Introduction
In vitro fertilization (IVF) is a first-line
treatment for female infertility. It involves egg collection,
fertilization of the eggs with sperm in the laboratory, followed by
the transfer of one or more viable embryos to the uterus, with the
hope of successful pregnancy. In 2012, an estimated 5 million
children were born from IVF (1).
During IVF, it is necessary to collect and fertilize numerous eggs
in order to increase the number of high-quality embryos available
for transfer, and multi-follicular development is induced by
hormonal stimulation. In women without ovarian function,
17β-estradiol (E2) and progesterone are the two hormones that need
to be provided to achieve a receptive endometrial environment
(2). Clinical evidence has
suggested that endometrial receptivity during controlled ovarian
hyperstimulation is reduced, which may increase the circulating
levels of E2 to supraphysiological levels (3,4).
Supraphysiological serum levels of E2 may exert a negative effect
on endometrial receptivity, but without affecting embryo quality
(5–7). Using a step-down protocol to decrease
the plasma levels of E2 prior to embryo transfer may improve the
implantation rate (8).
Nevertheless, the mechanisms underlying the effects of
supraphysiological levels of E2 on endometrial receptivity remain
unclear. Proteomic studies have revealed the differences between
receptive and non-receptive endometria during the implantation
window (9); however, the effect of
E2 is still poorly understood (8).
The roles of aquaporins (AQPs) in the reproductive
system have previously been examined (10–12).
AQPs serve functional and distinctive collaborative roles in
regulating the amount of water in the mammalian uterus and oviduct
(13,14). It has previously been demonstrated
that in the human endometrium, AQP2 is expressed in a
menstruation-dependent manner, suggesting that AQP2 may serve
physiological roles in uterine receptivity (15). In addition, AQP2 expression in
human endometrium is positively correlated with serum E2 levels,
suggesting that E2 may regulate AQP2 expression (15,16).
A previous study strongly suggested that AQP2 may be regulated by
E2 via an estrogen receptor-estrogen response element (ER-ERE) in
endometrial carcinoma cells (17).
However, whether E2 regulates AQP2 expression in normal human
endometrium has yet to be determined. In addition, the role of AQP2
in embryo implantation is largely unknown. Therefore, the present
study aimed to investigate whether E2 regulates AQP2 expression in
normal human endometrial tissues, and to determine the role of AQP2
in embryo implantation.
Patients and methods
Patients and sample collection
Normal endometrial samples were collected from women
seeking IVF and embryo transfer (n=113; median age, 32 years; age
range, 25–38 years) at the Women's Hospital (Hangzhou, China).
Women with endometrial abnormalities were excluded from this study.
Endometrial samples were collected from women undergoing
hysterectomies between August 2015 and February 2016 on days 6–15
(proliferative phase, n=38) or 16–28 (secretory phase, n=75) of the
menstrual cycle. None of the women were treated with cycle-altering
medication. Shortly after collection, the endometrial samples were
cut with a scalpel into pieces (size, 1.0×0.5 cm), and either snap
frozen in liquid nitrogen and stored at −70°C for the extraction of
protein and mRNA, or fixed with 4% paraformaldehyde for 24 h at 4°C
prior to pathological examination. According to Noyes pathological
diagnostic criteria (18),
endometrial samples were categorized as early-mid proliferative
(days 5–10), late proliferative (days 11–14), early secretory (days
15–18), mid-secretory (days 19–24) and late secretory (days 25–28)
phase groups following hematoxylin and eosin staining. Briefly, the
sections were prepared at room temperature and were stained with
hematoxylin (0.5%) and eosin (0.5%) for 25–28 min at room
temperature, after which, they were examined using a light
microscope (Olympus Corporation, Tokyo, Japan).
The present study was approved by the Ethics
Committee of the School of Medicine of Zhejiang University
(Hangzhou, China). Written informed consent was obtained from each
patient prior to tissue collection and enrolment in the present
study.
Tissue culture
Endometrial explant culture was performed as
previously described (19).
Briefly, endometrial explants in the proliferative phase from
hysterectomy specimens were minced using a scalpel (2–3 mm), placed
in a 12-well dish with the luminal epithelial surface facing
upward, and cultured in RPMI-1640 containing 10% charcoal-stripped
fetal bovine serum (FBS; Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA) at 37°C in a humidified environment containing 5%
CO2. After 24 h, the tissues (n=5) were exposed to
ethanol (control; same volume as in the E2 groups) or various
concentrations of E2 (10−9 to 10−5 M; cat.
no. E2257; Sigma-Aldrich Merck KGaA, Darmstadt, Germany) for an
additional 24 h at 4°C. Rat kidney tissues were collected from a
4-week old male Sprague-Dawley rat (weight, 200 g; Shanghai SLAC
Laboratory Animal Co., Ltd., Shanghai, China) and were used as
positive controls for AQP expression, as previously described
(20). The rat was routinely
maintained and received no prior treatment; kidney tissues were
cultured in the same manner as endometrial tissues.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following treatment, endometrial tissues were either
snap frozen in liquid nitrogen and stored at −70°C for the
extraction of RNA and protein, or processed to obtain 4-µm
sections. For RNA extraction, endometrial tissues (50–100 mg) were
homogenized in 1 ml RNAiso Plus (Sangon Biotech Co., Ltd.,
Shanghai, China). The supernatant was collected, mixed with 200 µl
trichloromethane and agitated for 30 sec. After centrifugation at
240 × g for 15 min at 4°C, the supernatant was collected and mixed
with an equal volume of isopropyl alcohol. The mixture was
centrifuged at 240 × g for 10 min at 4°C, and the precipitate was
dissolved in 75% ethanol. Total RNA was extracted after being
centrifuged at 120 × g for 5 min at 4°C, and was dissolved in DEPC
water. Total RNA was processed using DNase to remove DNA
contamination. Subsequently, cDNA was synthesized using the Reverse
Transcriptase kit (Takara Bio, Inc., Otsu, Japan); the RNA was
heated to 37°C for 60 min, followed by incubation at 95°C for 5
min. Eventually, the cDNA was stored at −20°C for further analysis.
