Introduction
The cytokine interleukin (IL)-17A, produced by T
helper cells (Th17), CD8-positive T cells, neutrophils, γδT cells,
and natural killer cells, plays an important role in host pathogen
defense by activating the inflammatory response, which recruits
neutrophils to fight infection (1). Notably, the chronic inflammatory
microenvironment established by IL-17A and other inflammatory
cytokines may facilitate chronic diseases in the liver, including
non-alcoholic steatohepatitis (2),
and hepatocellular carcinoma (HCC) initiation and progression
(3,4). For example, IL-17A knockout
significantly reduced tumor incidence from 65 to 20% in diethyl
nitrosamine-induced HCC models (5). In addition to the role of tumor
initiation, IL-17A may also have an impact on tumor proliferation,
angiogenesis, metastasis and chemoresistance (6). IL-17A activates STAT3 and NF-κB
signaling pathways, regulating the expression of a panel of genes
and aggravating tumor malignancy (7). IL-17A not only promotes tumor growth
in animal models (5,8), but also predicts poor prognosis in a
large portion of clinical reports, including gastric, lung and
breast cancers, and HCC (9–12).
We have previously reported that elevated serum IL-17A levels were
positively correlated with a larger tumor size and an increased
risk of early recurrence of HCC after curative hepatectomy
(13). The number of intratumoral
Th17 cells positively correlates with microvessel density and poor
survival, suggesting that IL-17A may stimulate tumor growth by
promoting angiogenesis (14).
While IL-17A does not have a direct role on
endothelial cell proliferation, it may stimulate the production and
secretion of pro-angiogenic factors from both tumor cells and
fibroblasts in some tumor types (15). IL-17A stimulates vascular
endothelial growth factor A (VEGFA) production and promotes
angiogenesis in non-small-cell lung cancer (16), gastric cancer (17) and colorectal cancer (18). In addition to VEGFA, some CXC
chemokines have been shown to play important roles in promoting
tumoral angiogenesis via the receptor C-X-C chemokine receptor type
2 (CXCR2). CXCR2 is a 7-transmembrane G protein-coupled receptor
that mediates chemotaxis during an immune response. Upregulation of
CXCR2 has been reported in inflammatory diseases including
rheumatoid arthritis, psoriasis and atherosclerosis (19). In addition to expression on a
subset of leukocytes, CXCR2 is also present on microvascular
endothelial cells, and has been shown to be involved in endothelial
cell chemotaxis and angiogenesis (20). The CXC chemokines with a glutamic
acid-leucine-arginine (ELR) motif such as CXCL1/2/3/5/6/8 can bind
to CXCR2 and are pro-angiogenic (21). The expression of IL-8 (CXCL8) is
correlated with vascularity in gastric cancers (22), and CXCL1/3/5/8 augments
angiogenesis and tumorigenesis in renal cell carcinoma (23). IL-8 secreted from activated
stellate cells also promotes angiogenesis in HCC (24). In particular, IL-17A has been shown
to promote the CXCR2-dependent angiogenesis and tumor growth of
non-small cell lung cancer, while it has no effect on VEGFA
secretion (25).
Since the IL-17A receptor is extensively expressed
in liver cell types including liver cancer cells, it is of interest
to determine the direct role of IL-17A in these cells. Previous
studies have suggested that IL-17A may influence HCC cell survival
and migration (26,27). However, it remains unclear whether
IL-17A can regulate the production of VEGFA or other pro-angiogenic
factors directly from liver cancer cells, thereby contributing to
angiogenesis. The present study examined the effect of IL-17A on
Huh7.5 and HepG2 cells. The results indicated that IL-17A
stimulated the secretion of angiogenic CXC chemokines and promoted
CXCR2-dependent endothelial chemotaxis.
Materials and methods
Cell cultures
The liver cancer cell line HepG2 (28,29)
was supplied by American Type Culture Collection and Huh7.5
(30) cells were gifted by Dr.
Aihua Zheng (Chinese Academy of Sciences). The endothelial cell
HUVEC was a gift from Dr. Peigang Wang (Capital Medical
University). The cell lines were authenticated by short tandem
repeat profiling (Beijing Center for Physical and Chemical
Analysis). These cells were cultured in Dulbecco's modified Eagle's
medium (DMEM; high glucose; Hyclone; GE Healthcare Life Sciences)
supplemented with 10% fetal bovine serum (FBS; Biological
Industries) and 105 U/l penicillin and 100 mg/l
streptomycin (Hyclone; GE Healthcare Life Sciences). When
indicated, 100 ng/ml recombinant IL-17A (R&D Systems, Inc.) was
added to the culture media.
For IL-17A or enhanced green fluorescent protein
(EGFP) overexpressing cells, pcDH-EF1-IL17A-IRES-puromycin or
pcDH-EF1-EGFP-IRES-puromycin were constructed using a pcDH vector
(System Biosciences). Construction was performed by GenScript and
was sequenced to be correct. For pseudovirus production, lentiviral
vector, pLP1, pLP2 and pVSVG (System Biosciences) were
co-transfected into 293T cells using X-tremeGENE HP (Roche
Diagnostics). After 12 h, the media was removed and fresh media was
added. The supernatants were collected at 48 h post-transfection
and filtered through 0.45 µm filters to remove the cell debris.
HepG2 or Huh7.5 cells were cultured with the pseudovirus-containing
media with polybrene (8 µg/ml; Sigma Aldrich; Merck KGaA) for 2
days and the stably transduced cells were selected using 5 µg/ml
puromycin (Shanghai Qcbio Science & Technologies Co.,
Ltd.).
ELISA
Cells (1×105) were cultured in 6-well
plates for 24 h. The cell-free supernatant was collected and stored
at −70°C. The concentrations of IL-17A, VEGFA, CXCL10 and CXCL2
were measured using commercially available ELISA kits for IL-17A
(cat. no. CHE0054; Beijing 4A Biotech Co., Ltd.), VEGFA (cat. no.
SEA143Hu; Cloud-Clone Corp.), CXCL10 (cat. no. SEA371Hu;
Cloud-Clone Corp.) and CXCL2 (cat. no. SEB603Hu; Cloud-Clone
Corp.).
Cell proliferation assay
Cell proliferation was measured using a Cell
Counting Kit-8 (CCK-8) kit (Dojindo Molecular Technologies, Inc.).
In this assay, cells were cultured in 96-well plates
(2×103 cells/well) for 0, 1, 3, 5 or 7 days. CCK-8 (10
µl) was added to each well and the cells were cultured for 2 h at
37°C. Then, the absorbance was measured at 450 nm using a
microplate reader (Thermo Electron Corporation). All tests were
performed in triplicate.
