Introduction
Stroke, which can be classified into ischemic stroke
and hemorrhagic stroke, is a disabling disease (1). Ischemic stroke is a condition caused
by the occlusion of a blood vessel supplying the brain and
represents >80% of all stroke cases worldwide (2–4).
Therefore, further investigation into the pathogenesis of ischemic
stroke, and the development of safe and effective intervention
strategies to reduce cerebral injury induced by cerebral
ischemia-reperfusion is required (5).
The brain is a sensitive organ to ischemia and
hypoxia, with the potential that cerebral ischemia can lead to
brain cell necrosis or apoptosis (6). Therefore, timely thrombolysis, rapid
and effective reconstruction of microvascular collateral
circulation, the recovery of blood reperfusion in the ischemic
region and penumbra are required in cases of cerebral ischemia
(7). However, reperfusion after
ischemia can also lead to ischemia-reperfusion injury (IRI)
(7). A number of inflammatory
factors exist in the focal ischemic area at the time of cerebral
ischemia-reperfusion. In addition, activation and infiltration of
inflammatory cells, as well as the synthesis and secretion of
adhesion molecules that initiate a cascade reaction, which enhance
and promote each other, converting cerebral ischemic injury into
inflammatory injury through an inflammatory signaling pathway
(8). Consequently, the
inflammatory response plays an important role in the mechanism of
cerebral IRI (9). A number of
pro-inflammatory cytokines, chemokines and white blood cell
adhesion molecules are upregulated after local cerebral ischemia;
the pathogenesis and pathological development of which rely on
cellular signal transduction pathways (8). Some reactions can only take place
after various extracellular stimulatory signals are transduced into
cells (8). Typically, the
Toll-like receptor (TLR) signaling pathway and the nuclear
factor-κB (NF-κB) signaling pathway are involved in the
transduction of cerebral ischemia-reperfusion inflammation
(7,10).
MicroRNAs (miRNAs/miRs) are important endogenous
non-coding small RNAs, which are highly conserved throughout
evolution (11). miRNAs are
involved in a number of biological processes, including
proliferation, differentiation, apoptosis, metabolism and
development (12). Previous
studies have discovered that specific miRNAs play important roles
in ischemic cerebrovascular diseases, including in the genesis and
development of stroke, and are also involved in regulating the
protection and repair mechanism after cerebral injury (13,14).
Liu et al (15) reported
that miRNA-451 protected neurons against IRI-induced cell death.
Leecharoenkiat et al (16)
showed that miRNA-451 levels may be associated with
β-thalassemia/hemoglobin-E. The present study aimed to investigate
the role of miRNA-451 on cerebral ischemia-reperfusion and its
possible mechanism.
Materials and methods
Mouse model of cerebral
ischemia-reperfusion
Male C57BL/6J mice weighing 20–21 g were purchased
from Animal Experiment Center of Chongqing Medical University
(Chongqing, China) and housed at 22–23°C, 55–60% humidity with a 12
h light/dark cycle and freely access to food and water. Cerebral
ischemia-reperfusion was induced by middle cerebral artery
occlusion (MCAo). Mice were anesthetized with 50 mg/kg
pentobarbital (i.p.) and randomly assigned into two groups: Control
and cerebral ischemia-reperfusion model group. In the cerebral
ischemia-reperfusion model group, a midline neck incision was cut
and the left common and external carotid arteries were isolated,
and ligated with microvascular clips. To induce MCAo in the mice,
8-0 nylon monofilaments with silicon resin (180–190 µm) were
introduced through a small incision into the common carotid artery
and advanced by 9 mm distal to the carotid bifurcation. After 1 h,
reperfusion was initiated by withdrawing the monofilament. After 1
day, all mice were sacrificed for further study. The animal
experiments were approved by the Ethics Committee of The First
Affiliated Hospital of Chongqing.
Hematoxylin and eosin staining
The hippocampus was washed with PBS and fixed using
4% w/v paraformaldehyde in PBS for 24 h at room temperature. Tissue
samples were dehydrated in a graded ethanol series, cleared in
xylene, embedded in paraffin. Tissue samples were cut into 5
µm-thick sections and stained with hematoxylin sassy for 10 min and
eosin for 30 sec at room temperature. Images were captured under
fluorescence microscopy (magnification, ×200; Leica Microsystems
GmbH).
ELISA kits. Serum samples or cell samples were
collected at 2000 g for 10 min at 4°C. These samples were used to
measure tumor necrosis factor-α (TNF-α, H052), interleukin (IL)-1β
(H002), IL-6 (H007) and IL-18 (H015) levels using ELISA kits
(Nanjing Jiancheng Biology Engineering Institute).
Cell culture
Neuro-2a cells (Shanghai Cell Bank of Chinese
Academy of Sciences) were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% heat-inactivated fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 mg/ml streptomycin at 37°C with 5%
CO2. miRNA-451 (5′-AAACCGUUACCAUUACUGAGUU-3′),
anti-miRNA-451 and control negative mimics were transfected into
cells (1×105 cells) at a final concentration of 50
nmol/l using Lipofectamine® 2000 Reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. To induce an in vitro model of cerebral
ischemia-reperfusion, 48 h after transfection, cells were treated
with 100 ng/ml lipopolysaccharide (LPS) for 6 h at 37°C.
At 24 h after transfection with anti-miRNA-451, TLR4
inhibitor TAK-242 (8 nM) was added to the cells for 24 h and they
were then treated with 100 ng/ml lipopolysaccharide (LPS) for 6 h
at 37°C.
At 24 h after transfection with anti-miRNA-451,
MyD88 inhibitor, ST2825 (5 µM) was added into cell for 24 h and
they were then treated with 100 ng/ml lipopolysaccharide (LPS) for
6 h at 37°C.
At 24 h after transfection with anti-miRNA-451 and
si-p65 (sc-422642, Santa Cruz Biotechnology, Inc.), cells were
treated with 100 ng/ml lipopolysaccharide (LPS) for 6 h at
37°C.
RNA isolation, reverse
transcription-quantitative (RT-q)PCR
miRNA was extracted from serum or cells samples
using the miRNeasy Mini kit (Qiagen, China Co., Ltd.). RT was
carried out using 50 ng miRNA using SuperScript III Reverse
Transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.) at 42°C
for 1 h and 82°C for 20 sec. RT-qPCR was carried out using SYBR
Green I (Invitrogen; Thermo Fisher Scientific, Inc.) and a
QuantStudio 6 Flex Realtime PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The sequence of the primers for RT-qPCR:
TLR4: 5′-CTTCCTCTCCCCCCGTAACC-3′ and 5′-GTAAGAAACCGAAGGAATCAAA-3′;
MyD88: 5′-GTTCTGTTGCAACAAATTGAT-3′ and
5′-CTTATCAATTTGTTGCAACGAAC-3′; p65: 5′-CTTCCAAGAAGAGCAGCGTG-3′ and
5′-GCCTGGTCCCGTGAAATACA-3′; hsa-miR-451:
5′-ACACTCCAGCTGGGAAACCGTTACCATTACT-3′ and
5′-CTGGTGTCGTGGAGTCGGCAA-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′. Data was analyzed using the
2−ΔΔCq method (17).
Reaction conditions were 95°C for 15 min, denaturation at 95°C for
30 sec, annealing at 60°C for 30 sec and extension at 72°C for 30
sec for a total of 40 cycles.
Microarray analysis
For microarray analysis, 500 ng mRNA was transcribed
using the Low Input Quick-amp labeling kit (Agilent Technologies,
Inc.) and labeled with Cy3 (Agilent Technologies, Inc). The cDNA
was hybridized onto Agilent SurePrint G3 Mouse GE Microarray Chips
(Agilent Technologies, Inc.) and quantified using Feature
Extraction 10.5.1.1 image analysis software (Agilent Technologies,
Inc.).