RT-qPCR was conducted on an Icycler™ Optical Module using the iQ™
SYBR® Green Supermix (both Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The primers used were as follows: AQP2 forward,
5′-TGGGCCATATGTGCTATGGAGA-3′ and reverse,
5′-AAGGACACTCAGGTGCCAGGA-3′ (amplicon size, 142bp); and as an
internal reference, GAPDH forward, 5′-CAGGGCTGCTTTTAACTCTGG-3′ and
reverse, 5′-TGGGTGGAATCATATTGGAACA-3′ (amplicon size, 102bp). For
qPCR, the following thermocycling program was adopted: Initial
denaturation at 94°C for 5 min; 30 cycles of denaturation at 94°C
for 30 sec, annealing at 65°C for 30 sec and extension at 72°C for
30 sec, and a final extension step at 72°C for 10 min. Data were
analyzed using the comparative quantification cycle (Cq) method
(21,22): Relative
expression=2−ΔΔCq, where ΔΔCq=ΔCq (treated group)-ΔCq
(control group), and ΔCq=Cq (sample)-Cq (internal control).
Immunofluorescence
Endometrial samples (1.0×0.5 cm) were fixed as
aforementioned, sectioned at 4 µm, permeabilized with Triton-X,
blocked with 1% bovine serum albumin (Hyclone; GE Healthcare Life
Sciences) at 4°C for 1 h and incubated with goat anti-AQP2 primary
antibody (1:100; cat. no. sc-9882; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) at 4°C overnight. The samples were subsequently
incubated with Alexa Fluor® 488 rabbit anti-goat
immunoglobulin G (IgG) secondary antibodies (1:200; cat. no.
A27012; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) for 2 h at room temperature. No counterstain was used. The
slides were analyzed under a Zeiss LSM 510Meta laser scanning
confocal microscope (Carl Zeiss AG, Oberkochen, Germany) or an
Olympus BX51TF fluorescence microscope (Olympus Corporation).
Western blot analysis
Western blotting was performed as described
previously, without modification (15). Briefly, the proteins were separated
by 8% SDS-PAGE and were transferred onto nitrocellulose membranes.
The membranes were blocked in skimmed milk at 4°C for 1 h and were
exposed to goat-anti AQP2 (1:500; cat. no. sc-9882; Santa Cruz
Biotechnology, Inc.) and goat-anti β-actin (1:500; cat no. sc-1616;
Santa Cruz Biotechnology, Inc.) polyclonal antibodies,
respectively, at 4°C overnight. Subsequently, membranes were
incubated with horseradish peroxidase-labeled rabbit anti-goat IgG
(1:2,000 for AQP2 and 1:5,000 for β-actin; cat. no. ZB-2306,
OriGene Technologies, Inc., Beijing, China) for 1 h at room
temperature with agitation. Visualization was performed with an
enhanced chemiluminescence detection reagent (Santa Cruz
Biotechnology, Inc.). The protein bands were analyzed with Quantity
One software 3.1 (Bio-Rad Laboratories, Inc.). β-actin was used as
a loading control.
AQP2 silencing by small interfering
RNA (siRNA) technology
siRNA sequences against AQP2 were generated using
the Silencer siRNA Construction kit (Ambion; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocols. The
siRNA sequences against AQP2 were as follows: Forward,
3′-GGUGGGUGGUAAGAGGGAATT-5′ and reverse,
3′-UUCCCUCUUACCACCCACCTG-5′. Control scrambled siRNAs were
purchased from Ambion (cat. no. AM4611; Thermo Fisher Scientific,
Inc.). Ishikawa cells (cat. no. 99040201) were inoculated at 100
cells/cm2 in minimum essential medium (MEM; both
Sigma-Aldrich; Merck KGaA) supplemented with 2 mM glutamine, 1%
non-essential amino acids and 5% FBS (Hyclone; GE Healthcare Life
Sciences) at 37°C in an atmosphere containing 5% CO2.
After reaching 70–80% confluence, the cells were transfected with
siRNA duplexes for 24 h using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. A final concentration of 5 nM AQP2 siRNA
was determined to maximally suppress target RNA expression. The
same concentration was used for scrambled siRNA transfection.
Evaluation of JAr spheroid attachment
onto Ishikawa cells
Attachment of the choriocarcinoma cell line, Jar
(HTB-144; American Type Culture Collection, Manassas, VA, USA) to
endometrial cells (Ishikawa) was quantified according to an
adhesion assay, as previously described (23–25).
Briefly, a suspension of 6×105 JAr cells in 6 ml
RPMI-1640 medium supplemented with 10% FBS was incubated at 37°C on
a gyratory shaker at 110 rpm, in order to generate multicellular
spheroids 3 days following culture initiation. Ishikawa cells were
inoculated at 100 cells/cm2 in 12-well plates containing
MEM supplemented with 2 mM glutamine, 1% non-essential amino acids
and 5% FBS (Hyclone; GE Healthcare Life Sciences) at 37°C in an
atmosphere containing 5% CO2. At 40% confluence,
Ishikawa cells were transfected with scrambled siRNA or siRNA (6
nM) duplexes targeting AQP2 for 24 h, as mentioned, and were then
treated with or without E2 (10−7 M) for 24 h at 37°C in
an atmosphere containing 5% CO2. Subsequently, spheroids
(60–200 µm) were transferred onto a confluent monolayer of Ishikawa
cells. The cultures were maintained in culture medium (RPMI-1640
medium with 10% FBS) for 1 h at 37°C in a 5% CO2
incubator. After 1 h, spheroid adhesion to the endometrial
monolayer was quantified by centrifugation of the 12-well plates
with the cell-spheroid surface facing down at 10 × g for 10 min at
4°C. The unattached spheroids were removed by centrifugation.
