Introduction
Breast cancer is the most frequently diagnosed
cancer and is considered to be the second leading cause of
cancer-associated mortality in women (1,2).
Breast cancer is routinely treated with surgery, hormone therapy,
chemotherapy, radiation therapy or a selective combination;
however, these current therapies are not completely effective, and
have also been reported to induce unwanted side effects and
toxicities (2).
Epidemiologic studies have demonstrated that
specific agents in dietary plants may serve tumor-suppressive and
therapeutic roles (3). Pertinent
to this, statistical studies have reported that dietary intake of
cruciferous vegetables may reduce the risk of cancer incidence and
progression (4). Isothiocyanates
(ITCs) are mainly derived from cruciferous vegetables, and may be
potentially used for the prevention of various types of cancer,
including lung, liver, breast and colon cancers (5,6).
Benzyl isothiocyanate (BITC) is a phytochemical that has
demonstrated preventive efficacy in cancer xenograft and transgenic
mouse models in the absence of notable side effects or toxicity
(7–10). Administration of BITC markedly
suppressed the incidence and stage of mammary hyperplasia and the
progression of carcinoma in mice (7,9,10).
In vitro studies have reported that BITC acts via diverse
mechanisms to induce antitumorigenic effects, including the
inhibition of cell proliferation and induction of apoptosis,
modulation of cell cycle arrest and inhibition of cancer cell
migration and invasion (11–13).
Additional studies revealed that BITC regulated various signaling
pathways; for example, BITC induced apoptosis by inhibiting the
phosphatidylinositide 3-kinase (PI3K) pathway, or by activating the
p53-liver kinase B1 (LKB1) and/or the p73-LKB1 pathways (11,12).
Additionally, BITC induced reactive oxygen species (ROS)-dependent
suppression of STAT3 protein expression, and inhibited migration
and invasion by affecting the MAPK signaling pathway and the
activity of matrix metalloproteinase-2/-9 enzymes (13–16).
Numerous studies investigated the effects of BITC on cancer
prevention and treatment; however, the underlying mechanisms of its
antitumor properties remain unclear.
Dysregulation of the Wnt signaling pathway is a
hallmark of several aggressive human cancers (17). At present, the Wnt signaling
pathway is considered an attractive area of research with regard to
cancer treatment and therapy (18). The key signaling molecule of this
pathway is β-catenin, whose activity is mainly regulated by a
destruction complex consisting of adenomatous polyposis coli (APC),
Axin2 and glycogen synthase kinase-3β (GSK-3β) (19). Wnt/β-catenin serves various roles
in numerous types of cancer cell in response to different
treatments, including the regulation of cell proliferation,
migration, apoptosis and epithelial-to-mesenchymal transition (EMT)
(20–22).
BITC inhibited human breast cancer cell
tumorigenesis via the suppression of the forkhead box H1
(FOXH1)-mediated Wnt/β-catenin pathway (22); however, whether BITC acts via the
Wnt signaling pathway in murine mammary carcinoma cells is unclear.
In the present study, the ability of BITC to inhibit the growth and
migration of murine breast cancer cells was investigated, as were
the associated mechanisms underlying this process.
Materials and methods
Reagents, cells and animals
BITC was purchased from Sigma-Aldrich (Merck KGaA).
SYBR Premix Ex Taq™ and PrimeScript™ RT kits were obtained from
Takara Bio, Inc. Antibodies specific for APC (cat. no. sc-896;
Santa Cruz Biotechnology, Inc.), β-catenin (cat. no. sc-7963; Santa
Cruz Biotechnology, Inc.), phosphorylated (p)-β-catenin (cat. no.
GTX50256; GeneTex, Inc.), Wnt2 (cat. no. ab109222; Abcam),
E-cadherin (cat. no. 3195; Cell Signaling Technology, Inc.),
vimentin (cat. no. GTX100619; GeneTex, Inc.), Axin2 (cat. no.
ab109307; Abcam), GSK-3β (cat. no. ab32391; Abcam), cyclin D1 (cat.
no. ab134175; Abcam), c-Myc (cat. no. ab32072; Abcam) and β-actin
(cat. no. TA-09; Zhongshan Jinqiao Bio-Technology Co., Ltd.) were
used in the present study. BITC was dissolved in dimethyl
sulfoxide.
The murine mammary carcinoma cell line 4T1-Luc
(stable transfection of the firefly luciferase gene) was purchased
from the Cold Spring Harbor Laboratory and cultured in DMEM
(HyClone; GE Healthcare Life Sciences) supplemented with 10% fetal
bovine serum (Biological Industries) in the presence of 1%
antibiotics (100 U/ml penicillin and 100 µg/ml streptomycin) and 2
mM L-glutamine at 37°C in a 5% CO2 humidified
atmosphere. The normal mammary epithelial cell line MCF-10A was
obtained from the American Type Culture Collection and was cultured
in DMEM/F-12 medium (Thermo Fisher Scientific, Inc.) supplemented
with 5% horse serum (Thermo Fisher Scientific, Inc.), 20 ng/ml
epidermal growth factor (PeproTech, Inc.), 100 ng/ml cholera toxin
(Sigma-Aldrich; Merck KGaA), 10 µg/ml human insulin, 100 U/ml
penicillin and 100 µg/ml streptomycin. The cells were maintained at
37°C and 5% CO2 in a humidified incubator.
Female BALB/c mice (specific pathogen-free grade, 8
weeks old, n=72 mice, 20–24 g) were provided by the Laboratory
Animal Center of Lanzhou University. The mice were housed in the
barrier system facilities of the Key Laboratory of Preclinical
Studies for New Drugs of Gansu Province (School of Basic Medical
Sciences, Lanzhou University), which specializes in preclinical
studies for novel drugs. Food and water was provided ad
libitum in the facility, and the animals were maintained in a
specific pathogen-free condition at 20–26°C, 40–70% humidity with a
12-h light/dark cycle. All the animal procedures were ethically
approved by the Laboratory Animal Science and Technology Management
Committee of the Lanzhou University School of Basic Medical
Sciences, and were conducted in accordance with the Guide for Care
and Use of Laboratory Animals (23).
Cell viability assay
A cell viability assay was performed as previously
described (24). Briefly, cells
were plated in 96-well plates at an initial density of
4×103 cells/well and cultured overnight. Subsequently,
the medium was replaced with fresh complete medium containing
various concentrations of BITC (0, 1, 2.5, 5, 10, 20 and 40 µM).
The cells were incubated for 24, 48 and 72 h. A total of 10 µl MTT
solution was then added to each well, followed by incubation for 4
h at 37°C. 10% SDS was added to each of the wells, and incubated
overnight at 37°C to dissolve the formazan crystals. Finally, the
absorbance values of each well were measured at 570 nm, and the
readings were quantified using a Powerwave X plate reader (Bio-Tek
Instruments, Inc.). Cell proliferation inhibitory rates were
calculated.
Clonogenicity assay
4T1-Luc cells were pretreated with various
concentrations of BITC (0, 1, 2.5, 5, 10 and 20 µM) for 24 h.
Subsequently, the cells were digested with 0.25% trypsin at 37°C
for 2–3 min, and single cell suspensions were seeded in 6-well
plates at a density of 1,000 cells/well. Following 2 weeks of
culture at 37°C in a 5% CO2 humidified atmosphere, the
cells in each well were washed with 1 ml phosphate buffered saline
(PBS) and fixed with 0.5 ml 100% methanol at room temperature for
30 min. Subsequently, the cells were stained with crystal violet
(0.1% in 20% methanol) at room temperature for 30 min. Excess
crystal violet was removed using dH2O. The colony
numbers were assessed visually using Olympus IX81 inverted
microscope (Olympus Corporation) at ×40 magnification, and only
colonies containing >50 cells with normal morphology were
counted.
Morphological characteristics
The cells were seeded into 6-well plates at a
density of 1×105 cells/well in 2 ml of medium.
Subsequently, they were treated with BITC as aforementioned and
observed under an inverted microscope (IX81; Olympus Corporation)
or mounted onto slides. The cells on the slides were stained using
Wright-Giemsa at room temperature for 15 min and observed under a
light microscope (AX80; Olympus Corporation).
Cells were sequentially fixed with 2% glutaraldehyde
at 4°C for 48 h and 1% OsO4 at 4°C for 90 min for
ultrastructural observation. Following dehydration, thin sections
(50–70 nm) embedded by resin were stained with uranyl acetate for
30 min and lead citrate for 15 min at room temperature for visual
investigation under a JEM 1230 transmission electron microscope
(JEOL, Ltd.). Images were acquired digitally from a randomly
selected pool of 10–15 fields for each condition at ×5,000
magnification.
