Introduction
Oral cancer is one of the major head and neck
malignant tumors. In terms of overall incidence, it is the sixth
most common form of cancer and carries a high risk of recurrence
(20–30%) (1). The annual worldwide
incidence was 300,000 new cases in 2012, with two-thirds occurring
in men (2). Although head and neck
cancers generally form in the pharynx or larynx, ~50% of all cases
involve the oral cavity (1). Such
cancers most commonly involve the tongue and occur in the gingiva,
buccal mucosa, floor of the mouth, palate, or lip. The significant
risk factors contributing to the etiology of oral cancer are
alcohol consumption and tobacco smoking (1). Histologically, >90% of oral
cancers are diagnosed as oral squamous cell carcinoma (OSCC)
(3). The 5-year survival rate of
patients with OSCC has remained at ~50% for several decades in
spite of advancements of treatment, including surgery, radiation,
and chemotherapy (4). The poor
survival rate is related to locoregional relapse and regional lymph
node metastasis (5). Thus, OSCC is
still a challenging disease to treat, and it is important to
develop a novel antiproliferative treatment method for OSCC.
Green tea is a common beverage worldwide and has
been studied regarding its usefulness for health (6,7). An
epidemiological study has revealed that green tea has preventive
effects against cancer (8). The
biological activity of green tea is attributed to its polyphenol
components (constituting 30% of dry weight) (9). Polyphenols in green tea include many
catechins-mainly epigallocatechin-3-gallate (EGCG),
epigallocatechin (EGC), epicatechin-3-gallate (ECG), and
epicatechin (EC)-whereas EGCG is the most effective polyphenol
(6). The use of EGCG has been
shown to suppress cancer progression in vitro and in animal
models: Not only the initiation but also progression or metastasis,
in several cancer types such as lung, liver, breast, colorectal,
prostate and skin cancer (9).
Nonetheless, to the best of our knowledge, few studies have
addressed the effect of EGCG on human OSCC cells, especially in an
experimental animal model. In this study, we evaluated the
influence of EGCG on a human OSCC cell line, HSC-3, then on an
in vivo xenograft mouse model, by investigating cell
proliferation and apoptosis. Finally, we discuss the therapeutic
potential of EGCG for oral-cancer therapy.
Materials and methods
Reagents
EGCG was purchased from Sigma-Aldrich (cat. no.
E4143), and cell titer 96® aqueous one solution
cytotocity assay (an MTS assay kit) from Promega. Propidium iodide
(PI) was acquired from Cayman Chemical (cat. no. 14289), whereas
ribonuclease A from Sigma-Aldrich (cat. no. R6513).
ApoScreen® Annexin V Apoptosis kit-FITC was bought from
Southern Biotech Birmingham, and Amplite™ Fluorimetric Caspase-3/7
Activity kit from AAT Bioquest. The In situ Apoptosis
Detection kit (TdT-mediated dUTP nick end labeling (TUNEL) assay)
was purchased from Takara Bio, Inc., a rabbit anti-Ki-67 monoclonal
antibody (cat. no. ab16667) from Abcam, and staurosporine and other
chemicals from Wako Pure Chemical Industries, Ltd.
Cell culture conditions
The HSC-3 cell line (purchased from the Japanese
Cancer Research Resources Bank, Tokyo, Japan) was used in this
study. This cell line consists of primary tumor cells originating
from a moderately differentiated squamous cell carcinoma (SCC) of
the human tongue with lymph node metastasis (3). This cell line is one of the most
commonly used for experimental study of OSCC with an allusion to
their origin and biological behavior (3). The cells were cultured in the
α-minimum essential medium (α-MEM; Invitrogen; Thermo Fisher
Scientific, Inc.) with 10% of fetal calf serum (FCS; BioWest,
Nuaillé, France). Penicillin (100 IU/ml) and streptomycin (100
mg/ml) (Invitrogen; Thermo Fisher Scientific, Inc.) were added to
the medium. The cells were grown at 37°C in a humidified atmosphere
containing 5% of CO2. The cells were subcultured every 3
days when confluence reached 80%.
The MTS assay
For this cytotoxity assay, 5×103 cells
were seeded in 96-well plates in α-MEM with 10% of FCS and cultured
for 24 h. Then, the cells were treated with various concentrations
of EGCG (0, 25, 50, 75 and 100 µM) in 100 µl of α-MEM with 1% of
FCS for 24, 48 and 72 h. Cell viability was assessed by the MTS
assay according to the manufacturer's instructions. Bioreduction of
tetrazolium was measured as absorbance at 490 nm on a 96-well plate
reader (SpectraMax M5; Molecular Devices), and the growth
inhibition rate was calculated.
Cell cycle analysis
A total of 106 cells were seeded in a 10
cm dish containing α-MEM with 10% of FCS and were cultured for 24
h. Then, the cells were incubated with or without 50 µM EGCG in
α-MEM with 5% of FCS for 24 h. The cells after treatment (0 and 24
h) were collected, washed with phosphate-buffered saline (PBS)
twice, and fixed with 70% ethanol at −20°C overnight. Then, the
cells were centrifuged and washed with PBS twice, resuspended in
900 µl of PBS containing 0.25 mg/ml ribonuclease A, and incubated
at 37°C for 30 min, followed by addition of PBS with 100 µl of 500
µg/ml PI, and incubated at 4°C for 15 min in the dark. Lastly, DNA
contents were analyzed on a flow cytometer (BD FACSCanto II; BD
Biosciences).
Annexin V-FITC analysis
The proportion (%) of apoptotic cells was determined
according to the manufacturer's manual by means of the Annexin V
Apoptosis kit-FITC and the flow cytometer. Namely, 106
cells were seeded in a 10 cm dish in α-MEM with 10% of FCS and
cultured for 24 h. Next, the cells were incubated with or without
50 µM EGCG in α-MEM with 1% of FCS for 6 h. Five hundred nM
staurosporine served as a positive control. The cells were
collected, washed with PBS twice, and resuspended in cold Annexin
binding buffer to a concentration of 106 cells/ml. Then,
10 µl of the Annexin V-FITC conjugate in a buffer was added to the
cells and incubated for 15 min at 4°C with protection from light.
Finally, we added 10 µl of a PI solution in a buffer and analyzed
the cells by flow cytometry. Annexin
V-FITC+/PI− cells are defined as early
apoptotic cells, and Annexin V-FITC+/PI+
cells are defined as late apoptotic cells.
Measurement of caspase-3 and −7
activities
These activities were determined with the Amplite™
Fluorimetric Caspase-3/7 Activity kit. A total of 5×104
cells per well were seeded in 96-well plates in α-MEM with 10% of
FCS and cultured for 24 h. Then, the cells were incubated with or
without 50 µM EGCG in α-MEM with 1% of FCS for 12 h. Five hundred
nM staurosporine served as a positive control. The cells were
rinsed once with ice-cold PBS and cultured in 100 µl of PBS.
Subsequently, 1 µl of a caspase-3 and −7 inhibitor (Ac-DEVD-CHO, a
synthetic tetrapeptide competitive inhibitor with a sequence
Ac-Asp-Glu-Val-Asp-CHO, which contains the amino acid sequence of
the PARP cleavage site) was added to selected samples as a negative
control. The cells were lysed with 100 µl of the caspase-3/7 assay
solution, and the plate was incubated for 2 h at room temperature
with protection from light. Cleavage substrate fluorescence was
measured on a 96-well plate reader at Excitation/Emission=350/450
nm (SpectraMax M5; Molecular Devices).