Cell viability assay
Transfected cells (1×103 cell/well) were
seeded in 96-well plates and incubated with 20 µl Cell Counting
Kit-8 (CCK-8, Beyotime Institute of Biotechnology) reagent at 37°C
for 2 h. The absorbance was measured at 450 nm using a microplate
reader (Bio-Rad Laboratories, Inc.).
Flow cytometry analysis of
apoptosis
Transfected cells (1×106 cells) were
washed with PBS and stained with 5 µl FITC-conjugated annexin-V and
5 µl propidium iodide (BD Biosciences) for 15 min at room
temperature. Apoptosis was measured using a FACS Canto instrument
(BD Biosciences).
Western blot analysis
Total protein was extracted using RIPA lysis buffer
(Beyotime Institute of Biotechnology) and the protein concentration
was quantified using a Bicinchoninic Acid Protein Assay kit
(Beyotime Institute of Biotechnology). For each sample, 50 µg
protein was resolved using SDS-PAGE on 10–12% gels, which was then
transferred onto PVDF membranes. Membranes were blocked with 5%
non-fat milk in TBS-Tween-20 (TBST) for 1 h at 37°C and incubated
overnight at 4°C with antibodies against TLR4 (sc-13593, 1:500;
Santa Cruz Biotechnology, Inc.), MyD88 (sc-74532, 1:500; Santa Cruz
Biotechnology, Inc.), NF-κB/p65 (sc-71677, 1:500; Santa Cruz
Biotechnology, Inc.) and GAPDH (sc-32233, 1:2,000; Santa Cruz
Biotechnology, Inc.). The membranes were washed with TBST and
incubated with a horseradish peroxidase-conjugated secondary
antibody for 1 h at room temperature (sc-2004, 1:2,000; Santa Cruz
Biotechnology, Inc.). Membranes were visualized using an EasyBlot
ECL kit (Sangon Biotech Co., Ltd.) and quantified using ImageJ 1.49
(National Institutes of Health).
Immunofluorescence
Transfected cells (1×106 cells) were
washed with PBS and fixed with 4% paraformaldehyde for 15 min at
room temperature Cells were permeabilized using 0.3% Triton X-100
for 15 min at room temperature and blocked with 5% BSA (Beyotime
Institute of Biotechnology) in PBS for 1 h at room temperature.
Cells were incubated with an antibody against TLR4 (sc-13593,
1:100; Santa Cruz Biotechnology, Inc.) at 4°C overnight. After
washing with PBS, cells were incubated with Alexa 594-conjugated
anti-mouse or anti-rabbit immunoglobulin G secondary antibodies
(sc-362282, 1:100; Santa Cruz Biotechnology, Inc.) for 1 h at room
temperature. After washing with PBS, cells were stained with DAPI
(1 mg/ml) for 30 min protected from light at room temperature.
Images were captured with a fluorescent microscope (×200, Leica
Microsystems GmbH).
Luciferase reporter assay
Network signal path was analyzed using http://www.targetscan.org/vert_71/. The 3′ UTR
region of TLR4 predicted to bind to anti-miR-451 was synthesized
into the pGL3 firefly luciferase reporter plasmid (Promega
Corporation). Cells (1×106 cells) were co-transfected
with 10 ng of anti-miRNA-451 and 10 ng of pGL3-TLR4 using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 48 h. Luciferase and Renilla signals were
evaluated using the Dual Luciferase Reporter Assay kit (Promega
Corporation).
Statistical analysis
All data are presented as the mean ± SEM using SPSS
21.0 (IBM Corp.), (n=3). Statistical analyses were performed using
ANOVA with Tukey's post hoc test and Student's t-test. P<0.05
was considered to indicate a statistically significant
difference.
Results
miRNA-451 expression in mice with
cerebral ischemia-reperfusion
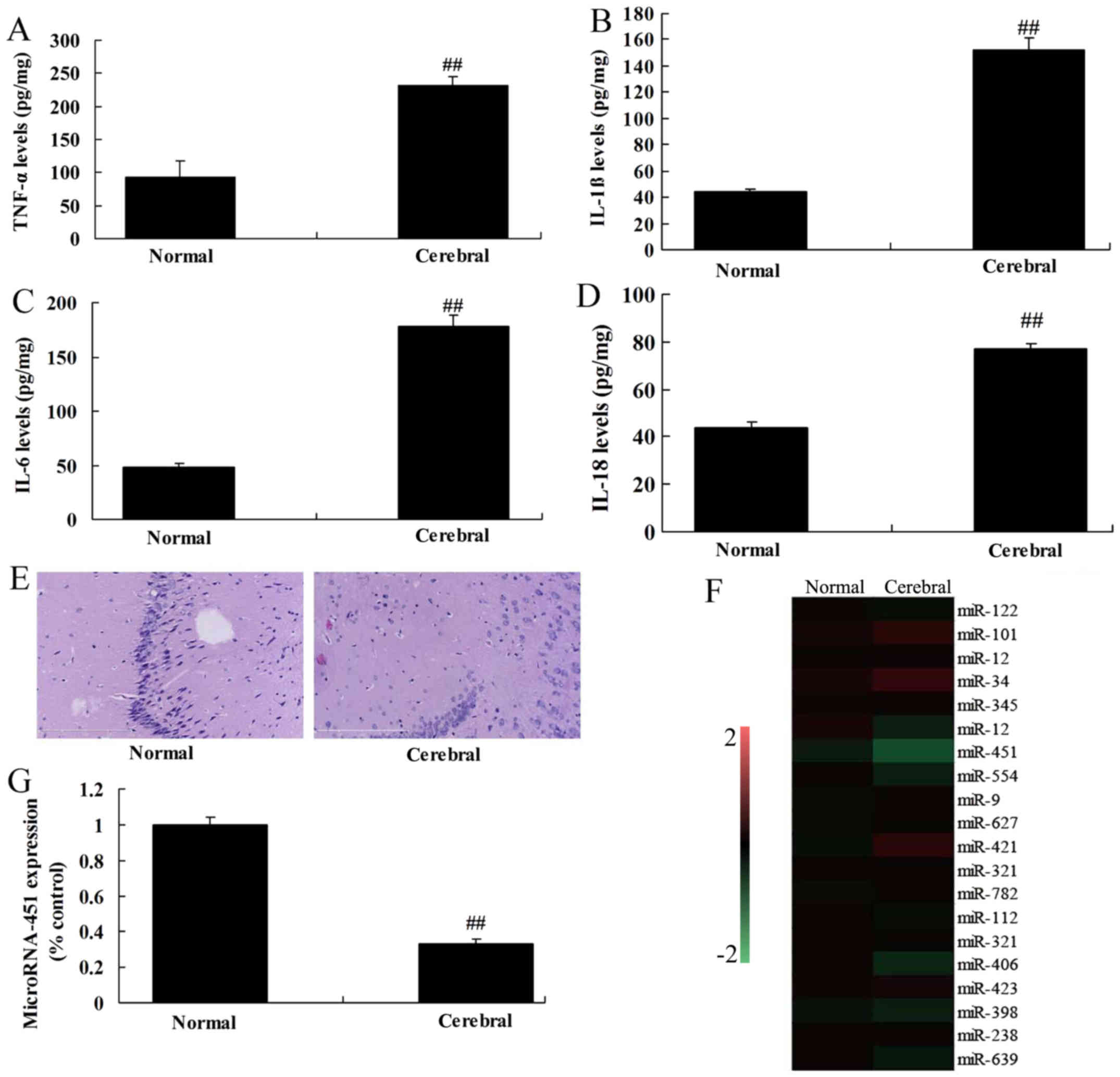
RT-qPCR was used to measure the level of miRNA-451
expression in rats with cerebral ischemia-reperfusion. The levels
of tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, IL-6 and
IL-18 were increased in rats with cerebral ischemia-reperfusion
compared with the control group (Fig.