Apoptotic and cell cycle analyses
The induction of apoptosis by BITC was determined
using an Annexin V/dead cell apoptosis kit (cat. no. V13241;
Invitrogen; Thermo Fisher Scientific, Inc.). Samples were gently
suspended in 100 µl binding buffer containing 5 µl of Annexin
V-FITC and 5 µl propidium iodide (PI), and further incubated for 15
min in the dark at room temperature. Finally, cells were suspended
in 500 µl binding buffer and were detected by flow cytometry (Epics
XL; Beckman Coulter, Inc.) and analyzed with Expo 32 software
(Beckman Coulter, Inc.). The apoptotic rate was determined for each
condition as follows: Apoptotic rate=(apoptotic rate early
apoptosis + apoptotic rate late apoptosis)
×100%.
Cells were treated with various concentrations of
BITC (0, 2.5, 5 and 10 µM) for 24 and 48 h, and then washed with
ice-cold PBS and fixed in cold 70% ethanol overnight. Prior to
analysis, cells were washed with PBS and incubated with PI for 30
min at room temperature in the dark. The DNA contents were analyzed
with Multicycle for Windows using an Epics XL flow cytometer
(Beckman Coulter, Inc.).
Mitochondrial transmembrane potential
(MTP) detection
Cells were washed with PBS and incubated with 1 µM
rhodamine 123 (Rh123) in the dark for 30 min at 37°C. The mean
fluorescence intensity (MFI) of Rh123-labeled cells was analyzed
with System II software version 3.0 (Epics XL; Beckman Coulter,
Inc.) using a flow cytometer (Epics XL; Beckman Coulter, Inc.) at
an excitation and emission wavelength of 488 and 525 nm,
respectively. A minimum of 1×104 events/sample were
acquired.
Wound healing assay
For the wound healing assay, cells were seeded in
6-well plates and grown in complete medium to ~100% confluence.
Subsequently, they were washed with serum-free medium and
serum-starved for 16 h. A 1-mm wide scratch was made across the
cell layer using a sterile pipette tip. The plates were
photographed immediately following scratching. Following treatment
of the cells with various concentrations of BITC (0, 1, 2.5, 5, 10
and 20 µM) for 0, 10 and 24 h, the plates were photographed at
identical locations to the initial images using Olympus IX81
inverted microscope (Olympus Corporation) at ×100
magnification.
Cell invasion and migration
assays
The ability of cells to penetrate a synthetic
basement membrane was assessed using a Matrigel-Boyden chamber
invasion assay (BD Biosciences). Transwell inserts (6.5 mm) fitted
with polycarbonate filters (8-µm pore size) were used. The upper
chambers of the wells were coated with Matrigel (BD Biosciences). A
total of 1×105 4T1-Luc cells were cultured in serum-free
medium containing treatments as aforementioned. The cells were
allowed to penetrate at 37°C for 24 h. The lower chambers of the
plate were filled with complete medium. Following 24 h of
incubation, the cells remaining in the upper part of the insert
membrane were removed by gentle scraping with a sterile cotton
swab. The cells that had invaded through the Matrigel to the bottom
of the insert were fixed in 4% paraformaldehyde at room temperature
for 30 min and stained with crystal violet (0.1% in 20% methanol)
at room temperature for 30 min. Five random fields were imaged
(magnification, ×100) using Olympus IX71 inverted microscope
(Olympus Corporation). All presented data were obtained from at
least three independent experiments performed in duplicate. The
migration assay was performed in the same manner as the invasion
assay in the absence of Matrigel.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
assay
RNA isolation and RT-qPCR were performed as
previously described (24). Total
RNA from the cells was isolated using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and reverse
transcribed to cDNA using a PrimeScript RT reagent kit purchased
from Takara Bio, Inc. according to the manufacture's protocol. qPCR
was performed using an SYBR Premix Ex Taq II kit. The following
primers were synthesised by Takara Bio, Inc.: APC, forward
5′-GAGAAACCCTGTCTCGAAAAAA-3′, reverse 5′-AGTGCTGTTTCTATGAGTCAAC-3′;
β-catenin, forward 5′-AGGGTGCTATTCCACGACTA-3′, reverse
5′-CACCCTTCTACTATCTCCTCCAT-3′; and β-actin, forward
5′-TGCTCCTCCTGAGCGCAAGTA-3′ and reverse
5′-CCACATCTGCTGGAAGGTGGA-3′. Gene expression levels were analyzed
using a Rotor-Gene 3000 quantitative PCR amplifier (Corbett Life
Science; Qiagen, Inc.). qPCR was conducted as follows: 10 sec at
95°C, followed by 40 cycles of denaturation at 95°C for 5 sec and
annealing at 60°C for 30 sec. β-actin was used as the internal
control. The relative expression levels of the genes were
determined by the 2−ΔΔCq method (25).
Western blot analysis
Western blot analysis was performed as previously
described. Proteins from cells were isolated using RIPA lysis
buffer (Beijing Solarbio Science & Technology Co. Ltd.), and
proteins from tumor tissues were dissolved in RIPA lysis buffer and
isolated using sonic disruption. The concentration levels were
determined using the BCA method. The proteins (40 µg) were
subsequently fractionated by 10% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes (EMD Millipore).
Immunodetection was performed by blocking the membranes in 0.1%
TBS-Tween 20 containing 5% nonfat milk at room temperature for 1 h,
followed by incubation with primary antibodies at 4°C overnight.
The primary antibodies used were as follows: APC (1:500), β-catenin
(1:500), p-β-catenin (1:500), Wnt2 (1:2,000), E-cadherin (1:1,000),
vimentin (1:1,000), Axin2 (1:2,000), GSK-3β (1:2,000), cyclin D1
(1:2,000), c-Myc (1:2,000) and β-actin (1:1,000). The following
morning, the membranes were incubated for 1 h at room temperature
with horseradish peroxidase conjugated goat anti-mouse (cat. no.
SA00001-1) or goat anti-rabbit (cat. no. SA00001-2) secondary
antibody (1:5,000; Proteintech Group, Inc.) and subsequently
developed using an Amersham Enhanced Chemiluminescence Western
blotting detection system (GE Healthcare Life Sciences) according
to the manufacturer's protocols.
Immunofluorescence assay
The cells were plated in 24-well plates at a density
of 20,000 cells/ml and allowed to adhere overnight. Following
treatment with BITC (0 and 5 µM) for 24 h, the cells were washed
with PBS and fixed with 4% paraformaldehyde at room temperature for
30 min. The cells were permeabilised with 0.4% Triton X-100 on ice
for 5 min, followed by blocking in 3% bovine serum albumin (cat.
no. H1130; Beijing Solarbio Science & Technology Co. Ltd.) at
room temperature for 1 h and incubation with primary antibodies at
4°C overnight. The primary antibodies used were as follows: APC
(1:100) and β-catenin (1:100). Following a final 4-h incubation at
room temperature with RBITC conjugated goat anti-mouse (1:500; cat.
no. SR131; Beijing Solarbio Science & Technology Co. Ltd.) or
fluorescein conjugated goat anti-rabbit (1:500; cat. no. SA00003-2;
Proteintech Group Inc.) secondary antibodies, the cells were
counterstained with DAPI (Beijing Solarbio Science & Technology
Co. Ltd.) at room temperature for 10 min and visualized with
confocal microscopy at ×1,000 magnification (in six
micro-fields).
In vivo xenograft mouse model
In vivo xenograft mouse models were
established based on previously described protocols (26,27).
4T1-Luc cells (2×106/ml) were implanted in the mammary
fat pads of female BALB/c mice (8 weeks old, n=32 mice) to
establish the orthotopic xenograft model. On the following day, the
mice were randomly grouped into four experimental groups (8
mice/group) and treated with oral gavage of vehicle (0.2 ml/20 g;
negative control), cisplatin (DDP; 2 mg/kg; positive control), 1.5
mg/kg BITC or 3 mg/kg BITC every day for 28 days.
To establish the metastatic model, the same density
of 4T1-Luc cells (2×106/ml) was injected into the tail
vein of female BALB/c mice (8 weeks old, n=40 mice). On the next
day, the mice were randomly grouped into four experimental groups
(10 mice/group). Mice from each group were treated the same as the
mice form the orthotopic xenograft study every other day for 35
days.
Every 7 days, D-luciferin (0.15 g/l) was injected
intraperitoneally, and the mice were analyzed using a Photon Imager
for luciferase activity using the Lumina II Living Image 4.0
software (PerkinElmer, Inc.). The mice were sacrificed following
the final imaging day. The tumors were collected, measured, weighed
and subjected to further analysis, including H&E staining,
immunohistochemistry (IHC) and western blot analyses.