TUNEL staining in vitro
in vitro apoptosis was determined by the
TUNEL assay using the In situ Apoptosis Detection kit
(Takara Bio, Inc.). A total of 5×104 cells per well were
seeded in 8-well slide chambers (Watson Bio Laboratory) in α-MEM
with 10% of FCS and were cultured for 24 h. After that, the cells
were incubated with or without 50 µM EGCG in α-MEM with 1% of FCS
for 24 h. Five hundred nM staurosporine served as a positive
control. The cells were rinsed once with ice-cold PBS and fixed
with 4% paraformaldehyde in PBS (pH 7.4). Thereafter, the
endogenous peroxide activity was eliminated with 0.3%
H2O2 methanol solution for 30 min at room
temperature. The cells were dipped in permeabilization buffer for 5
min at 4°C. The cells were incubated with 50 µl of a labeling
reaction mixture (consisting of TdT Enzyme 5 µl + Labeling Safe
Buffer 45 µl) in a 37°C humidified chamber for 90 min. Then, they
were reacted with 50 µl of an anti-FITC antibody conjugated with
horseradish peroxidase (not diluted, cat. no. MK503; Takara Bio,
Inc.) for 30 min at 37°C. Immunoreactivity was visualized by
immersing the sections in 3,3′-diaminobenzidine (DAB; Dako, Agilent
Technologies, Inc.) for 10 min at room temperature. Subsequently,
the sections were counterstained with 3% methyl green for 1 min,
dehydrated in 100% ethanol, which was replaced by Clear Plus (Falma
Co., Ltd., Tokyo, Japan). Apoptotic cells were counted at
magnification ×400. The proportion of apoptotic cells was
calculated by counting the TUNEL-positive cells and by dividing
this number by the total number of tumor cells and then multiplying
by 100%, in a minimum of seven microscopic fields under a light
microscope.
The in vivo xenograft murine
model
A total of 20 BALB/c nude (nu/nu) mice (5-week-old
females; CLEA Japan, Inc.) were used in this study. The mice were
kept in groups of three or four per cage and provided with food and
water ad libitum. The animals were maintained under the
following conditions: 12 h light/12 h dark cycle, 24°C±2°C, and
50±10% relative humidity (mean, range). A total of 5×106
HSC-3 cells in 50 µl of α-MEM supplemented with 10% FCS were mixed
with 50 µl of Matrigel (BD Biosciences) and implanted
subcutaneously into the back of anesthetized mice. Tumor-bearing
mice were subdivided into the EGCG group and control group. Two
weeks after HSC-3 cells implantation, 75 mg/kg EGCG or saline was
intraperitoneally injected via a syringe as described in previous
reports (10,11). EGCG was administered twice a week
for 4 weeks. The tumors were allowed to reach ~40 mm3 in
size. Body weight was recorded, and weight loss exceeding 20% was
assumed to be a humane endpoint for euthanasia. Tumor diameters
were measured with vernier calipers. Tumor volume was calculated by
means of the following formula: (0.5 × length × width2),
as described by Yoshida et al (12). The mice were then euthanized, and
the tumor was carefully removed along with the overlying skin and
the surrounding tissue. The specimens were fixed with 4%
paraformaldehyde and embedded in paraffin for histological
examination. All the animal experiments were permitted by the
Animal Ethics Committee of the University of Fukui (approval no.
27110) and followed the Guide for the Care and Use of Laboratory
Animals (National Institutes of Health, USA).
Immunohistochemistry
The tissue sections were deparaffinized in Clear
Plus for 15 min and rehydrated in ethanol solutions of descending
concentrations. For antigen retrieval, the sections were immersed
in Tris-EDTA buffer (pH 9.0) and incubated at 95°C for 30 min. The
endogenous peroxide activity was eliminated with 0.3%
H2O2 methanol solution for 30 min at room
temperature. Normal horse serum (2.5%) for 30 min at room
temperature was used to block nonspecific immunoreactions. The
sections were incubated with a rabbit anti-Ki-67 monoclonal
antibody (dilution 1:100) for 120 min at room temperature. The
sections were then incubated with ImmPRESS (peroxidase) polymer
anti-rabbit IgG reagent (cat. no. MP-7401; Vector Laboratories) for
30 min at room temperature. Immunoreactivity was visualized by
immersing the sections in DAB. Subsequently, the sections were
counterstained with hematoxylin for 30 sec at room temperature,
dehydrated in ethanol solutions with ascending concentration, and
finally incubated with Clear Plus at room temperature. The sections
were rinsed three times in PBS between all the steps. The
percentage of Ki-67-positive cells among all tumor cells was
determined by cell counting in at least seven microscopic fields of
vision (at magnification ×400) per histological section.
TUNEL staining in vivo
The TUNEL assay was carried out to evaluate
apoptosis using the In situ Apoptosis Detection kit (Takara
Bio, Inc.). Briefly, the sections were deparaffinized in Clear Plus
for 15 min, rehydrated in 100% ethanol for 15 min, and
permeabilized with 10 µg/ml proteinase K (Invitrogen; Thermo Fisher
Scientific, Inc.) for 10 min at room temperature. The endogenous
peroxide activity was eliminated with 0.3%
H2O2 methanol solution for 5 min at room
temperature. The sections were incubated with 50 µl of a labeling
reaction mixture (consisting of TdT Enzyme 5 µl + Labeling Safe
Buffer 45 µl) in a 37°C humidified chamber for 90 min. Then, they
were reacted with 70 µl of the anti-FITC antibody conjugated with
horseradish peroxidase (not diluted, cat. no. MK503; Takara Bio,
Inc.) for 30 min at 37°C. Immunoreactivity was visualized by
immersing the sections in DAB for 10 min at room temperature. Next,
the sections were counterstained with hematoxylin for 10 sec and
dehydrated in 100% ethanol, which was replaced by Clear Plus. EXCEL
Mount (Falma) served for mounting. The sections were rinsed three
times in PBS between all the steps. The proportion of apoptotic
cells was evaluated by counting the TUNEL-positive cells among all
tumor cells, avoiding necrotic tumor areas, in a minimum of seven
visual fields in each individual section under a light microscope
(at magnification ×400).
Statistical analysis
All statistical analyses were performed in the
StatMate software (version 1.1; ATMS Co., Ltd.; for Macintosh). The
measured values were presented as the mean with standard deviation
(SD). Differences between groups were analyzed with unpaired t test
(cell cycle, apoptosis cells in vitro, TUNEL staining in
vitro, tumor volume, body weight, Ki-67 staining in vivo
and TUNEL staining in vivo) or one-way ANOVA with
Tukey-Kramer's multiple comparison post-hoc test (cell viability
and caspase-3/7 activity). P<0.05 was considered to indicate a
statistically significant difference.
