Introduction
Chronic pancreatitis (CP) (ICD-10 code for recurrent
chronic pancreatitis: K86.1) is a progressive, irreversible disease
process characterized by chronic inflammation, glandular necrosis
and fibrosis, which leads to the impairment of pancreatic exocrine
and endocrine functions (1).
Patients with CP suffer from chronic abdominal pain, impaired
digestion, malnutrition, anorexia, diabetes and disease-related
complications, such as the formation of pseudocysts, which
negatively impact their quality of life (2). In the USA, the prevalence of CP has
been estimated to be 20–30 cases per 105 individuals in
the entire population (3,4). Although general and statistical
reports concerning the prevalence of CP do not exist in the
Republic of Korea, we predict that the prevalence of CP in Korea
may be 35–40 cases per 105 individuals from the reports
of Japanese individuals (5). The
mortality rate of CP patients is higher than that of the general
population, and the survival at 10 years after the onset of CP is
estimated at 69–80% (6,7). The leading cause of death includes
pancreatic cancer, with a higher incidence observed in CP patients
(8). The development of pancreatic
cancer may stem from the pancreatic inflammatory process, and it is
known that they have common risk factors such as alcohol and
smoking.
In the present study, a mouse model of repetitive
acute pancreatitis (RAP)-induced CP was used. This model is widely
used because of its ease of induction and good reproducibility. The
RAP mouse model is characterized histologically by extensive
inflammatory infiltrate, acinar cell atrophy and pancreatic
fibrotic lesions. Moreover, it exhibits the biochemical and
pathophysiological features of CP observed in humans, such as
fibrotic destruction of the pancreatic gland, and structural
impairment of exocrine and endocrine functions (9,10).
Accumulating genetic, clinical and experimental evidence supports
the hypothesis that CP is the result of multiple episodes of RAP
(11). Although many studies have
been conducted to elucidate the pathophysiology of CP, the
pathobiochemical and molecular mechanisms remain unclear.
Currently, no significant prophylactic or therapeutic agents are
clinically effective for the prevention of CP.
Pancreatic stellate cells (PSCs) play a crucial role
in the initiation and progression of pancreatic fibrogenesis in CP
(12). Under normal conditions,
these cells exist in the quiescent state. Once a cell is injured,
these quiescent cells convert to myofibroblast-like, α-smooth actin
(α-SMA)-positive cells that are highly proliferative and capable of
depositing fibrillar collagen in the interstitial spaces (13). The representative stimulatory
cytokine found on PSCs is transforming growth factor (TGF)-β, which
is generated by inflammatory mediators (14). TGF-β stimulates PSCs to synthesize
and secrete matrix proteins such as collagen and fibronectin
(9). Thus, the regulation of TGF-β
in the development of CP is a key factor for the treatment of
CP.
Piperine is a phenolic component of black pepper
(Piper nigrum) and long pepper (P. longum) (15), and has many beneficial biological
and pharmacological effects, such as anti-depressant,
hepatoprotective, anti-metastatic and anti-thyroid effects
(16). We previously reported the
protective effects of piperine against acute pancreatitis and
sepsis (17,18). However, currently, it is unclear
whether piperine can attenuate pancreatic fibrosis during CP.
Therefore, we aimed to determine the anti-inflammatory and
anti-fibrotic effects of piperine on cerulein-induced CP in mice.
We examined histological variations in the pancreas, and typical
characteristics such as activation of PSCs, deposition of
extracellular matrix (ECM), and production of cytokines (TNF-α,
IL-1β, IL-6 and TGF-β) and chemokines (CCL2 and CXCL2).
Furthermore, using primary mouse PSCs, the effects of piperine on
TGF-β-induced PSC activation and ECM formation were
demonstrated.
Materials and methods
Materials
Piperine, corn oil, cerulein, DAPI, hematoxylin,
eosin, xylene and ethanol were purchased from Sigma-Aldrich/Merck
KGaA. Easy-Blue™ Total RNA extraction kit and RIPA buffer were
purchased from iNtRON Biotechnology (Seongnam, Korea). Antibodies
against phospho-specific SMAD2/3 (cat. no. sc-11769), α-SMA (cat.
no. sc-32251), amylase (cat. no. sc-46657) and GAPDH (cat. no.
sc-32233) were purchased from Santa Cruz Biotechnology (Santa Cruz,
CA, USA). TGF-β (cat. no. ab66043) and collagen I (cat. no.
ab34710) were purchased from Abcam (Cambridge, MA, UK).
Phospho-specific SMAD1/5 (cat. no. 9516S), SMAD3 (cat. no. 9523S),
and SMAD5 (cat. no. 9517S) were purchased from Cell Signaling
Technology (Beverly, MA, USA).
Animals
All experiments were performed according to
protocols approved by the Animal Care Committee of Wonkwang
University (WKU15-18). In total, 180 female C57BL/6 mice (6–8 weeks
of age, weighing 15–20 g) were purchased from Orient Bio (Sungnam,
KyungKiDo, Korea). All animals were bred and housed in standard
shoebox cages in a climate-controlled room with an ambient
temperature of 23±2°C and a 12-h light-dark cycle for 7 days.
Animals were fed standard laboratory chow, allowed water ad
libitum, and randomly assigned to a control or experimental
group.
Experimental design
CP was induced by administering intraperitoneal
injections of cerulein (50 µg/kg, n=6 per group for three
experiments, total=18) six times at 1-h intervals, 5 times per
week, for a total of 3 weeks. In the pre-treatment groups, piperine
(1, 5, or 10 mg/kg, n=6 per group for three experiments, total=18)
or corn oil (control group, n=6 per group for three experiments,
total=18) was administrated orally 1 h before the first cerulein
injection, once a day, 5 times a week, for a total of 3 weeks. In
the post-treatment groups, piperine (10 mg/kg, n=6 per group for
three experiments, total=18) or corn oil was administered orally at
1 or 2 weeks after the first cerulein injection. Mice were
sacrificed at 24 h after the last cerulein injection via
CO2 asphyxiation. Pancreases were collected for further
analysis. All experiments were performed independently for three
times.
Histology
The pancreatic tissues were fixed overnight in 4%
paraformaldehyde, embedded in paraffin using standard methods, and
cut into serial sections (4-µm thick). The tissues were stained
with hematoxylin and eosin (H&E) for histological examination
via light microscopy. The tissue slides were blind-labeled and
reviewed by a pancreas pathologist who had no knowledge of the
experimental groups. The histopathological assessment of CP was
scored based on inflammation and glandular atrophy. Using a
previously described method (19),
a tissue section representing a minimum of 100 fields was examined
for each sample and scored on a scale of 0–3 (0 being normal and 3
being severe).