H&E staining
After the tumors were fixed in 4% formaldehyde at
4°C for 24 h, they were embedded in paraffin followed by the
routine method and sliced to 5 µm thickness. The sections were
dewaxed with 100% xylene for 15 min three times and hydrated using
a series of graded concentrations of ethanol (100% ethanol, 1 min;
100% ethanol, 1 min; 95% ethanol, 1 min; 90% ethanol, 1 min; 80%
ethanol, 1 min; 70% ethanol, 1 min; distilled water, 5 min) at room
temperature. Following deparaffinization and rehydration, the
sections were stained with hematoxylin at room temperature for 5
min, immersed five times in a solution of 1% HCl and 70% ethanol
and subsequently rinsed with distilled water. Then the sections
were stained with eosin at room temperature for 2 min. Following
dehydration, the sections were treated with 100% xylene at room
temperature for 2 min three times. A light microscope (AX80;
Olympus Corporation) was used to examine the histopathological
morphology at a magnification of ×400. The images were digitized
and quantified using ImageJ (National Institutes of Health).
IHC and terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL) assay
For IHC analysis, tumors were fixed in 4%
formaldehyde at 4°C for 24 h and embedded in paraffin. The sections
(5 µm) were subjected to antigen retrieval. Citrate buffer (pH 6.0)
was used to retrieve antigen as a repair solution. The antigen was
placed in an autoclave for 15 min. Following antigen retrieval, the
sections were incubated with 3% H2O2 at room
temperature for 10 min to block endogenous peroxidase activity and
1% bovine serum albumin (Wuhan Boster Biological Technology, Ltd.)
at room temperature for 1 h to block non-specific binding. Anti-APC
antibody (1:200) and anti-β-catenin antibody (1:200) were added
dropwise to the sections and incubated at 4°C overnight. According
to the protocol of an IHC three-step detection kit (SP9001 and
SP9002; Zhongshan Jinqiao Bio-Technology Co., Ltd.), the section
was incubated with 100 µl antibodies at room temperature for 1 h.
Subsequently, the sections were exposed to 3,3-diaminobenzidine
(cat. no. ZLI-9017; Zhongshan Jinqiao Bio-Technology Co., Ltd.) for
5–10 min and observed under a light microscope (AX80; Olympus
Corporation) at ×400 magnification. At least four random,
nonoverlapping representative images were captured using Image-Pro
Plus software, version 6.0 (Media Cybernetics, Inc.) for the
quantification of protein expression. The images corresponded to
each tumor section from the eight tumors of each group.
TUNEL assays were performed according to the
manufacturer's protocol (Roche Diagnostics). The 5 µm
paraffin-embedded sections were fully dewaxed and hydrated, soaked
in 3% H2O2 at room temperature for 30 min to
inhibit endogenous peroxidase activity. After rinsing in PBS three
times at room temperature for 5 min, the sections were treated with
proteinase K (Sigma-Aldrich; Merck KGaA) at 37°C for 15 min. After
rinsing in PBS three times at room temperature for 5 min, the
sections were soaked in the TdT buffer for 10 min and then
incubated at 37°C for 1 h in a moist chamber with 50 µl of the TdT
buffer containing TdT (Roche Diagnostics). After further rinsing in
PBS three times for 5 min, the sections were dipped in DAB at room
temperature for 3–4 min and the nuclei were counterstained with
hematoxylin buffer at room temperature for 10 sec. The samples were
detected and analyzed using an Olympus microscope (AX80; Olympus
Corporation) at ×400 magnification (in six micro-fields).
Statistical analysis
All experiments were performed in triplicates.
Statistical analyses were performed using Microsoft Excel software,
version 2016 (Microsoft Corporation). Significant differences were
analyzed using one-way ANOVA and a post-hoc Bonferroni test.
P<0.05 was considered to indicate a statistically significant
differences. The results were presented as the mean ± standard
deviation of triplicate experiments performed three times.
Results
BITC inhibits the proliferation of
murine mammary carcinoma cells in vitro and in vivo
MTT and clonogenicity assays were conducted to
assess the antitumorigenic effects of BITC on 4T1-Luc cells in
vitro. In response to BITC treatment, the survival of 4T1-Luc
cells was significantly inhibited in a concentration-dependent
manner (Fig. 1A). At
concentrations of BITC >10 µM, the survival rate was <50%. As
indicated in Fig. 1B, a
clonogenicity assay indicated that BITC significantly suppressed
the colony formation of 4T1-Luc cells. Consistent with the in
vitro results, BITC demonstrated antitumorigenic effects in
vivo. Following treatment with 1.5 or 3 mg/kg BITC, the growth
of 4T1-Luc cell tumors in BALB/c mice of the orthotopic xenograft
model was significantly inhibited, as determined by tumor volume
and estimated tumor weight (Fig.
1C). Every 7 days, the tumors were monitored dynamically using
an in vivo optical bioluminescence imaging system. The
results further indicated that BITC treatment inhibited tumor
growth in mice (Fig. 1D and E).
Following animal sacrifice, the tumors were separated. H&E
staining demonstrated that tumors from 3 mg/kg BITC-treated mice
displayed significantly decreased numbers of mitotic cells
(Fig. 1F). The necrosis phenomena
in BITC-treated mice were not as clear as the control mice. Thus,
we inferred that BITC may inhibit necrosis.
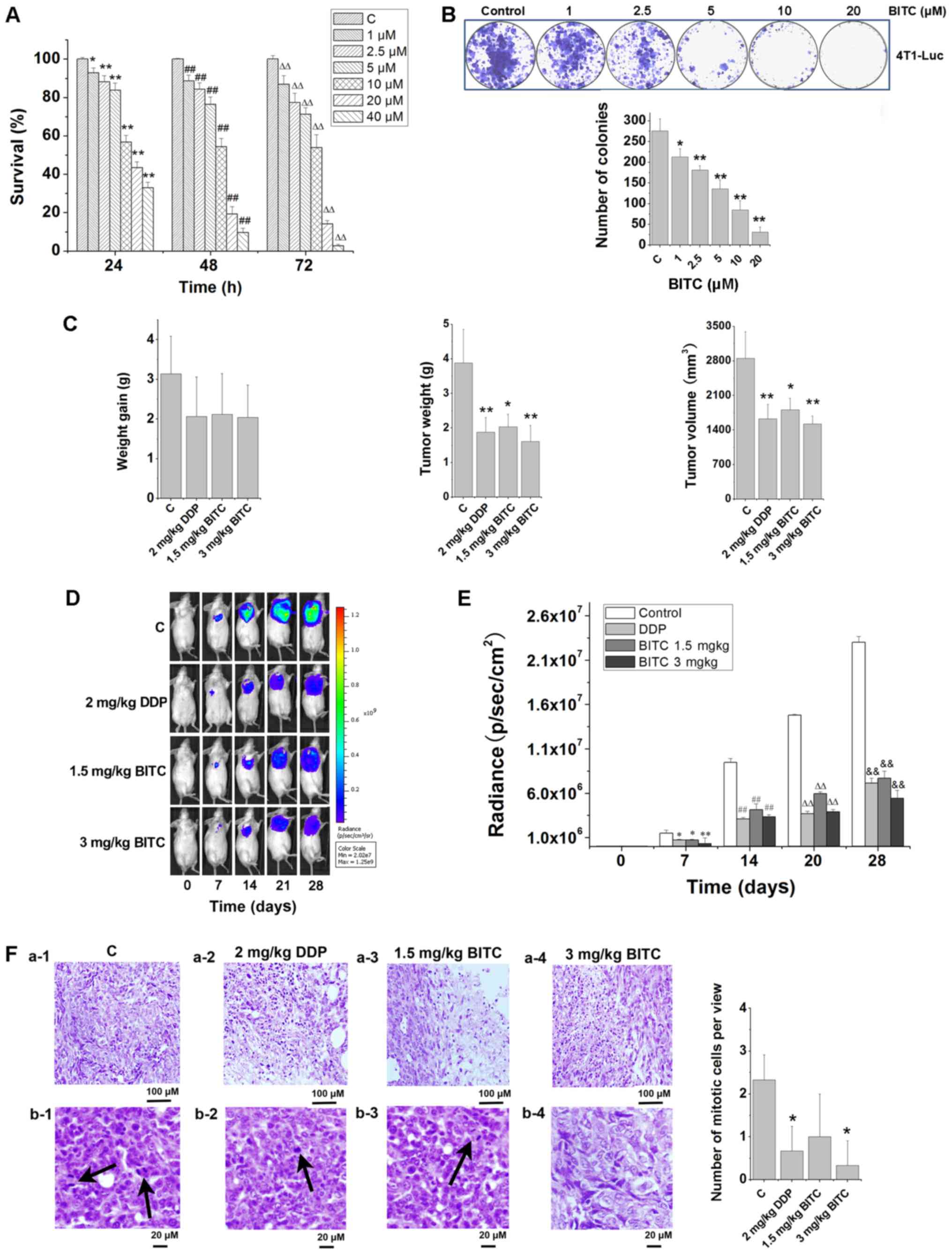 | Figure 1.BITC serves an antitumorigenic role
by inhibiting the proliferation and growth of 4T1-Luc cells in
vitro and in vivo. (A) BITC inhibited the proliferation
of 4T1-Luc cells in a concentration-dependent manner. The viability
of 4T1-Luc cells treated with various concentrations of BITC for
24, 48 and 72 h was detected using an MTT assay. N=3/group.