Results
EGCG suppresses HSC-3 cell
proliferation in vitro
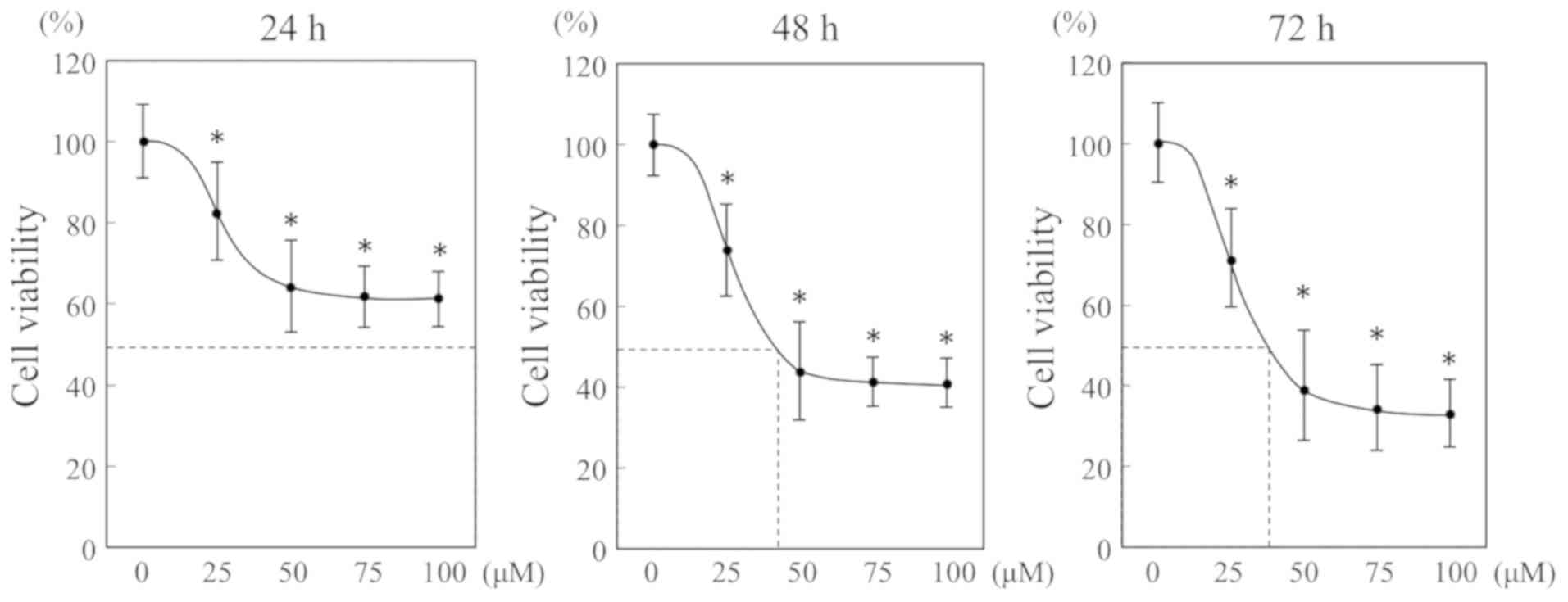
To examine the effect of EGCG on OSCC cells, HSC-3
cells were cultured with different concentration of EGCG for
various periods. Based on the cell viability, the inhibition rate
was calculated by means of the MTS assay. EGCG significantly
inhibited cell viability in a dose- and time-dependent manner
(Fig. 1). The IC50
value at 24, 48 and 72 h were >100, 43.2 and 39.3 µM,
respectively. There was no significant difference in inhibitory
effect at concentrations above 50 µM. Thus, we used 50 µM of EGCG
for the following experiments.
EGCG arrests HSC-3 cells in the G1
phase in vitro
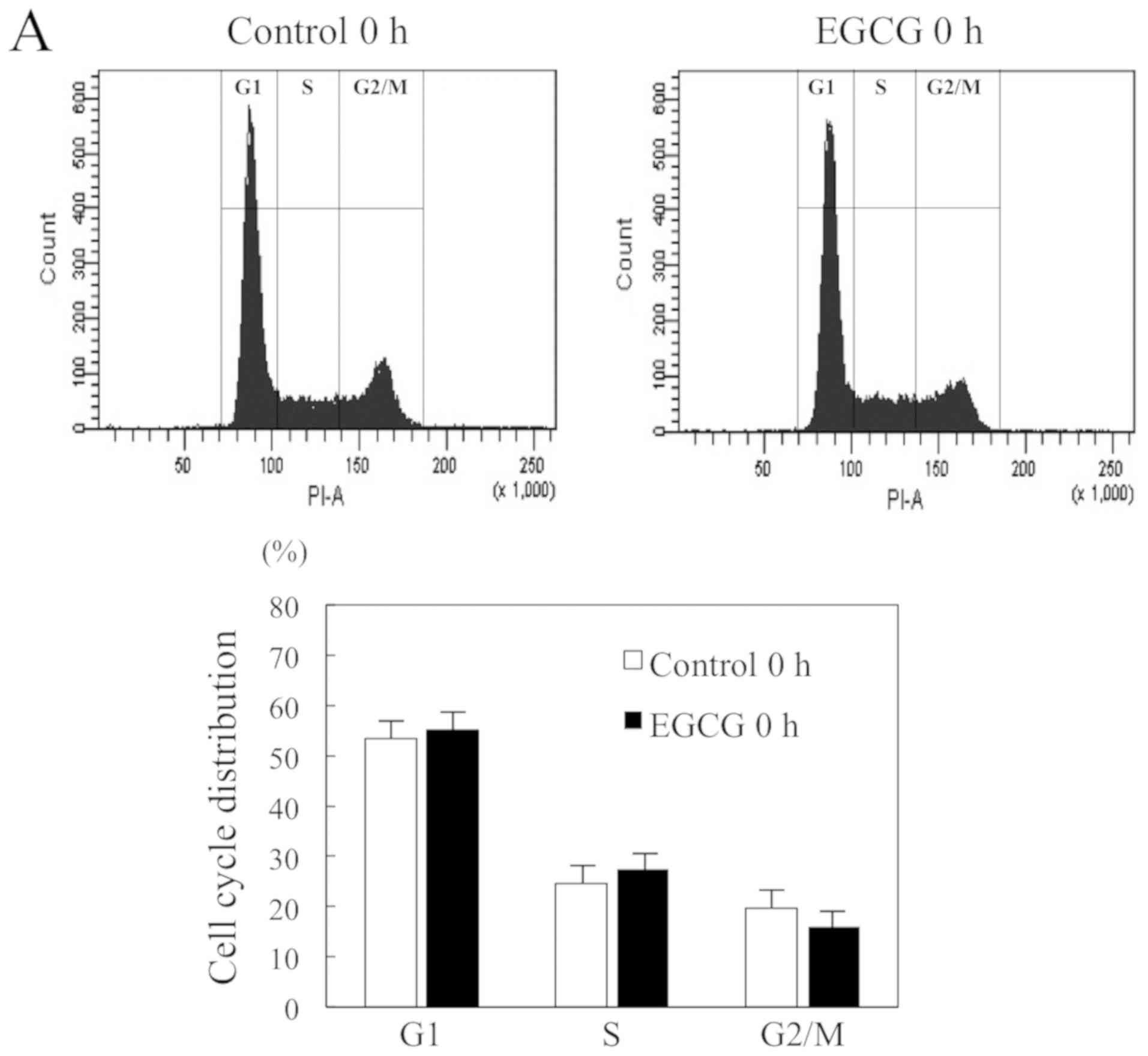
To identify the mechanism by which EGCG influences
cell viability, we performed cell cycle analysis. We examined the
DNA content of HSC-3 cells treated with or without WGCG treatment
at 0 and 24 h. The flow-cytometric analysis revealed a significant
increase in the percentage of G1 phase cells as compared to control
cells (64.3±4.5 vs. 46.2±4.7%), and a significant decrease in the
percentage of G2/M phase cells as compared with control cells
(11.6±5.1 vs. 26.0±5.5%) at 24 h (Fig.