Immunofluorescence
Immunofluorescence assays for α-SMA and TGF-β was
performed on frozen pancreatic tissue sections by using a cryostat
microtome. The tissues were permeabilized with 0.1% Triton X-100
for 15 min at room temperature (RT), and blocked with 3% BSA for 1
h at RT. The sections were then incubated with the primary
antibodies against α-SMA (1:500 dilution in 3% BSA; Santa Cruz
Biotechnology) and TGF-β (1:500 dilution in 3% BSA; Cell Signaling
Technology) at 4°C overnight followed by the fluorescence-labeled
secondary antibodies Alexa Fluor 594 goat anti-mouse (cat. no.
A11005) and Alexa Fluor 568 goat anti-rabbit (cat. no. A11011) at
RT for 1 h (1:2,000 dilution in 3% BSA; Invitrogen; Thermo Fisher
Scientific, Inc.). Nuclei were counterstained with DAPI (1:2,000
dilution in 3% BSA; Vector Laboratories, Burlingame, CA, USA) for 5
min at RT. Stained sections were visualized using a confocal laser
microscope (Olympus, Japan).
Collagen staining
Sirius Red was used for staining of the collagen
deposition. Deparaffinized and rehydrated slides were immersed in
saturated picric acid for 20 min and rinsed in tap water until the
yellow stain in the specimens was cleared. The slides were then
stained with 0.1% Sirius Red solution (0.5 g of Direct Red 80;
Sigma-Aldrich; Merck KGaA) in 500 ml of picric acid for 1 h, washed
in two changes of acidified water (5 ml glacial acetic acid in 1 l
of water), and counterstained with hematoxylin. The relative
intensity was measured using Leica microscopy software (Wetzlar,
Germany).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using an Easy-Blue™ Total
RNA extraction kit according to the manufacturer's instructions and
reverse transcription of RNA to cDNA was performed using an ABI
cDNA synthesis kit (Applied Biosystems, Foster City, CA, USA)
(conditions: 37°C for 1 h, followed by 95°C for 5 min). TaqMan
quantitative RT-PCR with an ABI StepOne Plus detection system was
performed according to the manufacturer's instructions (Applied
Biosystems; Thermo Fisher Scientific, Inc.). For each sample,
triplicate test reactions and a control reaction without reverse
transcriptase were analyzed for expression of the gene of interest
and to control for variations in the reactions. All qPCR data were
normalized to levels against the housekeeping gene hypoxanthine
guanine phosphoribosyltransferase (HPRT). Forward, reverse, and
probe oligonucleotide primers for multiplex real-time TaqMan PCR
were purchased from ABI (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Cycling conditions were 50°C for 2 min, 95°C for
10 min, followed by 40 cycles of 95°C for 10 sec and 60°C for 30
sec. The data were analyzed using StepOne™ software (version 2.3;
Applied Biosystems; Thermo Fisher Scientific, Inc.). The
2−ΔΔCq method was used to determine the relative mRNA
expression level (20).
Isolation of mouse PSCs
PSCs were isolated from the C57BL/6 mice via
digestion of the pancreatic tissue and Nycodenz®
(Sigma-Aldrich; Merck KGaA) density gradient centrifugation, as
described previously (11).
Briefly, freshly isolated mouse PSCs were cultured in DMEM/High
Glucose supplemented with 10% fetal bovine serum and 1%
penicillin-streptomycin at 37°C with 5% CO2. Passages 3
to 6 were used.
Western blot analysis
PSCs were harvested and then lysed with lysis buffer
(1% cocktail of protease inhibitor and 1% phosphatase inhibitor in
1X RIPA buffer). The cells were boiled in sample buffer [62.5 mM
Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate (SDS), 20% glycerol,
and 10% 2-mercaptoethanol]. The protein concentrations were
determined by bicinchoninic acid assay. Total cell proteins (20 µg)
were separated via 10% SDS-polyacrylamide gel and transferred to a
PVDF membrane (GE Healthcare Life Sciences, Little Chalfont,
Buckinghamshire, UK). The membranes were blocked with 5% skim milk
in PBS-Tween-20 (PBST) for 2 h at RT and incubated with primary
antibodies (1:1,000) at 4°C overnight: α-SMA, collagen I, GAPDH,
pSMAD2/3, pSMAD1/5, SMAD3, SMAD5. After washing thrice, the
membranes were incubated with HRP-conjugated goat anti-mouse
(1:5,000; cat. no. 1031-05; Southern Biotech, Birmingham, AL, USA)
or goat anti-rabbit (1:5,000; cat. no. NB7156; Novus Biologicals,
LLC, Littleton, CO, USA) IgG secondary antibodies for 1 h at room
temperature. The protein bands were visualized using an enhanced
chemiluminescence detection system (Amersham; GE Healthcare),
according to the manufacturer's recommended protocol. Capture of
protein bands and quantitative analysis were performed using
Quantity One® software version 4,6,6 (Bio-Rad
Laboratories, Inc, Hercules, CA, USA).
Immunohistochemical analysis
Immunohistochemical staining for amylase was
performed using DAB immunohistochemical kit (Dako, Cytomation,
Denmark; as shown in Fig.
S1).
Statistical analysis
Results are expressed as the mean ± standard error
of the mean (SEM). Significance was evaluated using a two-way
analysis of variance (ANOVA), with time and dose parameters.
Significant ANOVA tests were further examined by post hoc analysis,
using the Duncan method as a multiple comparison among groups.
Values of P<0.05 were accepted as statistically significant.