*P<0.05, **P<0.01 vs. C (24 h); ##P<0.01 vs. C
(48 h); ∆∆P<0.01 vs. C (72 h). (B) Clonogenicity of
4T1-Luc cells treated with various concentrations of BITC.
*P<0.05, **P<0.01 vs. Control. (C) BITC inhibits the growth
of xenograft tumors formed by 4T1-Luc cells in vivo. The
antitumorigenic actions of BITC and DDP were detected by measuring
(left to right): Weight gain; tumor weight; and tumor volume.
N=8/group. *P<0.05, **P<0.01 vs. C. (D) Tumors were
dynamically detected via bioluminescence in vivo imaging
every 7 days (n=8 mice/group). (E) Tumor growth was monitored via
bioluminescence in vivo imaging for 4 weeks, and the
radiance (p/sec/cm2) detected from the tumors was
analyzed. *P<0.05, **P<0.01 vs. Control (7 days);
##P<0.01 vs. Control (14 days);
∆∆P<0.01 vs. Control (21 days);
&&P<0.05 vs. Control (28 days). (F) Tumor
sections were analyzed via H&E staining. a-1-4, necrosis in
control, DDP, 1.5 mg/kg BITC and 3 mg/kg BITC group tissues. b-1-4,
mitotic cells in control, DDP, 1.5 mg/kg BITC and 3 mg/kg BITC
group tissues (×400). Arrows indicate mitotic cells. *P<0.05 vs.
C. Data are presented as the mean ± standard deviation. BITC,
benzyl isothiocyanate; C, control; DDP, cisplatin. |
BITC induces 4T1-Luc cell apoptosis in
vitro and in vivo
The induction of 4T1-Luc cell apoptosis following
BITC treatment was demonstrated by phase-contrast microscopy and
light microscopy using Right-Giemsa staining (Fig. 2A). The formation of the apoptotic
bodies was apparent in BITC-treated 4T1-Luc cells using
transmission electron microscopy (Fig.
2A). Flow cytometry using Annexin V-FITC and PI staining
indicated that BITC treatment induced apoptosis in 4T1-Luc cells,
with the apoptotic rate significantly increased following treatment
with 2.5, 5 and 10 µM BITC (Fig.
2B). Alterations in the MTP are also apoptotic indicators; the
use of Rh123 reveals the changes to the MTP. The results indicated
that the mitochondrial transmembrane potential was significantly
depolarized in BITC-treated 4T1-Luc cells compared with untreated
cells, suggesting that BITC treatment induced the apoptosis of
4T1-Luc cells (Fig. 2C). BITC
treatment also altered the cell cycle distribution of 4T1-Luc cells
(Fig. 2D). BITC treatment led to
cell cycle arrest and the accumulation of 4T1-Luc cells at the G2/M
phase compared with the cell cycle distribution of control-treated
4T1-Luc cells. In vivo, TUNEL staining of tumor sections
indicated that the number of TUNEL-positive apoptotic cells in
BITC-treated mice was significantly increased compared with in
non-treated mice (Fig. 2E and
F).
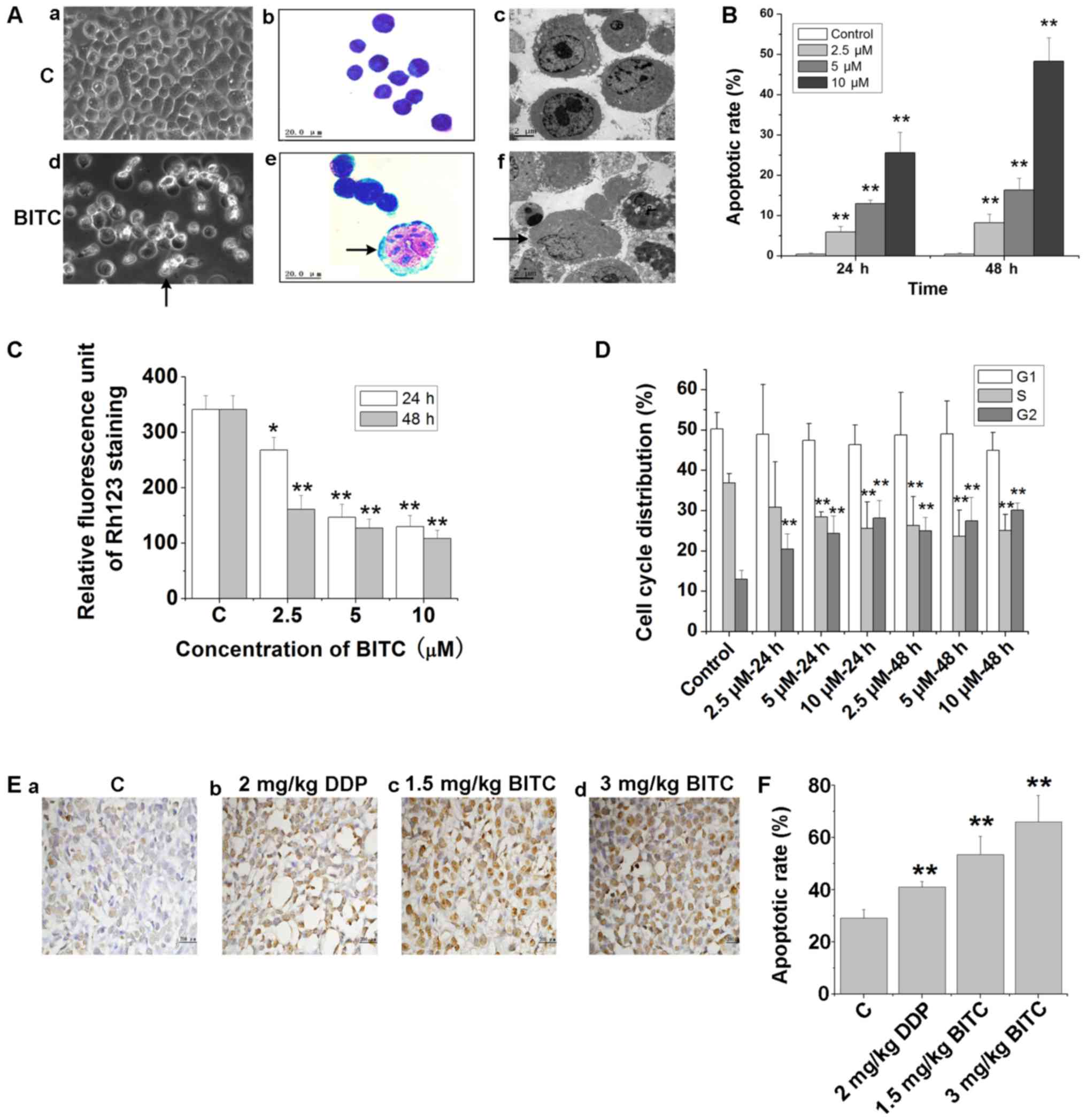 | Figure 2.BITC induces the apoptosis of 4T1-Luc
cells. (A) Morphological alterations of 4T1-Luc cells treated with
or without BITC. (a) and (d) Morphological changes detected by
phase contrast microscopy (×200). (b) Untreated and (e)
BITC-treated cells were stained by Wright-Giemsa staining and
recorded by light microscopy (×1,000). (c) and (f) Ultrastructural
changes of 4T1-Luc cells detected by transmission electron
microscopy (×5,000). Arrows indicate apoptotic cells. (B) 4T1-Luc
cells were treated with various concentrations of BITC for 24 and
48 h, and apoptosis was analyzed via Annexin V/PI double staining.
**P<0.01 vs. Control. (C) Mitochondrial transmembrane potential
of 4T1-Luc cells treated with various concentrations of BITC for 24
and 48 h was detected via flow cytometry using Rh123 staining.