RNA extraction and reverse
transcription-quantitative PCR
Total RNA was extracted using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. cDNA was synthesized from 1 µg total RNA using
FastKing RT kits (Tiangen Biotech Co., Ltd.), according to the
manufacturer's instructions. Briefly, genomic DNA was removed using
gDNA buffer at 42°C for 3 min and then reverse transcription was
performed at 42°C for 15 min followed by 95°C for 3 min. RT-qPCR
was performed on an ABI 7500 Real-Time PCR Systems (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using SYBR-Green
(Toyobo Life Sciences). The PCR thermocycling conditions were as
follows: 50°C incubation for 2 min, 95°C initial denaturation for
10 min, then 40 cycles of 95°C for 15 sec, 60°C for 30 sec and 72°C
for 30 sec. Fluorescence data were collected in the extension step
(72°C for 30 sec). All reactions were run in triplicate. The
housekeeping gene GAPDH served as an internal control. The
2−ΔΔCq method was used to determine expression levels
(31). The primer sequences are as
follows: CXCL1 forward (F), TCAATCCTGCATCCCCCATAGTTA and reverse,
(R) GTGAGCTTCCTCCTCCCTTCT; CXCL2 F, CGCATCGCCCATGGTTA and R,
ACAGCCACCAATAAGCTTCC; CXCL3 F, GCTTGTCTCAACCCCGCA and R,
CACCCTGCAGGAAGTGTCAATG; CXCL4 F, ATCGCACTGAGCACTGAGATCC and R,
GACCTGGGAGGTGGTCTTCAC; CXCL5 F, TTTACAGACCACGCAAGGAGT and R,
TTTCCTTGTTTCCACCGTCCA; CXCL6 F, AGCAAGTTTGTCTGGACCCG and R,
CAGAAAACTGCTCCGCTGAA; CXCL7 F, GGAAAGGAACCCATTGCAACC and R,
TCTGGGTCCAGGCAGATTTT; CXCL8 F, TCCTGATTTCTGCAGCTCTGT and R,
CCAGACAGAGCTCTCTTCCA; CXCL9 F, AAGCCCTTCCTGCGAGAAAA and R,
TTCACATCTGCTGAATCTGGGT; CXCL10 F, AAGCCAATTTTGTCCACGTGTT and R,
AGCACTGCATCGATTTTGCTC; CXCL11 F, TCGAAGCAAGCAAGGCTTAT and R,
TTCAGATGCTCTTTTCCAGGACT; CXCL12 F, AGTGTGCATTGACCCGAAGC and R,
GCAGGCCCTTCCCTAACAC; CXCL14 F, TGAAGCCAAAGTACCCGCAC and R,
TGACCTCGGTACCTGGACAC; VEGFA F, CTACCTCCACCATGCCAAGT and R,
GCAGTAGCTGCGCTGATAGA; VEGFR2 F, CAAGTGGCTAAGGGCATGGA and R,
ATTTCAAAGGGAGGCGAGCA; IL17RA F, CTGATGGGGACCCAAACCAC and R,
CCACAGGGTGAAGCTCACAC; and GAPDH F, CTGCACCACCAACTGCTTAG and R,
GAGCTTCCCGTTCAGCTCAG.
Transwell assay
Chemotaxis of HUVECs was evaluated using a transwell
assay. Briefly, HUVECs were cultured in DMEM with 2% FBS for 8 h.
Following this, 1×104 cells in DMEM with 2% FBS were
placed in the upper chamber with 8-µm pores (BD Biosciences)
pre-coated with 100 µg/ml Matrigel (BD Biosciences) for 30 min at
37°C. Cell-free media from liver cancer cells cultured with 2% FBS
for 24 h was added to the lower compartments of the chamber. For
the inhibitory assay, 400 nM CXCR2 inhibitor SB225002 (MedChem
Express) was added to the upper and lower compartments of the
chamber. HUVECs were cultured overnight, and then the cells on the
upper surface of the filters were wiped away using cotton swabs.
The filters were fixed with 4% paraformaldehyde/PBS for 10 min at
4°C, permeabilized with 0.2% Triton X-100/PBS for 10 min at 4°C and
stained with DAPI (Yeasen Shanghai Biotechnology Co., Ltd.) at room
temperature for 5 min. The number of invasive cells was observed
under a fluorescent microscope (BX51; Olympus Corporation) and
quantified by counting cells in five randomly selected high-powered
fields (magnification, ×400) in each well.
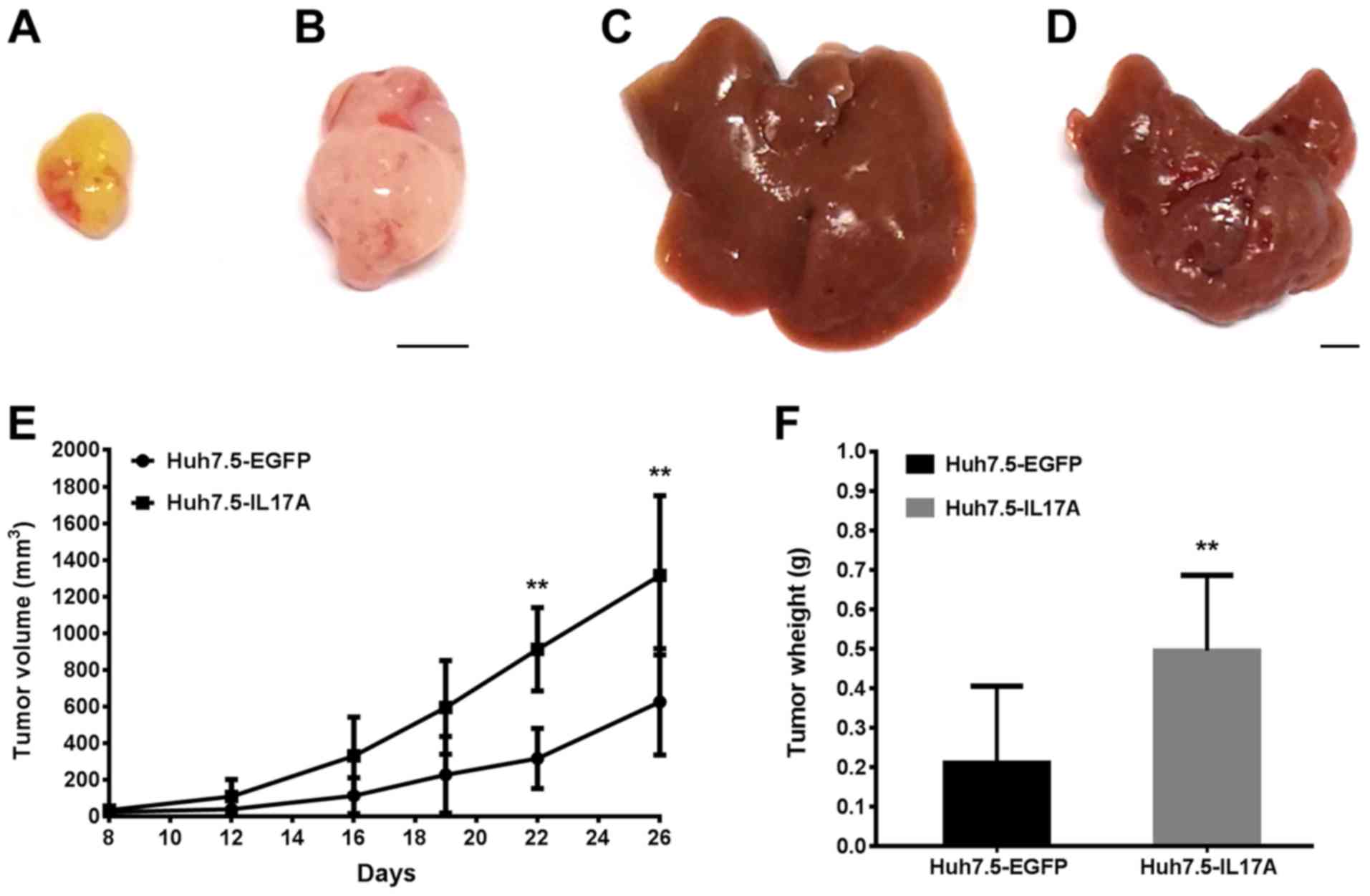
Xenograft tumor models
The Animal Studies Committee of China-Japan
Friendship Hospital approved all of the animal experiments in the
present study, which were performed according to the Principles of
Laboratory Animal Care (32). For
subcutaneous xenograft models, 6–8 week old female nude (nu/nu)
mice (Charles River Laboratories, Inc.), weighing 21.0±0.82 g were
used in the present study. Mice were maintained under
specific-pathogen-free conditions and had access to food and water
ad libitum. Mice were acclimatized to standardized
laboratory conditions for 1 week prior to experimentation (24±2°C;
50±10% relative humidity; 12 h light/dark cycle). Mice in the
control group were inoculated with 5×106 Huh7.5-EGFP
cells and the experimental group were inoculated with Huh7.5-IL17A
cells in 100 µl FBS-free culture medium in their backs. The
subcutaneous tumor volume was monitored at day 8, 12, 16, 19, 22
and 26 by measuring the tumor diameters (mm) along the largest (a)
and perpendicular (b) diameters, from which the tumor volume (V;
mm3) was calculated according to the formula: V=0.5 × a
× b2. For the liver metastasis model, the nude mice were
anesthetized with 240 mg/kg avertin (intraperitoneal;
Sigma-Aldrich; Merck KGaA). The skin was disinfected with 75%
alcohol and an oblique incision was made on the left side to pull
out the spleen. A total of 1×106 Huh7.5-EGFP cells in 25
µl FBS-free culture medium were injected into the spleen of the
control group and in the experimental group an equal number of
Huh7.5-IL17A cells in 25 µl FBS-free culture medium were injected
into the spleen. After injection and needle withdrawal, a dry
cotton swab was used for 2 min for hemostasis and cell leakage
oppression. Then, the spleen was replaced into the abdominal cavity
and the skin was sutured. The mice were euthanized and the tumors
were excised for histological analysis 3 and 4 weeks later for the
subcutaneous and liver metastasis models, respectively.