Results
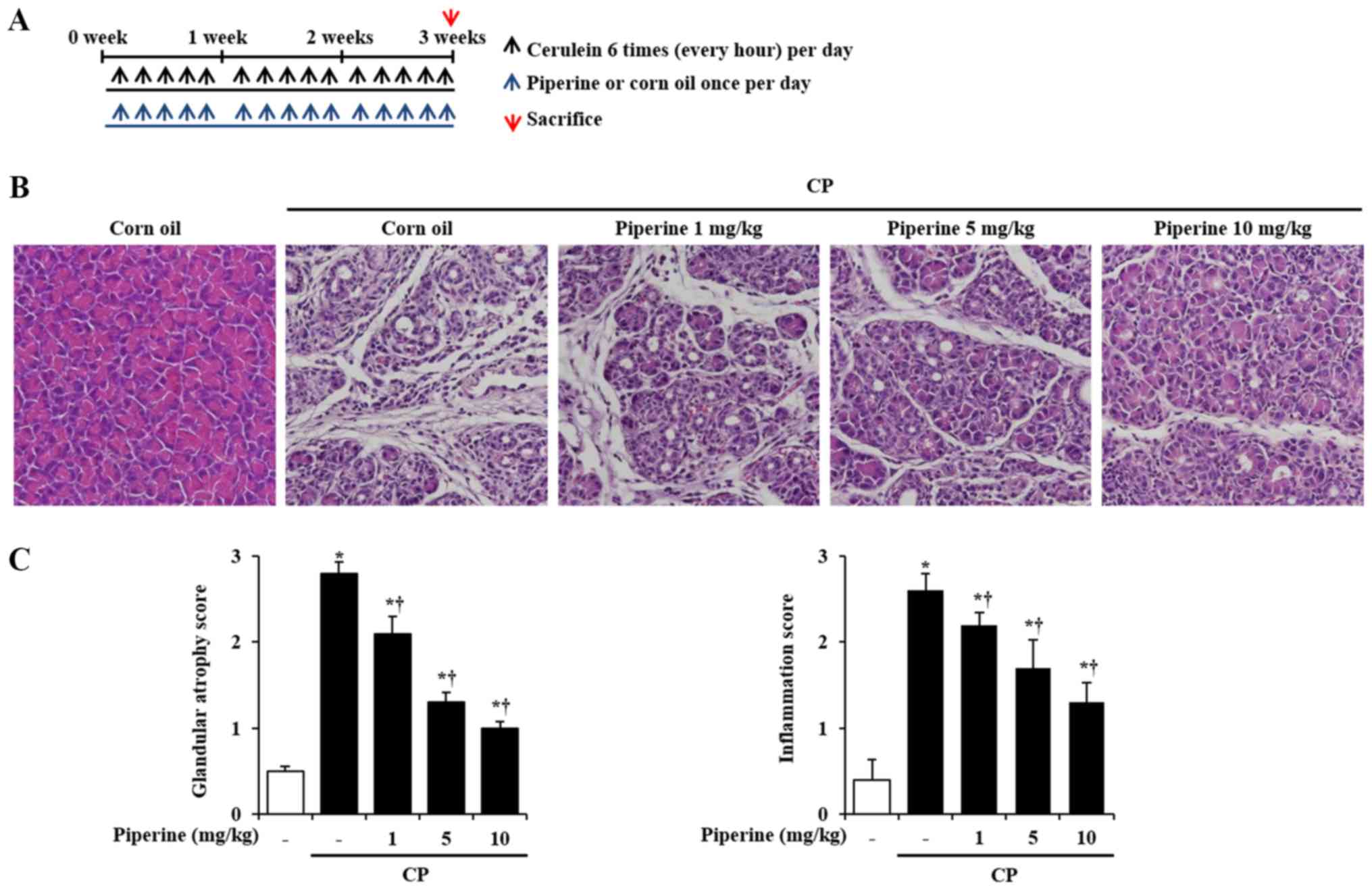
Morphological and histological
examination of the effects of piperine on the pancreases with
CP
To determine whether piperine can attenuate the
development of CP, the histological architecture of the pancreas
was investigated. Piperine had no adverse or harmful effects on
animals and no significant alterations were found between the corn
oil group and the piperine group with respect to the histology of
the pancreatic tissues and body weights (data not shown). In the
cerulein-induced CP mice, the histological features of the
pancreases showed severe pancreatic damage such as glandular
atrophy and inflammatory cell infiltration (Fig. 1B and C). However, these features
were inhibited by treatment with piperine in a dose-dependent
manner (Fig. 1B and C). To further
examine the effect of piperine on CP, the survival of pancreatic
acini, which secrete amylase, were assessed. Mice with CP showed
fewer pancreatic acini as indicated by the reduced levels of
amylase. However, piperine treatment significantly reduced acinar
cell death and destruction in the pancreases of the CP-induced mice
(Fig. S1).
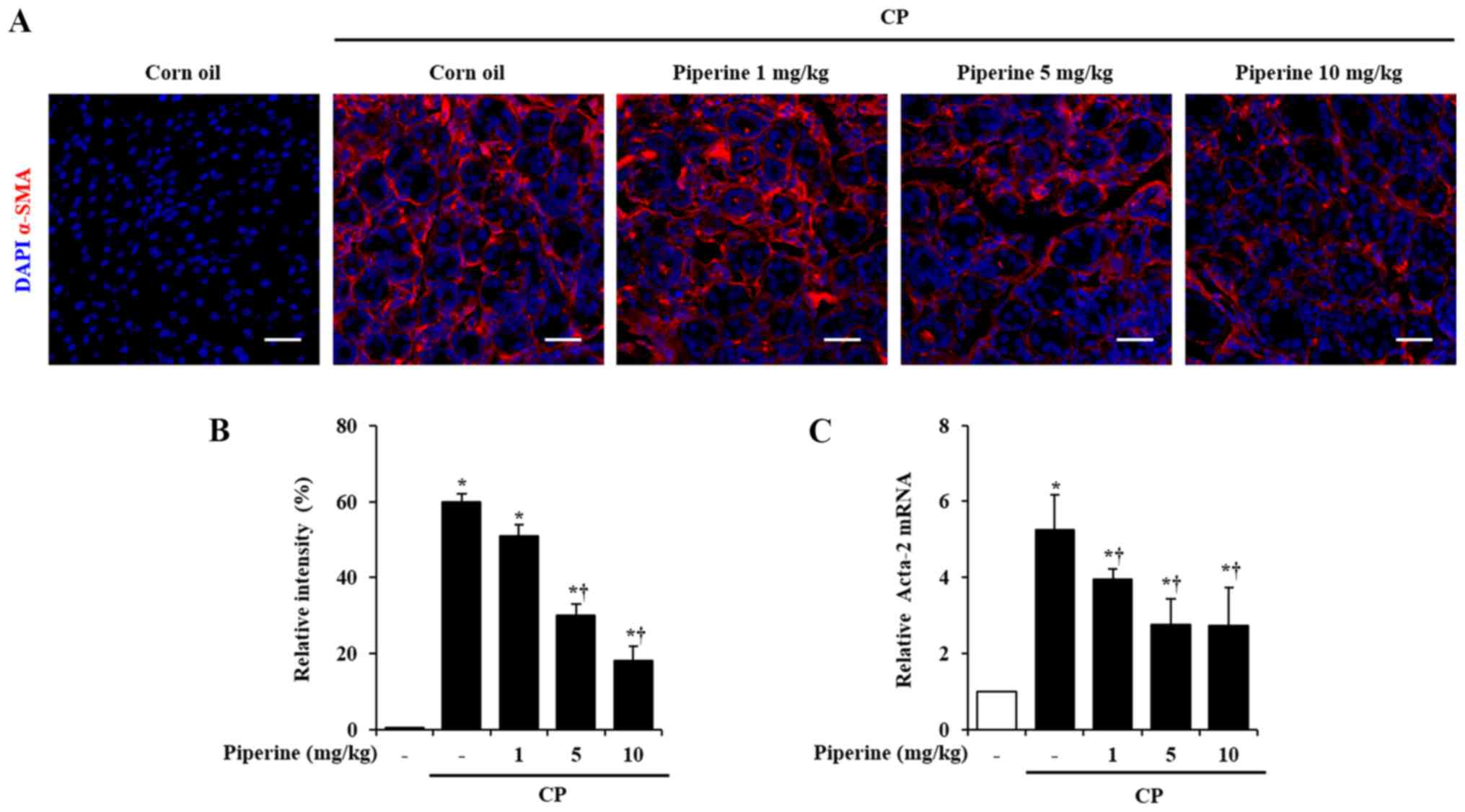
Effect of piperine on the activation
of PSCs during CP
Generally, PSCs are desmin-positive cells. When PSCs
are activated they transform into myofibrolast-like cells
expressing α-SMA (21). Therefore,
we examined the expression of α-SMA as a marker of activated PSCs
in the pancreas using immunofluorescence analysis. The
immunofluorescence analysis revealed that the number of
α-SMA-positive cells (in red color) were markedly increased in the
CP-induced mice. Piperine-treated mice showed decreased α-SMA
expression (Fig. 2A and B). To
confirm the above finding, the mRNA expression of Acta-2 for α-SMA
was assessed. Acta-2 level was markedly increased in the pancreas
of the CP-induced mice. However, this increase was significantly
reduced by treatment with piperine (Fig. 2C).