Attached spheroids were visualized under an anatomical microscope
(magnification, ×100) and were counted under a phase contrast
microscope (magnification, ×400). Results were expressed as a
percentage of the total number of spheroids (attachment rate
(%)=number of attached spheroids/total spheroids) × 100.
Statistical analysis
All experiments were performed three times. All data
were normally distributed and expressed as the means ± standard
error of the mean. One-way analysis of variance followed by a
Tukey's post hoc test was used to assess differences among groups.
SPSS 11.0 for Windows (SPSS, Inc., Chicago, IL, USA) was used for
statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Menstrual cycle-dependent expression
of AQP2 in human endometrium
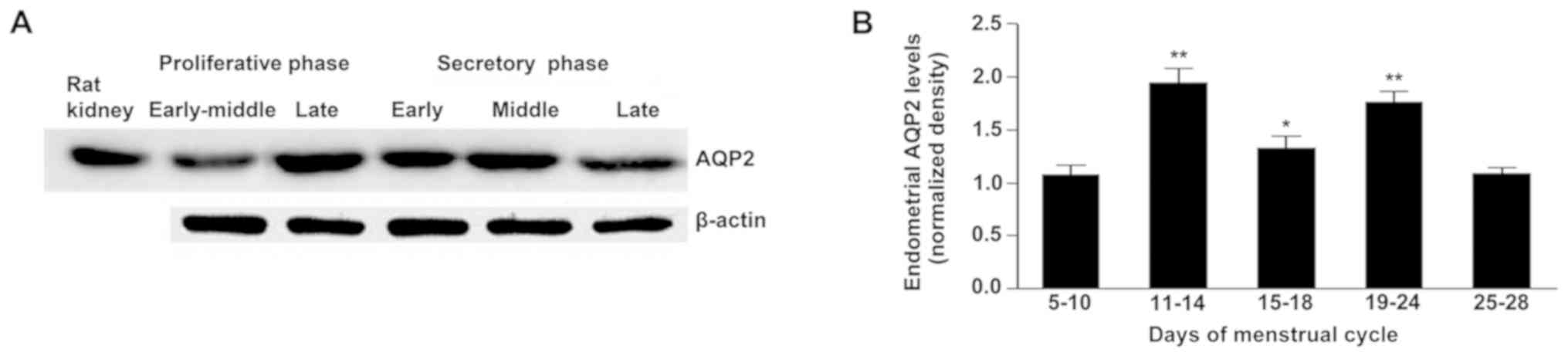
AQP2 mRNA and protein were expressed in human
endometrium, in a menstrual cycle-dependent manner (Fig. 1A). The lowest levels of AQP2
expression were observed in the early-mid proliferative and late
secretory phases (Fig. 1A and B).
The highest AQP2 expression levels were obtained in the late
proliferative and mid-secretory phases (Fig. 1A and B). E2 levels are altered
throughout the menstrual cycle (with a peak at day 12–13) (26) and these results suggested that AQP2
expression in the human endometrium may be E2-dependent.
Treatment with 10−7M E2
increases AQP2 expression in cultured human endometrium
