Introduction
In 2013, the World Health Organization (WHO)
reported that 235 million people suffered from asthma (1). The inadequate control of asthma is a
serious problem (2). Asthma is a
pulmonary disease that can lead to apnea and death due to excessive
mucus production, goblet cell hyperplasia, epithelial cell
shedding, basement membrane thickening, and eosinophil and
lymphocyte infiltration (3). The
different allergens that are the inducers of asthma can be
classified into two main categories: Indoor and outdoor allergens.
The indoor allergens include pet dander, domestic mites, and
cockroaches, while the outdoor allergens include tobacco smoke,
chemical irritants, and dust particles (4).
The re-uptake of the allergens can cause an
imbalance in the T-helper (Th) cells and eventually result in
asthma (3). Various studies have
revealed that Th type 1 (Th1)-related cytokines [interleukin
(IL)-12 and interferon (IFN)-γ], Th type 2 (Th2)-related cytokines
(IL-4, IL-5, and IL-13), and other proinflammatory cytokines
(IL-1β, IL-6, and TNF-α) are associated with asthma. Of these,
IFN-γ has been revealed to have an important role in suppressing
asthma induction by regulating the increase in the level of IgE
(5) and accelerating the
activation of T-bet, which is a Th1 cell transcription factor, in a
positive feedback loop (6,7). IL-12 has been demonstrated to
modulate the balance between Th1 and Th2 cells not only by
promoting Th1 cell proliferation and inhibiting Th2 cell
proliferation, but also by accelerating IFN-γ production (8,9).
IL-4 has been reported to upregulate the level of IgE, increase
eosinophil count (10), and induce
GATA-3 activation (11,12). IL-5 has been revealed to have
several functions related to the development, activation,
migration, and survival of eosinophils, in addition to augmentation
of IL-6 proliferation (9,13). IL-13 has been demonstrated to not
only modulate B cell activation but also induce airway remodeling,
which causes excessive mucus production, goblet cell hyperplasia,
epithelial cell shedding, basement membrane thickening, and
eosinophil and lymphocyte infiltration in asthma (14–17).
IL-6 is one of the proinflammatory cytokines which has been
revealed to promote the expression of IgE and modulate the
activation of Th cells. TNF-α, on the contrary, has been
demonstrated to recruit granulocytes and induce fibroblast
proliferation (18).
Inhaled corticosteroids are usually used for the
treatment of asthma and are often administered as a combination
therapy with long-acting β2-agonist or leukotriene
modulators, such as leukotriene receptor antagonist or
5-lipoxygenase inhibitor (19).
However, corticosteroids exhibit several severe adverse effects on
different systems, such as growth suppression in children (20); cataract and glaucoma; hypertension,
water retention, and hyperlipidemia in the cardiovascular system;
peptic ulcer and pancreatitis; and catatonia, decreased
concentration, agitation, insomnia, and behavioral changes
(21).
Effort has been escalated to identify effective and
safe drugs, especially natural products, for the treatment of
asthma (22–24). Allium hookeri is a plant
belonging to the Alliaceae family. It is commonly used for
culinary purposes in India, Thailand and China. The rhizome of this
plant is used in traditional medicine for protection against
hypertension (25). Recently,
several biological activities of A. hookeri have been
reported, such as antimicrobial effects (26), anti-inflammatory effects (27), and vascular growth factor
regulation in allergic rhinitis (28). The anti-asthmatic effects of A.
hookeri root extracts (AHRE), however, have not been studied
yet. Therefore, this study was conducted to evaluate the
anti-asthmatic activities of AHRE (30 and 300 mg/kg/day for 5 days)
in an ovalbumin (OVA)-induced asthma model using experimental
mice.
Materials and methods
Preparation of AHRE
Briefly, A. hookeri plants were collected in
May 2014 from near Naju (Korea). A voucher specimen (MNUCSS-SC-01)
was deposited at Mokpo National University (Muan, Korea). The roots
were separated, air-dried, powdered (250 g), and extracted twice
with 70% ethanol (1 l) at room temperature for 3 days. Following
filtration, the ethanol was evaporated and the extract was dried
and stored at −50°C. Yield of extraction was 9% (w/w).
Animal experiments
All the experiments were approved by the
Institutional Animal Care and Use Committee at Dongshin University,
Naju, Korea (approval no. 2014-08-03). Eighty female BALB/c mice
were purchased from Samtako Korea (Osan, Korea) and divided into
five groups that received different treatment drugs as follows: i)
Control, receiving vehicle (sterile tap water); ii) OVA for asthma
induction; iii) OVA for asthma induction plus 1 mg/kg/day
dexamethasone (positive control); iv) OVA for asthma induction plus
30 mg/kg/day AHRE; and v) OVA for asthma induction plus 300
mg/kg/day AHRE. Dexamethasone was selected as a positive control
since it is broadly used for the treatment of asthma.
On days 1 and 8, all the groups of mice except the
control were exposed to intraperitoneal OVA (20 µg) containing 1 mg
aluminum hydroxide hydrate (both from Sigma-Aldrich; Merck KGaA) as
an adjuvant in 500 µl saline. From days 21 to 25, the mice were
challenged once daily with 5% OVA for 30 min using a nebulizer
(NE-U17; OMRON Co. Ltd.) at a flow rate of 3 ml/min. During the
same 5-day period, the other groups of mice were also treated once
daily with oral doses of sterile tap water, dexamethasone, 30
mg/kg/day AHRE, and 300 mg/kg/day AHRE, respectively 1 h prior to
the OVA challenge. The control group was sensitized to OVA
according to the same procedure as the other groups, following
which they were exposed to saline and aluminum hydroxide hydrate by
a nebulizer for 5 consecutive days.
BALF and serum analysis
One day after the final treatment, the mice were
anesthetized with intraperitoneal injections of 50 mg/kg Zoeltil
100 (Virbac Corporation), and the tracheas were cannulated with
disposable animal feeding needles. Bronchoalveolar lavages were
performed with three 0.4 ml aliquots of cold phosphate-buffered
saline (PBS). The BALF samples were collected and immediately
centrifuged at 900 × g for 5 min (Sorvall Legend Micro 17R; Thermo
Fisher Scientific, Inc.). The cell pellets were re-suspended in PBS
for WBC and differential cell counts. The total WBC and
differential cell count were performed by a Hemavet Multispecies
Hematology System (Drew Scientific, Inc.). The serum IgE levels
were estimated using a specific mouse IgE enzyme-linked
immunosorbent assay kit (cat. no. 555248; BD Biosciences) according
to the manufacturer's instructions.
Histopathological examinations
The lung tissues were fixed in 10% (v/v)
formaldehyde solution, dehydrated in a graded concentration of
ethanol (99.9, 90, 80, and 70%), at room temperature for 1 month
and embedded in paraffin at room temperature for 2 weeks. The
paraffin-embedded lung tissues were then sectioned longitudinally
(5 µm) and in order to analyze semi-quantitatively glycopreoteins
expression they were stained with hematoxylin and eosin (H&E)
at room temperature for 6 min, and with Periodic acid-Schiff stain
at room temperature for 12 min. The images were captured with an
Axioscope A1 microscope (Carl Zeiss AG). In order to define the
change of histopathological morphology on mucous hypersecretion,
epithelial cell hyperplasia, and inflammatory cell infiltration
scores from 0 (none) to 3 (severe) were assigned.
Immunohistochemical examinations
Immunohistochemical examinations were performed by a
method previously described (24).