1A-D). H&E staining showed that the number of nerve cells
was reduced in the cerebral ischemia-reperfusion group compared
with the control group (Fig. 1E).
As shown in Fig. 1F and G,
miRNA-451 expression was significantly lower in the cerebral
ischemia-reperfusion group compared with the control group,
suggesting that downregulation of miRNA-451 is a potential risk
factor for cerebral ischemia-reperfusion.
Regulation of miRNA-451 affects the
expression of MyD88 and NF-κB/p65 in cerebral ischemia-reperfusion
by TLR4
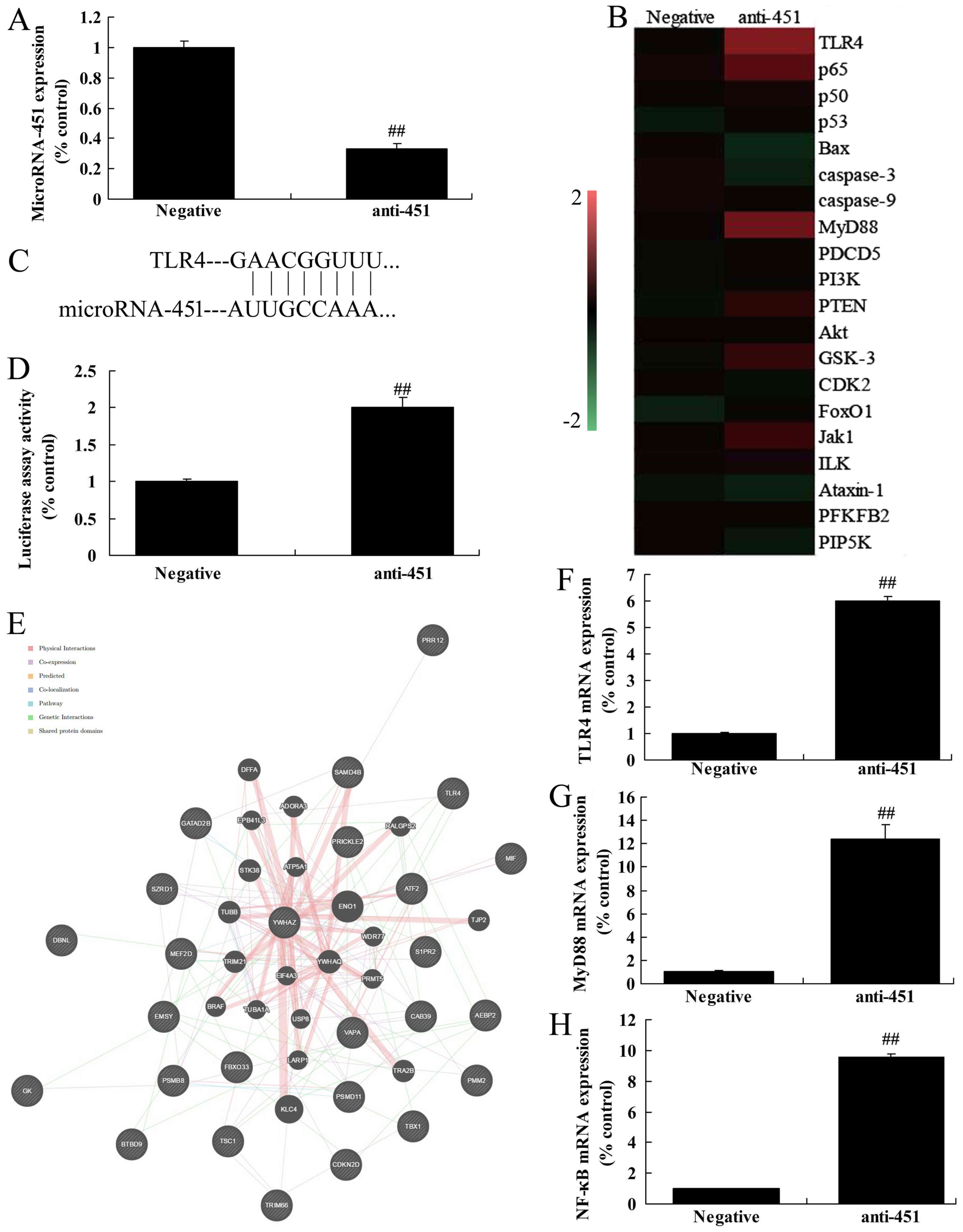
The mechanism of miRNA-451 on inflammation of
cerebral ischemia-reperfusion was evaluated by analyzing the
expression of MyD88 and NF-κB/p65. An anti-miRNA-451 mimic was used
to reduce the expression of miRNA-451 in vitro compared with
the negative control (Fig. 2A). A
heat map showed that the expression of TLR4, MyD88 and NF-κB/p65
were increased in vitro following the downregulation of
miRNA-451 compared with the negative group (Fig. 2B). Using putative miRNA-451 binding
sites in the 3′ untranslated region of TLR4 and a luciferase
reporter assay, it was found that the level of transcription was
higher following the downregulation of miRNA-451 (Fig. 2C and D). Network signal path
analysis using http://www.targetscan.org/vert_71/ revealed that TLR4
may be an important signaling pathway (Fig. 2E). RT-qPCR analysis showed that the