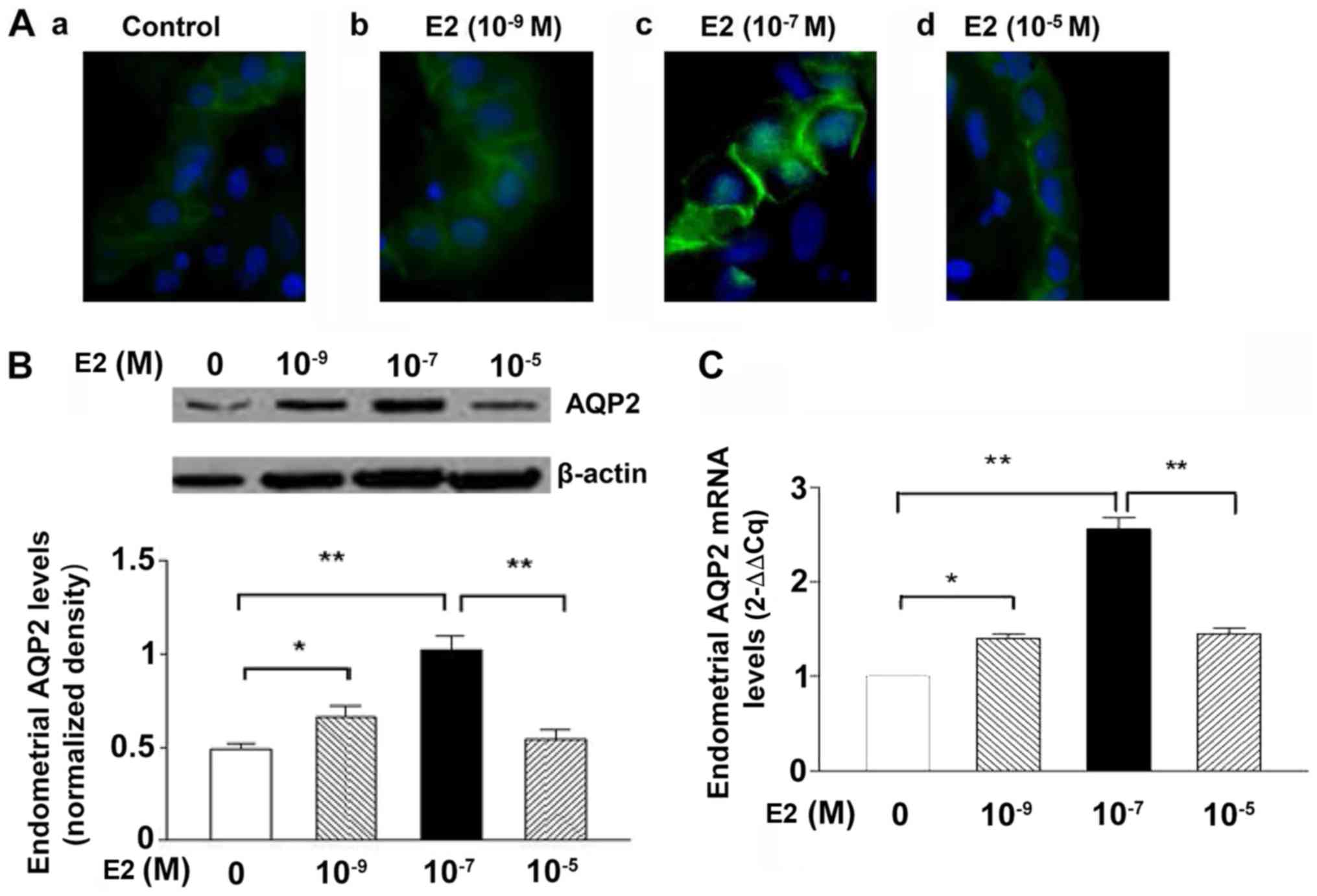
Immunofluorescence revealed that AQP2 was mainly
located in the glandular and luminal epithelia (Fig. 2Aa-d), in accordance with our
previous report (15). Stronger
immunofluorescence signals for AQP2 were observed in the
10−7 M E2 group (Fig.
2Ac); however, treatment with 10−5 M E2 notably
decreased the fluorescence of AQP2 (Fig. 2Ad) compared with 10−7 M
E2 (Fig. 2Ac). Western blotting
(Fig. 2B) and RT-qPCR (Fig. 2C) analyses revealed that
endometrial tissues in the 10−9 and 10−7 M
E2-treated groups exhibited significantly higher expression levels
of AQP2 compared with in the control group. AQP2 expression levels
were significantly reduced in response to treatment
with10−5 M E2 compared within response to
10−7 M E2 (Fig. 2B and
C). Therefore, 10−7 M E2 was selected for subsequent
experimentation to assess the effects of E2 on the endometrium.
Effects of AQP2 RNAi on Ishikawa
cells
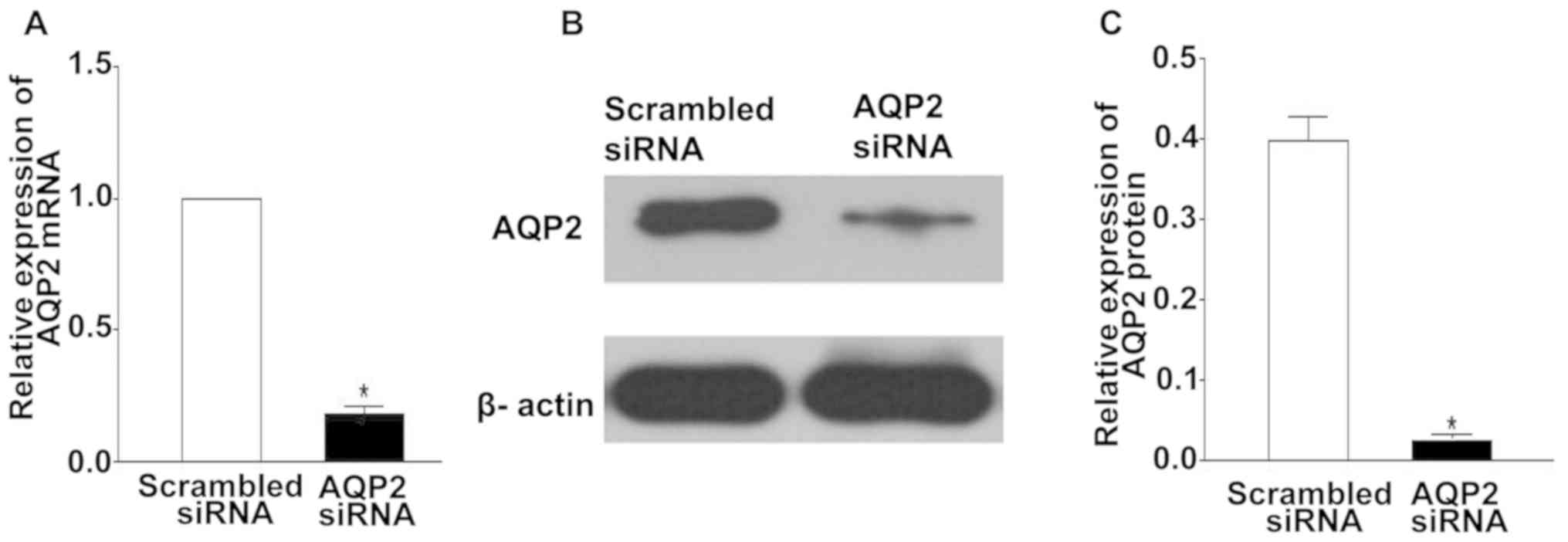
To confirm the silencing efficiency of AQP2 siRNA by
RT-qPCR and western blotting, Ishikawa cells were harvested 24 h
post-transfection with scrambled or AQP2 siRNA duplexes. Compared
with in the scrambled siRNA group, Ishikawa cells transfected with
AQP2 siRNA exhibited significant reductions in the mRNA and protein
expression levels of AQP2 [~81.0±2.1% (P<0.05) and ~72.0±1.9%
(P<0.05), respectively] (Fig.