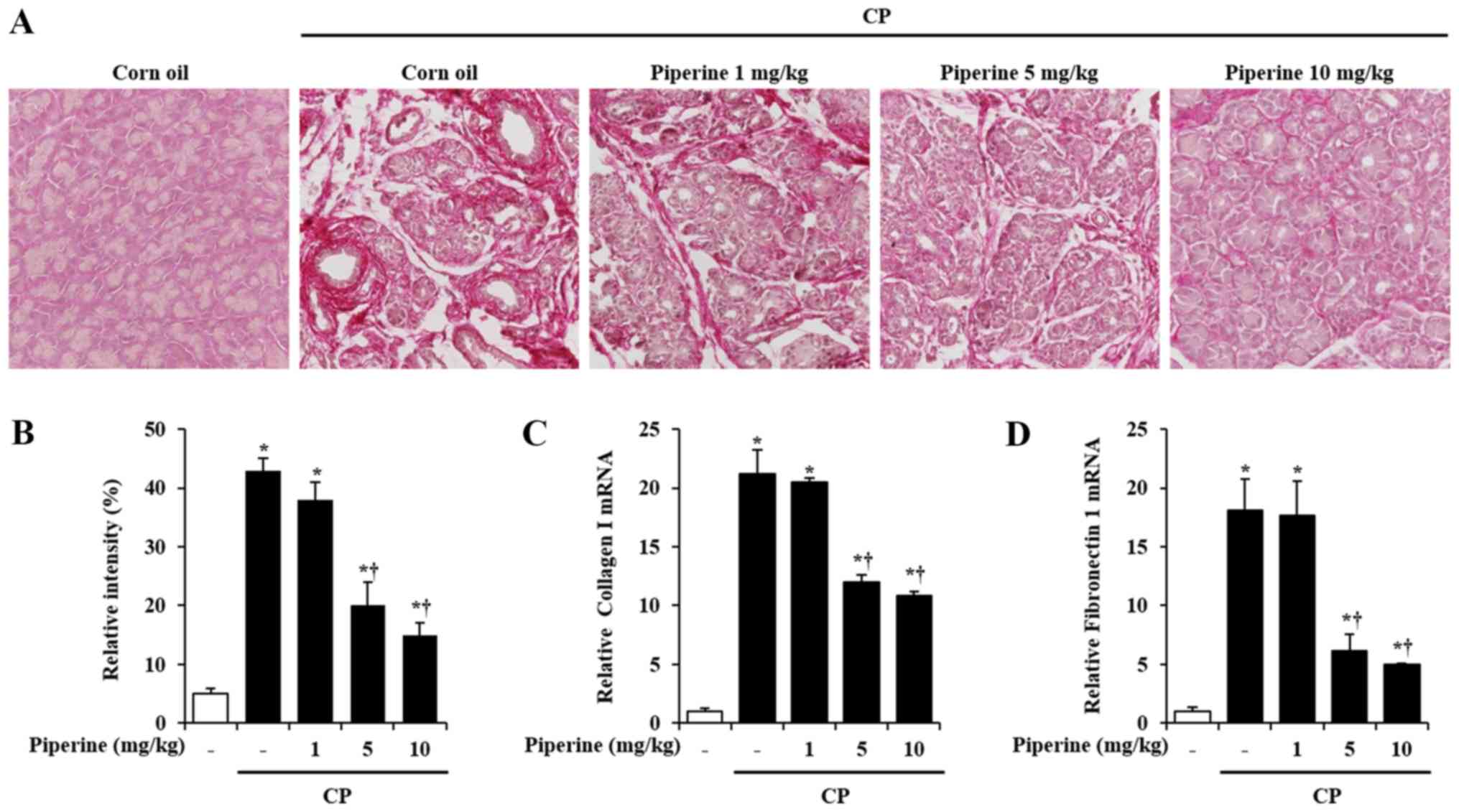
Effect of piperine on ECM production
during CP
ECM deposition such as collagen and fibronectin is
an important manifestation of CP. To determine the degree of
collagen deposition following piperine treatment, Sirius Red
staining was performed. As shown in Fig. 3A, CP-induced mice showed a marked
increase in collagen deposition in the pancreas. However, a
significant decrease in collagen deposition was observed in the
piperine-treated mice (Fig. 3A and
B). Furthermore, the mRNA levels of collagen I and fibronectin
1 were also reduced following treatment with piperine (Fig. 3C and D).