*P<0.05, **P<0.01 vs. C. (D) BITC induced cell cycle arrest
in 4T1-Luc cells. 4T1-Luc cells were treated as indicated and
subjected to cell cycle analysis. **P<0.01 vs. Control. (E)
Tumor sections were assessed via TUNEL staining to determine the
number of apoptotic cells (magnification, ×400). (F) Quantification
of TUNEL staining in tumor sections. **P<0.01 vs. Control. BITC,
benzyl isothiocyanate; C, control; DDP, cisplatin; PI, propidium
iodide; Rh123, rhodamine 123. |
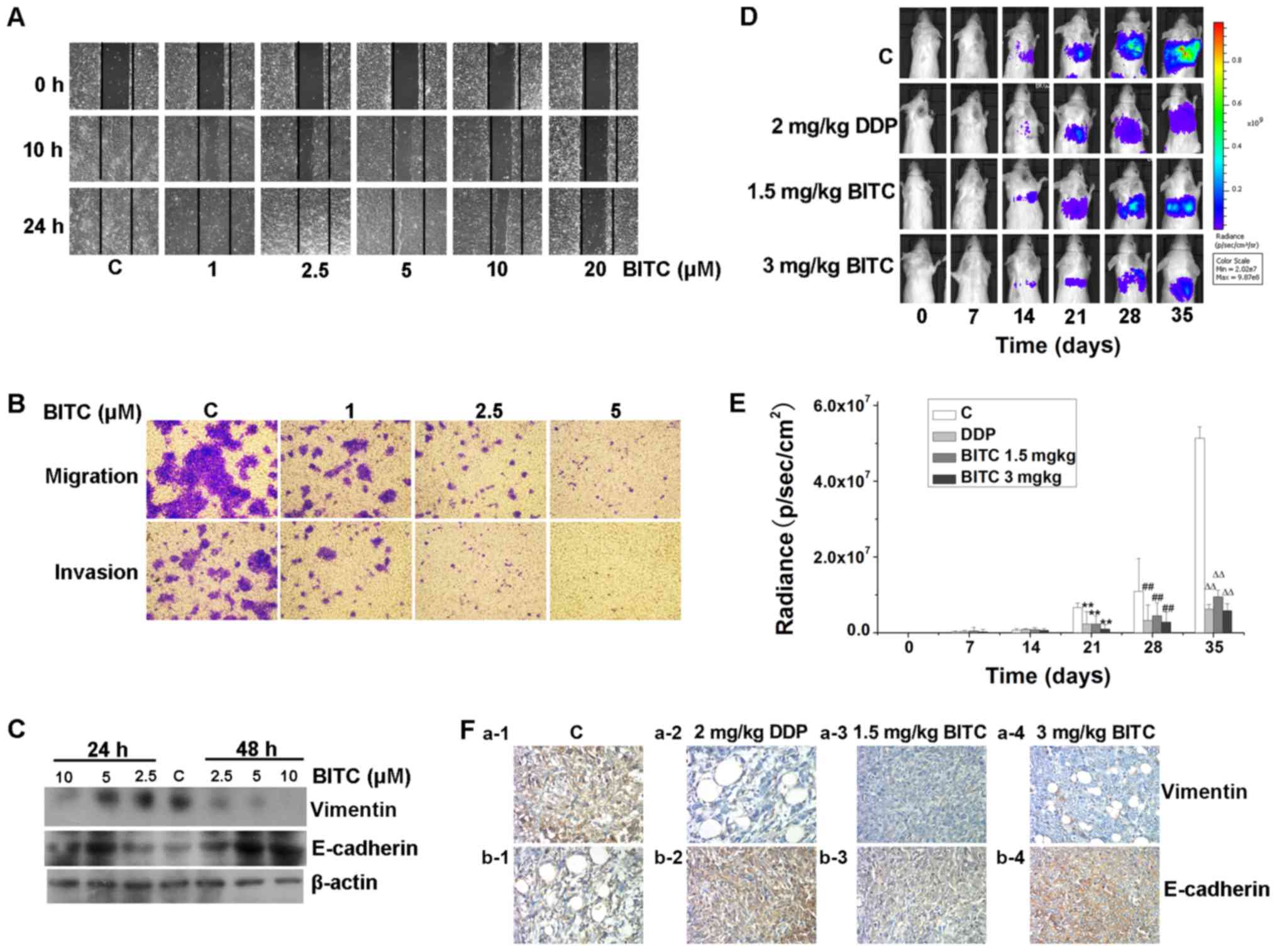
BITC suppresses the migration,
invasion and metastasis of 4T1-Luc cells by activating E-cadherin
and inhibiting the expression of vimentin
BITC treatment markedly inhibited the mobility of
4T1-Luc cells in a concentration-dependent manner (Fig. 3A). The effects of BITC were
assessed on the migration and invasion of cells. Transwell chambers
coated with collagen or Matrigel were used and as presented in
Fig. 3B, the migratory and
invasive abilities of 4T1-Luc cells were notably inhibited
following BITC treatment. Consistent with the in vitro
results, the in vivo studies further revealed that BITC
inhibited the metastasis of murine mammary carcinoma, as
demonstrated using an in vivo optical bioluminescence
imaging system (Fig. 3D and E).
The expression levels of the metastasis inhibitor E-cadherin were
increased, whereas the expression levels of the metastasis promoter
vimentin (28) were decreased in
BITC-treated cells and tissues (Fig.
3C and F). These results suggested that the
metastasis-inhibitory effects of BITC on murine mammary carcinoma
were mediated by upregulating E-cadherin and downregulating
vimentin expression levels.
BITC induces antitumorigenic effects
by regulating the Wnt/β-catenin pathway
The Wnt signaling pathway serves an important role
in carcinogenesis (17). The
expression levels of its signaling transducers were evaluated in
BITC-treated cells and tumor-bearing mice. Western blotting
indicated that the expression levels of APC and GSK-3β were notably
increased in 4T1-Luc cells following BITC treatment compared with
those in control cells (Fig. 4A),
whereas the expression of the tumor promoters β-catenin and Wnt2
was inhibited in response to BITC treatment. The expression levels
of Axin2 were markedly unaffected. Additionally, the
phosphorylation levels of β-catenin were markedly upregulated,
whereas those of the downstream factors of β-catenin, such as
cyclin D1 and c-Myc were downregulated (Fig. 4B). Immunofluorescence staining
demonstrated that the expression levels of APC were markedly
increased, whereas those of β-catenin were decreased (Fig. 4C). In addition, β-catenin protein
appeared to be partially transferred from the cytoplasmic to the
nuclear region (Fig. 4C). The mRNA
expression levels of APC and β-catenin were detected
via RT-qPCR analysis. The transcriptional levels of APC and
β-catenin in BITC-treated 4T1-Luc cells were significantly
increased and decreased, respectively (Fig. 4D), in accordance with the
aforementioned alterations observed at the translational level.
Western blotting indicated that the expression levels of APC and
β-catenin were notably increased and decreased, respectively in
tumors from BITC-treated mice compared with controls (Fig. 4E). IHC staining further indicated
that in BITC-treated mice, APC expression was upregulated, whereas
that of β-catenin was downregulated (Fig. 4F), suggesting that BITC may serve
antitumorigenic roles in murine mammary carcinoma cells by
regulating the Wnt/β-catenin pathway and the APC-GSK-3β degradation
complex.
