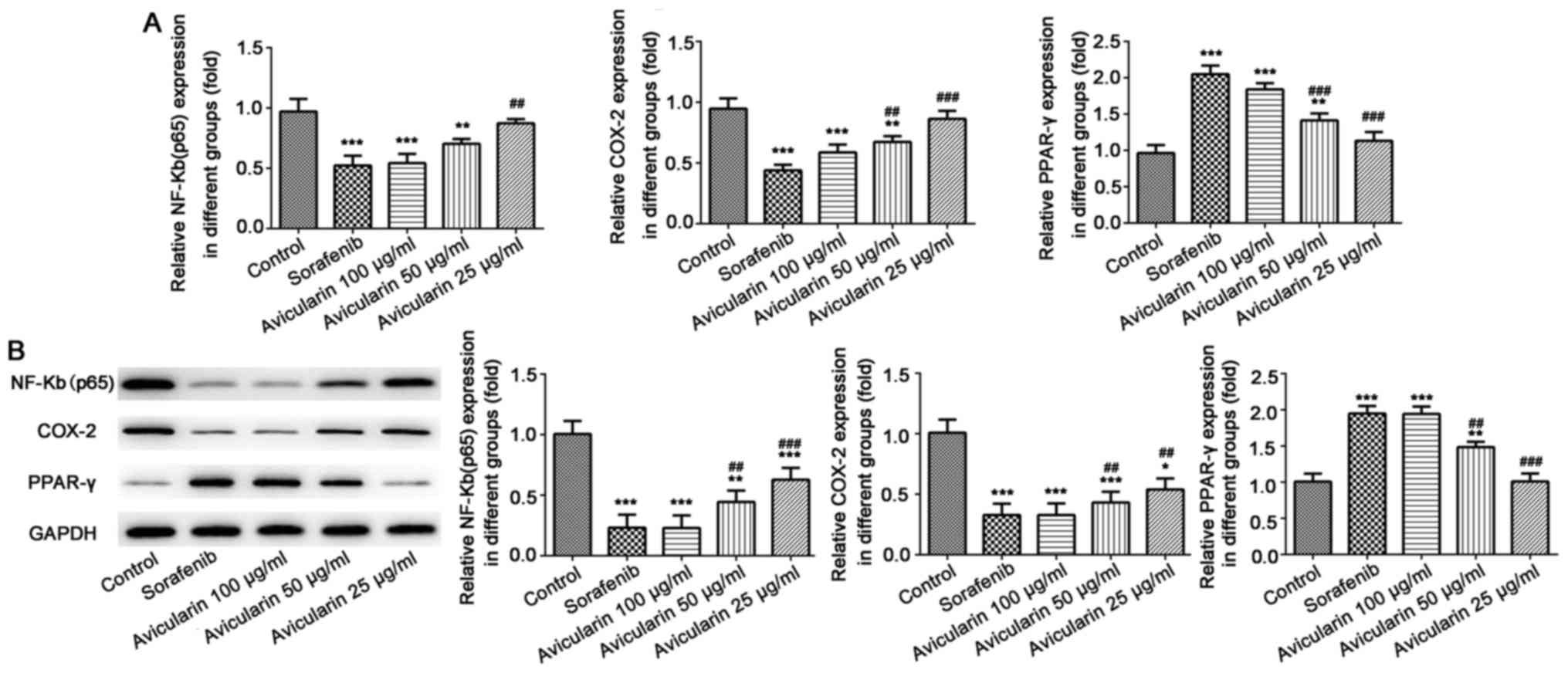 | Figure 4.Expression of NF-κB (p65), COX-2 and
PPAR-γ. (A) Following treatment with sorafenib and avicularin, the
mRNA levels of NF-κB (p65), COX-2 and PPAR-γ was measured by
reverse transcription-quantitative polymerase chain reaction
analysis. (B) Protein expression of NF-κB (p65), COX-2 and PPAR-γ
was measured by western blot analysis and quantified. Data is
presented as the mean ± standard deviation of three experiments.
*P<0.05, **P<0.01 and ***P<0.001, vs. control;
##P<0.01 and ###P<0.001, vs. sorafenib.