Effect of piperine on cytokine and
chemokine production during CP
It has been reported that TGF-β contributes to
chronic pancreatic injury and fibrosis (22). Therefore, the expression of TGF-β
was measured in pancreases of the CP-induced mice. Pancreatic TGF-β
was markedly increased by CP as previously reported (22); however, piperine treatment
significantly inhibited the TGF-β expression (Fig. 4A and B). The level of TGF-β mRNA
was also inhibited by piperine while corn oil-treated mice with CP
showed higher levels of TGF-β mRNA (Fig. 4C). In addition, cytokines such as
TNF-α, IL-1β, and IL-6 and chemokines such as CCL2 and CXCL2 are
important candidate factors that may activate PSCs to promote CP
(22,23). Thus, we investigated whether
administration of piperine is able to decrease the cytokine and
chemokine levels during CP. The results of this experiment
(Fig. 4D and E) demonstrated that
the treatment of piperine decreased the levels of cytokines such as
TNF-α, IL-1β, and IL-6 mRNA and chemokines such as CCL2 and CXCL2
mRNA (Fig. 4D and E).
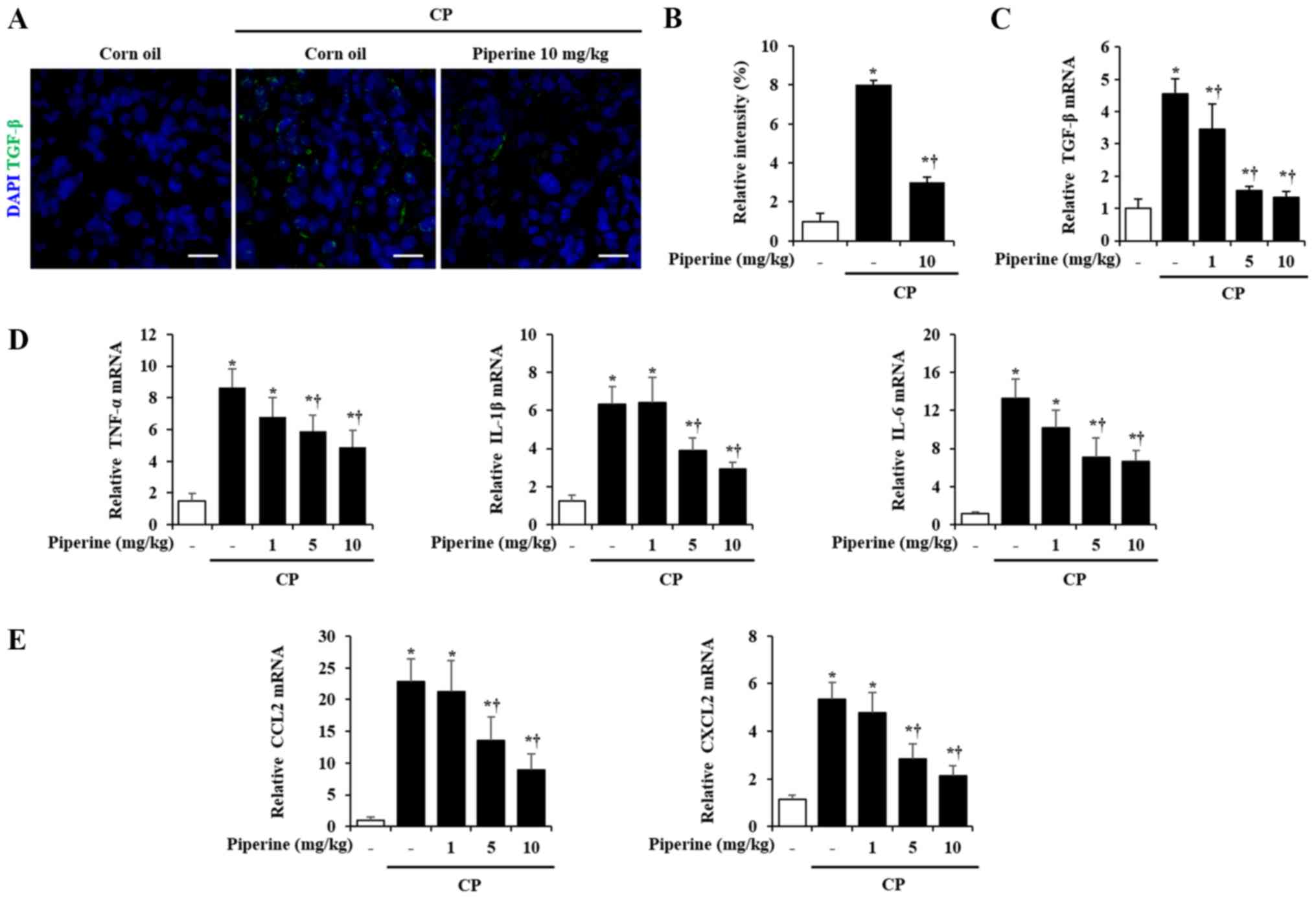 | Figure 4.Effect of piperine on cytokines
(TGF-β, TNF-α, IL-1β and IL-6 mRNA) and chemokines (CCL2 and CXCL2
mRNA) production during CP. (A) Confocal images of
immunofluorescence staining of TGF-β (green) and DAPI (blue). (B)
Relative intensity of TGF-β. (C) TGF-β mRNA was determined using
RT-qPCR. (C) TGF-β, (D) TNF-α, IL-1β and IL-6, (E) CCL2 and CXCL2
mRNA was determined using RT-qPCR. Data are represented as means ±
SEM for 6 mice in each group. Results are representative of three
experiments. *P<0.05 vs. corn oil alone; †P<0.05
vs. CP. Scale bar, 10 µm. CCL, C-C motif chemokine ligand; CXCL,
C-X-C motif chemokine ligand; CP, chronic pancreatitis; IL,
interleukin; TGF-β, transforming growth factor-β; TNF, tumor
necrosis factor; DAPI, 4′,6-diamidino-2-phenylindole. |
Effect of piperine on TGF-β-induced
PSC activation in the isolated PSCs
To evaluate the detailed beneficial effects of
piperine against CP, mouse PSCs, the major cells which regulate CP,
were isolated. As previous studies have shown that TGF-β promotes
PSC activation, and stimulates ECM synthesis (24,25),
we examined the effect of piperine on the TGF-β-induced expression
of fibrosis-related genes such as α-SMA, fibronectin 1 and collagen
I/III in isolated PSCs. Using real-time PCR analysis, we found
that, compared with the control group, mRNA levels of α-SMA,
fibronectin 1 and collagen I/III in the TGF-β treatment group were
notably increased. However, the increased mRNA expression by TGF-β
was significantly decreased in the presence of piperine (Fig. 5A-D). Likewise, protein levels of
α-SMA and collagen I were also decreased by piperine treatment
(Fig. 5E).