The de-paraffinized tissue sections were treated with 3% hydrogen
peroxide in methanol for 10 min to remove the endogenous
peroxidase. Antigen retrieval was carried out with sodium citrate
buffer (0.1 M) using the hot plate method. The slides were
incubated with normal serum to block the nonspecific bindings and
then incubated for 1 h at 4°C with different primary antibodies
(diluted 1:100 to 1:200), such as rat anti-mouse CD4 monoclonal
(cat. no. 14-9766; eBioscience; Thermo Fisher Scientific, Inc.),
rabbit anti-mouse CD8 polyclonal (cat. no. MBS551004; Mybiosource,
Sand Diego, CA, USA), rabbit anti-mouse CD19 polyclonal (cat. no.
250585; Abbiotec), rabbit anti-mouse CD68 polyclonal (cat. no.
bs-0649R; Bioss, Inc.), rat anti-mouse MHC class II monoclonal
(cat. no. sc-59318cat), rat anti-mouse IFN-γ monoclonal (cat. no.
sc-74104), goat anti-mouse IL-12 Ap35 polyclonal (cat. no.
sc-9350), rat anti-mouse IL-12p40 monoclonal (cat. no. sc-57258),
rat anti-mouse IL-4 monoclonal (cat. no. sc-73318), rabbit
anti-mouse IL-5 polyclonal (cat. no. sc-7887), goat anti-mouse
IL-13 polyclonal (sc-1776), goat anti-mouse IL-6 polyclonal (cat.
no. sc-1265; all from Santa Cruz Biotechnology, Inc.), and rabbit
anti-mouse TNF-α polyclonal antibody (cat. no. 3053R-100;
BioVision, Inc.). The slides were then incubated with a
biotinylated pan-specific secondary antibody for 10 min and reacted
with a streptavidin-peroxidase complex for 5 min (Universal Quick
HRP Kit; cat. no. PK-7800; Vector Laboratories, Inc.). The signals
were detected using 3,3-diaminobenzidine tetrahydrochloride
substrate chromogen solution, and the cells were counterstained
with Mayer's hematoxylin. The cells were imaged using an Axioscope
A1 microscope (Carl Zeiss AG). To determine the number of
positively stained cells, the cells in five randomly selected
non-overlapping fields (×200 magnification) of three separately
immunostained lung sections per mouse (n=8/group) were counted.
Immunofluorescence analysis
In order to localize the Th1 cell transcription
factor (T-bet), and Th2 cell transcription factor (GATA-3),
immunofluorescence analysis was conducted in four groups of mice
such as the control, the OVA for asthma induction, the OVA for
asthma induction plus dexamethasone, and the OVA dexamethasone plus
300 mg/kg/day AHRE groups. The steps prior to the antibody binding
step were similar to that of the immunohistochemistry method except
for rabbit anti-mouse T-bet (cat. no. orb7075; Biorbyt Ltd.) or
goat anti-mouse GATA-3 (cat. no. TA305795; OriGene Technologies,
Inc.) were used as primary antibodies for 1 h at a room
temperature. The slides were incubated for 2 h with FITC-conjugated
anti-rabbit IgG (cat. no. 315-095-003; Jackson Immunoresearch
Laboratories, Inc.) or Alexa Fluor 555-conjugated anti-goat IgG
antibody (cat. no. A-21127; Thermo Fisher Scientific, Inc.) and the
cells were counterstained with 4′,6-diamidino-2-phenylindole (DAPI)
(cat. no. 62249; Thermo Fisher Scientific, Inc.). The images were
obtained using a K1-Fluo confocal microscope (Nanoscope Systems,
Inc.).
Statistical analysis
The results were expressed as the mean ± standard
deviation (SD). The group differences were evaluated by one-way
analysis of variance, followed by Dunnett's multiple comparison
post hoc test. A P-value of <0.05 was considered to indicate a
statistically significant difference.
Results
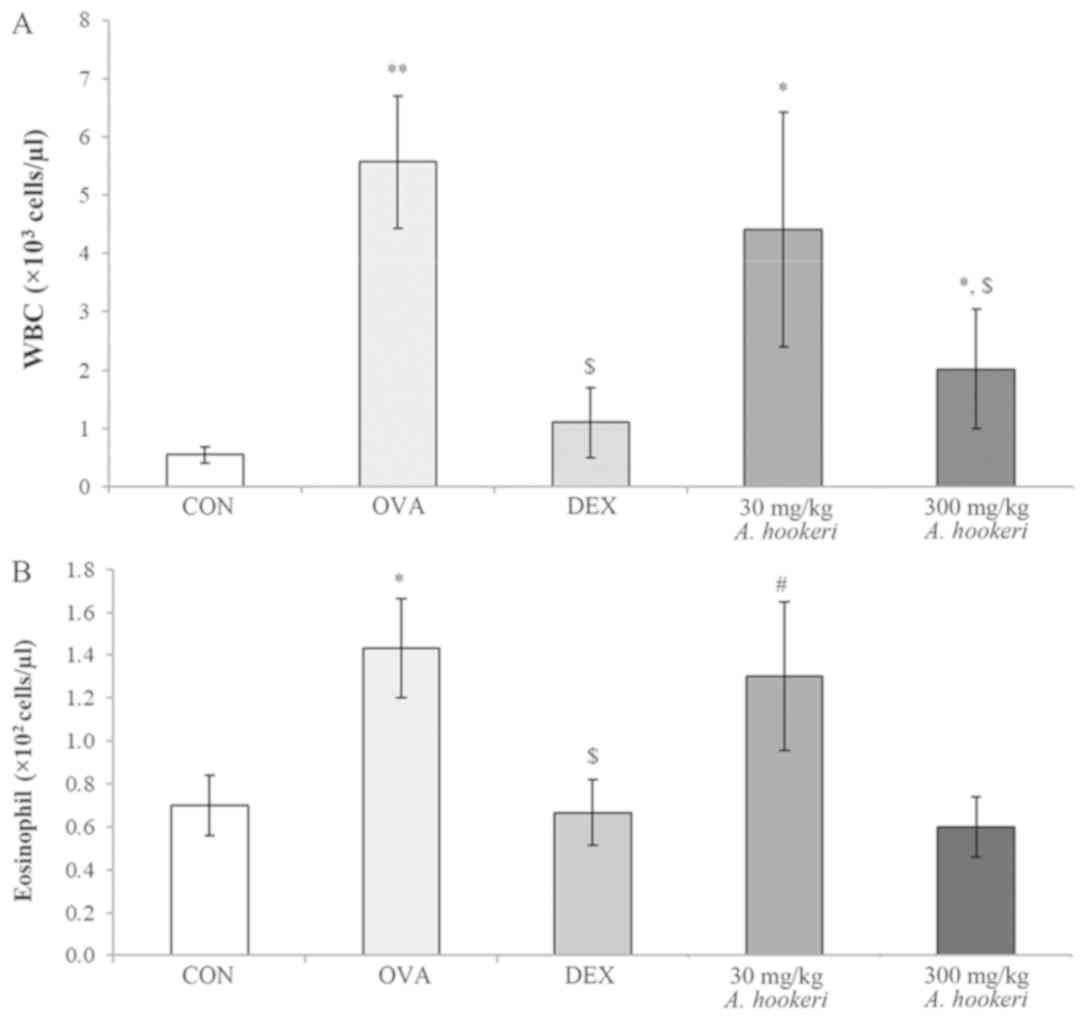
AHRE significantly suppresses WBC
count, especially eosinophils in bronchoalveolar lavage fluid
(BALF)
The WBC count, and especially the eosinophil count,
in BALF were significantly increased in the OVA-treated group
(Fig. 1). As revealed in Fig. 1A, the WBC count in the OVA-treated
group was significantly increased, but was considerably reduced by
dexamethasone treatment, which was selected as a positive control
since it is broadly used for asthma treatment. AHRE reduced the
OVA-induced increase in WBC count in a dose-dependent manner. The
eosinophil count was also reduced by AHRE (Fig. 1B). Based on the eosinophil count in
the OVA-treated group (100%), the reduction in the dexamethasone
group was 53% and that in the 300 mg/kg AHRE group was 58%.