2A and B). These results showed that EGCG can arrest HSC-3
cells at the G1 checkpoint of the cell cycle.
EGCG induces apoptosis of HSC-3 cells
in vitro
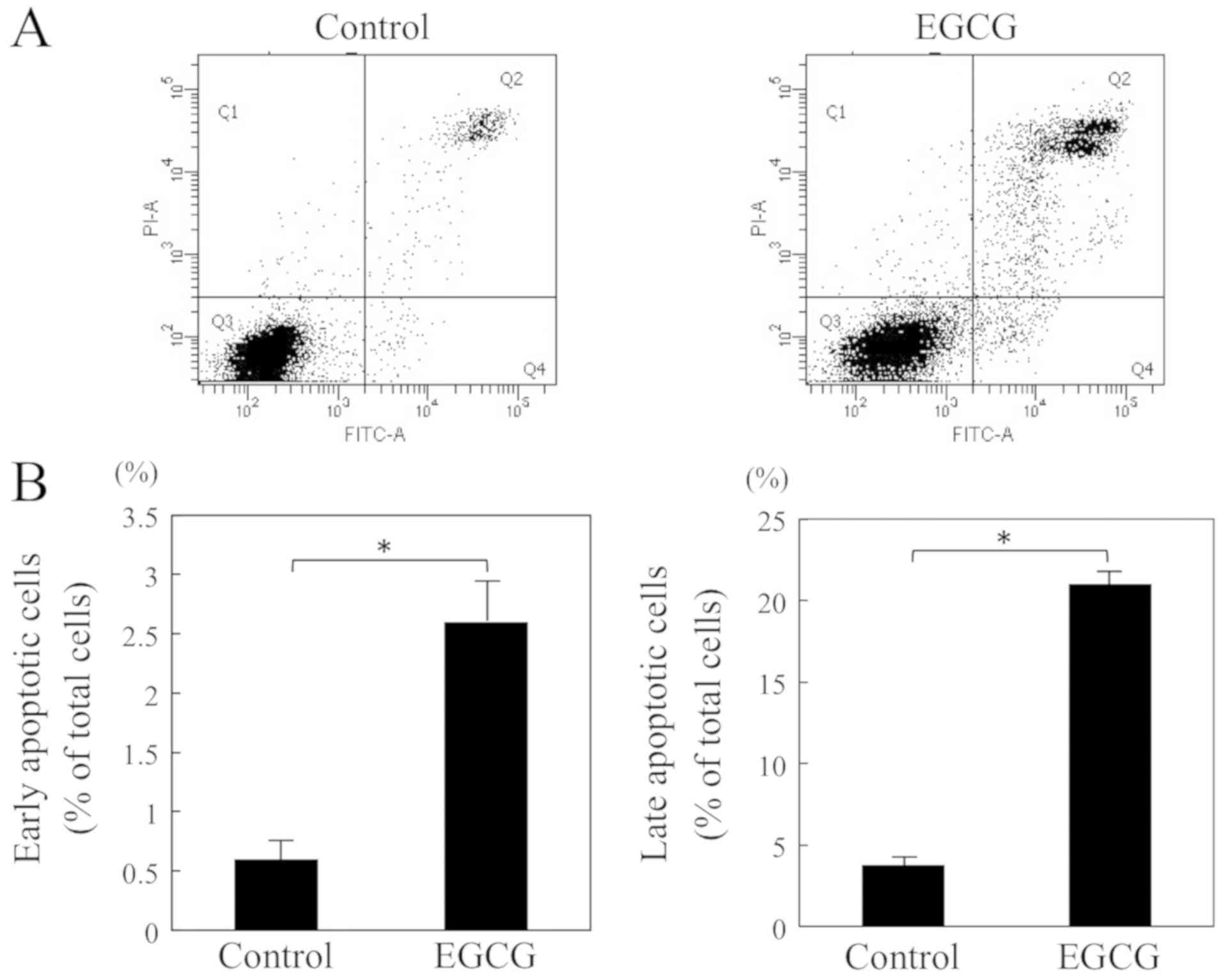
To further investigate the mechanism, we have
evaluated the effects of EGCG on three stages of apoptosis: The
early stage by Annexin V assay which estimates the expression of
phosphatidylserine on the outer leaflet of the cell membrane, the
middle stage by evaluation of the activities of caspase-3 and −7
(13,14), and the late stage by TUNEL assay
which detects DNA strand breaks. The cells were incubated with or
without EGCG for 6 h and then analyzed by flow cytometry with
Annexin V to determine the apoptotic rate. The treatment with EGCG
significantly increased the percentage of apoptotic cells as
compared to control cells (early apoptotic cells: 2.6±0.4 vs.
0.6±0.2% and late apoptotic cells: 21.0±1.2 vs. 3.8±0.6%) (Fig. 3A and B). We also assessed the
effects of EGCG on the activities of caspase-3 and −7. The
caspase-3 and −7 activities significantly increased after 12 h of
treatment with EGCG as compared to control cells (2,861.4±580.7 RFU
vs. 884.6±76.6 RFU) (Fig. 3C). The
caspase-3 and −7 inhibitor (Ac-DEVD-CHO) was used in this study,
and it significantly suppressed these activities. We also
investigated the effect of EGCG on apoptosis in vitro by the
TUNEL assay. The percentage of apoptotic cells of EGCG treatment
for 24 h was significantly greater than that in the control cell
group (2.8±1.1 vs. 0.3±0.5%) (Fig. 3D
and E). These results suggested that EGCG induces apoptosis in
HSC-3 cells.