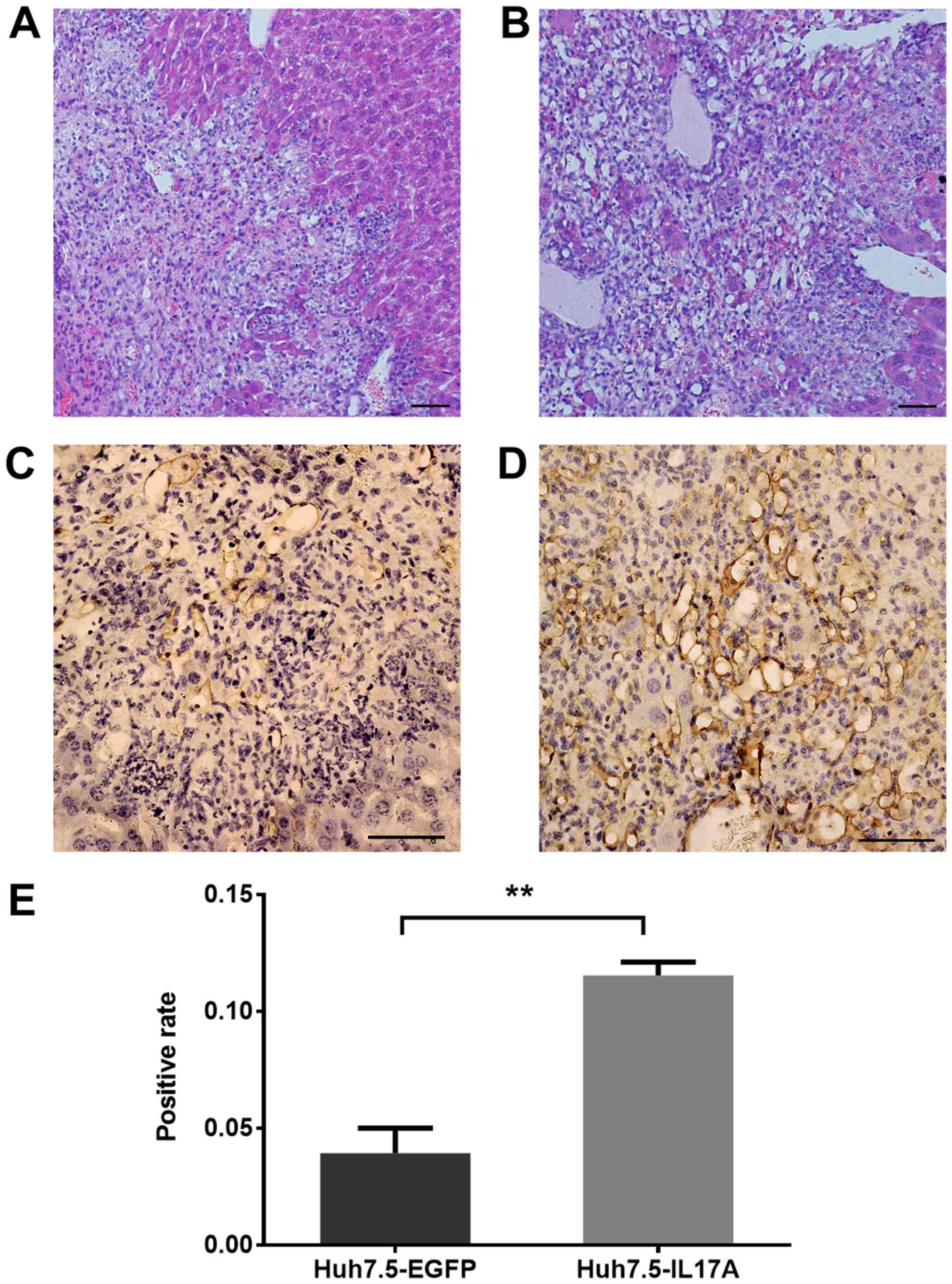
Histological analysis
Subcutaneous tumors or the livers were fixed in 4%
paraformaldehyde/PBS overnight at 4°C, and then prepared into 5 µm
paraffin sections for hematoxylin and eosin staining. The sections
were stained with hematoxylin for 5 min and eosin for 1 min at room
temperature. Immunohistochemistry was performed according to the
standard procedure. Briefly, the tumor slides were dewaxed in
xylene for 5 min twice, rehydrated in 100% ethanol for 5 min twice,
95% ethanol for 3 min twice and 70% ethanol for 1 min at room
temperature, and were washed with tap water. The slides were
subjected to antigen retrieval using 0.01 M citrate buffer (pH 6.0;
Beyotime Institute of Biotechnology) and boiling in the microwave
for 10 min, slides were then washed with tap water. The slides were
incubated in 3% H2O2 for 10 min at room
temperature and blocked in 3% goat serum (cat. no. ZLI-9022;
OriGene Technologies, Inc.) for 1 h at room temperature. The
primary rabbit anti-cluster of differentiation (CD)-31 antibody
(1:500; ab182981; Abcam) was applied to the slides overnight at
4°C. Sections were then incubated with goat-anti-rabbit secondary
antibody conjugated with horseradish peroxidase (ready to use; cat.
no. PV-6001; ZSGB-Bio) at room temperature for 30 min. Finally,
diaminobenzidine tetrachloride was used for color development and
the slides were counterstained with hematoxylin for 5 min at room
temperature. Slides were observed under a light microscope (BX53;
Olympus Corporation) at ×400 magnification and imaged using Canon
EOS600D camera (Canon, Inc.).