3).
AQP2 siRNA reduces spheroid
attachment
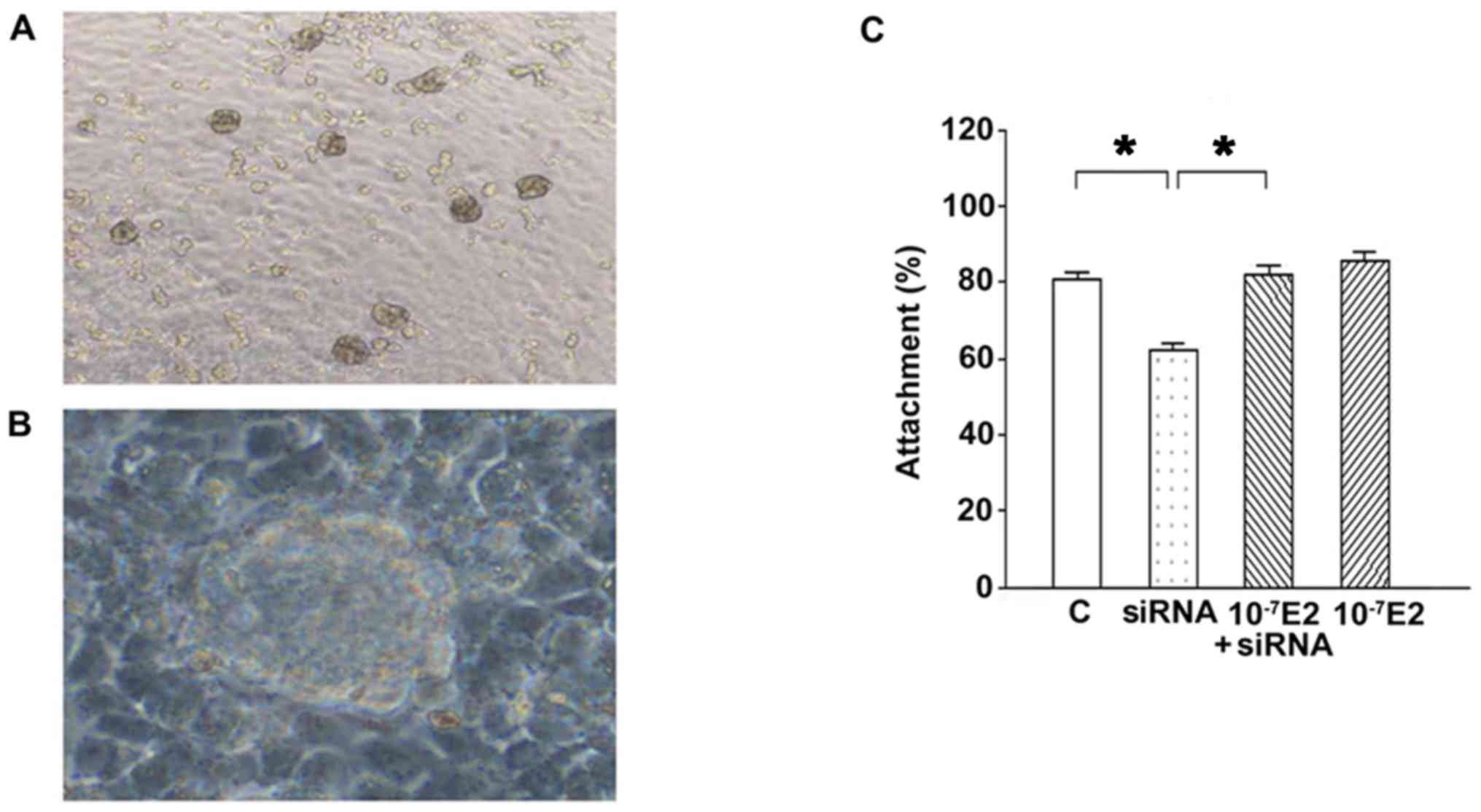
JAr spheroids at a diameter of 60–200 µm were used
to assess JAr cell attachment onto endometrial Ishikawa cells
(Fig. 4A). As presented in
Fig. 4B, knockdown of AQP2 by
siRNA resulted in a decreased attachment rate in the in
vitro attachment model. Attachment rates were: 80.6±2.2,
62.4±1.5, 82.1±2.2 and 85.4±2.4% in the scrambled siRNA (control),
AQP2 siRNA, 10−7 M E2 + AQP2 siRNA, and 10−7
M E2 groups, respectively. There were no differences between the
10−7 E2 and 10−7 E2 + siRNA groups, between
the 10−7 E2 and control groups, and between the
10−7 E2 + siRNA groups and control group (all
P>0.05); however, a significant difference was detected between
the siRNA and 10−7 E2 groups, between the siRNA and
10−7 E2 + siRNA groups, and between the siRNA and
control groups (all P<0.05). These results revealed that
knockdown of AQP2 by siRNA significantly decreased JAr spheroid
attachment compared with in the control group; however, this effect
was significantly reversed when AQP2 siRNA-transfected cells were
treated with 10−7 M E2 (Fig. 4C). These results indicated that the
expression of AQP2 may be associated with spheroid attachment and
E2 could improve spheroid attachment since significant differences
were observed between the siRNA and control groups, and between the
siRNA and 10−7 E2 + siRNA groups.