EGCG suppresses tumor growth of HSC-3
cells in vivo
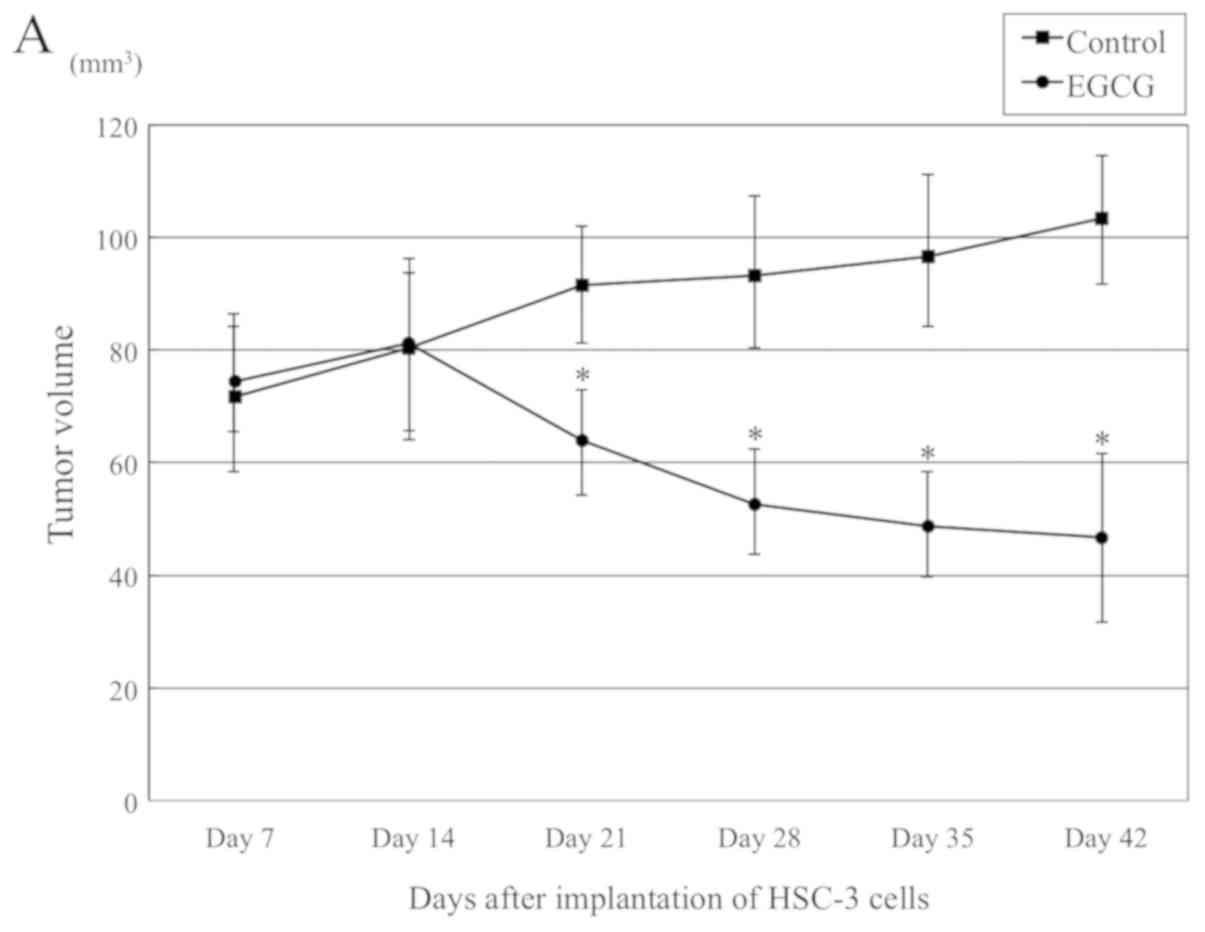
To confirm the in vitro findings above, we
examined the effects of EGCG on in vivo tumor growth. HSC-3
cells were implanted subcutaneously into the back of nude mice,
with monitoring for 6 weeks. Two weeks after HSC-3 cells
implantation, 75 mg/kg EGCG or saline was intraperitoneally
administered twice a week for 4 weeks. A significant difference was
observed from 1 week after the administration of EGCG. The
administration of EGCG for 4 weeks resulted in a 45.2% reduction in
tumor volume as compared with control animals (46.7±17.8 vs
103.4±12.4 mm3) (Fig.
4A-C). The weight of the mice did not decrease significantly,
and the average weight of the EGCG treatment group was 23.3±1.5 g,
almost equal to that of the control group (24.2±1.2 g) (Fig. 4D). In agreement with in
vitro data, EGCG significantly inhibited tumor growth in our
xenograft model.
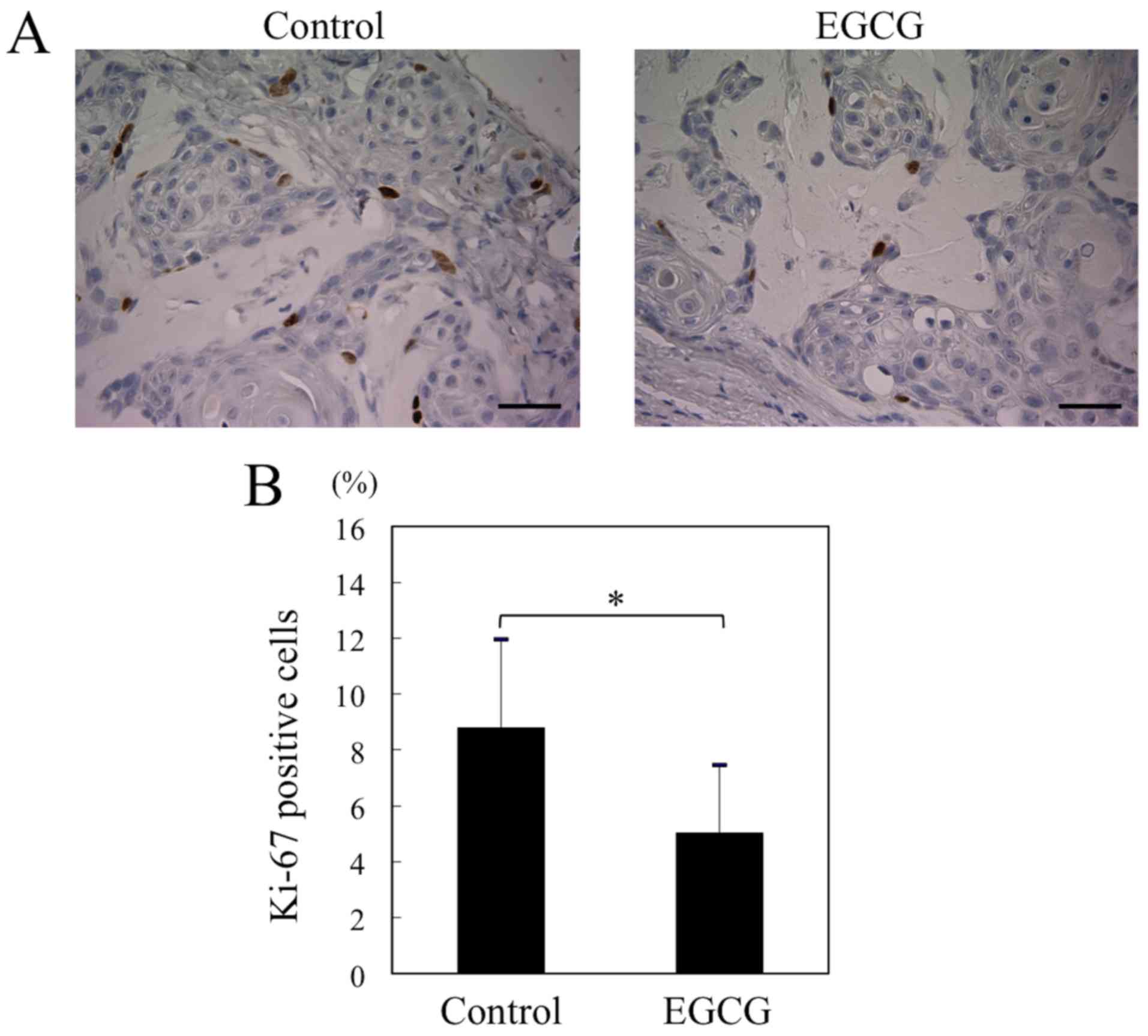
EGCG suppresses division of HSC-3
cells in vivo
Ki-67 is a 395 kDa nuclear antigen whose expression
is confined to the late G1, S, M and G2 phases, and its expression
is strictly associated with cell proliferation (15). We also evaluated the expression of
Ki-67 in tumor cells in vivo. There were significant
differences in mean Ki-67 expression between the EGCG treatment and
the control group (8.8±3.2 vs. 5.0±2.4%) (Fig. 5A and B). The results indicated an
inhibitory effect of EGCG on cell proliferation in the xenograft
tumors.