Statistical analysis
Sections showing positive staining for CD31 were
assessed in 6 fields of view (magnification, ×400) and the
percentage of positive staining was calculated with Image-Pro Plus
software (version 6.0; Media Cybernetics, Inc.). Student's t-test
was used to compare the statistical difference between two groups.
For data containing multiple groups, statistical analyses were
performed using one-way analysis of variance followed by the
Tukey's post hoc test using GraphPad Prism software (version 6.01;
GraphPad Software, Inc.). Data are presented as the mean ± SD.
P<0.05 was considered significant as indicated in the figure
legends.
Results
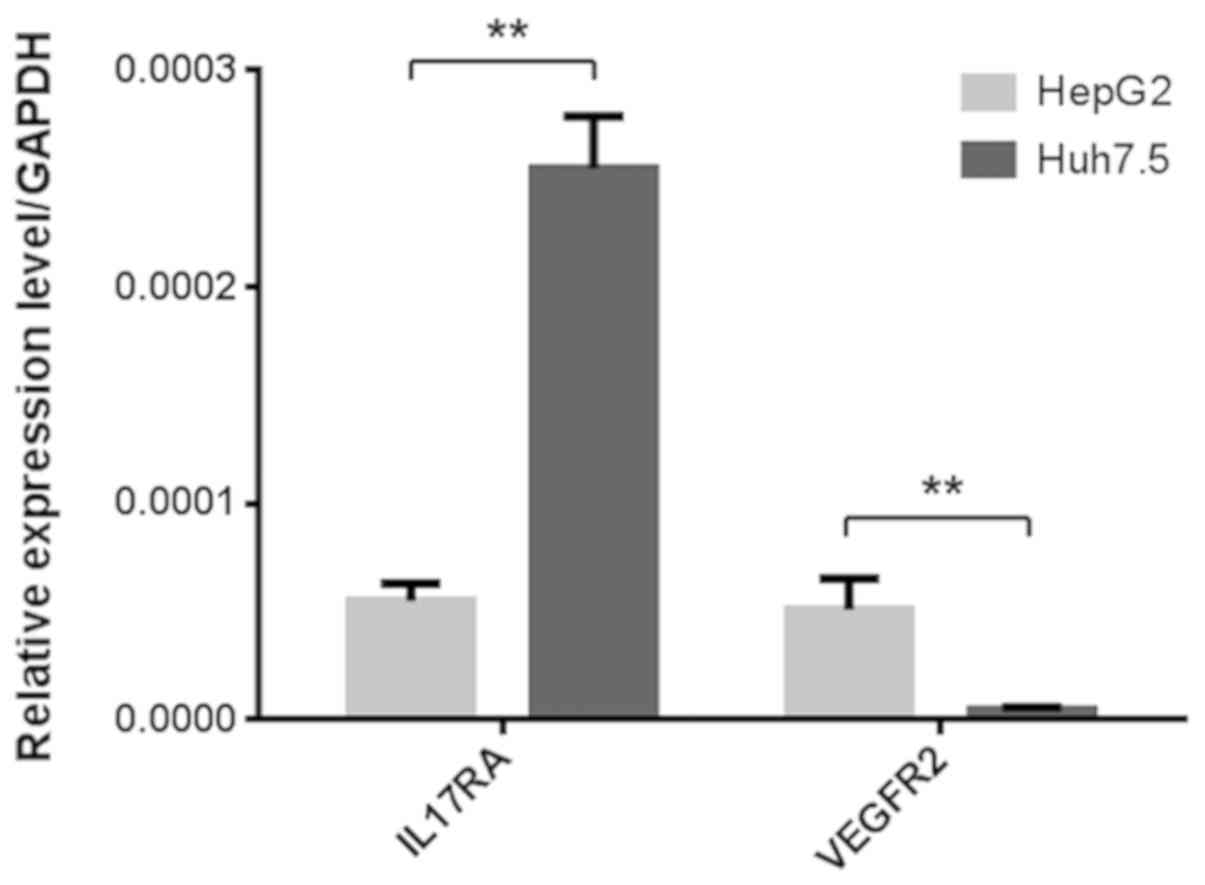
Expression of IL17RA and VEGFR2 on
liver cancer cells
Since IL-17A has been shown previously to upregulate
VEGFA in some tumor cells (16–18),
it was originally hypothesized that the potentially increased level
of VEGFA from liver cancer cells may act on its own VEGFR2 receptor
to stimulate cell proliferation. Hence, the present study first
examined the expression of IL17RA and VEGFR2 in Huh7.5 and HepG2
cells using RT-qPCR. IL17RA and VEGFR2 were expressed in both cell
lines, which was consistent with previous reports (33,34).
While VEGFR2 expression was relatively low in Huh7.5 cells, IL17RA
expression was significantly higher in Huh7.5 cells than in HepG2
cells (Fig. 1).
Overexpression of IL-17A does not
promote in vitro liver cancer cell proliferation
To study the effect of IL-17A on liver cancer cells,
the present study constructed IL-17A overexpressing cells using
lentiviral vectors. Although IL-17A is not naturally expressed in
liver cancer cells, this method allows for the study of the role of
IL-17 in the following in vivo tumor growth experiment, and
it has been commonly used in previous IL-17A studies (25,35,36).
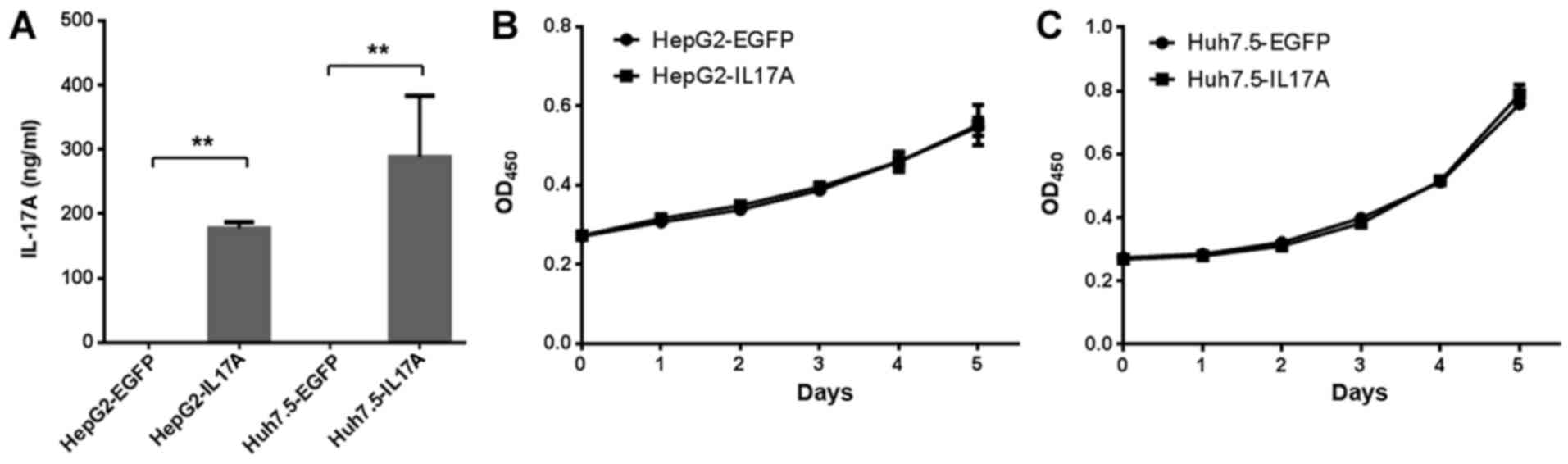
IL-17A secretion was detected at >100 ng/ml in Huh7.5-IL17A and
HepG2-IL17A cells but not Huh7.5-EGFP and HepG2-EGFP cells, when
the cell confluency was ~20% (Fig.
