Introduction
Malignant melanoma arises from melanocytes, which
are responsible for pigment production (1–3). The
incidence of melanoma has increased at an alarming rate and
patients with advanced malignancies exhibit poor prognoses, with an
average survival time of 3–11 months (4–8).
Melanoma can be removed via surgical resection in patients with
early diagnosis; however, melanoma has high metastatic potential
and treatment options for metastatic melanoma are limited (9–12).
Therefore, novel targets against melanoma are urgently required for
the identification of effective therapies.
Remodeling and spacing factor 1 (Rsf-1), also known
as hepatitis B X-antigen associated protein, is a subunit of RSF
(13,14). Rsf-1 protein is located in the
nucleus and binds to human sucrose nonfermenting protein 2 homolog
(hSNF2H), forming a chromatin remodeling complex (15). The Rsf-1/hSNF2H complex regulates
adenosine 5′-triphosphate-dependent chromatin remodeling and alters
the chromatin structure of nucleosomes (15,16),
which are required for biological processes, including activation
or repression of transcription, DNA replication and cell cycle
progression (17,18).
Rsf-1 overexpression has been reported in a number
of solid tumors, including breast cancer, ovarian cancer and oral
squamous cell carcinoma (19–25);
increased Rsf-1 expression was associated with poor prognosis in
bladder cancer (15) and
nasopharyngeal cancer (26).
Additionally, ectopic expression of Rsf-1 promoted cell and tumor
growth in a mouse xenograft model (27). Furthermore, Rsf-1 was associated
with paclitaxel resistance in ovarian cancer (28); however, there are no reports
concerning the expression profile of Rsf-1 in malignant melanoma.
The aim of the present study was to determine the status of Rsf-1
in malignant melanoma tissues, and the effects of Rsf-1 on the
biological behavior of melanoma cell lines.
Materials and methods
Patients and specimens
The present study was approved by the Ethics
Committee of China Medical University (Shenyang, China). Informed
consent was obtained from all patients. Melanoma and adjacent
normal specimens were obtained from 50 patients diagnosed with
malignant melanoma who underwent resection at The First Affiliated
Hospital of China Medical University (Shenyang, China) between
November 2009 and March 2012. Patients did not receive chemotherapy
or radiation therapy prior to surgical resection. Histological
classification was performed according to the American Joint
Committee on Cancer (29). There
were 20 female and 30 male patients, with an age range of 25–82
years (mean, 53.2±8.67 years).
Immunohistochemical staining
Tumor samples were obtained from The First
Affiliated Hospital of China Medical University. The samples were
fixed in 37% formaldehyde at room temperature for 18 h and embedded
in paraffin. Immunostaining was performed using the Elivision Plus
method (Fuzhou Maixin Biotech. Co., Ltd., Fuzhou, China). Sections
of 4-µm thickness were deparaffinized in xylene and rehydrated with
a graded alcohol series (100, 95, 80 and 70%). Sections were
permeabilized with Triton X-100 and then boiled in citrate buffer.
Sections were blocked with goat serum (Fuzhou Maixin Biotech Co.,
Ltd.) at room temperature for 20 min. Hydrogen peroxide (0.3%) was
used to block peroxidase activity. Sections were incubated with
rabbit anti-Rsf-1 polyclonal antibody (1:1,000; HPA046129,
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 4°C overnight.
Sections were then incubated for 2 h at 37°C with a biotinylated
anti-rabbit horseradish peroxidase (HRP) polymer (KIT-9902, Fuzhou
Maixin Biotech. Co., Ltd.). Sections were developed with
3,3′-diaminobenzidine plus from Fuzhou Maixin Biotech. Co., Ltd.
Sections were counterstained with hematoxylin at room temperature
for 2 min.
All tumor slides were analyzed by two independent
investigators randomly under a light microscope (magnification,
×400; BX53; Olympus Corporation, Tokyo, Japan). Immunostaining of
Rsf-1 was scored using a semi-quantitative scale by evaluating the
intensity and percentage of tumor cells. Nuclear immunostaining was
considered positive. The intensity of Rsf-1 staining was scored as
0 (no signal), 1 (moderate) or 2 (strong). Percentage scores were
assigned as 1 (1–25%), 2 (26–50%), 3 (51–75%) or 4 (76–100%)
(30). The scores of each tumor
sample were multiplied to provide a final score of 0–8; tumor
samples that scored 4–8 were considered to demonstrate Rsf-1
overexpression.
Cell culture and reagents
M14 cells with low Rsf-1 expression, and MV3 and
A375 cells with high Rsf-1 expression were purchased from the
American Type Culture Collection (Manassas, VA, USA). M14 and A375
cells were cultivated in Dulbecco's Modified Eagle's medium
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Invitrogen; Thermo
Fisher Scientific, Inc.). MV3 cells were cultured in RPMI-1640
medium (Invitrogen; Thermo Fisher Scientific, Inc.) supplemented
with 10% FBS. Cells were cultured under conditions of 37°C and 5%
CO2, and seeded at a density of 1×106
cells/ml. Then, cells were treated with cisplatin (final
concentration, 5 µM) following attachment of cells to plates at
37°C for 24 h. Additionally, M14 cells were treated with NF-κB
inhibitor (Bay11-7082; cat. no. S2913, Selleck Chemicals, Houston,
TX, USA) at a concentration of 10 µM for 12 h at 37°C.
Small interfering RNA (siRNA) and
plasmid transfection
Oligonucleotide pools of siRNA targeting Rsf-1 and
non-targeting siRNA (control siRNA) were purchased from GE
Healthcare Dharmacon, Inc. (Lafayette, CO, USA), and MV3 and A375
cells were transfected with 50 nM siRNA using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols. The
targeting sequences were as follows: Rsf-1 siRNA,
5′-GGAAAGACAUCUCUACUAU-3′; and control siRNA,
5′-GCGCGATAGCGCGAATATA-3′. pCMV6-Rsf-1 and control empty plasmids
were purchased from OriGene Technologies, Inc. (Rockville, MD,
USA), and M14 cells were transfected with 1 µg plasmid using
Lipofectamine 3000 according to the manufacturer's protocols.
Subsequent experiments were performed 48–72 h following
transfection.
Western blotting
Total protein from cells was extracted using Pierce™
Universal Nuclease for Cell Lysis (Pierce; Thermo Fisher
Scientific, Inc.) and quantified by the Bradford method. A total of
40 µg protein was separated by 8–12% SDS-PAGE. Samples were
transferred to polyvinylidene difluoride membranes (EMD Millipore),
blocked at room temperature for 1 h in 3% bovine serum albumin
(BioSharp Co., Hefei, China), and incubated overnight at 4°C with
antibodies against: Rsf-1 (1:2,000; cat. no. HPA046129,
Sigma-Aldrich; Merck KGaA), cyclin E (1:700; cat. no. 4129, Cell
Signaling Technology, Inc., Danvers, MA, USA), matrix
metalloproteinase-2 (MMP2; 1:1,000; cat. no. 4022, Cell Signaling
Technology, Inc.), IκB (1:1,000; cat. no. 9242, Cell Signaling
Technology, Inc.), phosphorylated (p)-IκB (1:1,000; cat. no. 9246,
Cell Signaling Technology, Inc.), nuclear factor
κ-light-chain-enhancer of activated B cells (NF-κB; 1:1,000; cat.
no. 4764, Cell Signaling Technology, Inc.), B-cell lymphoma (Bcl-2;
1:1,000; cat. no. 15071, Cell Signaling Technology, Inc.),
Bcl-2-associated X protein (Bax; 1:1,000; cat. no. 2774, Cell
Signaling Technology, Inc.), cellular inhibitor of apoptosis
protein 1 (cIAP1; 1:1,000; cat. no. 7065, Cell Signaling
Technology, Inc.), cIAP2 (1:1,000; cat. no. 3130, Cell Signaling
Technology, Inc.) and β-actin (1:2,000; cat. no. 4970, Cell
Signaling Technology, Inc.). Following incubation with
HRP-conjugated anti-mouse/rabbit IgG (1:1,000; cat nos. 7076/7074,
Cell Signaling Technology, Inc.) at 37°C for 2 h, proteins were
visualized using an enhanced chemiluminescence kit (Pierce; Thermo
Fisher Scientific, Inc.) and detected using a DNR Bio-Imaging
System (DNR Bio-Imaging Systems, Ltd., Neve Yamin, Israel).