NF-κB, nuclear factor-κB; COX-2, cyclooxygenase-2; PPARγ,
peroxisome proliferator-activated receptor γ. |
Discussion
HCC is a global public health problem that accounts
for a high rate of morality. Currently, liver resection, liver
transplantation and ablation remain the mainstream treatment, which
are only appropriate for those patients at an early stage and can
easily result in adverse effects. Therefore, to overcome these
deficiencies, sorafenib, approved by the FDA, has been shown to be
effective against several solid tumors and is a standard treatment
for HCC (14). Previous studies
have reported that sorafenib exerts its antitumor activities by
triggering cell apoptosis and inhibiting tumor angiogenesis through
the Raf, c-kit, vascular endothelial growth factor,
platelet-derived growth factor, fms-like tyrosine kinase 3 and
mitogen-activated protein kinase pathways (5,15).
In addition, it is reported that sorafenib can reduce Alzheimer's
disease pathology by reducing neuroinflammation through inhibition
of the expression of NF-κB and COX-2 (16). In the present study, it was found
that sorafenib also targets NF-κB and COX-2 to exert its
antineoplastic role. However, as the high cost of sorafenib makes
is difficult to afford, it remains essential to develop more
effective, safe and appropriate methods for the treatment of HCC.
In the present study, a novel therapy against HCC was investigated
and its mechanism of anticancer activity was examined. It was found
that avicularin inhibited the proliferation, migration and invasion
of Huh7 cells in HCC through influencing cell apoptosis and cycle
distribution on the regulation of NF-κB (p65), COX-2 and
PPAR-γ.
In oriental culture, numerous plants have been used
in folk medicine for thousands of years. Among abundant plants, the
flavonoid family, as a group of plant secondary metabolites with
multiple phenolic structures, is of particular interest and is
usually found in vegetables and fruits (17). It has been reported that numerous
flavonoids can enhance the efficacy and alleviate the adverse
effects of tumor therapies. Baicalein and silymarin, extracts of
Scutellaria baicalensis, have been shown to exert a
suppressive effect on tumors. The combination of baicalein and
silymarin can suppress HepG2 cell proliferation by increasing the
percentages of cells in the G0/G1 phase and decreasing those in the
S phase (18). Amentoflavone, a
flavonoid compound extracted from Selaginella tamariscina
Spring, reportedly exerts antineoplastic activity via the induction
of cell apoptosis and inhibition of glycolysis in HCC (19). Additionally, baicalein (20), tectorigenin (21) and other flavonoids have been shown
to protect cells against cancer progression though the activation
of proapoptotic and antiproliferative pathways or other pathways
(22). Avicularin
(quercetin-3-α-L-arabinofuranoside) belongs to a group of flavonoid
glycosides. It has been reported that avicularin has a protective
effect against human gastric cancer through inducing apoptosis
dependent on Bax and BCL-2-related ovarian killer (11). The present study focused on the
efficacy of avicularin on HCC. The data showed that avicularin
exerted marked anticancer activity in Huh7 cells in a
concentration-dependent manner. Avicularin at 100 µg/ml had a
marked suppressive effect on cell proliferation, migration and
invasion, similar to sorafenib. However, at a concentration at 25
µg/ml, avicularin had no effect on HCC.
NF-κB is a protein complex that controls the
transcription of DNA (23). It is
a key transcription factor that is closely associated with the
proliferation and apoptosis of cancer, and it also serves an
important role in the cell cycle, which is vital in determining the
degree of cellular proliferation and apoptosis. In the present
study, the cell population was increased in the G0/G1 phase but
decreased in the S phase. Cell apoptosis was improved following
treatment with avicularin. The mRNA level and protein expression
levels of p65 (a subunit of NF-κB) were significantly inhibited by
avicularin, which indicates that avicularin suppressed cell cycle
progression and promoted cell apoptosis via the inhibition of NF-κB
activity. The involvement of COX-2 in tumorigenesis in HCC has been
widely reported in several studies, and is also closely linked with
NF-κB. The promoter regions of the COX-2 gene in human and mice
harbor binding sites for NF-κB. Therefore the expression of COX-2
can be mediated via NF-κB (24).
Accordingly, the results of the present study showed that
avicularin markedly downregulated the mRNA and protein expression
levels of COX-2 in Huh7 cells. Therefore, it is possible that
avicularin exerts its anticancer activity on the inhibition of
COX-2 through NF-κB.