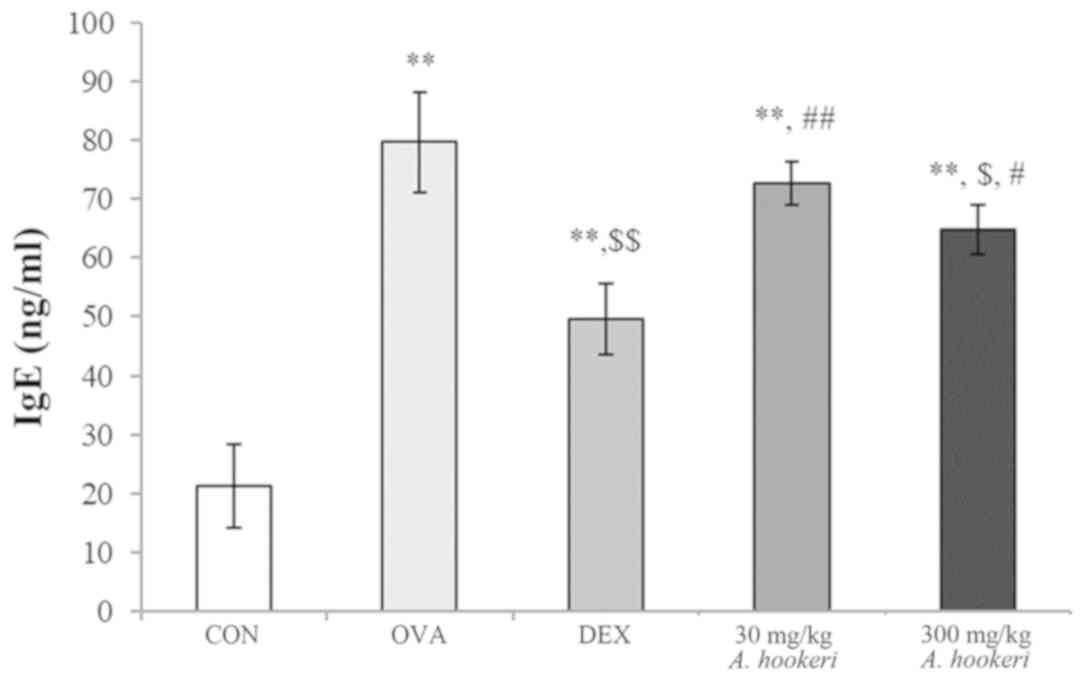
AHRE downregulates the OVA-induced
increase in serum IgE
The change in the level of IgE was similar to that
of WBC (Fig. 2). The level of IgE
in the OVA-treated group was significantly increased, which was
reduced by dexamethasone. AHRE decreased the levels of IgE in a
dose-dependent manner, and the effect of administering 300 mg/kg
AHRE was similar to that of dexamethasone treatment.
AHRE ameliorates asthmatic changes in
the lungs
In the lungs of the OVA-treated group, typical
histopathological changes associated with asthma were observed,
such as mucus hypersecretion, epithelial hyperplasia, goblet cell
hyperplasia, and eosinophil infiltration near the bronchioles and
vessels (Fig. 3A-b), when compared
with that in the lungs of the control group animals (Fig. 3A-a). Dexamethasone ameliorated the
OVA-induced morphological changes, as evidenced by the presence of
only a sparse eosinophilic infiltration (Fig. 3A-c). In the lungs of the 30 mg/kg
AHRE-treated group, asthma-like typical morphological changes were
observed (Fig. 3A-d), whereas, in
the 300 mg/kg AHRE-treated group, only a sparse eosinophilic
infiltration was observed (Fig.
3A-e) similar to that in the dexamethasone-treated group
(Fig. 3A-c).
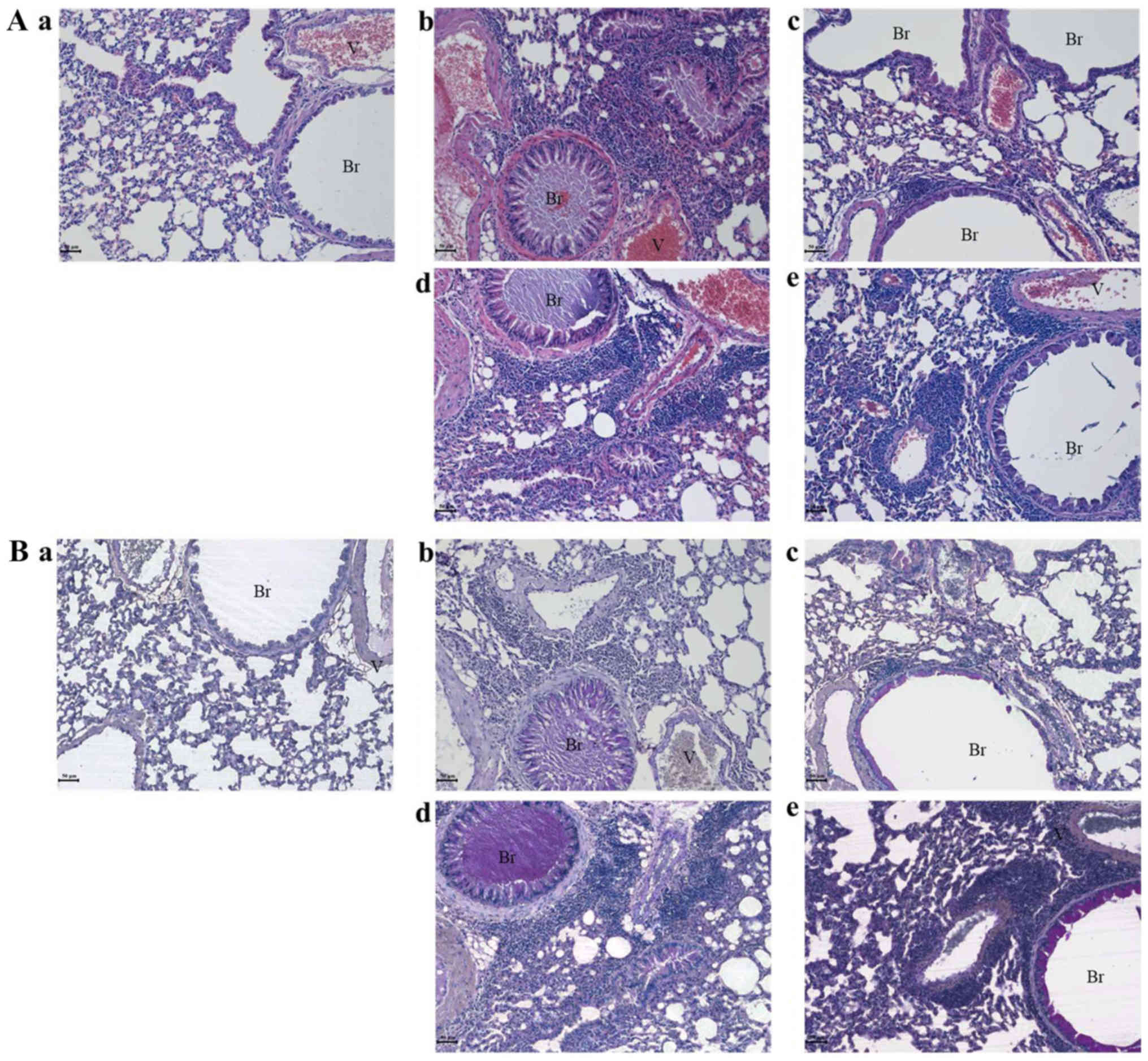 | Figure 3.AHRE ameliorates OVA-induced
asthmatic changes in the lungs in a dose-dependent manner. AHRE
ameliorated the asthmatic changes in mouse lungs, such as mucus
hypersecretion, eosinophil infiltration near the bronchioles and
vessels, epithelial cell hyperplasia, and goblet cell hyperplasia
as observed by (A) hematoxylin and eosin staining. (B) Periodic
acid-Schiff staining confirmed a protective effect by AHRE against
mucus hypersecretion (scale bar, 50 µm). Br, bronchiole; V, vessel;
a, vehicle control; b, asthma induction; c, dexamethasone; d, AHRE
30 mg/kg/day; and e, AHRE 300 mg/kg/day. AHRE, Allium
hookeri root extract; CON, control; OVA, ovalbumin; DEX,
dexamethasone. |
In order to assess mucus secretion, Periodic-Schiff
staining was performed (Fig. 3B).