Relative protein levels were quantified using ImageJ 1.8.0 software
(National Institutes of Health, Bethesda, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from MV3, M14 and A375 cells
using TRIzol® reagent (Thermo Fisher Scientific, Inc.).
Total RNA (500 ng) was then reverse-transcribed using PrimeScript
RT Master Mix (10X; Takara Biotechnology Co., Ltd., Dalian, China)
at 85°C for 2 min and 37°C for 30 min. qPCR was conducted using the
Reverse Transcription System kit (Promega Corporation, Madison, WI,
USA) according to the manufacturer's protocols. An ABI 7500
Real-Time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) was used for gene amplification, under the conditions of:
95°C for 2 min, and 40 cycles of 95°C for 2 sec and
annealing/extension at 60°C for 30 sec. A dissociation step was
performed at 60–95°C for 6 sec to generate a melting curve. β-actin
was used as the reference gene, and relative levels of gene
expression were represented as: ΔCq=Cq gene-Cq reference. The fold
change in gene expression was calculated using the
2−ΔΔCq method (31).
The experiment was performed in triplicate. The primers were as
follows: Rsf-1, forward 5′-GATACTATGCGTCTCCAGCCAA-3′, reverse,
5′-CAACTCGTTTCGATTTCTGACAA-3′; and β-actin, forward
5′-CCAACCGCGAGAAGATGACC-3′ and reverse,
5′-GATAGCACAGCCTGGATAGCAAC-3′.
MTT assay
A total of 5,000 cells were plated in 96-well plates
and cultured overnight, followed by the addition of 20 µl of 5
mg/ml MTT solution to each well; cells were incubated for 4 h at
37°C. The supernatant was removed from each well, and dimethyl
sulfoxide (150 µl) was added to dissolve the formazan crystals. The
absorbance was detected at 490 nm using a microplate reader
(Infinite F50; Tecan Group, Ltd., Mannedorf, Switzerland).
Colony formation assay
For colony formation, cells were seeded into three
6-cm cell culture dishes (~800 cells/dish) 48 h following
transfection. Cells were incubated for 14 days at 37°C. Plates were
washed with PBS and then stained with Giemsa at room temperature
for 10 min. The number of colonies with >50 cells was manually
counted under a light microscope (magnification, ×200; BX53).
Transwell invasion assay
A Transwell invasion assay was performed using a
24-well Transwell chamber with a pore size of 8 µm (Costar; Corning
Inc., Corning, NY, USA), and the inserts were coated with 20 µl
Matrigel (1:3; BD Biosciences, San Jose, CA, USA). After 48 h
following transfection, cells were trypsinized (0.25% trypsin) at
37°C for 30 sec and then transferred to the upper Matrigel-coated
chamber in 100 µl serum-free medium (1×105 cells/ml).
Medium (DMEM for M14 and A375 cells, RPMI-1640 for MV3 cells)
supplemented with 10% FBS was added to the lower chamber as the
chemoattractant. Cells were incubated for 18 h at 37°C.
Non-invading cells on the upper membrane surface were then removed
with a cotton tip, and the cells that passed via the filter were
fixed in 4% paraformaldehyde at room temperature for 20 min. Cells
were stained with hematoxylin at room temperature for 5 min. Cells
were observed under a light microscope (magnification, ×200; BX53).
The experiments were performed in triplicate.
Flow cytometry for cell cycle and
apoptosis analyses
Cells in 6-well plates were collected using tryptase
48 h following transfection. Cells were washed twice with PBS,
followed by resuspension in 250 µl binding buffer (BD Pharmingen;
BD Biosciences). Cells were fixed in 1% paraformaldehyde at 4°C
overnight and then stained with 5 mg/ml propidium iodide (PI) alone
or together with Annexin V/fluorescein isothiocyanate (BD
Pharmingen; BD Biosciences) at room temperature for 15 min for cell
cycle or apoptosis analysis, respectively. Incubation was performed
in the dark for 15 min. Flow cytometry was performed using flow
cytometer and analyzed using NovoExpress 1.2.5 software (ACEA
Biosciences, Inc.; Agilent Technologies, Inc., Santa Clara, CA,
USA). The apoptotic rate was calculated by adding the percentage of
early apoptotic (Annexin V-positive, PI-negative) and late
apoptotic cells (Annexin V-positive, PI-positive).
Detection of the mitochondrial
membrane potential (MMP)
The MMP was detected via the JC-1 staining method.
Briefly, cells (300 cells/µl) were harvested, washed with PBS and
incubated with 5 µM JC-1 (Cell Signaling Technology, Inc.) at 37°C
for 30 min in an incubator. Cells were then washed and analyzed
using a flow cytometer. Data were analyzed using NovoExpress 1.2.5
software.
Statistical analysis
SPSS version 16 for Windows (SPSS, Inc., Chicago,
IL, USA) was used for all statistical analyses. A χ2
test was used to examine potential associations between Rsf-1
expression and the clinicopathological features of patients with
melanoma. A Student's t-test was used to compare differences
between the control and treatment groups. Data were presented as
the mean ± standard deviation of at least three experiments.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of Rsf-1 in human malignant
melanoma
Rsf-1 expression in 50 cases of malignant melanoma
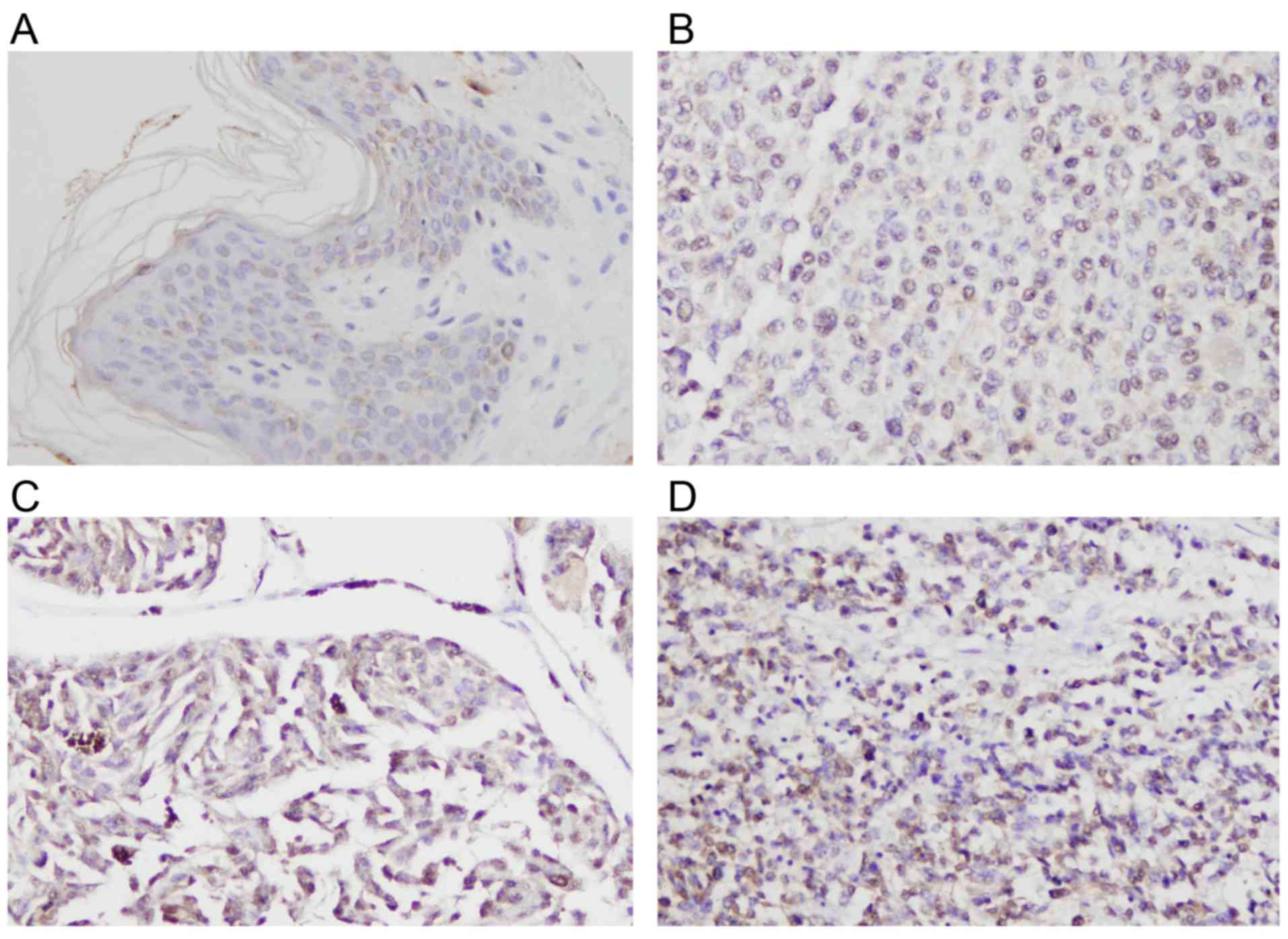
was determined by immunohistochemistry (Fig. 1). Normal skin tissue exhibited weak
or negative staining (Fig. 1A). In
total, 19/50 (38%) cases of skin melanoma demonstrated high Rsf-1
immunoreactivity (Rsf-1 overexpression, or an immunostaining score
of ≥4), which was localized to the nuclear compartment of tumor
cells (Fig. 1B-D). The association
between Rsf-1 expression and the clinicopathological
characteristics of patients with melanoma was analyzed (Table I). The frequency of Rsf-1
overexpression was increased in melanomas of advanced tumor, node
and metastasis (TNM) stages (III+IV vs. II, P=0.0494). The results
revealed that no significant association was observed between Rsf-1
expression and patient age (P=0.5655) and gender (P=0.122), or T
stage (P=0.8842).