2A). The effect of IL-17A on the proliferation of liver cancer
cells was then determined. Using a CCK-8 assay, the overexpression
of IL-17A did not significantly affect the cell proliferation rate
of Huh7.5 or HepG2 cells (Fig. 2B and
C).
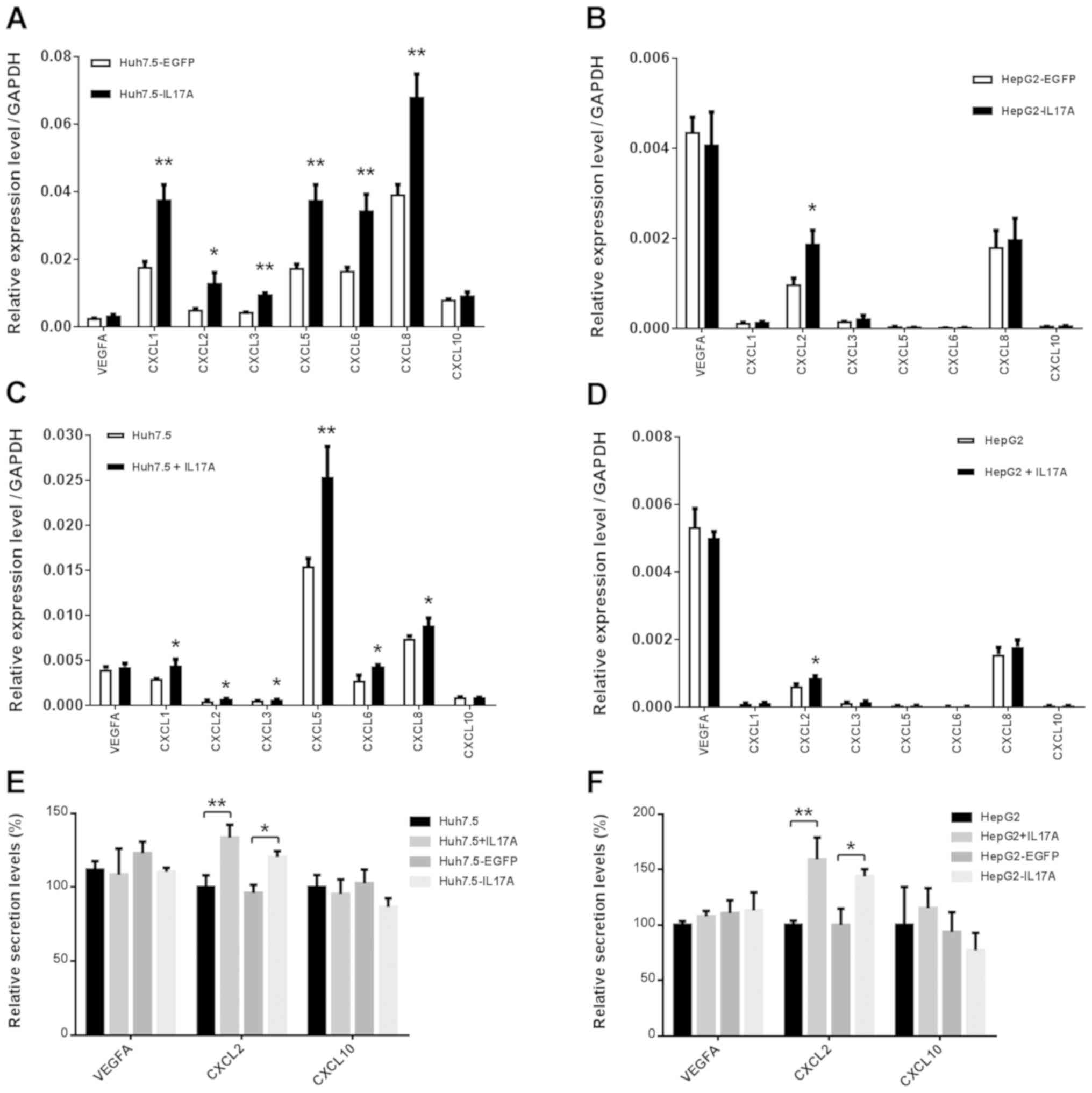
IL-17A upregulates the production of
proangiogenic CXC chemokines but not VEGFA in liver cancer
cells
Next, the present study examined the possibility
that IL-17A may upregulate VEGFA expression in liver cancer cells,
which may then in turn promote cell proliferation and angiogenesis.
Notably, the results revealed that the VEGFA expression was not
altered in IL-17A overexpressing cells, in either of the cell lines
tested (Fig. 3A and B). The
overexpression of IL-17A selectively and significantly upregulated
the expression of pro-angiogenic CXC chemokines CXCL1, CXCL2,
CXCL3, CXCL5, CXCL6 and CXCL8 in Huh7.5 cells, and CXCL2 in HepG2
cells, while the expression of the angiostatic chemokine CXCL10 was
unchanged (Fig. 3A and B). Other
CXC chemokines (data not shown) were expressed at extremely low
levels or not detected by RT-qPCR. These results are consistent
with those of recombinant IL-17A stimulation (Fig. 3C and D). The secretion of VEGFA,
CXCL2 and CXCL10 were further confirmed by ELISA in the presence or
absence of recombinant IL-17A, and in the IL-17A or EGFP
overexpressing cells (Fig. 3E and
F). These data suggest that the pro-angiogenic CXC chemokines
upregulated by IL-17A may promote angiogenesis in liver cancer.