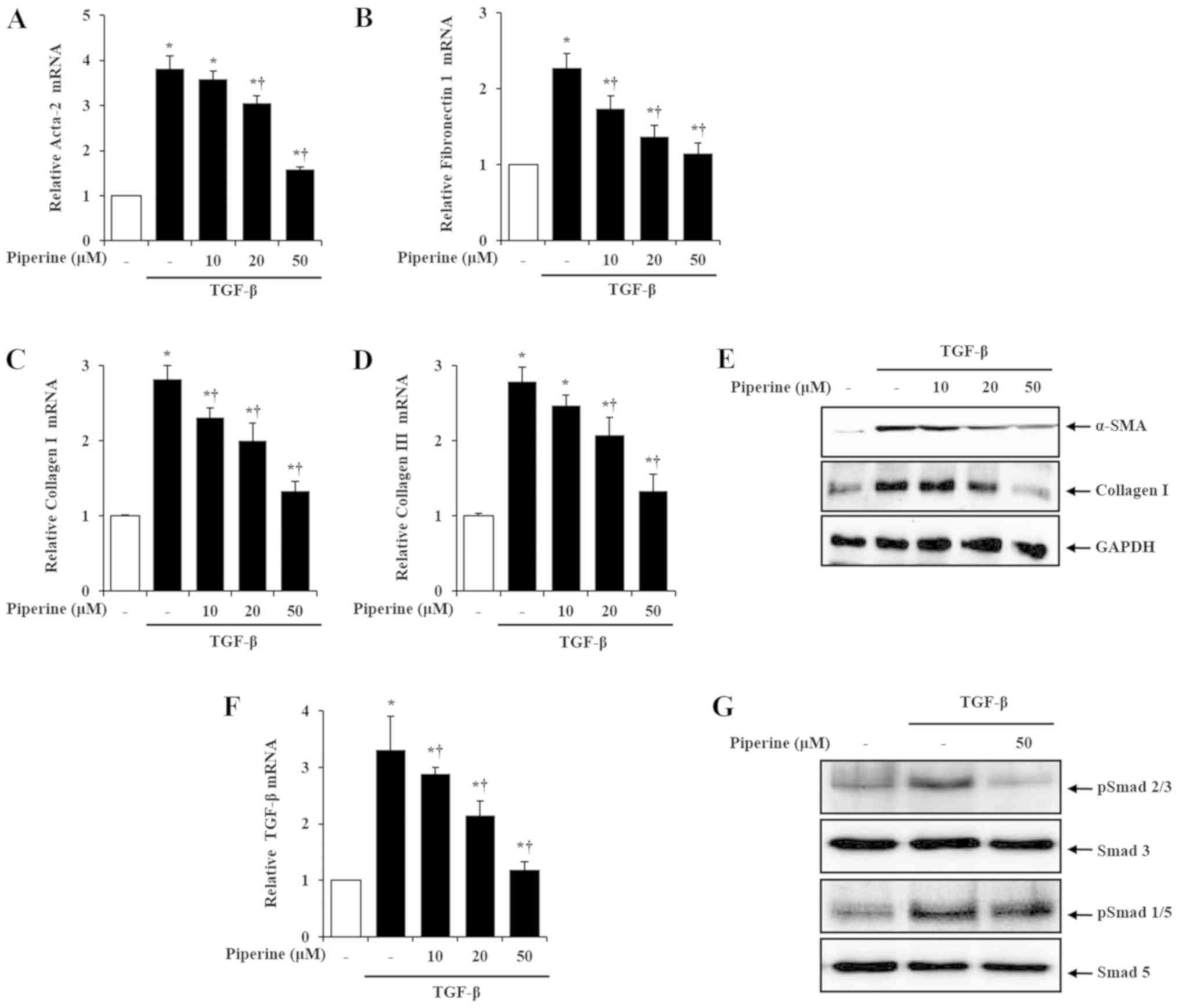 | Figure 5.Effect of piperine on TGF-β-induced
PSC activation and ECM production on the isolated PSCs. Mouse PSCs
were pretreated with piperine at various concentrations (10, 20, 50
µM) for 1 h, and then stimulated with TGF-β (0.5 ng/ml) for 24 h.
Levels of mRNA were quantified by RT-PCR for (A) α-SMA, (B)
fibronectin 1, (C) collagen I, (D) collagen III, (F) TGF-β. (E)
Protein levels of α-SMA and collagen I were assessed using western
blot analysis and GAPDH was used as a loading control. Mouse PSCs
were pretreated with piperine (50 µM) for 1 h followed by TGF-β
(0.5 ng/ml) for 30 min, and then whole cell lysates were harvested.
(G) The phosphorylation (p) levels of SMAD2/3 and SMAD 1/5 were
analyzed by western blot analysis and SMAD3 and SMAD5 were used as
loading controls. Data are represented as means ± SEM for 6 mice in
each group. Results are representative of three experiments.
*P<0.05 vs. saline alone; †P<0.05 vs. TGF-β.
TGF-β, transforming growth factor-β; PSCs, pancreatic stellate
cells; ECM, extracellular matrix; CP, chronic pancreatitis; α-SMA,
α-smooth muscle actin. |
TGF-β intracellular signaling is mediated and
modulated primarily by SMADs (26,27).
Thus, to ascertain the mechanisms responsible for the inhibition of
PSC activation by piperine, we firstly examined the expression of
TGF-β and SMAD. In accordance with previous reports, TGF-β and
SMAD2/3 and SMAD1/5 were increased in the activated PSCs. However,
in the presence of piperine, TGF-β expression and phosphorylation
of SMAD2/3 were significantly inhibited, while phosphorylation of
SMAD1/5 was not altered (Fig. 5F and
G).
Therapeutic effects of piperine during
CP
To examine the therapeutic effects of piperine on
CP, we administered piperine orally at 1 week or 2 week after the
onset of CP (Fig. 6A). Thus,
piperine was administered for 2 weeks (starting at 1 week after the
onset of CP) or 1 week (starting at 2 week after the onset of CP)
during CP. Then, the mice were sacrificed at 3 weeks after the
beginning of the first cerulein injection. Injury of the pancreas
such as glandular atrophy and inflammatory cell infiltration was
significantly attenuated by post-treatment of piperine during CP
(Fig. 6B and C). Furthermore, the
elevation of α-SMA, collagen deposition, fibronectin 1 and TGF-β
was inhibited by post-treatment of piperine during CP (Fig. 6D-K).