PPAR-γ is a member of the nuclear hormone receptor
superfamily. It is involved in the control of biological processes
associated with differentiation, growth, apoptosis and cell cycle
(25). The activity of PPAR-γ has
been shown to be inhibited in HCC (26). The overexpression of PPAR-γ can
inhibit cell proliferation in HCC, and the downregulation of PPAR-γ
has been associated with differentiation and poor prognosis in
patients in HCC (27). PPAR-γ is
also reported to be correlated with cell cycle arrest through p53
and p21 (28). Additionally,
PPAR-γ promotes cell apoptosis, which resulted in the inhibition of
cancer (29). Based on these
results, the present data showed that the expression of PPAR-γ was
significantly increased when the cells were treated with sorafenib
and avicularin. The anticancer activity of avicularin involved in
the inhibition of cell proliferation, migration and invasion, and
changes in cell apoptosis and cell cycle may partly depend on the
upregulation of PPAR-γ.
In conclusion, the results demonstrate an
antineoplastic role of avicularin of HCC in vitro. It was
clearly demonstrated that avicularin can inhibit cell
proliferation, migration and invasion in HCC through inducing
apoptosis and suppressing cell cycle progression. Additionally, the
decreased activity of NF-κB and COX-2 and increased activity of
PPAR-γ suggest that avicularin has an antineoplastic effect through
the regulation of these, and that NF-κB, COX-2 and PPAR-γ are vital
factors predicting the anticancer effect in HCC. Overall, the
results of the present study suggests that avicularin may be a
valuable therapeutic agent in the treatment of HCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Health and
Family Planning Commission of Zhejiang Province (grant no.
GZS2012031)
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JS was involved in the design of the experiment. ZM,
FL and YQ performed the experiments and analyzed the data. ZM and
YQ wrote the manuscript. JS reviewed and edited the manuscript. All
authors read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lu T, Seto WK, Zhu RX, Lai CL and Yuen MF:
Prevention of hepatocellular carcinoma in chronic viral hepatitis B
and C infection. World J Gastroenterol. 19:8887–8894. 2003.
View Article : Google Scholar
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang JD and Roberts LR: Hepatocellular
carcinoma: A global view. Nat Rev Gastroenterol Hepatol. 7:448–458.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang X, Feng K, Zhang Y, Li Z, Zhou F,
Dou H and Wang T: Sorafenib and DE605, a novel c-Met inhibitor,
synergistically suppress hepatocellular carcinoma. Oncotarget.
6:12340–12356. 2015.PubMed/NCBI
|
|
6
|
Cui X, Wang Y, Kokudo N, Fang D and Tang
W: Traditional Chinese medicine and related active compounds
against hepatitis B virus infection. Biosci Trends. 4:39–47.
2010.PubMed/NCBI
|
|
7
|
Williams RJ, Spencer JP and Rice-Evans C:
Flavonoids: Antioxidants or signalling molecules? Free Radic Biol
Med. 36:838–849. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Handoussa H, Osmanova N, Ayoub N and
Mahran L: Spicatic acid: A 4-carboxygentisic acid from Gentiana
spicata extract with potential hepatoprotective activity. Drug
Discov Ther. 3:278–286. 2009.PubMed/NCBI
|
|
9
|
Fujimori K and Shibano M: Avicularin, a
plant flavonoid, suppresses lipid accumulation through repression
of C/EBPα-activated GLUT4-mediated glucose uptake in 3T3-L1 cells.
J Agric Food Chem. 61:5139–5147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vo VA, Lee JW, Chang JE, Kim JY, Kim NH,
Lee HJ, Kim SS, Chun W and Kwon YS: Avicularin inhibits
lipopolysaccharide-induced inflammatory response by suppressing ERK
phosphorylation in RAW 264.7 macrophages. Biomol Ther (Seoul).
20:532–537. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo XF, Liu JP, Ma SQ, Zhang P and Sun WD:
Avicularin reversed multidrug-resistance in human gastric cancer
through enhancing Bax and BOK expressions. Biomed Pharmacother.