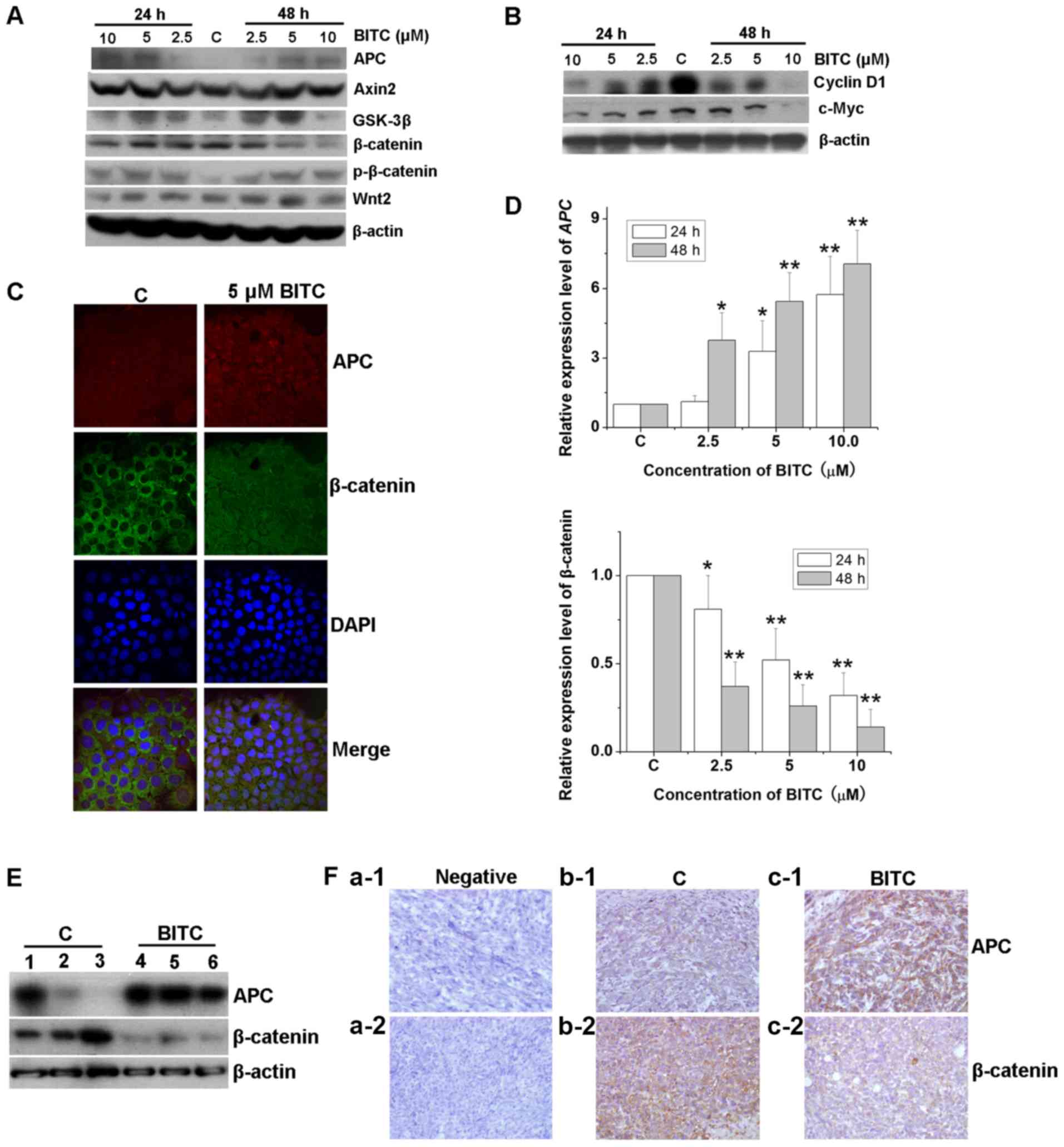 | Figure 4.BITC regulates the Wnt/β-catenin
pathway and the APC/β-catenin complex. 4T1-Luc cells were treated
with various concentrations of BITC as indicated for 24 and 48 h.
Total protein lysates were immunoblotted for (A) APC, Axin2,
GSK-3β, β-catenin, p-β-catenin and Wnt2, and (B) cyclin D1 and
c-Myc protein expression. β-actin was used as the internal control.
(C) Immunofluorescence assay was performed to evaluate the
expression and subcellular location of APC and β-catenin protein
(magnification, ×1,000). (D) 4T1-Luc cells were treated with
various concentrations of BITC as indicated for 24 and 48 h, and
total RNA was isolated and reverse transcribed. The mRNA expression
levels of APC and β-catenin were detected via reverse
transcription-quantitative PCR analysis. *P<0.05, **P<0.01
vs. C. (E) Protein lysates isolated from the tumors of control or
BITC-treated mice were subjected to western blot analysis for APC
and β-catenin. 1–6 indicated different mice. (F) Tumors from
control and BITC-treated mice were subjected to immunohistochemical
analysis using APC and β-catenin antibodies (magnification, ×400).
APC, adenomatous polyposis coli; BITC, benzyl isothiocyanate; C,
control; DAPI, 4′,6-diamidino-2-phenylindole; GSK-3β, glycogen
synthase kinase-3β. |
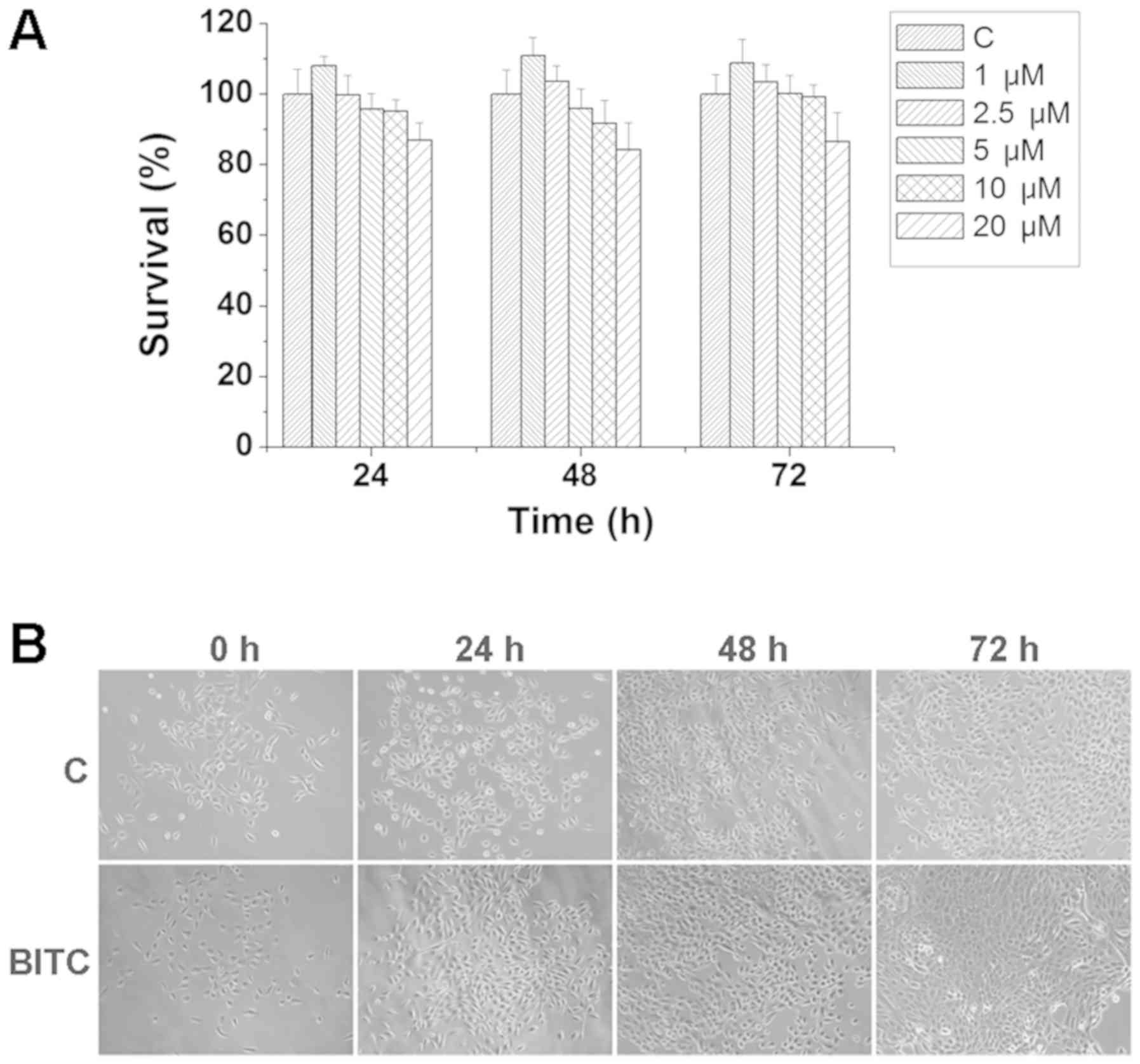
BITC does not induce toxicity or side
effects in the normal mammary epithelial cell line MCF-10A
To investigate whether BITC treatment affects normal
mammary epithelial cells, MCF-10A cells were treated with various
concentrations of BITC for 24, 48 and 72 h. As indicated in
Fig. 5A, BITC treatment did not
induce significant inhibitory effects on the viability of MCF-10A
cells. The morphological differences detected by phase contrast
microscopy indicated that MCF-10A cells treated with BITC exhibited
no morphological alterations compared with control cells (Fig. 5B).
Discussion
BITC is a compound found in cruciferous vegetables
that has been reported to possess antitumor properties (7–15).
Administration of BITC suppresses the incidence, growth and
metastasis of cancer cells (8,9).
Warin et al (7) reported
that BITC prevented mammary carcinogenesis in MMTV-neu mice. Kim
et al (9) demonstrated that
oral BITC treatment induced a significant reduction in the growth
of solid tumors. In the present study, it was demonstrated that the
growth of 4T1-Luc xenografts was significantly inhibited following
BITC treatment. The weight and the volume of the tumors in
BITC-treated and DDP-treated BALB/c mice were significantly
decreased compared with those in untreated mice. As 4T1 cells were
transfected with luciferase, tumor growth could be observed
conveniently and dynamically using an optical in vivo
bioluminescence imaging system. The results suggested that tumor
growth was significantly inhibited. These in vivo
experiments demonstrated that BITC induced antitumor effects in
murine mammary carcinoma.