downregulation of miRNA-451 induced the expression of mRNA encoding
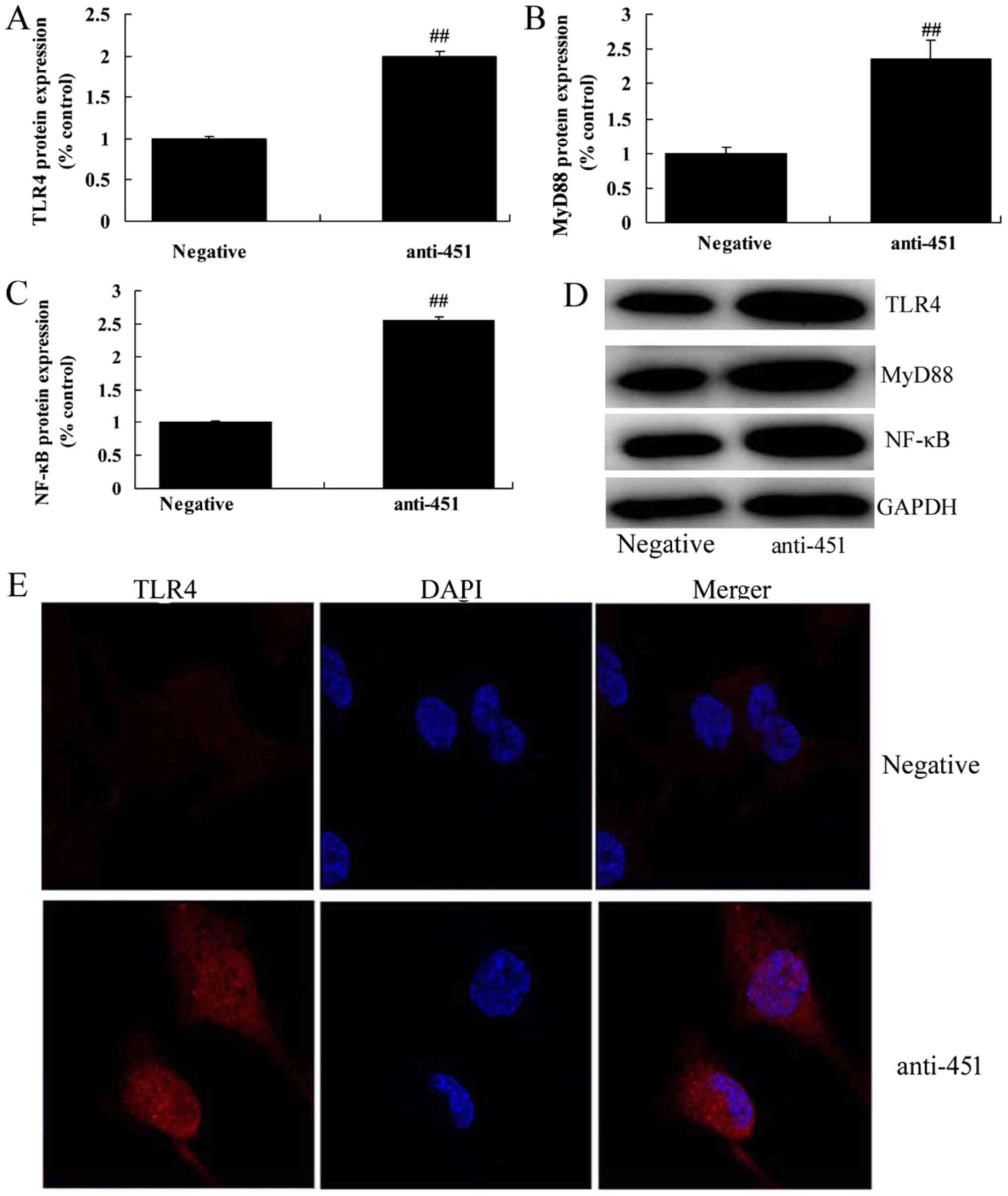
TLR4, MyD88 and NF-κB/p65 in vitro (Fig. 2F-H). Western blot analysis showed
that the downregulation of miRNA-451 also induced the expression of
TLR4, MyD88 and NF-κB/p65 at the protein level in vitro
(Fig. 3A-D). Immunofluorescence
also indicated that the downregulation of miRNA-451 induced the
protein expression of TLR4 in vitro (Fig. 3E). Transfection of cells with an
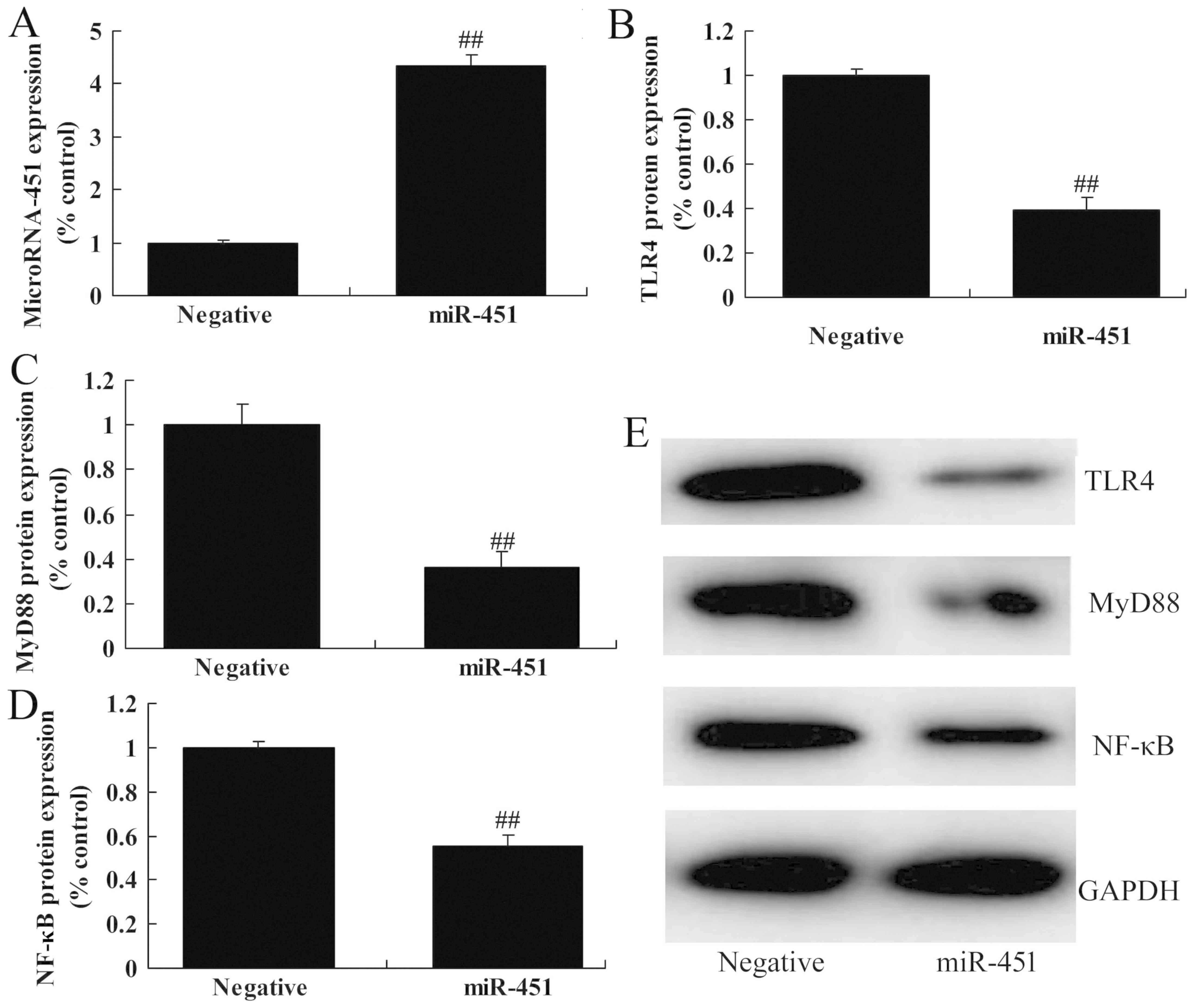
miRNA-451 mimic increased the expression of miRNA-451 in
vitro compared with the negative control (Fig. 4A). The upregulation of miRNA-451
suppressed the protein expression of TLR4, MyD88 and NF-κB/p65
in vitro (Fig. 4B-E). These
results indicated that miRNA-451 affected cerebral
ischemia-reperfusion-induced inflammation through the
TLR4/NF-κB/p65 signaling pathway.
Regulation of miRNA-451 affects
inflammation of cerebral ischemia-reperfusion
The effects of miRNA-451 on inflammation were
investigated further in the in vitro model of cerebral
ischemia-reperfusion. In comparison with the negative control,
overexpression of miRNA-451 reduced the levels of TNF-α, IL-1β,
IL-6 and IL-18. By contrast, the downregulation of miRNA-451
increased the levels of TNF-α, IL-1β, IL-6 and IL-18 (Fig. 5).