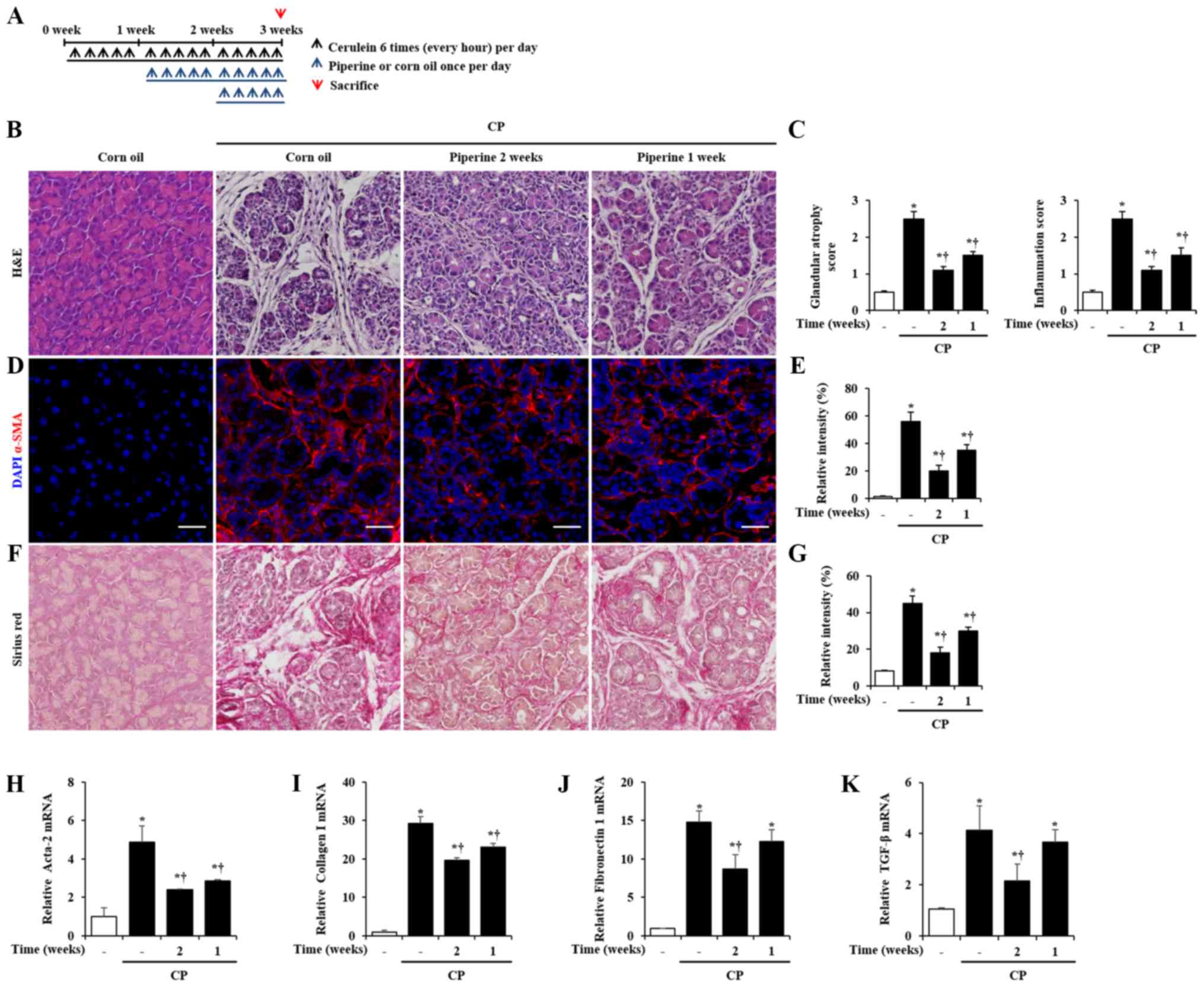 | Figure 6.Therapeutic effects of piperine on
cerulein-induced CP in mice. (A) CP was induced by the
administration of six intraperitoneal injections of cerulein (50
µg/kg) at 1-h intervals, 5 days per week, for a total of 3 weeks.
Piperine or corn oil was administrated orally 1 or 2 week after the
onset of the first cerulein injection, once a day, 5 days per week,
for a total of 2 or 1 week. Groups of mice were euthanized 3 weeks
after the beginning of the cerulein treatment. (B) Representative
H&E-stained pancreatic tissue sections. (C) Histological
sections of the pancreas were scored from 0 (normal) to 3 (severe)
for glandular atrophy and inflammation. (D) Immunofluorescence
staining of α-SMA (red) and DAPI (blue). (E) Relative intensity of
α-SMA. (F) Sirius red staining. (G) Relative intensity of Sirius
Red. The pancreatic mRNA levels of (H) Acta-2, (I) collagen I, (J)
fibronectin 1 and (K) TGF-β were determined using RT-qPCR. Data are
represented as means ± SEM for 6 mice in each group. Results are
representative of three experiments. *P<0.05 vs. corn oil alone;
†P<0.05 vs. CP. Original magnification, ×200. Scale
bar, 20 µm. CP, chronic pancreatitis; H&E, hematoxylin and
eosin; α-SMA, α-smooth muscle actin; TGF-β, transforming growth
factor-β; DAPI, 4′,6-diamidino-2-phenylindol. |
Discussion
Chronic pancreatitis (CP) is an irreversible process
that occurs through the activation of inflammatory cascades and
pancreatic fibrosis. According to the recurrent acute pancreatitis
(RAP) hypothesis, CP is the result of repeated episodes of AP. This
cascade of events is initiated by acinar cell injury. Patient can
recover from singular AP events. However, recurrent events amplify
the degree of injury and inflammation, interrupt the repair
process, and promote the progression of CP. In our previous report
(17), piperine, a natural
alkaloid found in black pepper, reduced the severity of AP as
determined by the reduction in pancreatic injury, acinar cell
death, neutrophil infiltration, digestive enzyme levels and
cytokine production. Therefore, we examined the effect of piperine
on the severity of CP using an RAP mouse model. In the present
study, it was shown that the administration of piperine reduced the
severity of CP as well as inflammation and fibrosis by inhibiting
TGF-β/SMAD signaling.
Piperine is a well-known phenolic compound that has
various biological functions. Various studies have shown that
piperine plays a role in inflammatory diseases such as neuronal
diseases (28), arthritis
(29), cardiovascular diseases
(30), gastrointestinal disorders
(31), sepsis (18) and acute pancreatitis (17). In a recent report (32), a high-carbohydrate, high-fat diet
supplemented with piperine reduced fibrosis in rat livers. In
addition, another study investigated the antioxidant effect of
curcumin with piperine in tropical pancreatitis (33). However, it remains unclear as to
whether piperine exerts preventive effects on inflammation and
fibrosis in the pancreas during CP. In our previous report
(17), there were no significant
differential effects between the oral and intraperitoneal
administration of piperine on the severity of acute pancreatitis.