 | Table I.Distribution of Rsf-1 status in
melanoma according to the clinicopathological characteristics of
patients. |
Table I.
Distribution of Rsf-1 status in
melanoma according to the clinicopathological characteristics of
patients.
| Clinicopathological
characteristics | Number of
patients | Rsf-1 low
expression | Rsf-1 high
expression | χ2 | P-value |
|---|
| Age (years) |
|
|
| 0.3302 | 0.5655 |
|
<60 | 34 | 22 | 12 |
|
|
|
≥60 | 16 | 9 | 7 |
|
|
| Sex |
|
|
| 2.3911 | 0.1220 |
|
Female | 20 | 15 | 5 |
|
|
|
Male | 30 | 16 | 14 |
|
|
| TNM stage |
|
|
| 3.8606 | 0.0494 |
| II | 43 | 29 | 14 |
|
|
|
III+IV | 7 | 2 | 5 |
|
|
| T stage |
|
|
| 0.0212 | 0.8842 |
|
T1-3 | 10 | 6 | 4 |
|
|
| T4 | 40 | 25 | 15 |
|
|
Rsf-1 promotes malignant melanoma cell
viability and invasion
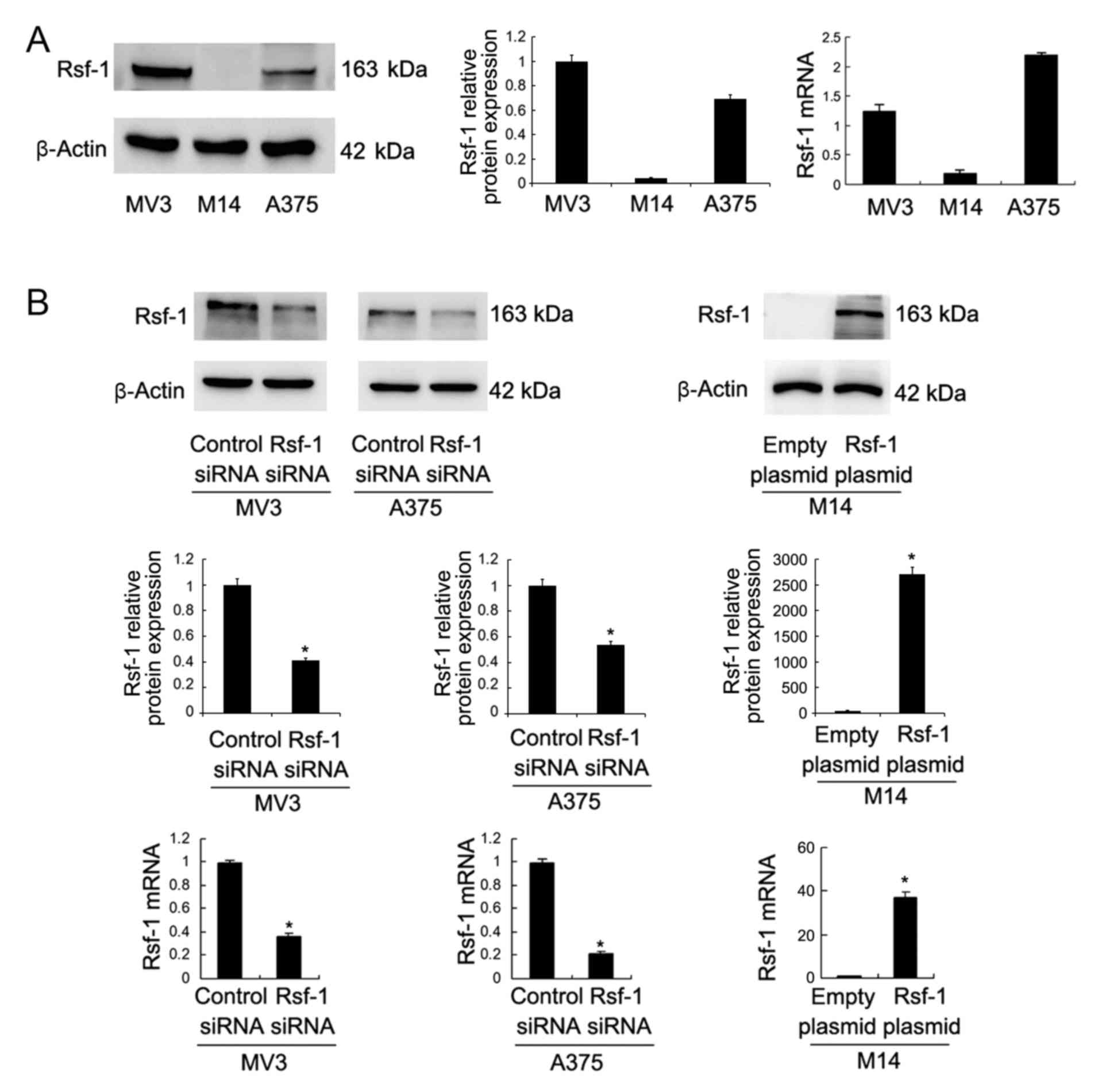
Rsf-1 expression levels in malignant melanoma cell
lines (MV3, M14 and A375) were investigated using western blotting
and RT-qPCR. It was revealed that the Rsf-1 expression levels were
low in M14 cells, and high in MV3 and A375 cell lines (Fig. 2A). To investigate the biological
roles of Rsf-1 in malignant melanoma, Rsf-1 siRNA-mediated
interference was performed in the MV3 and A375 melanoma cell lines,
whilst Rsf-1-encoding plasmid transfection was performed in the M14
cell line. As presented in Fig.