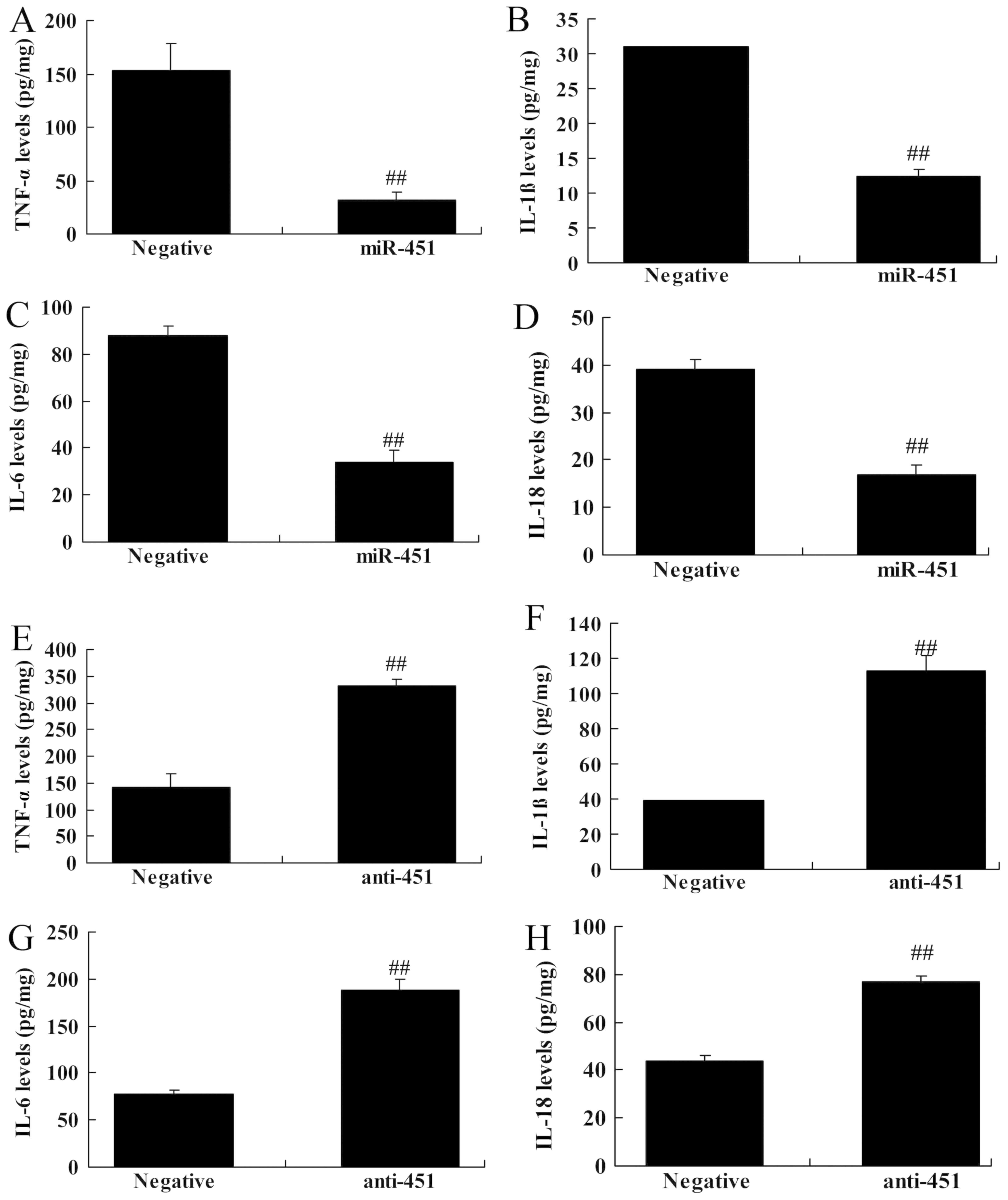 | Figure 5.Regulation of miRNA-451 affects
inflammation in cerebral ischemia-reperfusion. The expression
levels of (A) TNF-α, (B) IL-1β, (C) IL-6 and (D) IL-18 were
determined following the overexpression of miR-451. The expression
levels of (E) TNF-α, (F) IL-1β, (G) IL-6 and (H) IL-18 were
determined following the downregulation of miR-451.
##P<0.01 vs. negative. Negative, negative control;
miR-451, microRNA-451; anti-451, anti-microRNA-451; TLR4, Toll-like
receptor 4; MyD88, myeloid differentiation primary response protein
MyD88; NF-κB/p65, nuclear factor-κΒ; TNF-α, tumor necrosis
factor-α; IL, interleukin. |
Inhibition of TLR4 attenuates the
effect of miRNA-451 on the expression of TLR4/MyD88/NF-κB/p65 in
cerebral ischemia-reperfusion
Following the association between the level of
miRNA-451 and the TLR4/MyD88/NF-κB/p65 signaling pathway in
cerebral ischemia-reperfusion, the role of miRNA-451 in the
expression of TLR4, MyD88 and NF-κB/p65 in cerebral
ischemia-reperfusion was examined. The administration of the TLR4
inhibitor TAK-242 reduced the expression of TLR4, MyD88 and
NF-κB/p65 in cerebral ischemia-reperfusion following transfection
with miRNA-451, compared with cells transfected with anti-miRNA-451
but not exposed to TAK-242 (Fig.
6A-D). The levels of inflammatory factors, including TNF-α,
IL-1β, IL-6 and IL-18, were reduced in cells transfected with
anti-miRNA-451 and exposed to TAK-242 compared with cells
transfected with anti-miRNA-451 but not treated with TAK-242
(Fig. 6E-6H).