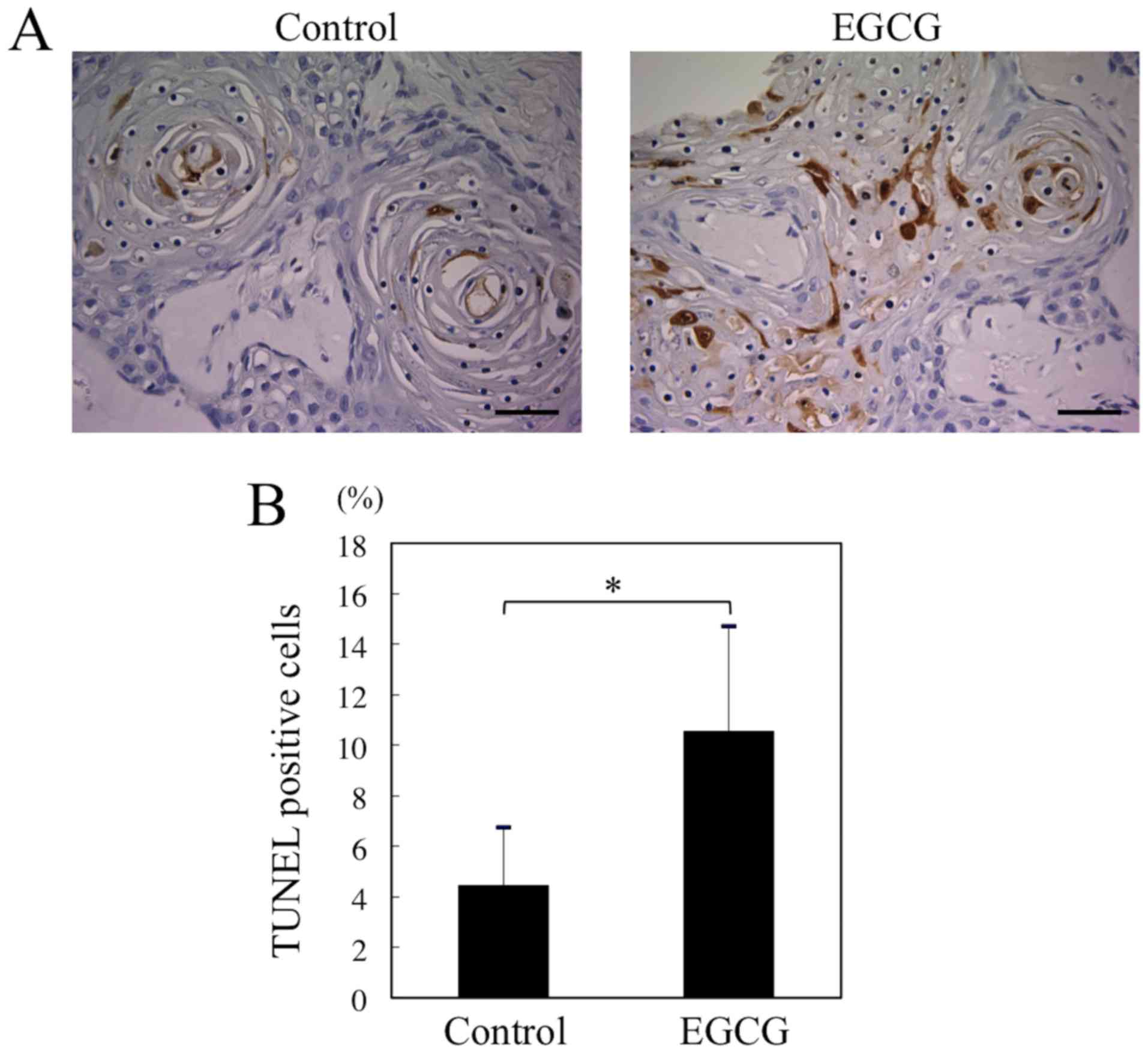
EGCG induces apoptosis of HSC-3 cells
in vivo
We investigated the effect of EGCG on apoptosis
in vivo by the TUNEL assay. The percentage of apoptotic
cells in the EGCG treatment group was significantly greater than
that in the control group (10.6±4.2 to 4.4±2.3%) (Fig. 6A and B). Thus, the EGCG treatment
can significantly induce apoptosis in the xenograft tumors.
Discussion
Oral cancer is one of the most common malignant
tumors in the head and neck region (5), and >90% of oral cancers are OSCC
(3). Patients with oral cancer
have a high mortality rate because of local invasion and distant
metastasis (5). The 5-year
survival rate of patients with early-stage disease is ~80%, and
this rate of late-stage disease is only ~20% (16). Because the most advanced oral
cancer is impossible to heal, blocking the process of
carcinogenesis is an important strategy for cancer management.
Green tea has been a popular beverage for many
centuries (7). The main
polyphenols in green tea are catechins, and the four main catechins
are EGCG (constitutes 59% of all catechins), EGC (19%), ECG
(13.6%), and EC (6.4%) (7).
Particularly, its major constituent, EGCG, has been demonstrated to
act on several key components of intracellular signaling pathways
associated with cell proliferation, differentiation, apoptosis,
inflammation, angiogenesis, and metastasis; however, these
molecular mechanisms are not completely characterized, and many
features have not been elucidated (17). We herein examined the possibility
that EGCG may have a therapeutic potential for human oral
cancer.