2B, Rsf-1 siRNA significantly downregulated Rsf-1 protein and
mRNA expression, whereas the Rsf-1 plasmid significantly
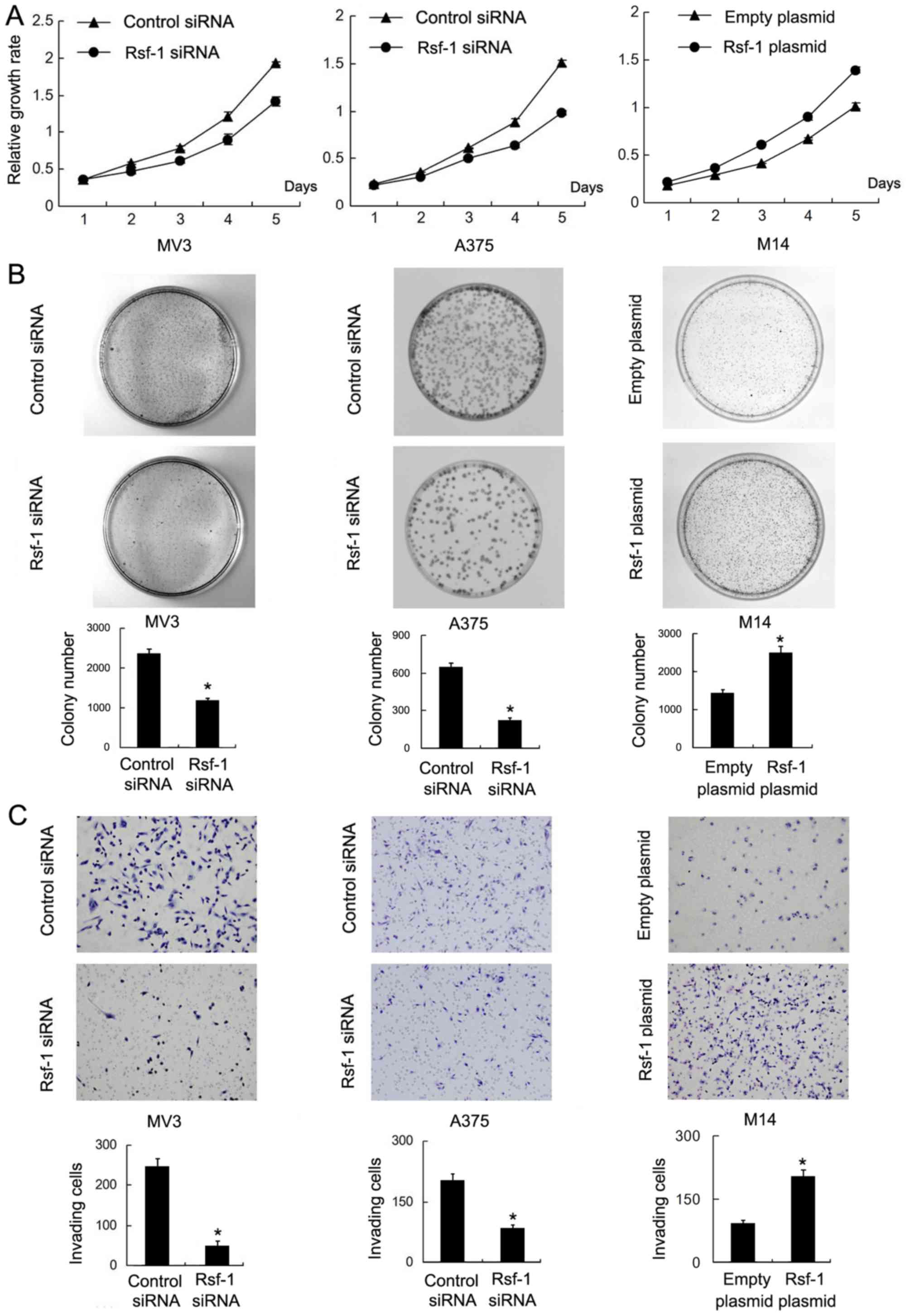
upregulated Rsf-1 expression. An MTT assay was performed to
investigate cell viability, which demonstrated that Rsf-1 depletion
in MV3 and A375 cells notably decreased cell viability compared
with the controls, whereas Rsf-1 overexpression in M14 cells
markedly promoted cell viability (Fig.
3A). A colony formation assay also revealed that Rsf-1
depletion significantly decreased colony number in the MV3 and A375
cell lines, while Rsf-1 overexpression significantly increased the
colony number in the M14 cell line compared with the control
(Fig. 3B). To determine the
effects of Rsf-1 on cell invasion, a Transwell invasion assay was
performed. The results revealed that the number of invasive cells
was significantly reduced following Rsf-1 depletion in the MV3
(control siRNA vs. Rsf-1 siRNA, 243±20 vs. 52±12 cells; P<0.05;
Fig. 3C) and A375 cell lines
(control siRNA vs. Rsf-1 siRNA, 214±15 vs. 90±8 cells; P<0.05)
compared with the control. Conversely, Rsf-1 overexpression
significantly increased the invasive ability of M14 cells compared
with the control (empty plasmid vs. Rsf-1 plasmid, 100±7 vs. 221±15
cells; P<0.05).
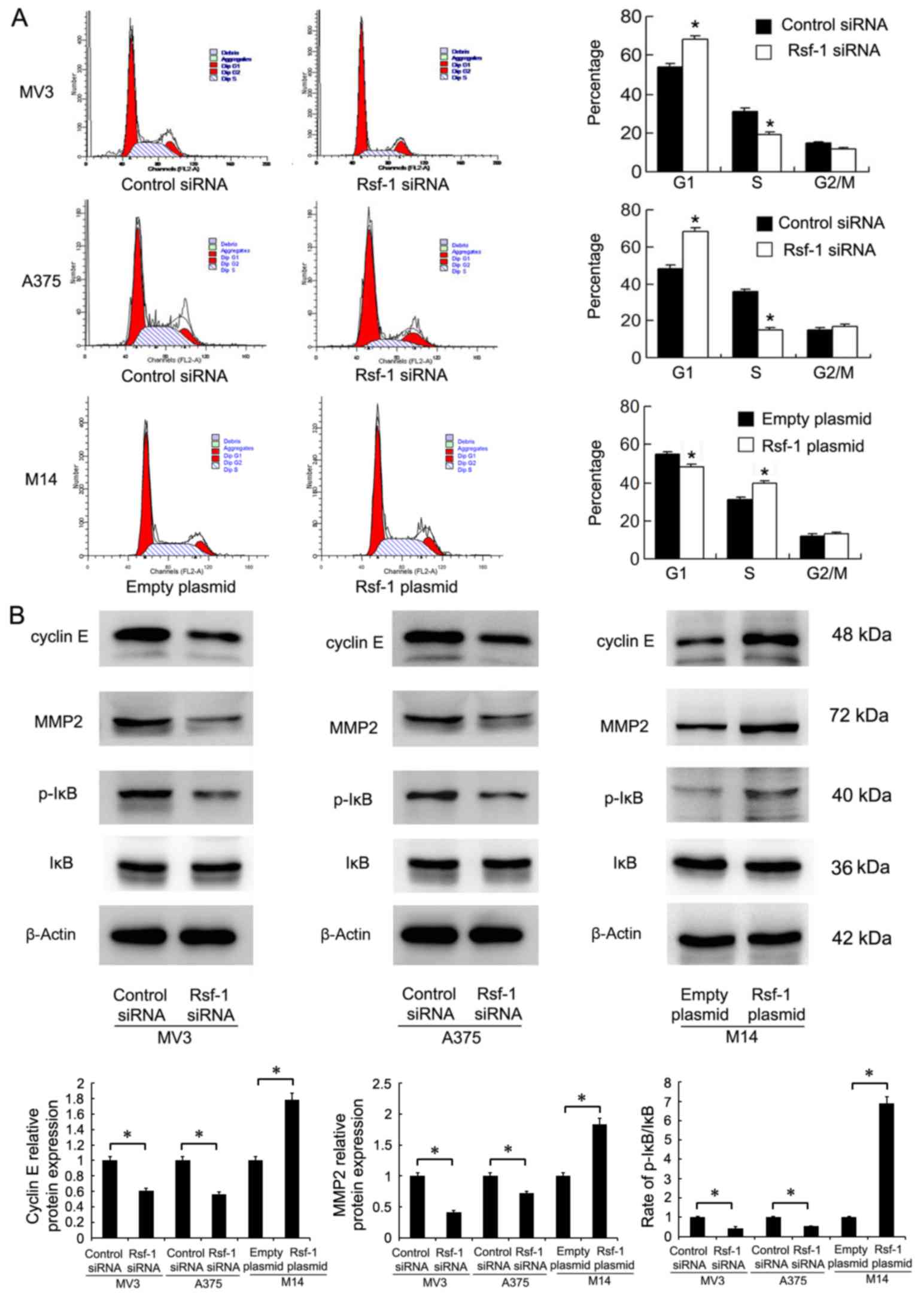
Rsf-1 regulates cell cycle progression
and associated protein expression
Cell cycle analysis was performed in melanoma cell
lines. Rsf-1 depletion in MV3 and A375 cells significantly
increased the percentage of cells in G1 phase and decreased that in
S phase compared with the control (Fig. 4A). Rsf-1 overexpression in M14
cells had opposing effects; the percentage of cells in G1 phase was
significantly reduced, while the percentage of cells in S phase
increased compared with the control, suggesting that Rsf-1
depletion suppresses G1/S cell cycle transition (Fig. 4A). To analyze the potential
molecular mechanisms underlying the effects of Rsf-1 on the cell
cycle, the expression of associated proteins was examined by
western blotting. As presented in Fig.