Discussion
E2 and AQP2 have been associated with endometrial
receptivity (5–7,10–12),
and E2 can directly regulate AQP2 expression in endometrial cancer
cells (15). Therefore, the
present study aimed to assess the role of AQP2 in embryo
implantation. The results revealed that AQP2 expression in human
endometrium maybe mediated by E2. In the present study, abnormally
high E2 levels attenuated AQP2 expression, which may be a potential
cause of impaired uterine receptivity. These results may be
beneficial for investigations regarding women undergoing IVF.
The present study reported temporal alterations in
the expression levels of AQP2 throughout the menstrual cycle; AQP2
was observed to be upregulated in human endometrium during the
implantation window (mid-secretory phase), supporting the findings
of previous studies (15,16). These findings suggested that AQP2
may serve physiological roles in uterine receptivity and affect
embryo attachment, which is the initial step of implantation. This
hypothesis was tested in vitro by knocking down AQP2 via
siRNA in the JAr-Ishikawa cell attachment assay (23–25).
In the present study, the Ishikawa cell line was selected as a
model for receptive endometrium as these cells possess
well-reported receptivity and express various
implantation-associated molecules (27), as well as estrogen and progesterone
receptors (28). JAr is a
trophoblastic choriocarcinoma cell line, which exhibits
cytotrophoblastic characteristics (29) and is able to attach to Ishikawa
cells in vitro (30). The
present study demonstrated that knockdown of AQP2 by siRNA in
Ishikawa cells significantly suppressed embryo attachment; this
finding further supported the hypothesis that AQP2 serves a key
role in uterine receptivity and embryo implantation. Our previous
study reported an association between AQP2 and E2 in the human
endometrium (15). Additionally,
the present study revealed the regulatory effects of E2 on AQP2.
Specifically, E2 had an effect on the expression of AQP2, but not
on JAr attachment to Ishikawa cells compared with the control
group. Therefore, these results suggested that E2 may have an
indirect effect on endometrial receptivity. These findings in
healthy women were in accordance with those of a recent report,
which demonstrated that impaired endometrial receptivity in
patients following controlled ovarian stimulation is associated
with reduced AQP2 expression (31).
Estrogens serve crucial roles in endometrial
cellular functions, including cell homeostasis, proliferation,
differentiation and vascularization (32,33).
ERs are expressed in endometrial glands and the stroma, and genomic
experiments have demonstrated that estrogens mainly exert their
effects via putative ER-EREs in promoter regions (34). A putative ER-ERE was identified in
the promoter region of the AQP2 gene (17). Recently, Chai et al
(35) compared the effects of high
serum E2 levels on endometrial steroid receptors in
gonadotropin-stimulated and natural cycles. Analysis revealed that
ER expression is significantly reduced in stimulated cycles
(35). Therefore, it may be
inferred that AQP2 expression and function may be attenuated in the
presence of highly elevated serum E2 levels; however, further
investigation is required.
Studies have reported significantly lower
implantation and pregnancy rates in cycles with high serum E2
concentrations (3–7); however, high serum E2 levels in fresh
IVF cycles may not affect implantation and pregnancy rates in
subsequent frozen-thawed embryo transfer cycles (36). Therefore, reduced implantation may
be due to an adverse endometrial environment resulting from high
serum E2 levels and not by the direct action of E2, suggesting that
high serum E2 levels may be an indirect cause of reduced
implantation (25,31,37–39).
The endometrium undergoes a series of precisely regulated
alterations under highly elevated serum estrogen levels induced by
the application of gonadotrophin in IVF cycles compared with
natural cycles (40,41). These changes include impaired
development of the endometrial glands (42), advanced stromal development
(39), desynchrony between glands
and the stroma (36), and early
expression of pinopodes (43). The
present study revealed that AQP2 expression was decreased under
abnormally high E2 levels in human endometrium. This finding
suggested a novel molecular mechanism may be involved in reducing
endometrial receptivity by elevating serum E2 levels in assisted
reproductive technologies, such as IVF. Physiological E2 levels are
closer to 10−7 M; however, at 10−5 M, E2 may
reduce AQP2 expression via ERs (44), which may indicate a different
mechanism from that of 10−7 M E2 used in the present
study. Conversely, a previous study proposed that very high serum
E2 levels may not be detrimental to the clinical outcomes of IVF
within a small sample (45).
Further investigation into the underlying mechanism of impaired
endometrial receptivity induced by AQP2 is required.
In conclusion, the present study demonstrated that
AQP2 expression in human endometrium may be regulated by E2. The
in vitro co-culture model using JAr and Ishikawa cells
suggested a notable role of AQP2 in uterine receptivity and embryo
implantation. Additionally, abnormal AQP2 expression may be a
potential cause of impaired uterine receptivity. The results of the
present study provided information regarding the causes of
decreased uterine receptivity in the context of controlled ovarian
hyperstimulation. These findings may provide an important marker
(AQP2) for endometrial receptivity in women undergoing IVF. Further
studies on the effects of AQP2 expression or polymorphisms on
female fertility are required.
Acknowledgements
This paper has been presented at the 2016 National
Symposium on the Advancement of Obstetrics and Gynecology in
Integrative Chinese and Western Medicine and 2016 Thesis and
Abstracts of the First Jiangsu, Zhejiang, and Zhejiang Integrative
Obstetrics and Gynecology Summit Forum.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 30700273, 81200429
and 81270702), the Natural Science Foundation of Zhejiang Province
(grant no. LQ14H040003) and the Fundamental Research Funds for the
Central Universities (grant no. 2015FZA7010).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and RH designed the study. WH, XC and XH
conducted the experiments. YH and RH participated in data analysis.
RH and WH wrote the manuscript. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the School of Medicine of Zhejiang University. Written
informed consent was obtained from each subject prior to
participation in this study.