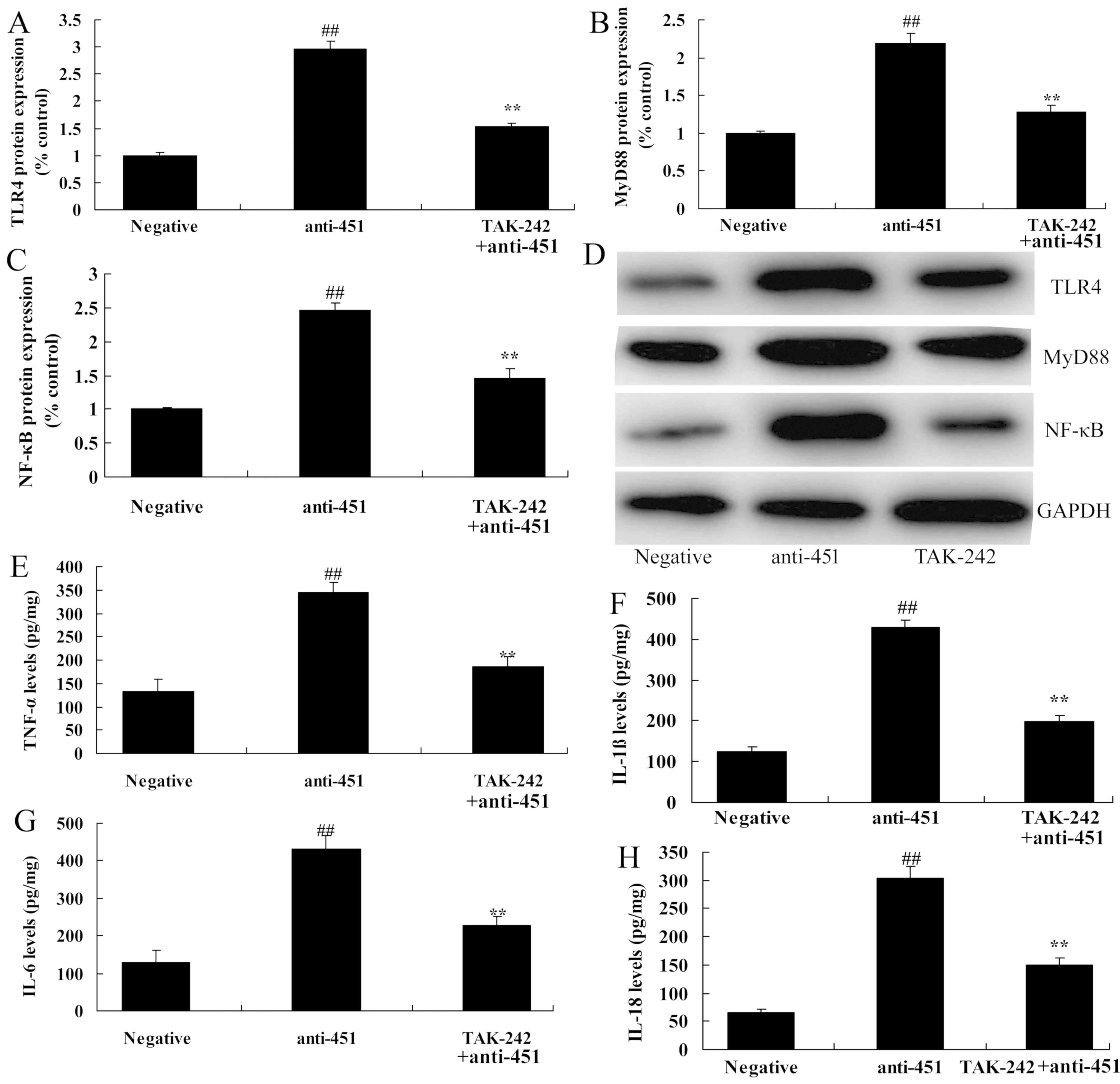 | Figure 6.Inhibition of TLR4 reduces the effect
of microRNA-451 on TLR4, MyD88, NF-κB/p65 protein expression in
cerebral ischemia-reperfusion. The expression level of (A) TLR4,
(B) MyD88 and (C) NF-κB/p65 were determined following transfection
with anti-451 and exposure to the TLR4 inhibitor TAK-242. (D)
Western blot analysis of TLR4, MyD88 and NF-κB/p65 protein
expression. The expression levels of (E) TNF-α, (F) IL-1β, (G) IL-6
and (H) IL-18 were determined following transfection with anti-451
and exposure to TAK-242. ##P<0.01 vs. negative;
**P<0.01 vs. anti-451. Negative, negative control; anti-451,
anti-microRNA-451; TLR4, Toll-like receptor 4; MyD88, myeloid
differentiation primary response protein MyD88; NF-κB/p65, nuclear
factor-κΒ; TNF-α, tumor necrosis factor-α; IL, interleukin. |
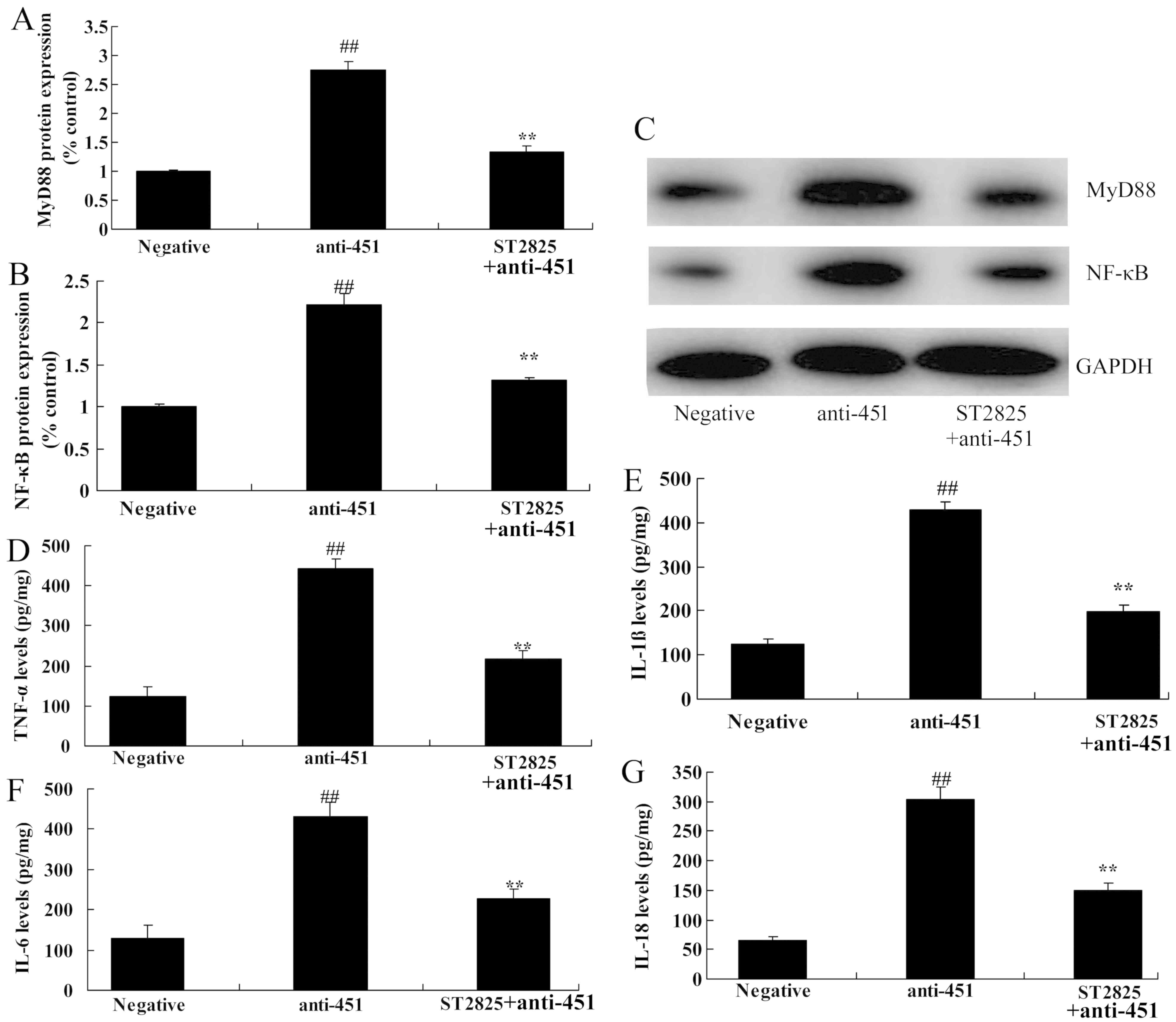
Inhibition of MyD88 attenuates the
effect of miRNA-451 on MyD88/NF-κB/p65 protein expression in
cerebral ischemia-reperfusion
To further explore the role of MyD88 and miRNA-451
in cerebral ischemia-reperfusion, the MyD88 inhibitor, ST2825, was
used. As shown in Fig. 7A-C,
ST2825 suppressed the effects of anti-miRNA-451 on the expression
of MyD88 and NF-κB/p65 in the in vitro model of cerebral
ischemia-reperfusion compared with cells transfected with
anti-miRNA-451 but not treated with ST2825. The inhibition of MyD88
also reduced the levels of TNF-α, IL-1β, IL-6 and IL-18 in cells
transfected with anti-miRNA-451 compared with cells transfected
with anti-miRNA-451 but not treated with ST2825 (Fig. 7D-G).