4B, the expression levels of MMP2, cyclin E and p-IκB were
decreased in Rsf-1-depleted MV3 and A375 cells compared with
control cells. Conversely, Rsf-1 overexpression upregulated MMP2,
cyclin E and p-IκB expression in M14 cells.
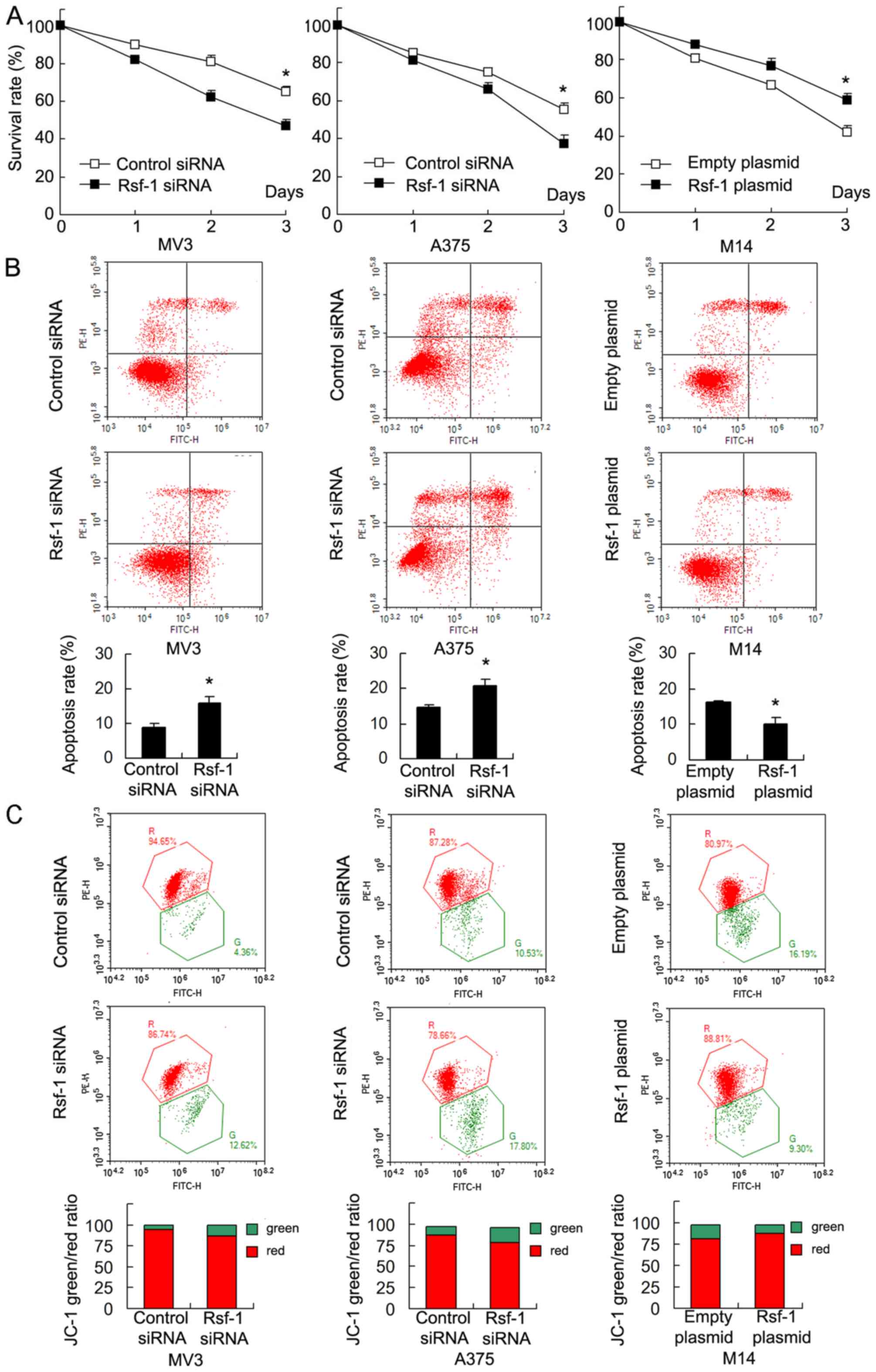
Rsf-1 enhances cisplatin resistance
and MMP
To investigate the role of Rsf-1 in the
chemoresistance of malignant melanoma cells, Rsf-1 depleted and
control cells were treated with cisplatin (5 µM). The results of
the MTT assay revealed that Rsf-1 siRNA significantly decreased
cell survival rate following 3 days of cisplatin treatment in MV3
and A375 cells compared with the control. Rsf-1 overexpression
significantly increased cell viability in M14 cells treated with
cisplatin (Fig. 5A). Furthermore,
apoptosis analysis revealed that the cell apoptotic rate was
significantly increased following Rsf-1 depletion in MV3 and A375
cells treated with cisplatin, and reduced in Rsf-1-overexpressing
M14 cells treated with cisplatin compared with the control
(Fig. 5B). Collectively, the
results demonstrated that Rsf-1 expression promotes cisplatin
resistance in melanoma cells.
As resistance to chemotherapeutic drugs is closely
associated with mitochondrial function, whether Rsf-1 affected the
MMP was investigated. JC-1 staining was used to monitor alterations
in MMP following cisplatin treatment. JC-1 staining exhibits red
fluorescence under normal conditions; however, green fluorescence
is observed when the MMP is depolarized following cisplatin
treatment. As presented in Fig.
5C, in M14 cells treated with cisplatin, Rsf-1 overexpression
notably decreased the percentage of cells exhibiting green
fluorescence, suggesting that Rsf-1 promoted mitochondrial membrane
polarization. Conversely, Rsf-1 depletion led to notable
depolarization of the MMP in MV3 and A375 cells treated with
cisplatin.