Patient consent for publication
Written informed consent was obtained from each
patient prior to tissue collection and enrolment in the present
study.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AQP2
|
aquaporin 2
|
|
Cq
|
quantification cycle
|
|
E2
|
17β-estradiol
|
|
ER
|
estrogen receptor
|
|
ER-ERE
|
estrogen receptor-estrogen response
element
|
|
IVF
|
in vitro fertilization
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
References
|
1
|
Adamson GD, Tabangin M, Macaluso M and
Mouzon J: The number of babies born globaylly after treatment with
the assisted reproductive technologies (ART). Fertil Steril.
100:S422013. View Article : Google Scholar
|
|
2
|
Paulson RJ, Hatch IE, Lobo RA and Sauer
MV: Cumulative conception and live birth rates after oocyte
donation: Implications regarding endometrial receptivity. Hum
Reprod. 12:835–839. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Absalan F, Ghannadi A and Kazerooni M:
Reproductive outcome following thawed embryo transfer in management
of ovarian hyperstimulation syndrome. J Reprod Infertil.
14:133–137. 2013.PubMed/NCBI
|
|
4
|
Li MQ and Jin LP: Ovarian stimulation for
in vitro fertilization alters the protein profile expression in
endometrial secretion. Int J Clin Exp Pathol. 6:1964–1971.
2013.PubMed/NCBI
|
|
5
|
Davar R, Janati S, Mohseni F, Khabazkhoob
M and Asgari S: A comparison of the effects of transdermal
estradiol and estradiol valerate on endometrial receptivity in
frozen-thawed embryo transfer cycles: A randomized clinical trial.
J Reprod Infertil. 17:97–103. 2016.PubMed/NCBI
|
|
6
|
Kara M, Kutlu T, Sofuoglu K, Devranoglu B
and Cetinkaya T: Association between serum estradiol level on the
hCG administration day and IVF-ICSI outcome. Iran J Reprod Med.
10:53–58. 2012.PubMed/NCBI
|
|
7
|
Arslan M, Bocca S, Arslan EO, Duran HE,
Stadtmauer L and Oehninger S: Cumulative exposure to high estradiol
levels during the follicular phase of IVF cycles negatively affects
implantation. J Assist Reprod Genet. 24:111–117. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ullah K, Rahman TU, Pan HT, Guo MX, Dong
XY, Liu J, Jin LY, Cheng Y, Ke ZH, Ren J, et al: Serum estradiol
levels in controlled ovarian stimulation directly affect the
endometrium. J Mol Endocrinol. 59:105–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dominguez F, Garrido-Gomez T, Lopez JA,
Camafeita E, Quinonero A, Pellicer A and Simón C: Proteomic
analysis of the human receptive versus non-receptive endometrium
using differential in-gel electrophoresis and MALDI-MS unveils
stathmin 1 and annexin A2 as differentially regulated. Hum Reprod.
24:2607–2617. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang HF, He RH, Sun CC, Zhang Y, Meng QX
and Ma YY: Function of aquaporins in female and male reproductive
systems. Hum Reprod Update. 12:785–795. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu C, Jiang Z, Bazer FW, Johnson GA,
Burghardt RC and Wu G: Aquaporins in the female reproductive system
of mammals. Front Biosci (Landmark Ed). 20:838–871. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Day RE, Kitchen P, Owen DS, Bland C,
Marshall L, Conner AC, Bill RM and Conner MT: Human aquaporins:
regulators of transcellular water flow. Biochim Biophys Acta.
1840:1492–1506. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aralla M, Borromeo V, Groppetti D, Secchi
C, Cremonesi F and Arrighi S: A collaboration of aquaporins handles
water transport in relation to the estrous cycle in the bitch
uterus. Theriogenology. 72:310–321. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Skowronski MT, Skowronska A and Nielsen S:
Fluctuation of aquaporin 1, 5, and 9 expression in the pig oviduct
during the estrous cycle and early pregnancy. J Histochem Cytochem.
59:419–427. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He RH, Sheng JZ, Luo Q, Jin F, Wang B,
Qian YL, Zhou CY, Sheng X and Huang HF: Aquaporin-2 expression in
human endometrium correlates with serum ovarian steroid hormones.
Life Sci. 79:423–429. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hildenbrand A, Lalitkumar L, Nielsen S,
Gemzell-Danielsson K and Stavreus-Evers A: Expression of aquaporin
2 in human endometrium. Fertil Steril. 86:1452–1458. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zou LB, Zhang RJ, Tan YJ, Ding GL, Shi S,
Zhang D, He RH, Liu AX, Wang TT, Leung PC, et al: Identification of
estrogen response element in the aquaporin-2 gene that mediates
estrogen-induced cell migration and invasion in human endometrial
carcinoma. J Clin Endocrinol Metab. 96:E1399–E1408. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Noyes RW, Hertig AT and Rock J: Dating the
endometrial biopsy. Am J Obstet Gynecol. 122:262–263. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Horne AW, King AE, Shaw E, McDonald SE,
Williams AR, Saunders PT and Critchley HO: Attenuated sex steroid
receptor expression in fallopian tube of women with ectopic
pregnancy. J Clin Endocrinol Metab. 94:5146–5154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fushimi K, Uchida S, Hara Y, Hirata Y,
Marumo F and Sasaki S: Cloning and expression of apical membrane
water channel of rat kidney collecting tubule. Nature. 361:549–552.