Mucus hypersecretion, also known as mucus plugs, was observed in
the bronchioles (Fig. 3B-b) in the
OVA-treated group, when compared with the control (Fig. 3B-a). Dexamethasone (Fig. 3B-c) and AHRE, in a dose-dependent
manner, (Fig. 3B-d and-e)
decreased the OVA-induced mucus hypersecretion. Especially, in the
300 mg/kg AHRE-treated group, mucus was almost undetected in the
bronchioles and was observed only near the epithelium (Fig. 3B-e).
In order to compare the morphological change level
of each group, quantitative analysis was conducted and the results
revealed that AHRE effectively suppressed the OVA-induced
histopathological changes in the lung (Table I).
 | Table I.The quantitative results of
histopathological changes by AHRE. |
Table I.
The quantitative results of
histopathological changes by AHRE.
| Groups | Mucous
hypersecretion (0–3) | Epithelial cell
hyperplasia (0–3) | Inflammatory cell
infiltration (0–3) |
|---|
| CON | 0.3±0.46 | 0.3±0.46 | 0.3±0.46 |
| OVA |
2.8±0.46b |
2.9±0.35b |
2.8±0.46b |
| DEX |
0.4±0.52c |
0.4±0.52c |
0.4±0.52c |
| 30 mg/kg A.
hookeri |
2.8±0.46b,e |
2.9±0.35b,e |
2.4±0.52b,e |
| 300 mg/kg A.
hookeri |
0.5±0.53c |
1.0±0.76a,c |
1.4±0.52b–d |
AHRE suppresses expression of T
cell-related factors, macrophages, and major histocompatibility
complex (MHC) class II molecules in asthma
The immune modulatory effects of AHRE were
investigated by analyzing the changes in the population of T cells
(CD4+ Th cells, Fig.
4A, and CD8+ cytotoxic T cells, Fig. 4B), B cells (CD19+ cells,
Fig. 4C), macrophages
(CD68+ cells, Fig. 4D),
and MHC class II molecules (Fig.
4E). AHRE suppressed all these cells, and especially almost
completely inhibited the expression of CD68+ cells
(Fig. 4D-e), and MHC class
II+ molecules (Fig.
4E-e) in a dose-dependent manner. The expression of
CD4+ Th cells (Fig.
4A-e), CD8+ cytotoxic T cells (Fig. 4B-e), and CD68+ cells
(Fig. 4D-e) in the 300 mg/kg
AHRE-treated group was lower than in the dexamethasone-treated
group (Fig. 4A-c, B-c, and D-c).
The expression of CD19+ B cells (Fig. 4C-e) and MHC class II+
molecules (Fig. 4E-e) in the 300
mg/kg AHRE-treated group was similar to that in the
dexamethasone-treated group (Fig.
4E-c).
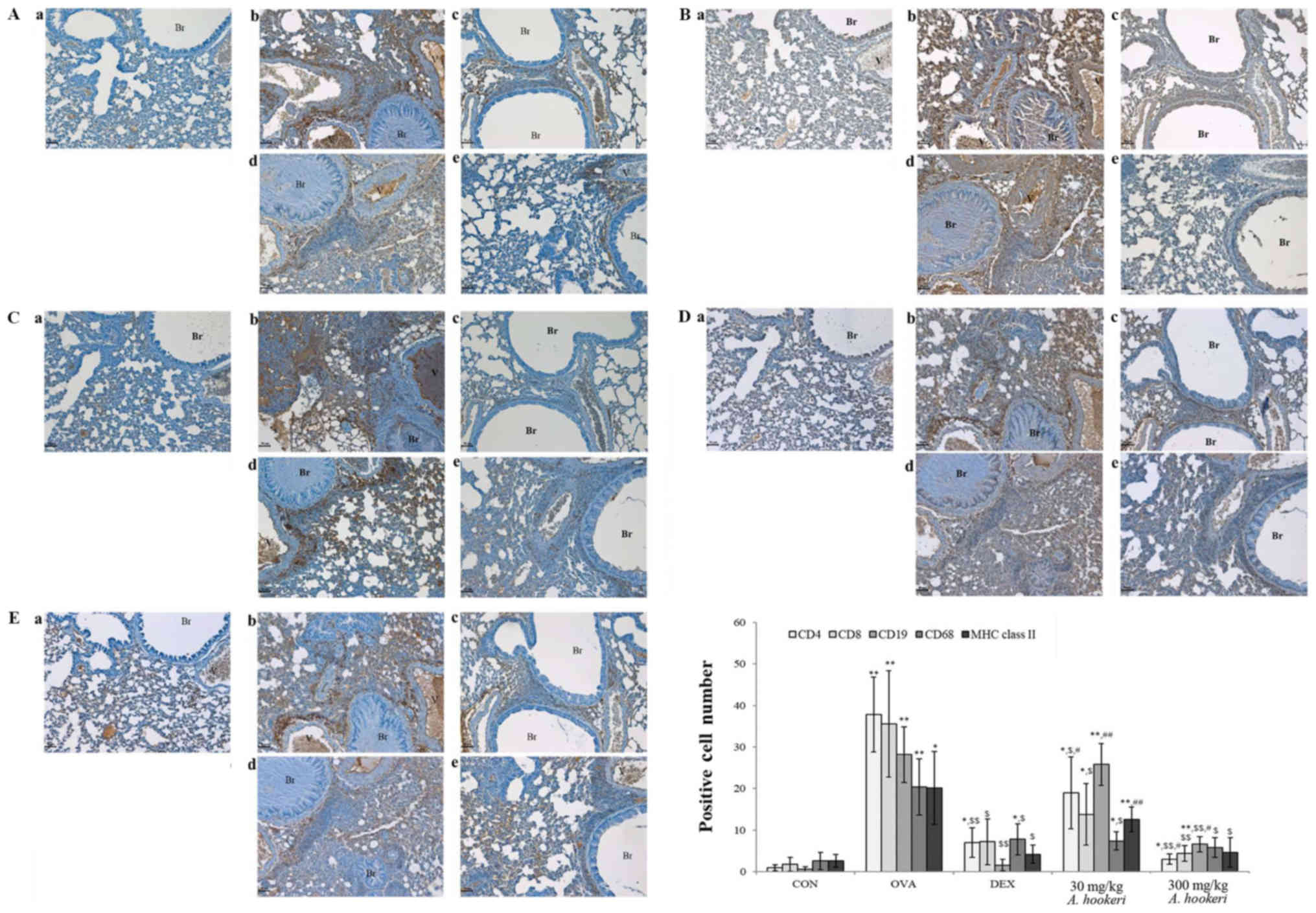 | Figure 4.AHRE suppresses the OVA-induced
increase in the expression of (A) CD4+ Th cells, (B)
CD8+ cytotoxic T cells, (C) CD19+ B cells,
(D) CD68+ cells, and (E) MHC class II+
molecules in a dose-dependent manner. The immune positive cells
were counted in five randomly selected non-overlapping fields
(×200, magnification) from three separately immunostained lung
sections per mouse (scale bar, 50 µm); a, vehicle control; b,
asthma induction; c, dexamethasone; d, AHRE 30 mg/kg/day; and e,
AHRE 300 mg/kg/day. *P<0.05 compared with the control,
**P<0.001 compared with the control, $P<0.05
compared with asthma induction (OVA-treated group),
$$P<0.001 compared with asthma induction (OVA-treated
group), #P<0.05 compared with dexamethasone, and
##P<0.001 compared with dexamethasone. AHRE,
Allium hookeri root extract; CON, control; OVA, ovalbumin;
DEX, dexamethasone. |
AHRE suppresses the expression of
Th1-related cytokines (IFN-γ and IL-12p40) but not IL-12p35
Asthma is a chronic respiratory disorder caused not
only by the imbalance between Th1- and Th2-related cytokines
(29,30) but also by changes in the activation
of several inflammatory mediators (13,18).