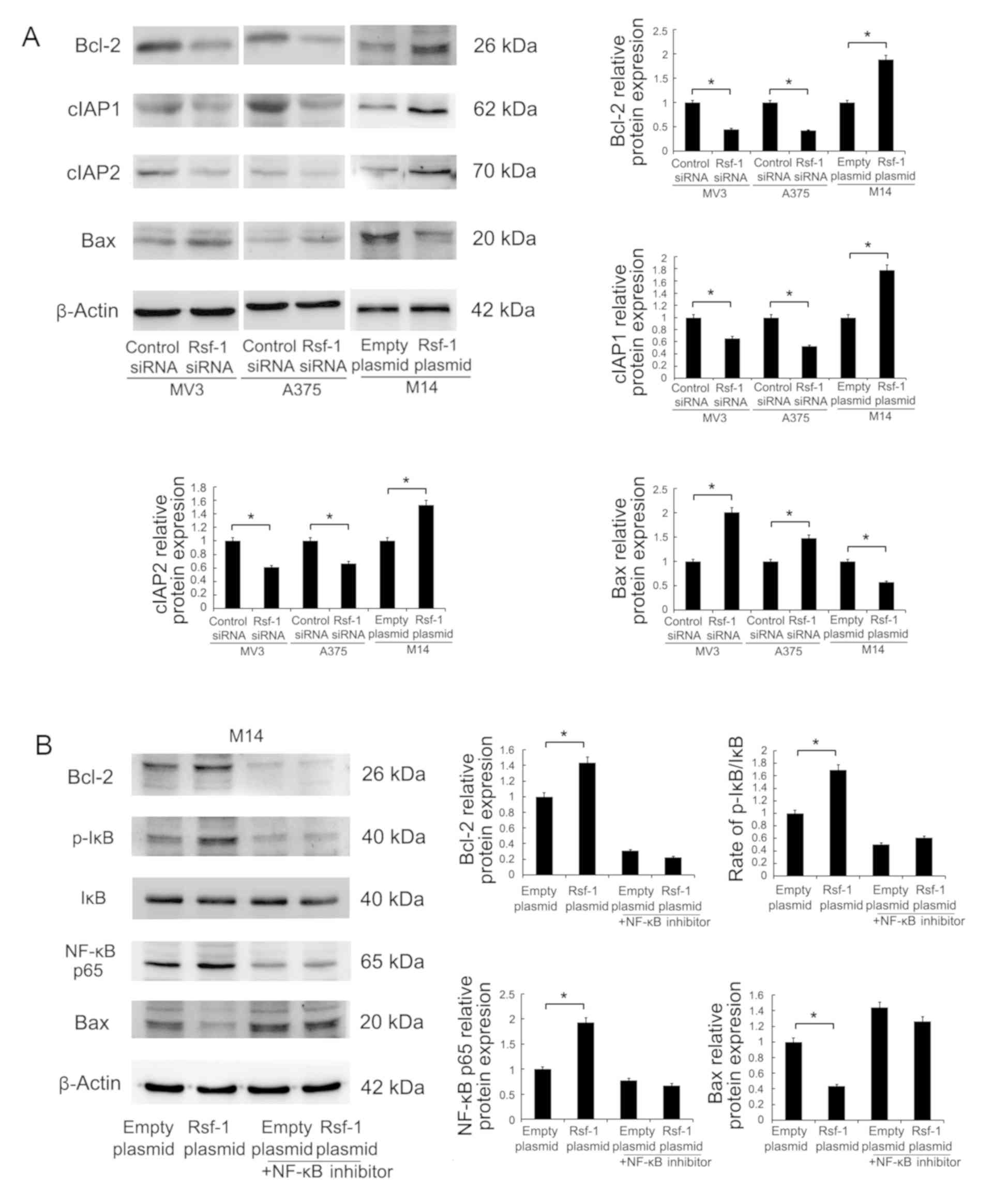
Rsf-1 regulates Bcl-2 expression via
NF-κB signaling
Furthermore, Rsf-1-induced alterations in
apoptosis-associated protein expression were investigated via
western blot analysis (Fig. 6A).
Rsf-1 depletion downregulated Bcl-2, cIAP1 and cIAP2 expression
levels, and upregulated Bax expression in MV3 and A375 cell lines
compared with the control; opposing effects were observed in
Rsf-1-overexpressing M14 cells. As Rsf-1 positively regulates
p-IκB, and Bcl-2 expression was reported as associated with NF-κB
signaling (32), whether Rsf-1
regulated the activity of Bcl-2 via its effects on NF-κB was
investigated. To validate this, M14 cells were treated with NF-κB
inhibitor (10 µM). As presented in Fig. 6B, NF-κB inhibition significantly
downregulated p-IκB and NF-κB p65 protein levels in control and
Rsf-1 plasmid-transfected M14 cells. Furthermore, treatment with
the NF-κB inhibitor eliminated the effects of Rsf-1 upregulation on
Bcl-2 expression.
Discussion
Previous studies have reported that Rsf-1
overexpression occurs in numerous cancers, including ovarian
cancer, breast cancer, nasopharyngeal carcinoma, non-small cell
lung cancer, gastric adenocarcinoma and colon cancer (20–23,25,26,28,33–35);
however, its involvement in melanoma has not been investigated. In
the present study, the expression of Rsf-1 was analyzed in 50
malignant melanoma specimens via immunohistochemistry.
Overexpression of Rsf-1 was detected in 19 cases, which was
positively associated with advanced TNM stage. As surgical therapy
is not the preferred treatment for patients with advanced
melanomas, particularly stage IV melanoma, the number of stage III
and IV melanoma specimens analyzed was markedly lower than that for
stage II melanoma. It was observed that the incidence of Rsf-1
overexpression was notably higher in melanomas with advanced TNM
stage (III and IV vs. II). Accordingly, overexpression of Rsf-1 was
reported to be associated with advanced TNM stage, nodal metastasis
and poorly differentiated tumor cells in other cancers (20,22,23,33,34).
Thus, Rsf-1 tends to be overexpressed in advanced stage melanomas,
suggesting its association with the malignant progression of
melanoma cells. To the best of our knowledge, the present study is
the first to demonstrate the clinical significance of Rsf-1 in
melanoma.
Rsf-1 has been reported to regulate cell behaviors
including proliferation, invasion and cell cycle progression
(16,26,36–39).
A previous study reported that Rsf-1 depletion significantly
decreased the proliferation rate and colony formation ability in
colon cancer cell lines HT29 and HCT116 (23). Rsf-1 depletion also inhibited
proliferation in lung cancer cells (21). The present findings support these
previous reports, demonstrating that Rsf-1 depletion decreased cell
viability and colony number, while its overexpression promoted
viability. For cell invasion, it was observed that the number of
invading cells decreased significantly following Rsf-1 depletion,
but markedly increased following overexpression of Rsf-1, which was
consistent with previous reports demonstrating that Rsf-1 depletion
inhibited invasiveness in the prostate cancer cell line DU145
(20) and lung cancer cells
(30).
The regulatory effect of Rsf-1 on cell viability
suggested that Rsf-1 may serve an important role in cell cycle
progression. The present study revealed that Rsf-1 depletion
enhanced the percentage of G1 phase cells and downregulated that of
S phase, demonstrating that Rsf-1 can facilitate G1/S transition.
Western blotting revealed that Rsf-1 depletion decreased the levels
of MMP2, cyclin E and p-IκB, consistent with previous reports of
MMP2 downregulation following Rsf-1 depletion in lung cancer cells
(30). MMP2 is a member of the
matrix metalloproteinase family, the members of which are involved
in various pathological and physiological processes, including
cancer cell growth, invasion and metastasis, suggesting that Rsf-1
regulates melanoma invasion via MMP2 (40). However, the possibility that other
effects regulated by Rsf-1 may also be responsible for its effects
on invasion cannot be excluded.
Transfection of cell lines exhibiting high Rsf-1
expression with siRNA targeted against Rsf-1 also increased the
rate of apoptosis, which may also contribute to the decreased
invasive ability of cells following Rsf-1 depletion. Cyclin E
serves an essential role in fundamental biological processes,
including cell cycle control and DNA replication (41–43).
Sheu et al (13) revealed
that cyclin E1 interacts with the first 441 amino acids of Rsf-1,
and that their interaction promotes G1-S transition. Additionally,
Rsf-1 depletion downregulated cyclin E in hepatocellular carcinoma
(25). These reports further
support the findings of the present study.
Furthermore, the present study proposed that Rsf-1
positively regulated the chemoresistance of melanoma cells, which
has not been previously reported, to the best of our knowledge. In
cells treated with cisplatin, MTT and Annexin V/PI analysis were
performed to examine the effects of Rsf-1. The cell survival rate
decreased, while the apoptotic rate increased significantly
following Rsf-1 depletion. The role of Rsf-1 in chemoresistance has
been indicated in various cancers including ovarian cancer
(28), lung cancer (44) and glioma (36); however, the association between
Rsf-1 and mitochondrial regulation has not yet been reported.
Mitochondrial function serves an important role in the development
of chemoresistance. Depolarization of the MMP induces apoptosis via
the mitochondria-dependent pathway (45). It was demonstrated that Rsf-1
depletion depolarized the MMP, with opposing effects observed
following Rsf-1 overexpression in M14 cells. To the best of our
knowledge, the present study is the first to report of the
association between the role of Rsf-1 in chemoresistance and the
regulation of mitochondrial function.