First, we examined the effectiveness of EGCG against
the cultured OSCC cell line, HSC-3. After exposure to EGCG, HSC-3
cells manifested suppression of cellular proliferation and were
arrested in the G1 phase of the cell cycle. Elattar and Virji have
demonstrated a significant inhibitory effect of EGCG on the growth
and proliferation of OSCC cells (SCC-25), where high doses of EGCG
exert an inhibitory effect on DNA synthesis (18). Masuda et al have reported
that EGCG inhibits cell growth and causes G1 arrest of the cell
cycle in two cell lines derived from human head and neck squamous
cell carcinoma (HNSCC): YCU-H891 and YCU-N861 (19). In the present study, these reported
effects of EGCG on OSCC and HNSCC could also be observed in HSC-3
cells in vitro. Apoptosis is a form of programmed cell death
and plays an important role in the regulation of cell homeostasis
of eukaryotes (20). Apoptosis can
proceed via the mitochondria-mediated (intrinsic) pathway
(including Bax, Bcl-2, and Bcl-XL proteins) and a death
receptor-mediated (extrinsic) pathway (including Fas/CD95 and FADD)
(20), and can be induced by
agents such as drugs used for chemotherapy (21). Masuda et al reported that
EGCG induces apoptosis in two cell lines of human HNSCC. Cell death
in both cell lines was found to be related to downregulation of the
antiapoptotic Bcl-XL and Bcl-2 proteins and an increase
in the amount of the proapoptotic Bax protein (19). Lin et al have demonstrated
dose- and time-dependent apoptosis of EGCG-treated HNSCC cells (SAS
and Cal-27) (22). Apoptosis is
induced through the expression of Fas/CD95. In line with increased
Fas/CD95 expression, EGCG inhibits STAT3 phosphorylation,
translocation to the nucleus, and leads to downregulation of the
target gene products of STAT3, such as Bcl-2, Mcl-1, VEGF and
cyclin D1 (22). Thus, EGCG can
induce apoptosis through effects on the mitochondria-mediated and
the death receptor-mediated pathway. Caspases are a family of
intracellular cysteine proteases and are known to perform an
important function in the initiation and execution of apoptosis
(23). The mitochondria-mediated
and death receptor-mediated pathways converge at the level of
caspase-3 activation (20),
whereas caspase-3 and −7 serve as effectors of apoptotic cells
(13,14). We evaluated the effect of EGCG on
caspase-3 and −7 activities, and the results suggested that the
proapoptotic effect of EGCG in HSC-3 cells is mediated by
activation of those caspases. We may need to verify the detailed
mechanism of EGCG's action on cell cycle arrest and apoptosis as
well as its effect on the molecular pathway. Nevertheless, our
results confirmed that EGCG significantly inhibits HSC-3 cell
proliferation by at least affecting the cell cycle and apoptosis
in vitro. In this study, we did not examine the toxicity of
EGCG on normal cells. Previous reports have shown that EGCG had no
effect on normal cells. Yamamoto et al reported that a high
dose of EGCG (~200 µM) caused reactive oxygen species production
and apoptosis only in oral SCC, but not in normal epithelial cells
in vitro (24). Chen et
al reported that tea polyphenols potently induced apoptotic
cell death and cell cycle arrest in tumor cells without affecting
the normal cell counterparts (25). Further research will be required to
evaluate the differential effect of EGCG on cancer cells and normal
cells.
To develop effective therapeutic strategies,
reliable experimental models are important. An in vivo
xenograft mouse model is one of the reliable experimental models
for the development of more effective treatments with evaluation of
cytotoxic effects (26). Second,
we examined the significance of EGCG effects on tumor growth by
means of the xenograft mouse model. Nude mice were
intraperitoneally injected with or without 75 mg/kg EGCG. Our in
vivo experiments indicated that, compared to the control group,
there was substantially diminished tumor growth in the EGCG-treated
group, with inhibition of cell division (Ki-67) and induction of
apoptosis (TUNEL assay). Many in vitro assays involving an
OSCC cell line have shown the effects of EGCG, but few researchers
have reported in vivo xenograft models (Table I) (27–29).
To our knowledge, this is the first report that documents
antiproliferative and proapoptotic effects of EGCG on oral cancer
cells in vivo. Although there was a difference in the
cancer-suppressive effects of EGCG between the in vitro and
in vivo experiments, this difference might be attributed to
the OSCC cell environment and concentration of EGCG.
 | Table I.Studies investing the effects of tea
contents on human oral malignancy in xenograft mouse model. |
Table I.
Studies investing the effects of tea
contents on human oral malignancy in xenograft mouse model.
|
|
| Administration of
tea extract |
|
|
|---|
|
|
|
|
|
|
|---|
| Author (year) | OSCC cell | Content | Rote | Dose and
duration | Results | (Refs.) |
|---|
| Chen et al
(2011) | SCC-9 (human tongue
SCC) | EGCG | Oral
administration | 10–20 mg/kg, every
day for 44 days | Inhibition of the
tumor growth | (27) |
| Chang et al
(2012) | SCC-4 (human tongue
SCC) | Black tea
extract | Oral
administration | 25, 50 mg/kg, every
day for 45 days | Inhibition of the
tumor growth | (28) |
| Hwang et al
(2013) | YD-10B (human oral
SCC) | EGCG | Intra-peritoneal
injection | 20 mg/kg, every
other day for 28 days | Inhibition of the
tumor growth. Inhibition the phosphorylation of Src, CTTN, FAK
proteins. Inhibition of the expression of MT1-MMP, MMP-2,
MMP-9. | (29) |
| Present study
(2019) | HSC-3 (human oral
SCC) | EGCG | Intra-peritoneal
injection | 75 mg/kg, twice a
week for 28 days | Inhibition of the
tumor growth. Increase in apoptotic index, and decrease in
proliferation index. |
|
In animal experimental models, weight loss is the
marker of adverse effects or toxicity of a pharmacological agent.
In our study, the weight in the EGCG-treated group of mice was
almost equal to that of the control group. Thus, our study also
revealed that EGCG exerted a milder adverse effects, while it was
still able to induce apoptosis and cell cycle arrest in OSCC cells.
Due to its pharmacological properties and reduced adverse effects,
EGCG may be a promising agent and a novel approach to oral-cancer
therapy.
In this study, a time and dose discrepancy in the
cancer-suppressive effects of EGCG was observed between the in
vitro and in vivo experiments. We have used a range of
EGCG concentrations from 25 to 100 µM for up to 3 days in the in
vitro experiment, whereas 75 mg/kg EGCG was intraperitoneally
administered twice a week for up to 28 days in the in vivo
experiment. Generally, the concentration of EGCG used in cell
culture experiments (20–100 µM) is much higher than the plasma and
tissue concentrations observed in the cancer-prevention studies
conducted in mice (usually <0.5 µM) (30,31).
Although the in vivo concentration in this study was low,
the prolonged exposure might have produced significant effects.
This seems consistent with our observation in vitro that
treatment of cancer cells with EGCG for a prolonged period (3 days)
increases the extent of inhibition of cell viability compared to
treatment for 1 or 2 days. The concentration and time of onset of
effects may be clinically relevant. There is a possibility that the
difference in the environment of the tumor cells might also have
played a role. The difficulty in studying the biological activities
of EGCG lies with correlating the in vitro biological
effects with the proposed mechanisms based on the in vivo
study results (31). Thus, the
inhibitory activities of EGCG against carcinogenesis should be
demonstrated both in in vitro and in vivo
experiments.
It may also not be obvious whether the information
obtained from cell lines with high EGCG concentrations can be
extrapolated to cancer prevention in humans. However, the
anticancer effect of EGCG has been demonstrated in epidemiological
studies and in clinical trials (1,9,32).
One epidemiological study has shown a protective effect of green
tea against malignant tumors. In a study of 8552 Japanese adults,
daily drinking >10 cups of green tea per day decreased the risk
and delayed the onset of cancers compared to those who drank less
than 3 cups per day (8). One
clinical trial showed that green tea extract (2,000-2,500 mg/day)
administration to smokers for 4 weeks reduces DNA damage in oral
keratinocytes. In addition, cell growth is inhibited, the
percentage of cells in the S phase decreases, cells accumulate in
the G1 phase, and apoptotic markers are upregulated. Therefore,
regular consumption of green tea or administration of green tea
extracts could be beneficial for the prevention of oral cancer in
humans.
In conclusion, we demonstrated that EGCG induces
cell cycle arrest and apoptosis in human OSCC cells, resulting in
antiproliferative effects in vitro and in vivo: In a
mouse model, significant growth inhibition of the OSCC tumor was
observed in EGCG-treated mice without a loss of body weight. Thus,
we believe that EGCG is a potential anticancer agent for OSCC
therapy. The mechanisms underlying the anticancer effects of EGCG
seem to be complex. Further research, especially molecular and
clinical, is needed to elucidate the usefulness of EGCG for
oral-cancer therapy.