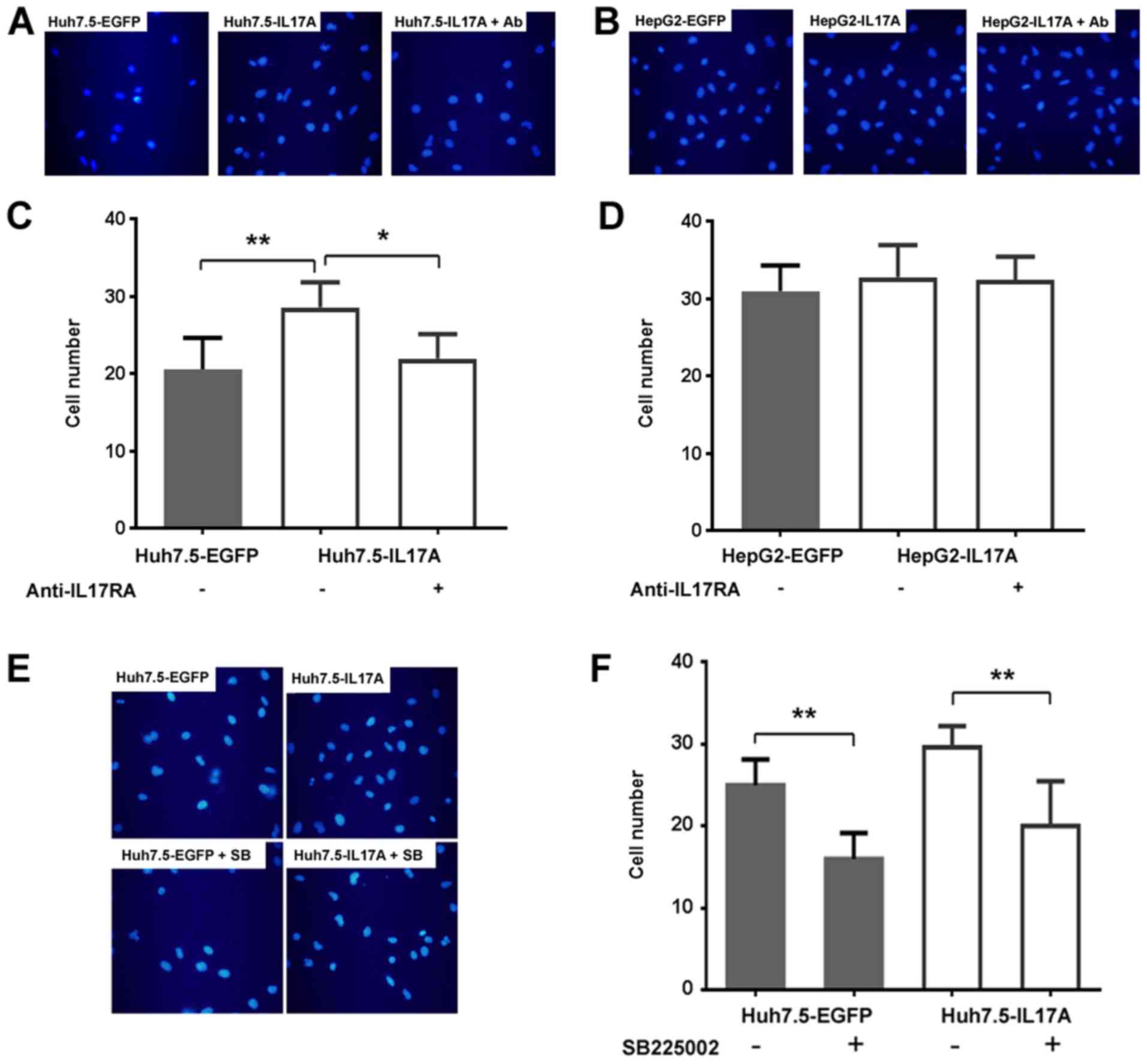
IL-17A-expressing Huh7.5 cells promote
endothelial chemotaxis in a CXCR2-dependent manner
As CXC chemokines do not directly stimulate
endothelial proliferation, but increase endothelial cell invasion,
the present study examined the chemotaxis effect of the
supernatants from IL-17A overexpressing liver cancer cells on
endothelial invasion using Transwell assays. The supernatant from
Huh7.5-IL17A cells significantly promoted HUVEC invasion compared
with Huh7.5-EGFP cells, and this promotion could be inhibited by
IL-17RA antibody (Fig. 4A and C),
indicating that the enhanced invasion of HUVECs was mediated by the
IL-17A-IL-17RA interaction in liver cancer cells. Since the
production of CXC chemokines did not respond to IL-17A in HepG2
cells as strongly as in Huh7.5 cells (Fig. 3A-D), it was unsurprising that the
overexpression of IL-17A in HepG2 did not significantly promote
HUVEC invasion in the same manner as observed in Huh7.5 cells
(Fig. 4B and D). Next, the present
study tested whether the CXCL-CXCR2 axis was responsible for the
enhanced chemotaxis effect in Huh7.5 cells by using the CXCR2
inhibitor SB225002. The addition of SB225002 to the assay
significantly suppressed HUVEC invasion of the supernatant from
Huh7.5-IL17A cells, indicating that IL-17A-stimulated endothelial
chemotaxis is dependent on CXCR2 signaling. At the same time,
SB225002 also inhibited the endothelial invasion of the supernatant
from Huh7.5-EGFP cells, indicating that the basal level of CXC
chemokines secreted from Huh7.5 cells may also function through
CXCR2 in the absence of IL-17A (Fig.
4E and F).
IL-17A promotes the in vivo growth and
angiogenesis of Huh7.5 cells in a mouse model
As Huh7.5 cells responded to IL-17A stimulation more
so than HepG2 cells based on CXC chemokine production and
endothelial cell chemotaxis, the present study performed the
following experiments using Huh7.5 cells. To study the in
vivo biological effect of IL-17A on liver cancer progression,
the present study determined Huh7.5 tumor growth in both
subcutaneous and liver xenograft models with or without IL17A
overexpression. It has been shown previously that the mouse and
human chemokine genes are closely related (37), with the human CXCR2 gene being
functionally equivalent to mouse CXCR2 (19). Furthermore, mouse CXCR2 has been
shown to respond to CXCL-chemokines from human cancer cell
xenografts (25,38). Therefore, it is possible that
IL-17A-stimulated CXC-chemokines from Huh7.5 cells could act
through mouse CXCR2 to promote angiogenesis and tumor growth in a
mouse model. Both the subcutaneous inoculation and intraspleen
injection produced solid tumors of Huh7.5 under the skin and in the
liver, respectively. Consistent with our hypothesis, the expression
of IL-17A significantly promoted the growth of Huh7.5 tumors in
both models compared with controls (Figs. 5, and 6A and B). The intensity of blood vessel
(CD31) staining was higher in the IL-17A-expressing group than the
EGFP-expressing group in the orthotopic models (Fig. 6C-E), and this result was similar in
the subcutaneous models (data not shown). Since IL-17A did not
directly augment cell proliferation in the in vitro assays,
the in vivo data suggests that IL-17A in the tumor
microenvironment promotes tumor growth by enhancing
angiogenesis.
Discussion
In the present study, it was demonstrated that
IL-17A could stimulate the secretion of angiogenic CXC chemokines
from liver cancer cells, which may recruit endothelial cells to the
tumor cells in a CXCR2-dependent manner. Tumor angiogenesis was
also promoted by IL-17A expression in vivo. The CXC
chemokines can be classified as angiogenic or angiostatic
predominantly based on the presence or absence of an ELR motif. The
angiogenic CXC chemokines include CXCL1, CXCL2, CXCL3, CXCL5,
CXCL6, CXCL8 and CXCL12, and the angiostatic chemokines include
CXCL4, CXCL9, CXCL10, CXCL11 and CXCL14 (21). IL-17A was shown to increase the
expression of CXCL1, CXCL2, CXCL3, CXCL5, CXCL6 and CXCL8 in Huh7.5
cells and upregulated CXCL2 in HepG2 cells. Additionally,
angiostatic CXC chemokines were not affected by IL-17A in both cell
lines. IL-17A has been reported to stimulate VEGFA production and
promote angiogenesis in several cancer cell lines (16–18)
and it has been shown previously that IL-17A does not affect VEGFA
production but rather stimulates CXCL1, CXCL5, CXCL6 and CXCL8
production in non-small cell lung cancer cell lines (25). The results of the present study are
consistent with those of Numasaki et al (25) in that IL-17A promoted angiogenesis
and was more dependent on the CXCL-CXCR2 axis than on VEGFA. Taken
together, this suggests that the effect of IL-17A on VEGFA
production may be dependent on the cancer cell type.