It was revealed that expression of the pro-apoptotic
protein Bax increased, while the levels of anti-apoptotic proteins,
including cIAP1, cIAP2 and Bcl-2 decreased significantly following
Rsf-1 depletion, as reported in previous studies (46–48);
Rsf-1 overexpression induced opposing effects. cIAP1 and cIAP2 are
members of the IAP family, which regulate apoptosis and
chemoresistance (49).
The NF-κB signaling pathway is induced via
activation of IκB, and is involved in numerous biological
processes, including cell growth, tumorigenesis and apoptosis
(50). Bcl-2 is a downstream
effector of NF-κB, and serves as an important anti-apoptotic
mediator in melanoma (51,52). The present study proposed that
Rsf-1 could positively regulate the NF-κB pathway via upregulation
of p-IκB. NF-κB signaling was considered particularly noteworthy
for two reasons. A previous study using Ingenuity Pathways Analysis
Systems revealed that various molecular hubs including NF-kB,
extracellular signal-regulated kinase (ERK) and protein kinase B
(Akt) were identified in an Rsf-1-regulated gene network (28). In addition, analysis of numerous
other signaling pathways was conducted, including p-ERK and p-Akt
(data not shown); however, significant alterations were not
observed in the expression profile of these proteins (data not
shown). Notable alterations in p-IκB expression were observed.
Thus, the NF-κB pathway was selected for further study, and its
importance was confirmed via the use of an NF-κB inhibitor. Rsf-1
overexpression failed to induce Bcl-2 upregulation in cells treated
by NF-κB inhibitor, supporting the association between Rsf-1 and
Bcl-2 in melanoma cells.
There are two novel points to highlight based upon
the findings of the present study. The clinical significance of
Rsf-1, which has not been previously reported in melanoma, was
demonstrated in this study. Additionally, the role of Rsf-1 in
chemosensitivity was associated with mitochondrial function. In
conclusion, the present study demonstrated that Rsf-1 is
overexpressed in malignant melanoma, and may contribute to the
proliferation, invasion and cell cycle progression of malignant
cells by modulating the expression of MMP2, cyclin E and NF-κB.
Furthermore, Rsf-1 may regulate chemoresistance and MMP in melanoma
cells, with concomitant alterations in cIAP1, cIAP2, Bax and Bcl-2
protein expression. Thus, Rsf-1 may serve as a potential
therapeutic target in the treatment of malignant melanoma.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JH and LF performed the experiments, evaluated the
data, drafted the manuscript and prepared the figures. QL made
significant contributions towards the design of the study,
evaluated the data and drafted the manuscript. All authors reviewed
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of China Medical University (Shenyang, China). Informed
consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Merrill SJ, Subramanian M and Godar DE:
Worldwide cutaneous malignant melanoma incidences analyzed by sex,
age, and skin type over time (1955–2007): Is HPV infection of
androgenic hair follicular melanocytes a risk factor for developing
melanoma exclusively in people of European-ancestry?
Dermatoendocrinol. 8:e12153912016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li H, Pedersen L, Nørgaard M, Ulrichsen
SP, Thygesen SK and Nelson JJ: The occurrence of non-melanoma
malignant skin lesions and non-cutaneous squamous-cell carcinoma
among metastatic melanoma patients: An observational cohort study
in Denmark. BMC Cancer. 16:2952016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nahar VK, Allison Ford M, Brodell RT,
Boyas JF, Jacks SK, Biviji-Sharma R, Haskins MA and Bass MA: Skin
cancer prevention practices among malignant melanoma survivors: A
systematic review. J Cancer Res Clin Oncol. 142:1273–1283. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peterson M, Albertini MR and Remington P:
Remington, incidence, survival, and mortality of malignant
cutaneous melanoma in wisconsin, 1995–2011. WMJ. 114:196–201.
2015.PubMed/NCBI
|
|
5
|
Johnson-Obaseki SE, Labajian V, Corsten MJ
and McDonald JT: Incidence of cutaneous malignant melanoma by
socioeconomic status in Canada: 1992–2006. J Otolaryngol Head Neck
Surg. 44:532015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dzambova M, Sečníková Z, Jiráková A,
Jůzlová K, Viklický O, Hošková L, Göpfertovà D and Hercogová J:
Malignant melanoma in organ transplant recipients: Incidence,
outcomes, and management strategies: A review of literature.
Dermatol Ther. 29:64–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brewer JD, Shanafelt TD, Call TG, Cerhan
JR, Roenigk RK, Weaver AL and Otley CC: Increased incidence of
malignant melanoma and other rare cutaneous cancers in the setting
of chronic lymphocytic leukemia. Int J Dermatol. 54:e287–e293.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang HY, Feng HL, Wang L, Chou P and Wang
PF: The Incidence, prevalence, and survival of malignant melanoma
in Taiwan. Value Health. 17:A7402014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nowak-Sadzikowska J, Walasek T, Jakubowicz
J, Blecharz P and Reinfuss M: Current treatment options of brain
metastases and outcomes in patients with malignant melanoma. Rep
Pract Oncol Radiother. 21:271–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schmid-Wendtner M and Wendtner CM:
Treatment of metastatic malignant melanoma. Dtsch Med Wochenschr.
141:10002016.(In German). PubMed/NCBI
|
|
11
|
Kozovska Z, Gabrisova V and Kucerova L:
Malignant melanoma: Diagnosis, treatment and cancer stem cells.
Neoplasma. 63:510–517. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Harries M, Malvehy J, Lebbe C, Heron L,
Amelio J, Szabo Z and Schadendorf D: Treatment patterns of advanced
malignant melanoma (stage III–IV)-A review of current standards in
Europe. Eur J Cancer. 60:179–189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sheu JJ, Choi JH, Guan B, Tsai FJ, Hua CH,
Lai MT, Wang TL and Shih IeM: Rsf-1, a chromatin remodelling
protein, interacts with cyclin E1 and promotes tumour development.
J Pathol. 229:559–568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hanai K, Furuhashi H, Yamamoto T, Akasaka
K and Hirose S: RSF governs silent chromatin formation via histone
H2Av replacement. PLoS Genet. 4:e10000112008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang PI, Wu LC, Sheu JJ, Wu TF, Shen KH,
Wang YH, Wu WR, Shiue YL, Huang HY, Hsu HP, et al: Rsf-1/HBXAP
overexpression is independent of gene amplification and is
associated with poor outcome in patients with urinary bladder
urothelial carcinoma. J Clin Pathol. 65:802–807. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Min S, Jo S, Lee HS, Chae S, Lee JS, Ji JH
and Cho H: ATM-dependent chromatin remodeler Rsf-1 facilitates DNA
damage checkpoints and homologous recombination repair. Cell Cycle.
13:666–677. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goldfarb DM, Gukova LA, Chernin LS,
Avdienko ID, Mnatsakanian GG, Kushner IC, Kuznetsova VN and
Strachova TS: Rsf mutants of Escherichia coli HfrC defective in the
production of the factor stimulating recombination in conjugation.
Mol Gen Genet. 129:295–310. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Iwasa H, Kuroyanagi H, Maimaiti S, Ikeda
M, Nakagawa K and Hata Y: Characterization of RSF-1, the
Caenorhabditis elegans homolog of the Ras-association domain family
protein 1. Exp Cell Res. 319:1–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fang FM, Li CF, Huang HY, Lai MT, Chen CM,
Chiu IW, Wang TL, Tsai FJ, Shih IeM and Sheu JJ: Overexpression of
a chromatin remodeling factor, RSF-1/HBXAP, correlates with
aggressive oral squamous cell carcinoma. Am J Pathol.
178:2407–2415. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li H, Zhang Y, Zhang Y, Bai X, Peng Y and
He P: Rsf-1 overexpression in human prostate cancer, implication as
a prognostic marker. Tumour Biol. 35:5771–5776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Q, Dong Q and Wang E: Rsf-1 is
overexpressed in non-small cell lung cancers and regulates cyclinD1
expression and ERK activity. Biochem Biophys Res Commun. 420:6–10.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin CY, Tian YF, Wu LC, Chen LT, Lin LC,
Hsing CH, Lee SW, Sheu MJ, Lee HH, Wang YH, et al: Rsf-1 expression
in rectal cancer: With special emphasis on the independent
prognostic value after neoadjuvant chemoradiation. J Clin Pathol.