Acknowledgements
Not applicable.
Funding
This study was supported by a Grant-in-Aid for
Scientific Research (grant nos. JP19791515, JP22890074, JP2472196
and JP15K11240) from the Japan Society for the Promotion of
Science, Japan.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HYoshim made substantial contributions to conception
and design, and acquisition of data and drafting the manuscript.
HYoshid made substantial contributions to conception and design and
acquisition of data. SM, TR and KO performed the experiments. MO
and SY analyzed the data. TK, MK and KS wrote the manuscript and
also made substantial contributions to conception of data, analysis
of data and critically revising the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were permitted by the Animal
Ethics Committee of the University of Fukui (approval no.
27110).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Iriti M and Varoni EM: Chemopreventive
potential of flavonoids in oral squamous cell carcinoma in human
studies. Nutrients. 5:2564–2576. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rivera C: Essentials of oral cancer. Int J
Clin Exp Pathol. 8:11884–11894. 2015.PubMed/NCBI
|
|
4
|
Schwartz JL, Baker V, Larios E and Chung
FL: Molecular and cellular effects of green tea on oral cells of
smokers: A pilot study. Mol Nutr Food Res. 49:43–51. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang B, Zhang S, Yue K and Wang XD: The
recurrence and survival of oral squamous cell carcinoma: A report
of 275 cases. Chin J Cancer. 32:614–618. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chu C, Deng J, Man Y and Qu Y: Green tea
extracts epigallocatechin-3-gallate for different treatments.
Biomed Res Int. 2017:56156472017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Narotzki B, Reznick AZ, Aizenbud D and
Levy Y: Green tea: A promising natural product in oral health. Arch
Oral Biol. 57:429–435. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Imai K, Suga K and Nakachi K:
Cancer-preventive effects of drinking green tea among a Japanese
population. Prev Med. 26:769–775. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lecumberri E, Dupertuis YM, Miralbell R
and Pichard C: Green tea polyphenol epigallocatechin-3-gallate
(EGCG) as adjuvant in cancer therapy. Clin Nutr. 32:894–903. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jung YD, Kim MS, Shin BA, Chay KO, Ahn BW,
Liu W, Bucana CD, Gallick GE and Ellis LM: EGCG, a major component
of green tea, inhibits tumour growth by inhibiting VEGF induction
in human colon carcinoma cells. Br J Cancer. 84:844–850. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Urusova DV, Shim JH, Kim DJ, Jung SK,
Zykova TA, Carper A, Bode AM and Dong Z: Epigallocatechin-gallate
suppresses tumorigenesis by directly targeting Pin1. Cancer Prev
Res (Phila). 4:1366–1377. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshida H, Yoshimura H, Matsuda S, Ryoke
T, Kiyoshima T, Kobayashi M and Sano K: Effects of peritumoral
bevacizumab injection against oral squamous cell carcinoma in a
nude mouse xenograft model: A preliminary study. Oncol Lett.
15:8627–8634. 2018.PubMed/NCBI
|
|
13
|
Kim CS, Cho SH, Chun HS, Lee SY, Endou H,
Kanai Y and Kim DK: BCH, an inhibitor of system L amino acid
transporters, induces apoptosis in cancer cells. Biol Pharm Bull.
31:1096–1100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moon SM, Yun SJ, Kook JK, Kim HJ, Choi MS,
Park BR, Kim SG, Kim BO, Lee SY, Ahn H, et al: Anticancer activity
of Saussurea lappa extract by apoptotic pathway in KB human oral
cancer cells. Pharm Biol. 51:1372–1377. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Scholzen T and Gerdes J: The Ki-67
protein: From the known and the unknown. J Cell Physiol.
182:311–322. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sinevici N and O'Sullivan J: Oral cancer:
Deregulated molecular events and their use as biomarkers. Oral
Oncol. 61:12–18. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singh BN, Shankar S and Srivastava RK:
Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms,
perspectives and clinical applications. Biochem Pharmacol.
82:1807–1821. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Elattar TM and Virji AS: Effect of tea
polyphenols on growth of oral squamous carcinoma cells in vitro.
Anticancer Res. 20:3459–3465. 2000.PubMed/NCBI
|
|
19
|
Masuda M, Suzui M and Weinstein IB:
Effects of epigallocatechin-3-gallate on growth, epidermal growth
factor receptor signaling pathways, gene expression, and
chemosensitivity in human head and neck squamous cell carcinoma
cell lines. Clin Cancer Res. 7:4220–4229. 2001.PubMed/NCBI
|
|
20
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaufmann SH and Earnshaw WC: Induction of
apoptosis by cancer chemotherapy. Exp Cell Res. 256:42–49. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin HY, Hou SC, Chen SC, Kao MC, Yu CC,
Funayama S, Ho CT and Way TD: (−)-Epigallocatechin gallate induces
Fas/CD95-mediated apoptosis through inhibiting constitutive and
IL-6-induced JAK/STAT3 signaling in head and neck squamous cell
carcinoma cells. J Agric Food Chem. 60:2480–2489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamamoto T, Hsu S, Lewis J, Wataha J,
Dickinson D, Singh B, Bollag WB, Lockwood P, Ueta E, Osaki T and
Schuster G: Green tea polyphenol causes differential oxidative
environments in tumor versus normal epithelial cells. J Pharmacol
Exp Ther. 307:230–236. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen D, Milacic V, Chen MS, Wan SB, Lam
WH, Huo C, Landis-Piwowar KR, Cui QC, Wali A, Chan TH and Dou QP:
Tea polyphenols, their biological effects and potential molecular
targets. Histol Histopathol. 23:487–496. 2008.PubMed/NCBI
|
|
26
|
Mery B, Rancoule C, Guy JB, Espenel S,
Wozny AS, Battiston-Montagne P, Ardail D, Beuve M, Alphonse G,
Rodriguez-Lafrasse C and Magné N: Preclinical models in HNSCC: A
comprehensive review. Oral Oncol. 65:51–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen PN, Chu SC, Kuo WH, Chou MY, Lin JK
and Hsieh YS: Epigallocatechin-3 gallate inhibits invasion,
epithelial-mesenchymal transition, and tumor growth in oral cancer
cells. J Agric Food Chem. 59:3836–3844. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang YC, Chen PN, Chu SC, Lin CY, Kuo WH
and Hsieh YS: Black tea polyphenols reverse
epithelial-to-mesenchymal transition and suppress cancer invasion
and proteases in human oral cancer cells. J Agric Food Chem.
60:8395–8403. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hwang YS, Park KK and Chung WY:
Epigallocatechin-3 gallate inhibits cancer invasion by repressing
functional invadopodia formation in oral squamous cell carcinoma.
Eur J Pharmacol. 715:286–295. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang CS and Wang X: Green tea and cancer
prevention. Nutr Cancer. 62:931–937. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang CS, Sang S, Lambert JD and Lee MJ:
Bioavailability issues in studying the health effects of plant
polyphenolic compounds. Mol Nutr Food Res. 52 (Suppl 1):S139–S151.
2008.PubMed/NCBI
|
|
32
|
Sheth SH, Johnson DE, Kensler TW and
Bauman JE: Chemoprevention targets for tobacco-related head and
neck cancer: Past lessons and future directions. Oral Oncol.
51:557–564. 2015. View Article : Google Scholar : PubMed/NCBI
|