The changes in the asthma-related inflammatory mediators, such as
Th1-related cytokines (IFN-γ, IL-12p40, and IL-12p35), (Fig. 5) and Th2-related cytokines (IL-4,
IL-5, and IL-13) (Fig. 6) were
evaluated. All of these factors are usually secreted and detected
near the bronchioles and pulmonary vessels. In asthma, an imbalance
exists between the Th1- and Th2-related factors and the quantity of
Th1-related cytokines may be lower than that of the Th2-related
ones. In the lungs of the OVA-treated group, the expression of the
Th1-related cytokines, such as IFN-γ, IL-12p40, and IL-12p35 was
increased (Fig. 5A-b, B-b, and
C-b) compared to the control, whereas, the expression was
significantly less in the dexamethasone-treated group (Fig. 5A-c, B-c, and C-c) compared to the
OVA-treated group. AHRE decreased the expression of IFN-γ (Fig. 5A) and IL-12p40 (Fig. 5B) in a dose-dependent manner
compared to the OVA-treated group. The 300 mg/kg AHRE-treated group
almost completely inhibited the expression of these cytokines
(Fig. 5A-e). Although the
expression of IL-12p35 was affected by AHRE, a dose-dependent
effect was not observed (Fig. 5C)
compared to the OVA-treated group. And the activation of
transcription factors that regulate T-bet and induce GATA-3 were
analyzed (Fig. S1). In order to
activate the Th1 cell and Th2 cell transcription factors, T-bet and
GATA-3 exist in the nucleus, whereas, during the state of
inactivation, they localize in the cytoplasm. The distribution of
T-bet in the control, dexamethasone group, and 300 mg/kg AHRE group
were detected in the cytoplasm (Fig.
S1B-a, -c, and-d), whereas, in the OVA-treated group it was in
the nucleus (Fig. S1B-b). The
distribution of GATA-3 was similar to that of T-bet (Fig. 1SC-a to -d). AHRE inhibited T-bet
and GATA-3 activation similar to that by dexamethasone (Fig. S1).
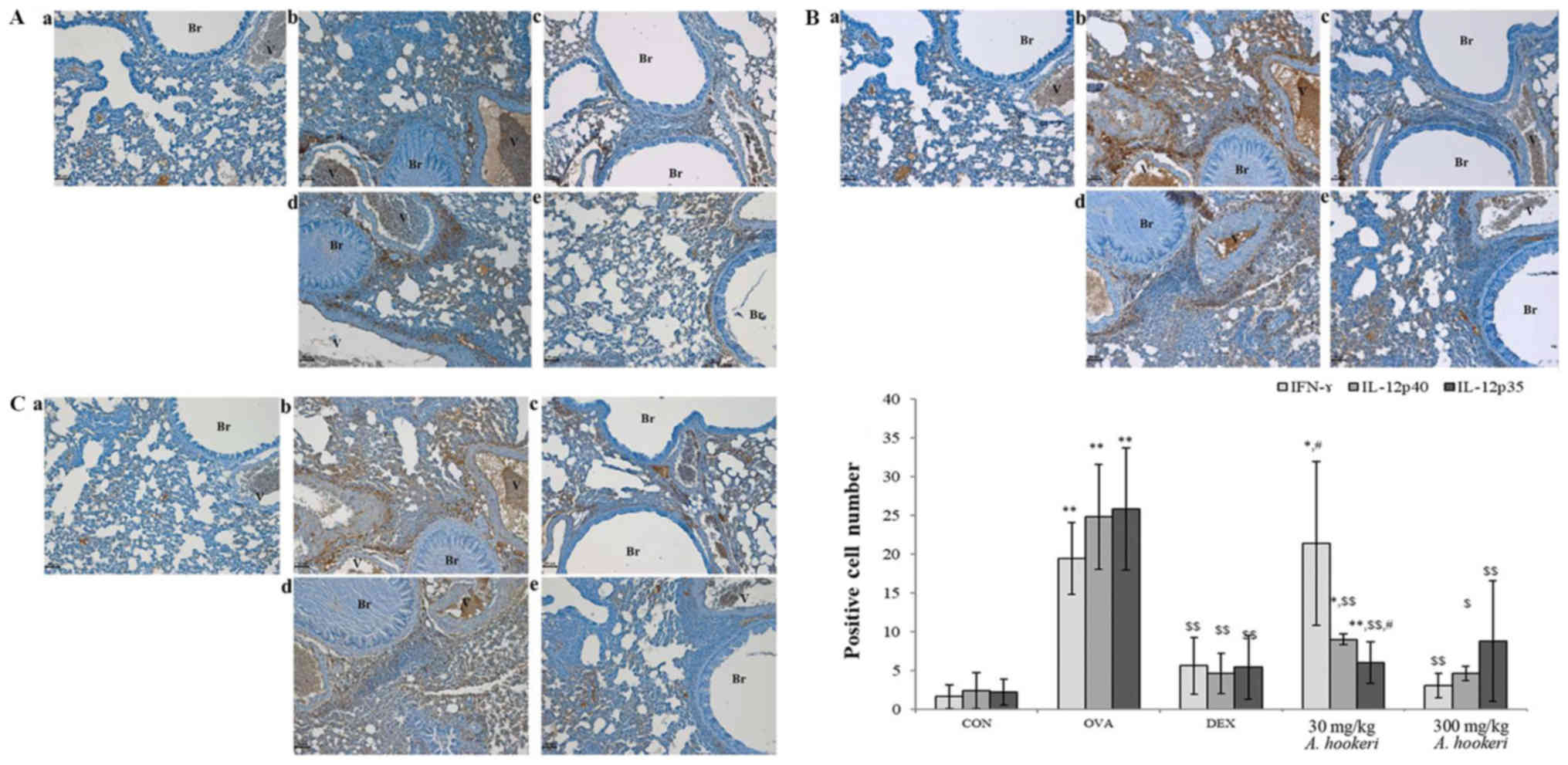 | Figure 5.AHRE reduces the expression of (A)
IFN-γ, and (B) IL-12p40 but not (C) IL-12p35 in the lungs. The
immune-positive cells were counted in five randomly selected
non-overlapping fields (×200, magnification) from three separately
immunostained lung sections per mouse (scale bar, 50 µm); a,
vehicle control; b, asthma induction; c, dexamethasone; d, AHRE 30
mg/kg/day; and e, AHRE 300 mg/kg/day. *P<0.05 compared with the
control; **P<0.001 compared with the control;
$P<0.05 compared with asthma induction (OVA-treated
group); $$P<0.001 compared with asthma induction
(OVA-treated group); and #P<0.05 compared with
dexamethasone. AHRE, Allium hookeri root extract; CON,
control; OVA, ovalbumin; DEX, dexamethasone. |
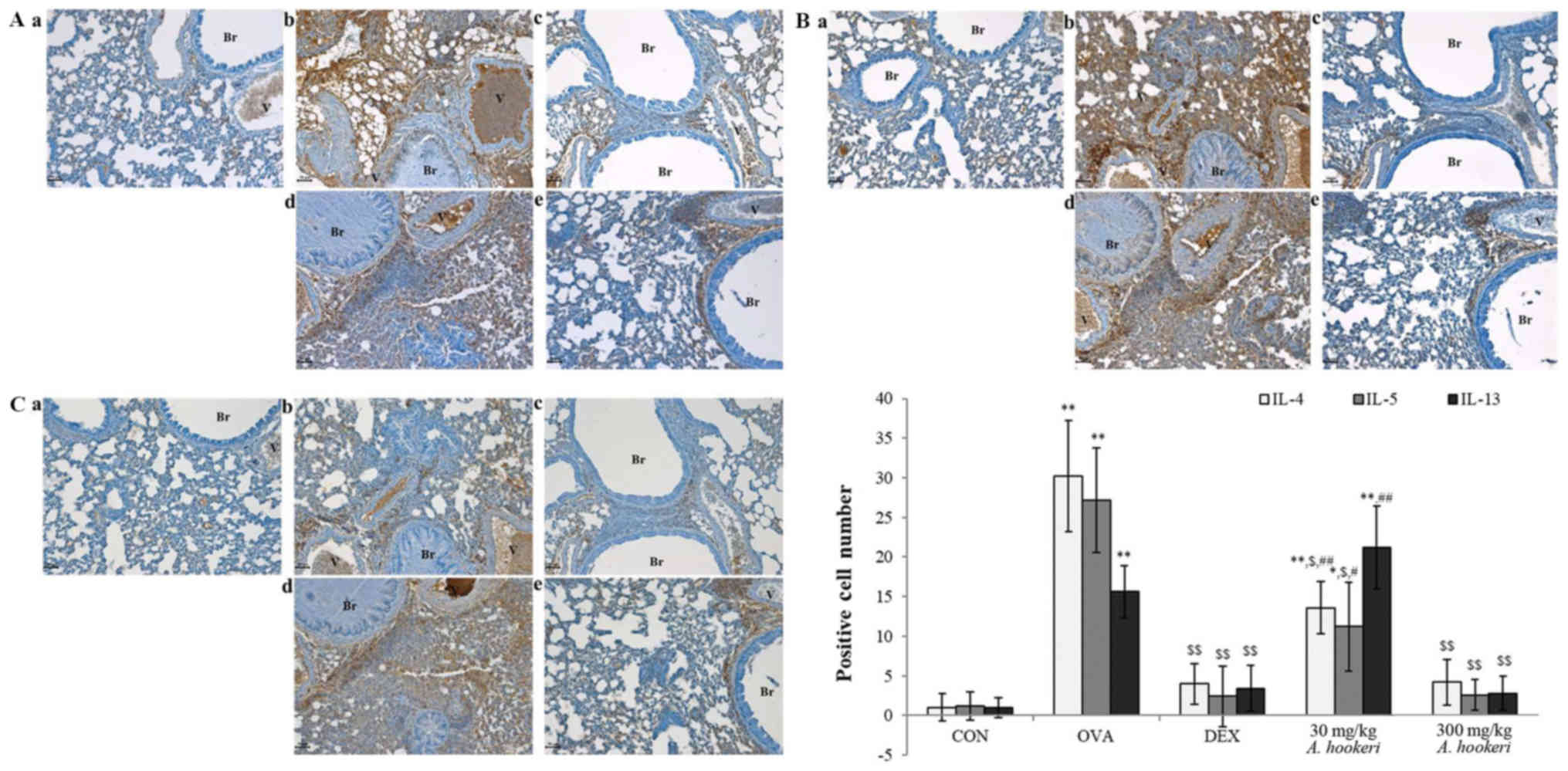 | Figure 6.AHRE significantly inhibits the
expression of (A) IL-4, (B) IL-5, and (C) IL-13 in the lungs in a
dose-dependent manner. The immune-positive cells were counted in
five randomly selected non-overlapping fields (×200, magnification)
from three separately immunostained lung sections per mouse (scale