103:67–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Srivastava S, Somasagara RR, Hegde M,
Nishana M, Tadi SK, Srivastava M, Choudhary B and Raghavan SC:
Quercetin, a natural flavonoid interacts with DNA, Arrests cell
cycle and causes tumor regression by activating mitochondrial
pathway of apoptosis. Sci Rep. 6:240492016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lang L: FDA approves sorafenib for
patients with inoperable liver cancer. Gastroenterology.
134:3792008. View Article : Google Scholar
|
|
15
|
Hahn O and Stadler W: Sorafenib. Curr Opin
Oncol. 18:615–621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Echeverria V, Burgess S, Gamble-George J,
Zeitlin R, Lin X, Cao C and Arendash GW: Sorafenib inhibits nuclear
factor kappa B, decreases inducible nitric oxide synthase and
cyclooxygenase-2 expression, and restores working memory in APPswe
mice. Neuroscience. 162:1220–1231. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nijveldt RJ, van Nood E, van Hoorn DE,
Boelens PG, van Norren K and van Leeuwen PA: Flavonoids: A review
of probable mechanisms of action and potential applications. Am J
Clin Nutr. 74:418–425. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen CH, Huang TS, Wong CH, Hong CL, Tsai
YH, Liang CC, Lu FJ and Chang WH: Synergistic anti-cancer effect of
baicalein and silymarin on human hepatoma HepG2 cells. Food Chem
Toxicol. 47:638–644. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu B and Yu S: Amentoflavone suppresses
hepatocellular carcinoma by repressing hexokinase 2 expression
through inhibiting JAK2/STAT3 signaling. Biomed Pharmacother.
107:243–253. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang RR, Zhang S, Qi JA, Wang ZD, Li J,
Liu PJ, Huang C, Le XF, Yang J and Li ZF: Preferential inhibition
of hepatocellular carcinoma by the flavonoid Baicalein through
blocking MEK-ERK signaling. Int J Oncol. 41:969–978. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang CP, Ding H, Shi DH, Wang YR, Li EG
and Wu JH: Pro-apoptotic effects of tectorigenin on human
hepatocellular carcinoma HepG2 cells. World J Gastroenterol.
18:1753–1764. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garcia ER, Gutierrez EA, de Melo FCSA,
Novaes RD and Goncalves RV: Flavonoids effects on hepatocellular
carcinoma in murine models: A systematic review. Evid Based
Complement Alternat Med. 2018:63289702018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gilmore TD: Introduction to NF-kappaB:
Players, pathways, perspectives. Oncogene. 25:6680–6684. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim JH, Na HK, Pak YK, Lee YS, Lee SJ,
Moon A and Surh YJ: Roles of ERK and p38 mitogen-activated protein
kinases in phorbol ester-induced NF-kappaB activation and COX-2
expression in human breast epithelial cells. Chem Biol Interact.
171:133–141. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koeffler HP: Peroxisome
proliferator-activated receptor gamma and cancers. Clin Cancer Res.
9:1–9. 2003.PubMed/NCBI
|
|
26
|
Vara D, Morell C, Rodriguez-Henche N and
Diaz-Laviada I: Involvement of PPARgamma in the antitumoral action
of cannabinoids on hepatocellular carcinoma. Cell Death Dis.
4:e6182013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li S, Li J, Fei BY, Shao D, Pan Y, Mo ZH,
Sun BZ, Zhang D, Zheng X, Zhang M, et al: MiR-27a promotes
hepatocellular carcinoma cell proliferation through suppression of
its target gene peroxisome proliferator-activated receptor γ. Chin
Med J (Engl). 128:941–947. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Galli A, Ceni E, Mello T, Polvani S,
Tarocchi M, Buccoliero F, Lisi F, Cioni L, Ottanelli B, Foresta V,
et al: Thiazolidinediones inhibit hepatocarcinogenesis in hepatitis
B virus-transgenic mice by peroxisome proliferator-activated
receptor gamma-independent regulation of nucleophosmin. Hepatology.
52:493–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu J, Shen B, Chu ES, Teoh N, Cheung KF,
Wu CW, Wang S, Lam CN, Feng H, Zhao J, et al: Inhibitory role of
peroxisome proliferator-activated receptor gamma in
hepatocarcinogenesis in mice and in vitro. Hepatology.
51:2008–2019. 2010. View Article : Google Scholar : PubMed/NCBI
|