BITC exhibited diverse mechanisms of action in
various types of cancer. Previous studies revealed that BITC
inhibited neoplasm formation by activating or inhibiting various
signaling pathways and/or processes, including the impairment of
ROS production, disruption of mitochondrial function, inhibition of
the PI3K, regulation of the MAPK signaling pathway and activation
of the p53 pathway (11,12,14,15,29,30).
Kim et al (9) demonstrated
that BITC induced murine mammary cell apoptosis, reduced the number
of pulmonary tumor nodules and decreased the total pulmonary
metastatic volume. Additionally, BITC induced alterations in the
expression levels of indicators of apoptosis and metastasis
(9); however, the authors did not
determine the underlying mechanism of actions of BITC in 4T1 cells.
In the present study, it was revealed that the expression levels of
several factors involved in the Wnt signaling pathway were altered
in murine mammary carcinoma cells in response to BITC treatment.
The expression levels of the proteins APC and GSK-3β were
increased, whereas the expression levels of β-catenin were
decreased, indicating that BITC activated the degradation complex
and regulated the Wnt signaling pathway in order to perform its
tumor-preventive and tumor-inhibitory roles.
The dysregulation of the Wnt signaling pathway is a
hallmark of various types of cancer with an aggressive phenotype
(31). The Wnt signaling
transduction pathway has been reported to serve a vital role in
homeostasis via multiple modes of action (32). The β-catenin-dependent canonical
pathway has been extensively studied compared with other signaling
pathways. APC, GSK-3β and β-catenin form a destruction complex,
which regulates the activity of β-catenin. The destruction complex
phosphorylates β-catenin, inducing a reduction in its levels in the
cytoplasmic and nuclear regions, and resulting in the
downregulation of its downstream targets, including cyclin D1 and
c-Myc (17,19,32).
The results of the present study indicated that BITC promoted the
phosphorylation of β-catenin and decreased its levels. In response
to BITC, β-catenin might target the genes cyclin D1 and c-Myc, and
downregulated their expression, though further investigation is
required. The present findings were consistent with results
reported in previous studies regarding the ability of BITC
treatment to reduce the expression levels of FOXH1 (22), which in turn significantly
increased the expression levels of β-catenin, cyclin D1 and c-Myc
proteins in MCF7 and MDA-MB-231 cells (22). By regulating the induction of the
Wnt/β-catenin pathway via FOXH1, BITC exposure suppressed cell
growth and invasion in human breast cancer cells (22).
Curcumin is a naturally occurring phenolic compound,
which regulates the Wnt/β-catenin pathway via metastasis associated
protein 1 and suppresses proliferation and invasion in non-small
cell lung cancer (33). Caudatin
is a C-21 steroidal glycoside isolated from Chinese herbs, which
can induce apoptosis in gastric cancer cells by modulating the
Wnt/β-catenin signaling (34). In
the present study, it was reported that BITC inhibited
proliferation, induced cell cycle arrest and apoptosis, and
disrupted the MTP in murine mammary carcinoma cells by regulating
the Wnt/β-catenin signaling pathway. It was revealed that BITC
inhibited the proliferation of 4T1-Luc cells in a
concentration-dependent manner, and that it induced cell cycle
arrest in 4T1-Luc cells at the G2/M phase. In addition, it was
further demonstrated that BITC induced apoptosis in murine mammary
carcinoma cells in vitro. To validate these findings, the
induction of apoptosis in tumor samples was investigated. TUNEL
staining demonstrated that the apoptotic rate was significantly
increased in BITC-treated tumor samples. The expression levels of
β-catenin in tumor samples treated with BITC were significantly
decreased, whereas those of APC were increased. Collectively, the
results suggested that BITC inhibited tumor growth and induced
apoptosis in murine mammary carcinoma in vitro and in
vivo, and that the Wnt/β-catenin pathway was involved in the
antitumor effects of BITC. As previously reported (7–10),
BITC exerts antitumor properties in the absence of notable side
effects or toxicity. Consistent with this, it was demonstrated that
BITC did not significantly affect the survival or morphology of
normal mammary epithelial cells in vitro.
A previous study reported that
homeodomain-interacting protein kinase-2 knockdown induced Wnt
signaling activation and β-catenin nuclear localisation, promoted
cell invasion and decreased E-cadherin expression (35). Paired-related homeobox 1 is an EMT
inducer that has been demonstrated to promote EMT in gastric cancer
cells via the activation of the Wnt/β-catenin signaling pathway,
and the regulation of the EMT molecular markers E-cadherin and
vimentin (36). These studies
indicated that the Wnt signaling pathway promoted metastasis. In
the present study, wound healing, Matrigel-invasion and migration
assays were performed, and it was observed that BITC inhibited the
migration of murine mammary carcinoma cells. This was further
supported by the in vivo results. Western blot analyses
indicated that the expression levels of E-cadherin were increased,
and those of vimentin were decreased following BITC treatment.
Thus, the present findings suggested that BITC may inhibit the
metastasis of murine mammary carcinoma cells by regulating the
Wnt/β-catenin signaling pathway, and by regulating the expression
levels of E-cadherin and vimentin.
In conclusion, the present study demonstrated that
BITC induced antitumorigenic effects, inhibiting proliferation,
inducing cell cycle arrest, inhibiting metastasis and inducing the
apoptosis of murine mammary carcinoma cells, but did not induce
toxicity or side effects in normal mammary epithelial cells. In
addition, the data indicated that the Wnt/β-catenin signaling
pathway was activated in response to BITC treatment, providing
insight into the molecular mechanisms underlying the effects of
BITC on murine mammary carcinoma 4T1-Luc cells in vitro and
in vivo. Despite the current data on the antitumor actions
of BITC, further studies involving knockdown or overexpression
models of the Wnt/β-catenin signaling pathway are required to
demonstrate the exact roles of the Wnt/β-catenin pathway in the
induction of apoptosis or the inhibition of metastasis mediated by
BITC. As BITC was reported to phosphorylate β-catenin, decrease
β-catenin levels, and downregulate Wnt2, cyclin D1 and c-Myc, it
was suggested that BITC exerted its antitumor properties by
regulating the Wnt/β-catenin signaling pathway; however, previous
studies indicated that the expression levels of cyclin D1, c-myc
and β-catenin were regulated by specificity protein (Sp)
transcription factors during antitumor processes (37). Therefore, whether BITC regulates Sp
to mediate its antitumorigenic effects also requires further
investigation.
Acknowledgements
The authors would like to thank Dr Dimiter Avtanski
of Hofstra Northwell School of Medicine (New York, USA) for his
meticulous language editing and revision of the manuscript.
Funding
The present work was supported by the Fundamental
Research Funds of the Central Universities from the Lanzhou
University (grant no. lzujbky-2017-142), the Science and Technology
Planning Project from Chengguan District, Lanzhou, Gansu Province,
China (grant no. 2016-7-11) and the Natural Science Fund of Gansu
(grant no. 17JR5RA192). These funds were used to design the study
and collect the data.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HLW made substantial contributions towards the
conception and design of the study, revised the paper critically
and provided approval for the final version to be submitted. BX
planned the study, designed and performed the experiments, acquired
the data, analyzed the results and drafted the manuscript. LZ, LLG,
HL, SYF, WJF, LL, JC, BW and LLF participated in performing the
experiments, interpreting the data, and drafting and revising the
manuscript.
Ethics approval and consent to
participate
All the animal procedures were ethically approved by
the Laboratory Animal Science and Technology Management Committee
of the Lanzhou University School of Basic Medical Sciences, and
conducted in accordance with the Guide for Care and Use of
Laboratory Animals (8th Edition).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Avtanski DB, Nagalingam A, Bonner MY,
Arbiser JL, Saxena NK and Sharma D: Honokiol inhibits
epithelial-mesenchymal transition in breast cancer cells by
targeting signal transducer and activator of transcription
3/Zeb1/E-cadherin axis. Mol Oncol. 8:565–580. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nagalingam A, Arbiser JL, Bonner MY,
Saxena NK and Sharma D: Honokiol activates AMP-activated protein
kinase in breast cancer cells via an LKB1-dependent pathway and
inhibits breast carcinogenesis. Breast Cancer Res. 14:R352012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen H and Liu RH: Potential mechanisms of
action of dietary phytochemicals for cancer prevention by targeting
cellular signaling transduction pathways. J Aqric Food Chem.
66:3260–3276. 2018. View Article : Google Scholar
|
|
4
|
Gründemann C and Huber R: Chemoprevention
with isothiocyanates-from bench to bedside. Cancer Lett. 414:26–33.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Soundararajan P and Kim JS:
Anti-carcinogenic glucosinolates in cruciferous vegetables and
their antagonistic effects on prevention of cancers. Molecules.