bar, 50 µm); a, vehicle control; b, asthma induction; c,
dexamethasone; d, AHRE 30 mg/kg/day; and e, AHRE 300 mg/kg/day.
*P<0.05 compared with the control; **P<0.001 compared with
the control; $P<0.05 compared with asthma induction
(OVA-treated group); $$P<0.001 compared with asthma
induction (OVA-treated group); #P<0.05 compared with
dexamethasone; and ##P<0.001 compared with
dexamethasone. AHRE, Allium hookeri root extract; CON,
control; OVA, ovalbumin; DEX, dexamethasone. |
AHRE downregulates the expression of
Th2-related cytokines (IL-4, IL-5, and IL-13)
The change in the expression of the Th2-related
cytokines was similar to that of the Th1-related ones (Fig. 6). Although IL-4, IL-5, and IL-13
were barely detected in the control group (Fig. 6A-a, B-a, and C-a), OVA increased
the expression of these cytokines (Fig. 6A-b, B-b, and C-b) compared to the
control, and dexamethasone suppressed all of them (Fig. 6A-c, B-c, and C-c) compared to the
OVA-treated group. AHRE suppressed the expression of Th2-related
cytokines such as IL-4 (Fig. 6A),
IL-5 (Fig. 6B), and IL-13
(Fig. 6C) in a dose-dependent
manner compared to the OVA-treated group. Especially, 300 mg/kg
AHRE effectively suppressed the expression of IL-4 (Fig. 6A-e) and IL-5 (Fig. 6B-e) compared to the OVA-treated
group, similar to that by dexamethasone.
In addition to the changes of Th1-/Th2-related
cytokines the changes of proinflammatory cytokines were analyzed.
The effects of AHRE on proinflammatory cytokines, such as TNF-α and
IL-6 that play important roles in the immune regulation of asthma
were studied (Fig. S2). TNF-α was
overexpressed by OVA (Fig.
S2A-b), but significantly suppressed by dexamethasone (Fig. S2A-c). The expression of TNF-α was
decreased by AHRE in a dose-dependent manner. Especially, 300 mg/kg
AHRE suppressed the expression of TNF-α (Fig. S2A-e) to a level similar to that
achieved by dexamethasone treatment. IL-6 expression was increased
by OVA treatment (Fig. S2B-b),
whereas, dexamethasone inhibited it (Fig. S2B-c). Although the expression of
IL-6 was suppressed by AHRE, a dose-dependent effect was not
observed.
Discussion
Asthma is a type 1 hypersensitivity reaction, which
is caused by exposure to specific allergens and is related to the
activation of IgE (29). The
allergens can be classified into two main categories according to
IgE dependence (31). One of the
categories stimulates IgE production and includes pollens, animal
dander, house-dust-mite particles, venom, and foods, such as
peanuts, fish, milk, and egg. The other category is not related to
IgE and includes reactions such as allergic contact dermatitis. The
OVA-induced asthma model is related to IgE production. This model
depicts not only a surge in the immune cell populations and
inflammatory mediators but also an imbalance between the Th1- and
Th2-related factors including proinflammatory cytokines (22).