1993. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schefe JH, Lehmann KE, Buschmann IR, Unger
T and Funke-Kaiser H: Quantitative real-time RT-PCR data analysis:
Current concepts and the novel ‘gene expression's CT difference’
formula. J Mol Med (Berl). 84:901–910. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hohn HP, Linke M and Denker HW: Adhesion
of trophoblast to uterine epithelium as related to the state of
trophoblast differentiation: In vitro studies using cell lines. Mol
Reprod Dev. 57:135–145. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Uchida H, Maruyama T, Ohta K, Ono M, Arase
T, Kagami M, Oda H, Kajitani T, Asada H and Yoshimura Y: Histone
deacetylase inhibitor-induced glycodelin enhances the initial step
of implantation. Hum Reprod. 22:2615–2622. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Y, Kodithuwakku SP, Ng PY, Chai J, Ng
EH, Yeung WS, Ho PC and Lee KF: Excessive ovarian stimulation
up-regulates the Wnt-signaling molecule DKK1 in human endometrium
and may affect implantation: An in vitro co-culture study. Hum
Reprod. 25:479–490. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marsh EE, Shaw ND, Klingman KM,
Tiamfook-Morgan TO, Yialamas MA, Sluss PM and Hall JE: Estrogen
levels are higher across the menstrual cycle in African-American
women compared with Caucasian women. J Clin Endocrinol Metab.
96:3199–3206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hannan NJ, Paiva P, Dimitriadis E and
Salamonsen LA: Models for study of human embryo implantation:
Choice of cell lines? Biol Reprod. 82:235–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nishida M: The Ishikawa cells from birth
to the present. Hum Cell. 15:104–117. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Apps R, Murphy SP, Fernando R, Gardner L,
Ahad T and Moffett A: Human leucocyte antigen (HLA) expression of
primary trophoblast cells and placental cell lines, determined
using single antigen beads to characterize allotype specificities
of anti-HLA antibodies. Immunology. 127:26–39. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Heneweer C, Schmidt M, Denker HW and Thie
M: Molecular mechanisms in uterine epithelium during trophoblast
binding: The role of small GTPase RhoA in human uterine Ishikawa
cells. J Exp Clin Assist Reprod. 2:42005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang D, Xu G, Zhang R, Zhu Y, Gao H, Zhou
C, Sheng J and Huang H: Decreased expression of aquaporin 2 is
associated with impaired endometrial receptivity in controlled
ovarian stimulation. Reprod Fertil Dev. 28:499–506. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smith SK: Regulation of angiogenesis in
the endometrium. Trends Endocrinol Metab. 12:147–151. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gupta SC, Kim JH, Prasad S and Aggarwal
BB: Regulation of survival, proliferation, invasion, angiogenesis,
and metastasis of tumor cells through modulation of inflammatory
pathways by nutraceuticals. Cancer Metastasis Rev. 29:405–434.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Klinge CM, Jernigan SC, Mattingly KA,
Risinger KE and Zhang J: Estrogen response element-dependent
regulation of transcriptional activation of estrogen receptors
alpha and beta by coactivators and corepressors. J Mol Endocrinol.
33:387–410. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chai J, Lee KF, Ng EH, Yeung WS and Ho PC:
Ovarian stimulation modulates steroid receptor expression and
spheroid attachment in peri-implantation endometria: Studies on
natural and stimulated cycles. Fertil Steril. 96:764–768. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu Ng EH, Yeung WS, Yee Lan Lau E, So WW
and Ho PC: High serum oestradiol concentrations in fresh IVF cycles
do not impair implantation and pregnancy rates in subsequent
frozen-thawed embryo transfer cycles. Hum Reprod. 15:250–255. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Forman R, Fries N, Testart J,
Belaisch-Allart J, Hazout A and Frydman R: Evidence for an adverse
effect of elevated serum estradiol concentrations on embryo
implantation. Fertil Steril. 49:118–122. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fossum GT, Davidson A and Paulson RJ:
Ovarian hyperstimulation inhibits embryo implantation in the mouse.
J In Vitro Fert Embryo Transf. 6:7–10. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Noci I, Borri P, Coccia ME, Criscuoli L,
Scarselli G, Messeri G, Paglierani M, Moncini D and Taddei G:
Hormonal patterns, steroid receptors and morphological pictures of
endometrium in hyperstimulated IVF cycles. Eur J Obstet Gynecol
Reprod Biol. 75:215–220. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kalem Z, Kalem MN and Gurgan T: Methods
for endometrial preparation in frozen-thawed embryo transfer
cycles. J Turk Ger Gynecol Assoc. 17:168–172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Valbuena D, Jasper M, Remohi J, Pellicer A
and Simon C: Ovarian stimulation and endometrial receptivity. Hum
Reprod (2 Suppl 14). S107–S111. 1999. View Article : Google Scholar
|
|
42
|
Xu LZ, Gao MZ, Yao LH, Liang AJ, Zhao XM
and Sun ZG: Effect of high ovarian response on the expression of
endocrine gland-derived vascular endothelial growth factor
(EG-VEGF) in peri-implantation endometrium in IVF women. Int J Clin
Exp Pathol. 8:8902–8911. 2015.PubMed/NCBI
|
|
43
|
Rashidi B, Soleimani Rad JI, Roshangar L
and Alizadeh Miran R: Evaluation of pinopodes expression on the
mouse endometrium immediately before implantation by treatment with
HMG/HCG and sildenafil citrate administration. Iran J Basic Med
Sci. 15:1091–1096. 2012.PubMed/NCBI
|
|
44
|
Cheema MU, Irsik DL, Wang Y, Miller-Little
W, Hyndman KA, Marks ES, Frøkiær J, Boesen EI and Norregaard R:
Estradiol regulates AQP2 expression in the collecting duct: A novel
inhibitory role for estrogen receptor alpha. Am J Physiol Renal
Physiol. 309:F305–F317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chenette PE, Sauer MV and Paulson RJ: Very
high serum estradiol levels are not detrimental to clinical outcome
of in vitro fertilization. Fertil Steril. 54:858–863. 1990.
View Article : Google Scholar : PubMed/NCBI
|