Anti-angiogenesis therapy, especially anti-VEGF
signaling drugs, has produced benefits in clinical trials for many
types of cancers. However, they have also had limited benefits in
patients with HCC (39,40), suggesting that another target may
be required for HCC treatment. We previously reported that besides
a bigger tumor size, elevated pre-therapy serum levels of IL-17 may
serve as an additional risk factor for the early recurrence of HCC
(13). Zhang et al
(14) have demonstrated that the
number of intratumoral Th17 cells positively correlates with
microvessel density, suggesting that the accumulation of
intratumoral IL-17-producing cells may promote tumor progression
via the promotion of angiogenesis. These observations together are
indicative of an important role for IL-17A in the progression of
HCC and IL-17A-promoted angiogenesis may be a target for HCC
therapy. In addition, the observation that IL-17A released in the
tumor microenvironment in response to VEGF-blocking drugs triggers
inflammatory and VEGF-independent angiogenesis that in turn induces
drug resistance (41), further
supports our finding that IL-17A may excite chemokine-induced
angiogenesis independent of VEGF signaling.
CXCR2 is the putative receptor for pro-angiogenic
CXC chemokines on endothelial cells (20). Previous studies have shown that in
addition to mediating an inflammatory response, the CXC chemokine
receptors CXCR1 and CXCR2 play important roles in tumor progression
and metastasis; CXCR2 knockout mice significantly repress tumor
growth and angiogenesis (38,42).
In preclinical models of ovarian cancer, sorafenib treatment
markedly elevated the expression of pro-angiogenic CXC chemokines,
and CXCR2 inhibitor treatment stabilized the progression of
sorafenib-resistant tumors (43),
indicating that the CXCL-CXCR2 pathway may be important in
compensating for the VEGF signaling pathway suggesting that the
simultaneous blockage of these two pathways could improve the
outcomes of anti-angiogenic tumor therapy. In the present study,
blocking of CXCR2 significantly reduced endothelial chemotaxis to
the Huh7.5 supernatant. Since CXC chemokines were produced at the
basal level even without IL-17A stimulation, the CXCR2 inhibitor
not only inhibited IL-17A-stimulated chemotaxis, but also
chemotaxis in the absence of IL-17A, indicating that CXCR2 may be a
potential target for anti-angiogenesis in liver cancer.
The present study also demonstrated that different
liver cancer cell lines may respond differently to IL-17A. Although
IL-17A did not affect the growth rate of Huh7.5 and HepG2 cells,
the expression of CXC chemokines in Huh7.5 cells increased much
more than in HepG2 cells in the presence of IL-17A, and the
stimulatory effect of IL-17A on endothelial chemotaxis was stronger
in Huh7.5 cells. Since the expression of IL17RA was much higher in
Huh7.5 cells than in HepG2 cells, it would be of interest to
investigate whether the increased response to IL-17A observed in
Huh7.5 cells was due to the higher expression of the receptor. In
addition, the expression levels of VEGFA and VEGFR2 were higher in
HepG2 cells than Huh7.5 cells, and HepG2 exhibited slightly
stronger endothelial cell chemotaxis without IL-17A stimulation.
Whether the different expression ratio of IL17RA and VEGFR2
regulates angiogenesis and tumor growth in vitro and in
vivo may possess further clinical implications, and may be
investigated in future work.
Acknowledgements
The authors would like to thank Dr Aihua Zheng
(Chinese Academy of Sciences, Beijing, China) for generously
providing the Huh7.5 cells, and Dr Peigang Wang (Capital Medical
University, Beijing, China) for providing the HUVEC cells.
Funding
The present study was supported by Youth Foundation
of China-Japan Friendship Hospital (grant no. 2015-1-QN-19).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL, ZW and ZY designed the experiment. LL acquired
the funding. LL and XL performed the animal experiments. HS and SW
performed the cell experiments and analyzed the data. HT and YS
performed the PCR experiments. SS and LX performed the ELISA
experiments. JH and WZ performed the histological examinations. ZW
and ZY supervised the research group. ZW wrote and revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The Animal Studies Committee of China-Japan
Friendship Hospital approved all of the animal experiments in the
present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ouyang W, Kolls JK and Zheng Y: The
biological functions of T helper 17 cell effector cytokines in
inflammation. Immunity. 28:454–467. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marengo A, Rosso C and Bugianesi E: Liver
cancer: Connections with obesity, fatty liver, and cirrhosis. Annu
Rev Med. 67:103–117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gomes AL, Teijeiro A, Buren S, Tummala KS,
Yilmaz M, Waisman A, Theurillat JP, Perna C and Djouder N:
Metabolic Inflammation-Associated IL-17A causes Non-alcoholic
steatohepatitis and hepatocellular carcinoma. Cancer Cell.
30:161–175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
He D, Li H, Yusuf N, Elmets CA, Li J,
Mountz JD and Xu H: IL-17 promotes tumor development through the
induction of tumor promoting microenvironments at tumor sites and
myeloid-derived suppressor cells. J Immunol. 184:2281–2288. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sun C, Kono H, Furuya S, Hara M, Hirayama
K, Akazawa Y, Nakata Y and Fujii H: Interleukin-17A plays a pivotal
role in chemically induced hepatocellular carcinoma in mice. Dig
Dis Sci. 61:474–488. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang B, Kang H, Fung A, Zhao H, Wang T and
Ma D: The role of interleukin 17 in tumour proliferation,
angiogenesis, and metastasis. Mediators Inflamm. 2014:6237592014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Grivennikov SI and Karin M: Dangerous
liaisons: STAT3 and NF-kappaB collaboration and crosstalk in
cancer. Cytokine Growth Factor Rev. 21:11–19. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Yi T, Zhang W, Pardoll DM and Yu
H: IL-17 enhances tumor development in carcinogen-induced skin
cancer. Cancer Res. 70:10112–10120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang B, Rong G, Wei H, Zhang M, Bi J, Ma
L, Xue X, Wei G, Liu X and Fang G: The prevalence of Th17 cells in
patients with gastric cancer. Biochem Biophys Res Commun.
374:533–537. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen WC, Lai YH, Chen HY, Guo HR, Su IJ
and Chen HH: Interleukin-17-producing cell infiltration in the
breast cancer tumour microenvironment is a poor prognostic factor.
Histopathology. 63:225–233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu C, Hao K, Yu L and Zhang X: Serum
interleukin-17 as a diagnostic and prognostic marker for non-small
cell lung cancer. Biomarkers. 19:287–290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang X, Weng W, Xu W, Wang Y, Yu W, Tang
X, Ma L, Pan Q, Wang J and Sun F: Prognostic significance of
interleukin 17 in cancer: A meta-analysis. Int J Clin Exp Med.
7:3258–3269. 2014.PubMed/NCBI
|
|
13
|
Wu J, Du J, Liu L, Li Q, Rong W, Wang L,
Wang Y, Zang M, Wu Z, Zhang Y and Qu C: Elevated pretherapy serum
IL17 in primary hepatocellular carcinoma patients correlate to
increased risk of early recurrence after curative hepatectomy. PLoS
One. 7:e500352012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang JP, Yan J, Xu J, Pang XH, Chen MS,
Li L, Wu C, Li SP and Zheng L: Increased intratumoral
IL-17-producing cells correlate with poor survival in
hepatocellular carcinoma patients. J Hepatol. 50:980–989. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Numasaki M, Fukushi J, Ono M, Narula SK,
Zavodny PJ, Kudo T, Robbins PD, Tahara H and Lotze MT:
Interleukin-17 promotes angiogenesis and tumor growth. Blood.