The expression of macrophages (CD68+
cells) has been revealed to be elevated (32) in asthma. The macrophages in the
respiratory system can be divided into M1 or M2 subsets according
to their role and cytokine profile (33). The M1 subset is mainly related to
anti-inflammatory effects and releases IL-12 (34), while the macrophage M2 subset
regulates inflammation and produces TNF-α (35). The alveolar macrophage contains
both the M1 and M2 subsets of macrophages. The present results
revealed that AHRE suppressed CD68+ cell expression (M2
subset).
The MHC gene family is subdivided into class I and
II in humans and mice. The MHC class II molecules initiate immune
reactions related to CD4+ cells (Th cells), whereas, the
MHC class I molecules initiate reactions related to CD8+
cells (cytotoxic T cells) (36–38).
The MHC class II molecules have important roles in the regulation
of Th cell and cytotoxic T-cell activation, and Th cell modulation
(balance between Th1 cells and Th2 cells) is important in asthma.
AHRE may control asthma through the downregulation of
CD4+ cells (Fig.
4E).
Asthma is caused by various cytokines and not by any
single factor (11). Its
pathogenic mechanism was reported to be due to the imbalance of
Th1- and Th2-related factors. IL-4 and IL-13 play key roles in
asthma induction (39) and are
responsible for the typical morphological changes such as mucus
hypersecretion, epithelial cell hyperplasia, goblet cell
hyperplasia, eosinophil infiltration, as well as the physiological
changes such as B cell activation in the pulmonary system
associated with asthma. AHRE suppressed the expression of IL-4 and
IL-13 in a dose-dependent manner. IL-4 expression was significantly
downregulated to a level similar to that of the
dexamethasone-treated group. Although AHRE suppressed most of the
asthma-related cytokines, such as IFN-γ, IL-12p40, IL-4, IL-5, and
IL-13, the mode of action may be related to T-bet (Th1 cell
transcription factor), GATA-3 (Th2 cell transcription factor),
CD68+ cells (TNF-α producer), and T-cell recognition
involving MHC class II+ molecules and CD68.
Recently, Jang et al (39) reported that AHRE suppressed
inflammation in the macrophage cells, which was induced by
lipopolysaccharide through nuclear factor (NF)-κB and
cyclooxygenase (COX)-2 downregulation. Based on these results, it
could be inferred that the hyperactivated immune cells, such as B
cells, and T cells, may be decreased by AHRE owing to its
downregulatory effects on NF-κB and COX-2.
Inhaled corticosteroids are extensively used for the
treatment of asthma. However, they exhibit numerous adverse
effects, such as growth suppression in children (20) catatonia, decreased concentration,
agitation, insomnia, and abnormal behaviors (21). Several efforts have been undertaken
to identify new drugs for treating asthma, which have no or less
adverse effects. In the present study, AHRE ameliorated
asthma-related changes as evidenced by the Th1-/Th2-related
inflammatory cytokine levels and histopathological changes. AHRE
was revealed to suppress T-cell recognition, which is an important
step in asthma induction, by the downregulation of CD68+
and MHC class II+ molecules.
In previous studies, we have identified ferulic
acid, linoleic acid, and cinnamic acid from the roots of AHRE
(40,41). Major compounds were identified as
linoleic acid ethyl ester (8.02%), hexacosane (7.5%). In addition,
minor compounds were identified as ferulic acid (0.17%) and
cinnamic acid (1.99%). It was reported that ferulic acid controls
Th2-response by decreasing lung inflammation, eosinophil
infiltration, mucus production, and serum IgE level. It also
decreases chemokines and cytokines, such as CCL5, CCL11, CCL20,
IL-4, IL-5, IL-13, IL-25, and IL-33 in the lungs. Linoleic acid is
one of the major constituents of AHRE. We previously reported that
the anti-inflammatory effects of AHRE were due to the antioxidant
and anti-inflammatory activities of linoleic acid (41). Xu et al demonstrated that
cinnamic acid suppressed the production of pro-inflammatory
cytokines and expression of NLRP3, caspase-1, and IL-1β mRNA
proteins, and also reversed the lipopolysaccharide-induced
histopathological changes in the lungs and the spleen (42).
From the results of the present study, it can be
concluded that the active compounds, such as ferulic acid, cinnamic
acid, and linoleic acid present in AHRE demonstrated effective
anti-asthmatic activities. AHRE has been used for culinary purposes
for a long time and is confirmed to be safe. Hence, AHRE can be
used as a natural source drug for treating asthma.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SHB and JHS revised the manuscript and conducted the
animal study and analysis, CSB analyzed the samples, BK read the
histopathological results, and SSC and DHP statistically analyzed
the data and revised manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All the experiments were approved by the
Institutional Animal Care and Use Committee at Dongshin University,
Naju, Korea (approval no. 2014-08-03).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
World Health Organization. Asthma Fact
Sheet No 307. 2013, . https://www.who.int/en/news-room/fact-sheets/detail/asthma31–August.
2017
|
|
2
|
Slejko JF, Ghushchyan VH, Sucher B, Globe
DR, Lin SL, Globe G and Sullivan PW: Asthma control in the United
States, 2008–2010: Indicators of poor asthma control. J Allergy
Clin Immunol. 133:1579–1587. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kay AB: Allergy and allergic diseases.
First of two parts. N Engl J Med. 344:30–37. 2000. View Article : Google Scholar
|
|
4
|
American Academy of Allergy,
AsthmaImmunology and Asthma statistics. 14–April. 2019, http://www.aaaai.org/about-the-aaaai/newsroom/asthma-statistics.aspx
|
|
5
|
Stirling RG and Chung KF: New
immunological approaches and cytokine targets in asthma and
allergy. Eur Respir J. 16:1158–1174. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Szabo SJ, Kim ST, Costa GL, Zhang X,
Fathman CG and Glimcher LH: A novel transcription factor, T-bet,
directs Th1 lineage commitment. Cell. 100:655–669. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu J, Jankovic D, Oler AJ, Wei G, Sharma
S, Hu G, Guo L, Yagi R, Yamane H, Punkosdy G, et al: The
transcription factor T-bet is induced by multiple pathways and
prevents an endogenous Th2 cell program during Th1 cell responses.
Immunity. 37:660–673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Manetti R, Parronchi P, Giudizi MG,
Piccinni MP, Maggi E, Trinchieri G and Romagnani S: Natural killer
cell stimulatory factor (interleukin 12 (IL-12)) induces T helper
type 1 (Th1)-specific immune responses and inhibits the development
of IL-4-producing Th cells. J Exp Med. 177:1199–1204. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mattes J, Yang M, Mahalingam S, Kuehr J,
Webb DC, Simson L, Hogan SP, Koskinen A, McKenzie AN, Dent LA, et
al: Intrinsic defect in T cell production of interleukin (IL)-13 in
the absence of both IL-5 and eotaxin precludes the development of
eosinophilia and airways hyperreactivity in experimental asthma. J
Exp Med. 195:1433–1444. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mosmann TR and Coffman RL: TH1 and TH2
cells: Different patterns of lymphokine secretion lead to different
functional properties. Annu Rev Immunol. 7:145–173. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Barnes PJ: Immunology of asthma and
chronic obstructive pulmonary disease. Nat Immunol. 8:183–192.