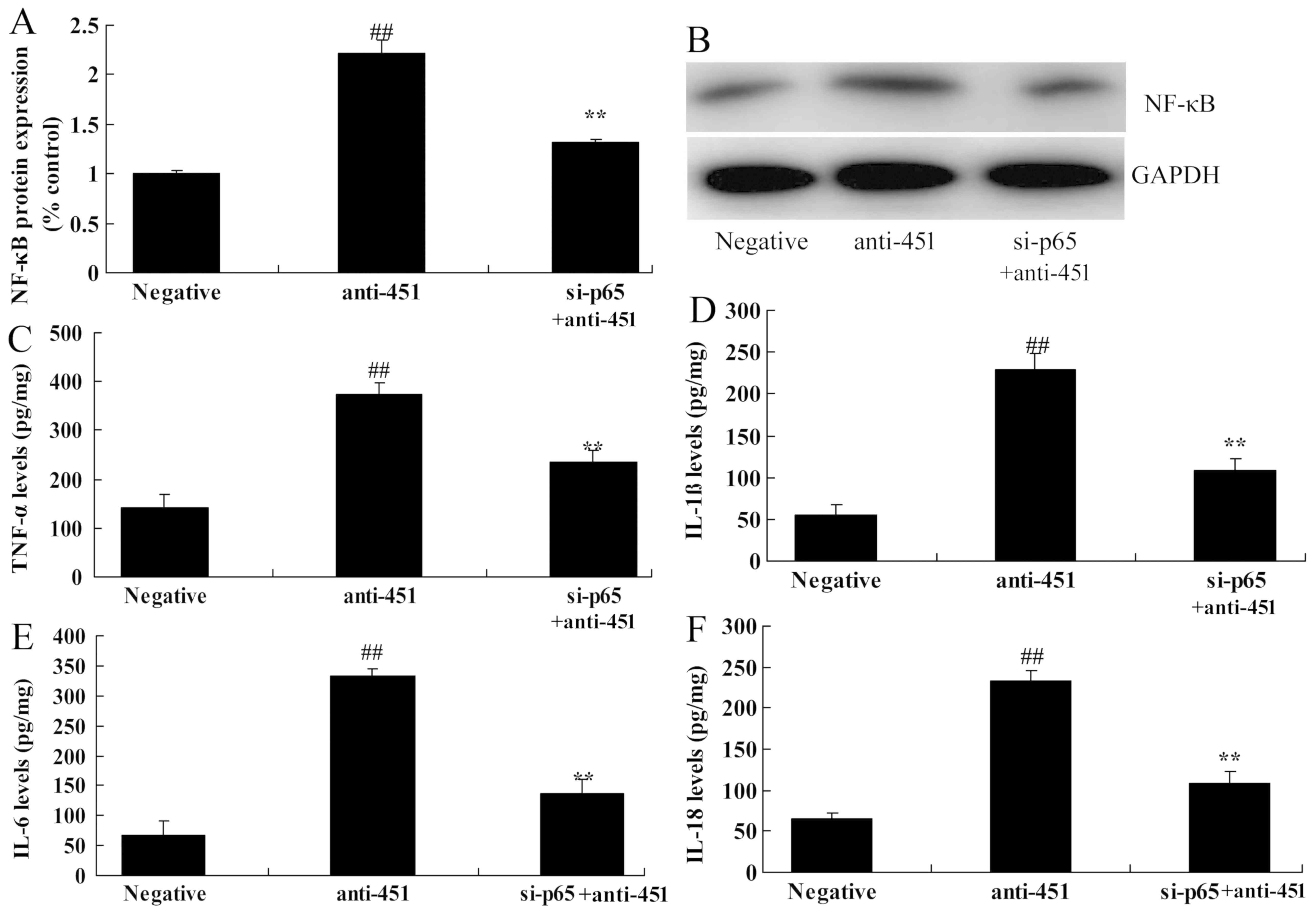
Inhibition of NF-κB/p65 attenuates the
effect of miRNA-451 on NF-κB/p65 protein expression in cerebral
ischemia-reperfusion
To further explore the role of NF-κB/p65 in and
miRNA-451 in cerebral ischemia-reperfusion, small interfering
(si)RNA targeting p65 (si-p65) was used. As shown in Fig. 8A and B, si-p65 suppressed the
effects of anti-miRNA-451 on the protein expression of NF-κB/p65 in
the in vitro model of cerebral ischemia-reperfusion compared
with cells transfected with anti-miRNA-451 but not with si-p65. The
reduction in the expression of NF-κB/p65 also reduced the levels of
TNF-α, IL-1β, IL-6 and IL-18 in cells transfected with
anti-miRNA-451 compared with cells transfected with anti-miRNA-451
but not with si-p65 (Fig.
8C-F).
Discussion
Cerebrovascular accident is also referred to as a
stroke, is characterized by high morbidity, high mortality, high
disability and high recurrence rates (18). The number of patients with
cerebrovascular disease, dominated by ischemic cerebrovascular
disease, has been increasing in China (19). Survival is associated with physical
disability, intellectual disability and a reduced quality of life,
which pose economic and social burdens on family and society
(19,20). The development of cerebral ischemia
is associated with a complicated mechanism and has diverse clinical
manifestations; therefore, its pathogenesis and prevention will
remain an important focus for medical research in the future.
miRNAs are a type of highly conserved non-coding small RNAs that
can become abnormally expressed in peripheral blood during the
pathogenesis of a disease (19,20).
miRNAs have been reported to play important roles in the genesis
and development of ischemic cerebrovascular disease (19,20).
In the present study, it was demonstrated that miRNA-451 expression
was downregulated in a mouse model of cerebral
ischemia-reperfusion. The downregulation of miRNA-451 increased
inflammation in an in vitro model of cerebral
ischemia-reperfusion. Wang et al (21) showed that miRNA-451 inhibited the
proliferation of synovial fibroblasts and the secretion of
inflammatory cytokines in rheumatoid arthritis.
Ischemic cerebrovascular disease results from local
cerebral blood flow obstruction, which can cause death or permanent
disability (10). Following a high
degree of cerebral ischemia, reperfusion cannot promote the
recovery of metabolism and other functions in the ischemic area,
but instead may aggravate the cerebral injury (22). The mechanism of cerebral IRI is
complex, including inflammatory responses, oxidative stress-induced
lipid peroxide reactions, metabolic disorders, toxic effects of
excitatory amino acids and apoptosis (23). In the present study, it was
demonstrated that the downregulation of miRNA-451 induced the
expression of TLR4, MyD88 and NF-κB/p65 in an in vitro model
of cerebral ischemia-reperfusion. Sun and Zhang (24) reported that miRNA-451 elevation
relieves inflammatory pain by targeting TLR4.
TLRs are members of the pathogen-related molecular
pattern receptor family, which can identify and bind to conserved
sequences of pathogenic micro-organisms (10). TLRs also recognize bacterial
endotoxin/LPS and induce inflammatory responses (10). It has been found that TLRs are also
involved in cerebral ischemia reperfusion inflammatory injury
(10). The brain is a sterile
organ and cerebral inflammatory injury is mainly induced by the TLR
pathway (22). The recognition of
DNA and protein by TLRs occurs in normal mice and mice with
cerebral IRI, which suggested that DNA and protein recognition by
TLR2 and TLR4 is higher in mice with cerebral IRI than normal mice
(22). The activity of NF-κB can
be regulated after the activation of TLRs, and NF-κB can upregulate
the expression of inflammatory factors, thus inducing inflammatory
injury (23). Furthermore, the
present study demonstrated that the downregulation of miRNA-451
induced the expression of TLR4, MyD88 and NF-κB/p65 in an in
vitro model of cerebral ischemia-reperfusion. Ren et al
(25) reported that ferulic acid
protected against cerebral ischemia-reperfusion-induced injury
through antioxidant and anti-apoptotic mechanisms. Liu et al
(26) reported that 3′-daidzein
sulfonate sodium inhibited neuronal apoptosis induced by cerebral
ischemia-reperfusion.
NF-κB is a member of the Rel protein family and is
an important signal transduction molecule involved in the
inflammatory response (27). NF-κB
is also an important transcription factor, which can be activated
by a number of factors, including inflammatory factors and
cytokines, and by calcium overload in the case of cerebral ischemia
(28). Activated NF-κB can induce
the expression of cytokines, adhesion molecules and inflammatory
enzymes, which gives rise to an inflammatory response, resulting in
brain edema and nerve cell injury (27). In the present study, the inhibition
of TLR4 and MyD88, or the knockdown of NF-κB, has been shown to
reduce the effect of miRNA-451 downregulation on inflammation in an
in vitro model of cerebral ischemia-reperfusion. Sun et
al (29) showed that miR-451
suppressed the expression of NF-κB-mediated pro-inflammatory
molecules in diabetic nephropathy. Liu et al (15) reported that miRNA-451 protected
neurons against IRI-induced cell death.
In conclusion, the present study found that
miRNA-451 expression was downregulated in a mouse model of cerebral
ischemia-reperfusion. The results from the present study suggested
that the downregulation of miRNA-451 reduces inflammation in an
in vitro model of cerebral ischemia-reperfusion through the
TLR4/MyD88/NF-κB signaling pathway. miRNA-451 may be a target for
the development of therapeutic agents for use in the treatment of
cerebral ischemia-reperfusion.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
XS designed the experiments; WL, MD, LC and LF
performed the experiments; XS analyzed the data and wrote the
manuscript.
Ethics approval and consent to
participate
Animal experiments were approved by the Ethics
Committee of The First Affiliated Hospital of Chongqing.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dippel DW, Majoie CB, Roos YB, van der
Lugt A, van Oostenbrugge RJ, van Zwam WH, Lingsma HF, Koudstaal PJ,
Treurniet KM, van den Berg LA, et al: Influence of device choice on
the effect of intra-arterial treatment for acute ischemic stroke in
MR CLEAN (multicenter randomized clinical trial of endovascular
treatment for acute ischemic stroke in the Netherlands). Stroke.