23(pii): E29832018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jutooru I, Guthrie AS, Chadalapaka G,
Pathi S, Kim K, Burghardt R, Jin UH and Safe S: Mechanism of action
of phenethylisothiocyanate and other reactive oxygen
species-inducing anticancer agents. Mol Cell Biol. 34:2382–2395.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Warin R, Chambers WH, Potter DM and Singh
SV: Prevention of mammary carcinogenesis in MMTV-neu mice by
cruciferous vegetable constituent benzyl isothiocyanate. Cancer
Res. 69:9473–9480. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ni WY, Hsiao YP, Hsu SC, Hsueh SC, Chang
CH, Ji BC, Yang JS, Lu HF and Chuang JG: Oral administration of
benzyl-isothiocyanate inhibits in vivo growth of subcutaneous
xenograft tumors of human malignant melanoma A375.S2 cells. In
Vivo. 27:623–626. 2013.PubMed/NCBI
|
|
9
|
Kim KJ, Hong JE, Eom SJ, Lee JY and Park
JH: Oral administration of benzyl-isothiocyanate inhibits solid
tumor growth and lung metastasis of 4T1 murine mammary carcinoma
cells in BALB/c mice. Breast Cancer Res Treat. 130:61–71. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim M, Cho HJ, Kwon GT, Kang YH, Kwon SH,
Her S, Park T, Kim Y, Kee Y and Park JH: Benzyl isothiocyanate
suppresses high-fat diet-stimulated mammary tumor progression via
the alteration of tumor microenvironments in obesity-resistant
BALB/c mice. Mol Carcinog. 54:72–82. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu X, Takano C, Shimizu T, Yokobe S,
Abe-Kanoh N, Zhu B, Nakamura T, Munemasa S, Murata Y and Nakamura
Y: Inhibition of phosphatidylinositide 3-kinase ameliorates
antiproliferation by benzyl isothiocyanate in human colon cancer
cells. Biochem Biophys Res Commun. 491:209–216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xie B, Nagalingam A, Kuppusamy P, Muniraj
N, Langford P, Győrffy B, Saxena NK and Sharma D: Benzyl
Isothiocyanate potentiates p53 signaling and antitumor effects
against breast cancer through activation of p53-LKB1 and p73-LKB1
axes. Sci Rep. 7:400702017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu M, Li W, Dong X, Chen Y, Lu Y, Lin B,
Guo J and Li M: Benzyl-isothiocyanate induces apoptosis and
inhibits migration and invasion of hepatocellular carcinoma cells
in vitro. J Cancer. 8:240–248. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kasiappan R, Jutooru I, Karki K, Hedrick E
and Safe S: Benzyl isothiocyanate (BITC) induces reactive oxygen
species-dependent repression of STAT3 protein by down-regulation of
specificity proteins in pancreatic cancer. J Biol Chem.
291:27122–27133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lai KC, Hsiao YT, Yang JL, Ma YS, Huang
YP, Chiang TA and Chung JG: Benzyl isothiocyanate and phenethyl
isothiocyanate inhibit murine melanoma B16F10 cell migration and
invasion in vitro. Int J Oncol. 51:832–840. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma YS, Hsiao YT, Lin JJ, Liao CL, Lin CC
and Chung JG: Phenethyl iosthiocyanate (PEITC) and Benzyl
isothiocyanate (BITC) inhibit human melanoma A375.S2 cell migration
and invasion by affecting MAPK signaling pathway in vitro.
Anticancer Res. 37:6223–6234. 2017.PubMed/NCBI
|
|
17
|
Duchartre Y, Kim YM and Kahn M: The Wnt
signaling pathway in cancer. Crit Rev Oncol Hematol. 99:141–149.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee MA, Kim WK, Park HJ, Kang SS and Lee
SK: Anti-proliferative activity of hydnocarpin, a natural lignan,
is associated with the suppression of Wnt/β-catenin signaling
pathway in colon cancer cells. Bioorg Med Chem Lett. 23:5511–5514.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schmitz Y, Rateitschak K and Wolkenhauer
O: Analysing the impact of nucleo-cytoplasmic shuttling of
β-catenin and its antagonists APC, Axin and GSK3 on Wnt/β-catenin
signaling. Cell Signal. 25:2210–2221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bilir B, Kucuk O and Moreno CS: Wnt
signaling blockage inhibits cell proliferation and migration, and
induces apoptosis in triple-negative breast cancer cells. J Transl
Med. 11:2802013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu QQ, Chen K, Ye Q, Jiang XH and Sun YW:
Oridonin inhibits pancreatic cancer cell migration and
epithelial-mesenchymal transition by suppressing Wnt/β-catenin
signaling pathway. Cancer Cell Int. 16:572016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Y, Zhang L, Meng Y and Huang L: Benzyl
isothiocyanate inhibits breast cancer cell tumorigenesis via
repression of the FoxH1-Mediated Wnt/β-catenin pathway. Int J Clin
Med. 8:17601–17611. 2015.
|
|
23
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals. 8th.
National Academies Press (US); 2011
|
|
24
|
Chen J, Cheng J, Yi J, Xie B, Lin L, Liu
Z, Zhao H, Wang B, Ai Z, Yang Y and Wei H: Differential expression
and response to arsenic stress of MRPs and ASAN1 determine
sensitivity of classical multidrug-resistant leukemia cells to
arsenic trioxide. Leuk Res. 50:116–122. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kocatürk B and Versteeg HH: Orthotopic
injection of breast cancer cells into the mammary fat pad of mice
to study tumor growth. J Vis Exp. Feb 8–2015.doi: 10.3791/51967.
View Article : Google Scholar
|
|
27
|
Rios Garcia M, Steinbauer B, Srivastava K,
Singhal M, Mattijssen F, Maida A, Christian S, Hess-Stumpp H,
Augustin HG, Müller-Decker K, et al: Acetyl-CoA Carboxylase
1-dependent protein acetylation controls breast cancer metastasis
and recurrence. Cell Metab. 26:842–855.e5. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou J, Tao D, Xu Q, Gao Z and Tang D:
Expression of E-cadherin and vimentin in oral squamous cell
carcinoma. Int J Clin Exp Pathol. 8:3150–3154. 2015.PubMed/NCBI
|
|
29
|
Huang SH, Wu LW, Huang AC, Yu CC, Lien JC,
Huang YP, Yang JS, Yang JH, Hsiao YP, Wood WG, et al: Benzyl
isothiocyanate (BITC) induces G2/M phase arrest and apoptosis in
human melanoma A375.S2 cells through reactive oxygen species (ROS)
and both mitochondria-dependent and death receptor-mediated
multiple signaling pathways. J Agric Food Chem. 60:665–675. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hedrick E, Cheng Y, Jin UH, Kim K and Safe
S: Specificity protein (Sp) transcription factors Sp1, Sp3 and Sp4
are non-oncogene addiction genes in cancer cells. Oncotarget.
7:22245–22256. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Valencia A, Román-Gómez J, Cervera J, Such
E, Barragán E, Bolufer P, Moscardó F, Sanz GF and Sanz MA: Wnt
signaling pathway is epigenetically regulated by methylation of Wnt
antagonists in acute myeloid leukemia. Leukemia. 23:1658–1666.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mahmood S, Bhatti A, Syed NA and John P:
The microRNA regulatory network: A far-reaching approach to the
regulate the Wnt signaling pathway in number of diseases. J Recept
Signal Transduct Res. 36:310–318. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu Y, Wei C and Xi Z: Curcumin suppresses
proliferation and invasion in non-small cell lung cancer by
modulation of MTA1-mediated Wnt/β-catenin pathway. In Vitro Cell
Dev Biol Anim. 50:840–850. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Zhang X, Liu X, Tan Z, Yang C, Ding
X, Hu X, Zhou J, Xiang S, Zhou C and Zhang J: Caudatin induces cell
apoptosis in gastric cancer cells through modulation of
Wnt/β-catenin signaling. Oncol Rep. 30:677–684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tan M, Gong H, Zeng Y, Tao L, Wang J,
Jiang J, Xu D, Bao E, Qiu J and Liu Z: Downregulation of
homeodomain-interacting protein kinase-2 contributes to bladder
cancer metastasis by regulating Wnt signaling. J Cell Biochem.
115:1762–1767. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guo J, Fu Z, Wei J, Lu W, Feng J and Zhang
S: PRRX1 promotes epithelial-mesenchymal transition through the
Wnt/β-catenin pathway in gastric cancer. Med Oncol. 32:3932015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pathi S, Jutooru I, Chadalapaka G, Nair V,
Lee SO and Safe S: Aspirin inhibits colon cancer cell and tumor
growth and downregulates specificity protein (Sp) transcription
factors. PLoS One. 7:e482082012. View Article : Google Scholar : PubMed/NCBI
|