2008. View
Article : Google Scholar
|
|
12
|
Yagi R, Zhu J and Paul WE: An updated view
on transcription factor GATA3-mediated regulation of Th1 and Th2
cell differentiation. Int Immunol. 23:415–420. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rincon M and Irvin CG: Role of IL-6 in
asthma and other inflammatory pulmonary diseases. Int J Biol Sci.
8:1281–1290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hershey GK: IL-13 receptors and signaling
pathways: An evolving web. J Allergy Clin Immunol. 111:677–690;
quiz 691. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rankin JA, Picarella DE, Geba GP, Temann
UA, Prasad B, DiCosmo B, Tarallo A, Stripp B, Whitsett J and
Flavell RA: Phenotypic and physiologic characterization of
transgenic mice expressing interleukin 4 in the lung: Lymphocytic
and eosinophilic inflammation without airway hyperreactivity. Proc
Natl Acad Sci USA. 93:7821–7825. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wills-Karp M, Luyimbazi J, Xu X, Schofield
B, Neben TY, Karp CL and Donaldson DD: Interleukin-13: Central
mediator of allergic asthma. Science. 282:2258–2261. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu Z, Homer RJ, Wang Z, Chen Q, Geba GP,
Wang J, Zhang Y and Elias JA: Pulmonary expression of
interleukin-13 causes inflammation, mucus hypersecretion,
subepithelial fibrosis, physiologic abnormalities, and eotaxin
production. J Clin Invest. 103:779–788. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hamid Q, Shannon J and Martin J:
Physiologic basis of respiratory diseaseCytokines and Chemokines in
Aasthma: An Overview. Tulic MK, Fiset PO, Muller Z and Hamid Q: BC
Decker Inc.; Hamilton: pp. 453–467. 2005
|
|
19
|
Mishra A, Yao X and Levine SJ: From
bedside to bench to clinic trials: Identifying new treatments for
severe asthma. Dis Model Mech. 6:877–888. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wise J: Corticosteroids for asthma may
suppress growth in children in first year of treatment, researchers
say. BMJ. 349:g46232014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ciriaco M, Ventrice P, Russo G,
Scicchitano M, Mazzitello G, Scicchitano F and Russo E:
Corticosteroid-related central nervous system side effects. J
Pharmacol Pharmacother. 4 (Suppl 1):S94–S98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bang MA, Seo JH, Seo JW, Jo GH, Jung SK,
Yu R, Park DH and Park SJ: Bacillus subtilis KCTC 11782BP-produced
alginate oligosaccharide effectively suppresses asthma via T-helper
cell type 2-related cytokines. PLoS One. 10:e01175242015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Seo JW, Cho SC, Park SJ, Lee EJ, Lee JH,
Han SS, Pyo BS, Park DH and Kim BH: 1′-Acetoxychavicol acetate
isolated from Alpinia galanga ameliorates ovalbumin-induced
asthma in mice. PLoS One. 8:e564472013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seo JH, Bang MA, Kim G, Cho SS and Park
DH: Erythronium japonicum attenuates histopathological lung
abnormalities in a mouse model of ovalbumin-induced asthma. Int J
Mol Med. 37:1221–1228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Singh KB and Nongmaithem M: Antioxidant
and free radical scavenging potential of Allium hookeri
Thwaites roots extract studied using in vitro models council for
innovative research. J Adv Biol. 4:276–285. 2014.
|
|
26
|
Li R, Wang YF, Sun Q and Hu HB: Chemical
composition and antimicrobial activity of the essential oil from
Allium hookeri consumed in Xishuangbanna, southwest China.
Nat Prod Commun. 9:863–864. 2014.PubMed/NCBI
|
|
27
|
Bae GC and Bae DY: The anti-inflammatory
effects of ethanol extract of Allium hookeri cultivated in
South Korea. Korea J Herbol. 27:55–61. 2012. View Article : Google Scholar
|
|
28
|
Kim HY, Nam SY, Hong SW, Kim MJ, Jeong HJ
and Kim HM: Protective effects of rutin through regulation of
vascular endothelial growth factor in allergic rhinitis. Am J
Rhinol Allergy. 29:e87–e94. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ariza A, Fernandez TD, Doña I, Aranda A,
Blanca-Lopez N, Melendez L, Canto G, Blanca M, Torres MJ and
Mayorga C: Basophil activation after nonsteroidal anti-inflammatory
drugs stimulation in patients with immediate hypersensitivity
reactions to these drugs. Cytometry A. 85:400–407. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
National Asthma Education and Prevention
Program: National asthma education and prevention program. Expert
panel report: Guidelines for the diagnosis and management of asthma
update on selected topics-2002. J Allergy Clin Immunol. 110 (Suppl
5):S141–S219. 2002.PubMed/NCBI
|
|
31
|
Galli SJ, Tsai M and Piliponsky AM: The
development of allergic inflammation. Nature. 454:445–454. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu J, Message SD, Qiu Y, Mallia P,
Kebadze T, Contoli M, Ward CK, Barnathan ES, Mascelli MA, Kon OM,
et al: Airway inflammation and illness severity in response to
experimental rhinovirus infection in asthma. Chest. 145:1219–1229.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mantovan A, Garlanda C and Locati M:
Macrophage diversity and polarization in atherosclerosis: A
question of balance. Arter Thromb Vasc Biol. 29:1419–1423. 2009.
View Article : Google Scholar
|
|
34
|
Magnan A, van Pee D, Bongrand P and
Vervloet D: Alveolar macrophage interleukin (IL)-10 and IL-12
production in atopic asthma. Allergy. 53:1092–1095. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gosset P, Tillie-Leblond I, Oudin S,
Parmentier O, Wallaert B, Joseph M and Tonnel AB: Production of
chemokines and proinflammatory and antiinflammatory cytokines by
human alveolar macrophages activated by IgE receptors. J Allergy
Clin Immunol. 103:289–297. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bal V, McIndoe A, Denton G, Hudson D,
Lombardi G, Lamb J and Lechler R: Antigen presentation by
keratinocytes induces tolerance in human T cells. Eur J Immunol.
20:1893–1897. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Frasca L, Marelli-Berg F, Imami N,
Potolicchio I, Carmichael P, Lombardi G and Lechler R:
Interferon-gamma-treated renal tubular epithelial cells induce
allospecific tolerance. Kidney Int. 53:679–689. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gaspari AA, Jenkins MK and Katz SI: Class
II MHC-bearing keratinocytes induce antigen-specific
unresponsiveness in hapten-specific Th1 clones. J Immunol.
141:2216–2220. 1988.PubMed/NCBI
|
|
39
|
Jang JY, Lee MJ, You BR, Jin JS, Lee SH,
Yun YR and Kim HJ: Allium hookeri root extract exerts
anti-inflammatory effects by nuclear factor-kB down-regulation in
lipopolysaccharide-induced RAW264.7 cells. BMC Complement Altern
Med. 17:1262017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim JE, Seo JH, Bae MS, Bae CS, Yoo JC,
Bang MA, Cho SS and Park DH: Antimicrobial Constituents from
Allium hookeri root. Nat Prod Commun. 11:237–238.
2016.PubMed/NCBI
|
|
41
|
Kim JE, Park KM, Lee SY, Seo JH, Yoon IS,
Bae CS, Yoo JC, Bang MA, Cho SS and Park DH: Anti-inflammatory
effect of Allium hookeri on carrageenan-induced air pouch
mouse model. PLoS One. 12:e01903052017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu F, Wang F, Wen T, Sang W, He X, Li L
and Zeng N: Protective effect of cinnamic acid in
endotoxin-poisoned mice. Phytother Res. 31:1946–1953. 2017.
View Article : Google Scholar : PubMed/NCBI
|