47:2574–2581. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim SK, Jang HM and Kim DY: The promoter
polymorphism of NFKB1 gene contributes to susceptibility of
ischemic stroke in Korean population. J Exerc Rehabil.
14:1096–1100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ceulemans AG, Zgavc T, Kooijman R,
Hachimi-Idrissi S, Sarre S and Michotte Y: The dual role of the
neuroinflammatory response after ischemic stroke: Modulatory
effects of hypothermia. J Neuroinflammation. 7:742010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Candelario-Jalil E: Injury and repair
mechanisms in ischemic stroke: Considerations for the development
of novel neurotherapeutics. Curr Opin Investig Drugs. 10:644–654.
2009.PubMed/NCBI
|
|
5
|
Kawakami S, Tahara Y, Noguchi T, Yagi N,
Kataoka Y, Asaumi Y, Nakanishi M, Goto Y, Yokoyama H, Nonogi H, et
al: Time to reperfusion in ST-segment elevation myocardial
infarction patients with vs. without pre-hospital mobile
telemedicine 12-lead electrocardiogram transmission. Circ J.
80:1624–1633. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao GC, Yuan YL, Chai FR and Ji FJ:
Effect of Melilotus officinalis extract on the apoptosis of brain
tissues by altering cerebral thrombosis and inflammatory mediators
in acute cerebral ischemia. Biomed Pharmacother. 89:1346–1352.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou L, Zhang J, Wang C and Sun Q:
Tanshinone inhibits neuronal cell apoptosis and inflammatory
response in cerebral infarction rat model. Int J Immunopathol
Pharmacol. 30:123–129. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bonestroo HJ, Nijboer CH, van Velthoven
CT, Kavelaars A, Hack CE, van Bel F and Heijnen CJ: Cerebral and
hepatic inflammatory response after neonatal hypoxia-ischemia in
newborn rats. Dev Neurosci. 35:197–211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Naderi Y, Sabetkasaei M, Parvardeh S and
Zanjani TM: Neuroprotective effect of minocycline on cognitive
impairments induced by transient cerebral ischemia/reperfusion
through its anti-inflammatory and anti-oxidant properties in male
rat. Brain Res Bull. 131:207–213. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li X, Su L, Zhang X, Zhang C, Wang L, Li
Y, Zhang Y, He T, Zhu X and Cui L: Ulinastatin downregulates TLR4
and NF-kB expression and protects mouse brains against
ischemia/reperfusion injury. Neurol Res. 39:367–373. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou J and Zhang J: Identification of
miRNA-21 and miRNA-24 in plasma as potential early stage markers of
acute cerebral infarction. Mol Med Rep. 10:971–976. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Li F, Zhao S, Luo Y, Kang J, Zhao
H, Yan F, Li S and Ji X: MicroRNA-124-mediated regulation of
inhibitory member of apoptosis-stimulating protein of p53 family in
experimental stroke. Stroke. 44:1973–1980. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen S, Wang M, Yang H, Mao L, He Q, Jin
H, Ye ZM, Luo XY, Xia YP and Hu B: LncRNA TUG1 sponges microRNA-9
to promote neurons apoptosis by up-regulated Bcl2l11 under
ischemia. Biochem Biophys Res Commun. 485:167–173. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Truettner JS, Alonso OF, Bramlett HM and
Dietrich WD: Therapeutic hypothermia alters microRNA responses to
traumatic brain injury in rats. J Cereb Blood Flow Metab.
31:1897–1907. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Q, Hu Y, Zhang M, Yan Y, Yu H and Ge
L: microRNA-451 protects neurons against ischemia/reperfusion
injury-induced cell death by targeting CELF2. Neuropsychiatr Dis
Treat. 14:2773–2782. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leecharoenkiat K, Tanaka Y, Harada Y,
Chaichompoo P, Sarakul O, Abe Y, Smith DR, Fucharoen S, Svasti S
and Umemura T: Plasma microRNA-451 as a novel hemolytic marker for
β0-thalassemia/HbE disease. Mol Med Rep. 15:2495–2502. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hill MD, Demchuk AM, Goyal M, Jovin TG,
Foster LD, Tomsick TA, von Kummer R, Yeatts SD, Palesch YY and
Broderick JP; IMS3 Investigators, : Alberta Stroke Program early
computed tomography score to select patients for endovascular
treatment: Interventional Management of Stroke (IMS)-III Trial.
Stroke. 45:444–449. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu Z, Fu Y, Tian D, Sun N, Han W, Chang
G, Dong Y, Xu X, Liu Q, Huang D and Shi FD: Combination of the
immune modulator fingolimod with alteplase in acute ischemic
stroke: A pilot trial. Circulation. 132:1104–1112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shan J, Sun L, Wang D and Li X: Comparison
of the neuroprotective effects and recovery profiles of isoflurane,
sevoflurane and desflurane as neurosurgical pre-conditioning on
ischemia/reperfusion cerebral injury. Int J Clin Exp Pathol.
8:2001–2009. 2015.PubMed/NCBI
|
|
21
|
Wang ZC, Lu H, Zhou Q, Yu SM, Mao YL,
Zhang HJ, Zhang PC and Yan WJ: MiR-451 inhibits synovial
fibroblasts proliferation and inflammatory cytokines secretion in
rheumatoid arthritis through mediating p38MAPK signaling pathway.
Int J Clin Exp Pathol. 8:14562–14567. 2015.PubMed/NCBI
|
|
22
|
Zhang P, Guo ZF, Xu YM, Li YS and Song JG:
N-Butylphthalide (NBP) ameliorated cerebral ischemia
reperfusion-induced brain injury via HGF-regulated TLR4/NF-κB
signaling pathway. Biomed Pharmacother. 83:658–666. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou H, Yang WS, Li Y, Ren T, Peng L, Guo
H, Liu JF, Zhou Y, Zhao Y, Yang LC and Jin X: Oleoylethanolamide
attenuates apoptosis by inhibiting the TLR4/NF-κB and ERK1/2
signaling pathways in mice with acute ischemic stroke. Naunyn
Schmiedebergs Arch Pharmacol. 390:77–84. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun X and Zhang H: miR-451 elevation
relieves inflammatory pain by suppressing microglial
activation-evoked inflammatory response via targeting TLR4. Cell
Tissue Res. 374:487–495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ren Z, Zhang R, Li Y, Li Y, Yang Z and
Yang H: Ferulic acid exerts neuroprotective effects against
cerebral ischemia/reperfusion-induced injury via antioxidant and
anti-apoptotic mechanisms in vitro and in vivo. Int J
Mol Med. 40:1444–1456. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu R, Zhong X, Zeng J, Huang Z, Li X,
Xiao H, Chen Q and Li D: 3′-Daidzein sulfonate sodium inhibits
neuronal apoptosis induced by cerebral ischemia-reperfusion. Int J
Mol Med. 39:1021–1028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kacimi R, Giffard RG and Yenari MA:
Endotoxin-activated microglia injure brain derived endothelial
cells via NF-κB, JAK-STAT and JNK stress kinase pathways. J Inflamm
(Lond). 8:72011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feola J, Barton A, Akbar A, Keillor J and
Johnson GVW: Transglutaminase 2 modulation of NF-κB signaling in
astrocytes is independent of its ability to mediate astrocytic
viability in ischemic injury. Brain Res. 1668:1–11. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun Y, Peng R, Peng H, Liu H, Wen L, Wu T,
Yi H, Li A and Zhang Z: miR-451 suppresses the NF-kappaB-mediated
proinflammatory molecules expression through inhibiting LMP7 in
diabetic nephropathy. Mol Cell Endocrinol. 433:75–86. 2016.
View Article : Google Scholar : PubMed/NCBI
|