101:2620–2627. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pan B, Shen J, Cao J, Zhou Y, Shang L, Jin
S, Cao S, Che D, Liu F and Yu Y: Interleukin-17 promotes
angiogenesis by stimulating VEGF production of cancer cells via the
STAT3/GIV signaling pathway in non-small-cell lung cancer. Sci Rep.
5:160532015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu X, Yang T, Liu X, Guo JN, Xie T, Ding
Y, Lin M and Yang H: IL-17 promotes tumor angiogenesis through
Stat3 pathway mediated upregulation of VEGF in gastric cancer.
Tumour Biol. 37:5493–5501. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu J, Duan Y, Cheng X, Chen X, Xie W,
Long H, Lin Z and Zhu B: IL-17 is associated with poor prognosis
and promotes angiogenesis via stimulating VEGF production of cancer
cells in colorectal carcinoma. Biochem Biophys Res Commun.
407:348–354. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mihara K, Smit MJ, Krajnc-Franken M,
Gossen J, Rooseboom M and Dokter W: Human CXCR2 (hCXCR2) takes over
functionalities of its murine homolog in hCXCR2 knockin mice. Eur J
Immunol. 35:2573–2582. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Addison CL, Daniel TO, Burdick MD, Liu H,
Ehlert JE, Xue YY, Buechi L, Walz A, Richmond A and Strieter RM:
The CXC chemokine receptor 2, CXCR2, is the putative receptor for
ELR+ CXC chemokine-induced angiogenic activity. J Immunol.
165:5269–5277. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Palacios-Arreola MI, Nava-Castro KE,
Castro JI, Garcia-Zepeda E, Carrero JC and Morales-Montor J: The
role of chemokines in breast cancer pathology and its possible use
as therapeutic targets. J Immunol Res. 2014:8497202014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kitadai Y, Haruma K, Sumii K, Yamamoto S,
Ue T, Yokozaki H, Yasui W, Ohmoto Y, Kajiyama G, Fidler IJ and
Tahara E: Expression of interleukin-8 correlates with vascularity
in human gastric carcinomas. Am J Pathol. 152:93–100.
1998.PubMed/NCBI
|
|
23
|
Mestas J, Burdick MD, Reckamp K, Pantuck
A, Figlin RA and Strieter RM: The role of CXCR2/CXCR2 ligand
biological axis in renal cell carcinoma. J Immunol. 175:5351–5357.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu B, Lin N, Zhang M, Zhu Y, Cheng H,
Chen S, Ling Y, Pan W and Xu R: Activated hepatic stellate cells
promote angiogenesis via interleukin-8 in hepatocellular carcinoma.
J Transl Med. 13:3652015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Numasaki M, Watanabe M, Suzuki T,
Takahashi H, Nakamura A, McAllister F, Hishinuma T, Goto J, Lotze
MT, Kolls JK and Sasaki H: IL-17 enhances the net angiogenic
activity and in vivo growth of human non-small cell lung cancer in
SCID mice through promoting CXCR-2-dependent angiogenesis. J
Immunol. 175:6177–6189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou Y, Wu PW, Yuan XW, Li J and Shi XL:
Interleukin-17A inhibits cell autophagy under starvation and
promotes cell migration via TAB2/TAB3-p38 mitogen-activated protein
kinase pathways in hepatocellular carcinoma. Eur Rev Med Pharmacol
Sci. 20:250–263. 2016.PubMed/NCBI
|
|
27
|
Li J, Lau GK, Chen L, Dong SS, Lan HY,
Huang XR, Li Y, Luk JM, Yuan YF and Guan XY: Interleukin 17A
promotes hepatocellular carcinoma metastasis via NF-κB induced
matrix metalloproteinases 2 and 9 expression. PLoS One.
6:e218162011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Knowles BB, Howe CC and Aden DP: Human
hepatocellular carcinoma cell lines secrete the major plasma
proteins and hepatitis B surface antigen. Science. 209:497–499.
1980. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lopez-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sumpter R Jr, Loo YM, Foy E, Li K,
Yoneyama M, Fujita T, Lemon SM and Gale M Jr: Regulating
intracellular antiviral defense and permissiveness to hepatitis C
virus RNA replication through a cellular RNA helicase, RIG-I. J
Virol. 79:2689–2699. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guide for the care and use of Laboratory
Animals. Eighth editor; Washington, D.C.: 2011
|
|
33
|
Huang J, Zhang X, Tang Q, Zhang F, Li Y,
Feng Z and Zhu J: Prognostic significance and potential therapeutic
target of VEGFR2 in hepatocellular carcinoma. J Clin Pathol.
64:343–348. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liao R, Sun J, Wu H, Yi Y, Wang JX, He HW,
Cai XY, Zhou J, Cheng YF, Fan J and Qiu SJ: High expression of
IL-17 and IL-17RE associate with poor prognosis of hepatocellular
carcinoma. J Exp Clin Cancer Res. 32:32013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tartour E, Fossiez F, Joyeux I, Galinha A,
Gey A, Claret E, Sastre-Garau X, Couturier J, Mosseri V, Vives V,
et al: Interleukin 17, a T-cell-derived cytokine, promotes
tumorigenicity of human cervical tumors in nude mice. Cancer Res.
59:3698–3704. 1999.PubMed/NCBI
|
|
36
|
Hu J, Ye H, Zhang D, Liu W, Li M, Mao Y
and Lu Y: U87MG glioma cells overexpressing IL-17 acclerate
early-stage growth in vivo and cause a higher level of CD31 mRNA
expression in tumor tissues. Oncol Lett. 6:993–999. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nomiyama H, Osada N and Yoshie O:
Systematic classification of vertebrate chemokines based on
conserved synteny and evolutionary history. Genes Cells. 18:1–16.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Singh S, Varney M and Singh RK: Host
CXCR2-dependent regulation of melanoma growth, angiogenesis, and
experimental lung metastasis. Cancer Res. 69:411–415. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sun W and Cabrera R: Systemic treatment of
patients with advanced, unresectable hepatocellular carcinoma:
Emergence of therapies. J Gastrointest Cancer. 49:107–115. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Raoul JL, Gilabert M, Adhoute X and
Edeline J: An in-depth review of chemical angiogenesis inhibitors
for treating hepatocellular carcinoma. Expert Opin Pharmacother.
18:1467–1476. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chung AS, Wu X, Zhuang G, Ngu H, Kasman I,
Zhang J, Vernes JM, Jiang Z, Meng YG, Peale FV, et al: An
interleukin-17-mediated paracrine network promotes tumor resistance
to anti-angiogenic therapy. Nat Med. 19:1114–1123. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Keane MP, Belperio JA, Xue YY, Burdick MD
and Strieter RM: Depletion of CXCR2 inhibits tumor growth and
angiogenesis in a murine model of lung cancer. J Immunol.
172:2853–2860. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Devapatla B, Sharma A and Woo S: CXCR2
Inhibition combined with sorafenib improved antitumor and
antiangiogenic response in preclinical models of ovarian Cancer.
PLoS One. 10:e01392372015. View Article : Google Scholar : PubMed/NCBI
|