65:687–692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu S, Dong Q and Wang E: Rsf-1
overexpression correlates with poor prognosis and cell
proliferation in colon cancer. Tumour Biol. 33:1485–1491. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maeda D, Chen X, Guan B, Nakagawa S, Yano
T, Taketani Y, Fukayama M, Wang TL and Shih IeM: Rsf-1 (HBXAP)
expression is associated with advanced stage and lymph node
metastasis in ovarian clear cell carcinoma. Int J Gynecol Pathol.
30:30–35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xie C, Fu L, Xie L, Liu N and Li Q: Rsf-1
overexpression serves as a prognostic marker in human
hepatocellular carcinoma. Tumour Biol. 35:7595–7601. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tai HC, Huang HY, Lee SW, Lin CY, Sheu MJ,
Chang SL, Wu LC, Shiue YL, Wu WR, Lin CM and Li CF: Associations of
Rsf-1 overexpression with poor therapeutic response and worse
survival in patients with nasopharyngeal carcinoma. J Clin Pathol.
65:248–253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sheu JJ, Choi JH, Yildiz I, Tsai FJ, Shaul
Y, Wang TL and Shih IeM: The roles of human sucrose nonfermenting
protein 2 homologue in the tumor-promoting functions of Rsf-1.
Cancer Res. 68:4050–4057. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Choi JH, Sheu JJ, Guan B, Jinawath N,
Markowski P, Wang TL and Shih IeM: Functional analysis of 11q13.5
amplicon identifies Rsf-1 (HBXAP) as a gene involved in paclitaxel
resistance in ovarian cancer. Cancer Res. 69:1407–1415. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Trinidad CM, Torres-Cabala CA, Curry JL,
Prieto VG and Aung PP: Update on eighth edition American Joint
Committee on Cancer classification for cutaneous melanoma and
overview of potential pitfalls in histological examination of
staging parameters. J Clin Pathol. 72:265–270. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang X, Fu L, Xue D, Zhang X, Hao F, Xie
L, He J, Gai J, Liu Y, Xu H, et al: Overexpression of Rsf-1
correlates with poor survival and promotes invasion in non-small
cell lung cancer. Virchows Arch. 470:553–560. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jang JH and Surh YJ: Bcl-2 attenuation of
oxidative cell death is associated with up-regulation of
gamma-glutamylcysteine ligase via constitutive NF-kappaB
activation. J Biol Chem. 279:38779–38786. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sheu JJ, Guan B, Choi JH, Lin A, Lee CH,
Hsiao YT, Wang TL, Tsai FJ and Shih IeM: Rsf-1, a chromatin
remodeling protein, induces DNA damage and promotes genomic
instability. J Biol Chem. 285:38260–38269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mao TL, Hsu CY, Yen MJ, Gilks B, Sheu JJ,
Gabrielson E, Vang R, Cope L, Kurman RJ, Wang TL and Shih IeM:
Expression of Rsf-1, a chromatin-remodeling gene, in ovarian and
breast carcinoma. Hum Pathol. 37:1169–1175. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Davidson B, Trope' CG, Wang TL and Shih
IeM: Expression of the chromatin remodeling factor Rsf-1 is
upregulated in ovarian carcinoma effusions and predicts poor
survival. Gynecol Oncol. 103:814–819. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao XC, An P, Wu XY, Zhang LM, Long B,
Tian Y, Chi XY and Tong DY: Overexpression of hSNF2H in glioma
promotes cell proliferation, invasion, and chemoresistance through
its interaction with Rsf-1. Tumour Biol. 37:7203–7212. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ren J, Chen QC, Jin F, Wu HZ, He M, Zhao
L, Yu ZJ, Yao WF, Mi XY, Wang EH and Wei MJ: Overexpression of
Rsf-1 correlates with pathological type, p53 status and survival in
primary breast cancer. Int J Clin Exp Pathol. 7:5595–5608.
2014.PubMed/NCBI
|
|
38
|
Chae S, Ji JH, Kwon SH, Lee HS, Lim JM,
Kang D, Lee CW and Cho H: HBxAPalpha/Rsf-1-mediated HBx-hBubR1
interactions regulate the mitotic spindle checkpoint and chromosome
instability. Carcinogenesis. 34:1680–1688. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hu BS, Yu HF, Zhao G and Zha TZ: High
RSF-1 expression correlates with poor prognosis in patients with
gastric adenocarcinoma. Int J Clin Exp Pathol. 5:668–673.
2012.PubMed/NCBI
|
|
40
|
Rotte A, Martinka M and Li G: MMP2
expression is a prognostic marker for primary melanoma patients.
Cell Oncol (Dordr). 35:207–216. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Santala S, Talvensaari-Mattila A, Soini Y
and Santala M: Cyclin E expression correlates with Cancer-specific
survival in endometrial endometrioid adenocarcinoma. Anticancer
Res. 35:3393–3397. 2015.PubMed/NCBI
|
|
42
|
Alsina M, Landolfi S, Aura C, Caci K,
Jimenez J, Prudkin L, Castro S, Moreno D, Navalpotro B, Tabernero J
and Scaltriti M: Cyclin E amplification/overexpression is
associated with poor prognosis in gastric cancer. Ann Oncol.
26:438–439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Deng W, Zhou Y, Tiwari AF, Su H, Yang J,
Zhu D, Lau VM, Hau PM, Yip YL, Cheung AL, et al: p21/Cyclin E
pathway modulates anticlastogenic function of Bmi-1 in cancer
cells. Int J Cancer. 136:1361–1370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li HC, Chen YF, Feng W, Cai H, Mei Y,
Jiang YM, Chen T, Xu K and Feng DX: Loss of the Opa interacting
protein 5 inhibits breast cancer proliferation through
miR-139-5p/NOTCH1 pathway. Gene. 603:1–8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen X, Wong JY, Wong P and Radany EH:
Low-dose valproic acid enhances radiosensitivity of prostate cancer
through acetylated p53-dependent modulation of mitochondrial
membrane potential and apoptosis. Mol Cancer Res. 9:448–461. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Matsuyama S, Palmer J, Bates A,
Poventud-Fuentes I, Wong K, Ngo J and Matsuyama M: Bax-induced
apoptosis shortens the life span of DNA repair defect Ku70-knockout
mice by inducing emphysema. Exp Biol Med (Maywood). 241:1265–1271.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gill C, Dowling C, O'Neill AJ and Watson
RW: Effects of cIAP-1, cIAP-2 and XIAP triple knockdown on prostate
cancer cell susceptibility to apoptosis, cell survival and
proliferation. Mol Cancer. 8:392009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vassina EM, Yousefi S, Simon D, Zwicky C,
Conus S and Simon HU: cIAP-2 and survivin contribute to
cytokine-mediated delayed eosinophil apoptosis. Eur J Immunol.
36:1975–1984. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gyrd-Hansen M and Meier P: IAPs: From
caspase inhibitors to modulators of NF-kappaB, inflammation and
cancer. Nat Rev Cancer. 10:561–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hussain AR, Ahmed SO, Ahmed M, Khan OS, Al
Abdulmohsen S, Platanias LC, Al-Kuraya KS and Uddin S: Cross-talk
between NFkB and the PI3-kinase/AKT pathway can be targeted in
primary effusion lymphoma (PEL) cell lines for efficient apoptosis.
PLoS One. 7:e399452012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Benimetskaya L, Ayyanar K, Kornblum N,
Castanotto D, Rossi J, Wu S, Lai J, Brown BD, Popova N, Miller P,
et al: Bcl-2 protein in 518A2 melanoma cells in vivo and in vitro.
Clin Cancer Res. 12:4940–4948. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Leiter U, Schmid RM, Kaskel P, Peter RU
and Krähn G: Antiapoptotic bcl-2 and bcl-xL in advanced malignant
melanoma. Arch Dermatol Res. 292:225–232. 2000. View Article : Google Scholar : PubMed/NCBI
|