Therefore, in the present study, we used intraperitoneal injections
of piperine. The pancreatic histological appearance of the
piperine-treated group was comparable with that of the
saline-treated group (data not shown). The 3 weeks of repeated
cerulein administrations were sufficient to induce CP in mice which
was proven by the presence of inflammation, pancreatic atrophy and
fibrosis (Fig. 1). The
administration of piperine inhibited inflammatory cell infiltration
and glandular atrophy, and increased the number of amylase-positive
cells in the pancreases of CP-induced mice (Fig. S1), suggesting that the inhibition
of inflammation and acinar cell death by piperine may improve
exocrine function.
Repetitive administration of cerulein aggravates
acinar cell injury, which ultimately results in pancreatic fibrosis
(34). During pancreatic fibrosis,
PSCs become activated by extra stimuli, and the activated PSCs
(α-SMA-positive cells) promote pancreatic fibrosis by producing and
secreting extracellular matrix (ECM) components such as collagen
and fibronectin (35–37). Thus, regulating the activation of
PSCs and ECM production from activated PSCs are crucial to treat
CP. In our present study, we confirmed the activation of PSCs by
α-SMA and ECM deposition such as collagen and fibronectin. While
corn oil-treated mice with CP showed a significant increase in
α-SMA-positive cells, piperine-treated mice with CP exhibited a
lower number of α-SMA positive cells and decreased ECM production
(Figs. 2 and 3). These results suggest that piperine
attenuates the severity of pancreatic fibrosis by inhibition of the
generation and activation of PSCs, and deactivation of PSCs by
piperine may lead to inhibition of exudates such as ECM from
activated PSCs.
TGF-β is a multipotent cytokine and plays an
integral role in regulating immune, inflammatory response, cell
growth and differentiation (38).
An excess of TGF-β in repeatedly injured sites is thus associated
with unresolved inflammation and fibrosis development. Previous
reports have previously shown that excessive TGF-β activity is the
cause of increased scarring and fibrosis in diseases including
hepatic cirrhosis (39) and
cardiac fibrosis (40). TGF-β may
be involved in pancreatic fibrogenesis, which is released after
cell injury and may be one of the pro-fibrogenic factors that
promote fibrotic responses in PSCs (34,36).
PSCs are activated upon exposure to cytokines, such as TGF-β, and
then α-SMA is expressed, fibronectin 1 levels increase, and
collagen is deposited near the fibrotic pancreatic tissues
(41). Thus, in our experiments,
we examined TGF-β as a stimulator of PSCs during CP. In the present
study, the mice with CP exhibited increased levels of TGF-β in
pancreatic tissue (Fig. 4A-C).
When mice were treated with piperine during CP, TGF-β production
was reduced in the mouse pancreas (Fig. 4A-C). These results suggest that
piperine may negatively regulate PSC activation by inhibition of
TGF-β production. Based on these in vivo findings, we also
directly examined the effect of piperine on TGF-β-induced PSC
activation and ECM production from PSCs. It was demonstrated that
piperine inhibited the expression of α-SMA, fibronectin 1 and
collagen I/III upon TGF-β treatment, which suggests that piperine
plays a key role in regulating fibrosis stimulated by TGF-β, as
well as in vivo data (Fig.
5A-E). We also aimed to examine the anti-fibrotic role of
piperine on human PSCs to provide clinical value. Yet, it was not
possible to obtain the human samples and permission for such a
clinical study from our institution. Although a study using human
PSCs was not carried out at this time, attainment of human samples
will be achieved in future studies.
In the TGF-β-mediated signaling pathway, SMAD
proteins are phosphorylated and activated by receptors and are
translocated into the nucleus where these complexes regulate
transcription of pro-fibrotic genes (42,43).
Among them, the TGF-β/SMAD2/3 signaling pathway is reported to play
a major role in the activation of PSCs. Studies have revealed that
TGF-β stimulates PSC activation in a SMAD2/3-dependent manner
(44). A previous study
demonstrated the ability to inhibit pancreatic fibrosis by blocking
the TGF-β/SMAD2 signaling pathway (45). On the basis of the above-mentioned
evidence, we investigated the effects of piperine on the expression
of signaling in the TGF-β/SMAD pathway. It was found that piperine
treatment inhibited pSMAD2/3, but not pSMAD1/5 in PSCs (Fig. 5F and G). These data suggest that
piperine reduces the expression of SMAD2/3 thereby downregulating
the expression of PSC activation and ECM production. In addition,
these results suggest that piperine exhibits its beneficial effects
on PSCs by regulating the TGF-β/SMAD pathway.
Taken together, this study demonstrated that
piperine prevents the progression of pancreatic fibrosis by
inhibiting the TGF-β/SMAD signaling pathway. Our findings suggest
that piperine has an anti-fibrotic effect against CP and may be
useful for the clinical management of pancreatic disorders.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korean government
(MEST) (grant nos. NRF-2017R1C1B2010031, NRF-2017R1A5A2015805 and
NRF-2019R1A2C2008814).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JWC, SKL, GSB and SJP made substantial contributions
to the conception and design of the study; JWC, SKL, MJK, DGK, JYS,
ZQZ, HJS and IJJ were involved in data analysis and interpretation;
JWC, SKL, HJS, GSB and SJP drafted the manuscript and revised it
critically for important intellectual content. All authors read and
approved the final manuscript, and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study was approved by the Animal Care
Committee of Wonkwang University (WKU15-18). All participants
provided informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
α-SMA
|
α-smooth muscle actin
|
|
BSA
|
bovine serum albumin
|
|
CCL
|
C-C motif chemokine ligand
|
|
CXCL
|
C-X-C motif chemokine ligand
|
|
CP
|
chronic pancreatitis
|
|
ECM
|
extracellular matrix
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
|
HPRT
|
hypoxanthine-guanine
phosphoribosyltransferase
|
|
H&E
|
hematoxylin and eosin
|
|
HRP
|
horseradish peroxidase
|
|
ICD
|
International Classification of
Disease
|
|
IHC
|
immunohistochemistry
|
|
IL
|
interleukin
|
|
PBST
|
phosphate-buffered saline/Tween 20
|
|
PSCs
|
pancreatic stellate cells
|
|
RAP
|
recurrent acute pancreatitis
|
|
RT
|
room temperature
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
SDS
|
sodium dodecyl sulfate
|
|
TGF-β
|
transforming growth factor-β
|
|
TNF
|
tumor necrosis factor
|
References
|
1
|
Apte M, Pirola R and Wilson J: The
fibrosis of chronic pancreatitis: New insights into the role of
pancreatic stellate cells. Antioxid Redox Signal. 15:2711–2722.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hobbs PM, Johnson WG and Graham DY:
Management of pain in chronic pancreatitis with emphasis on
exogenous pancreatic enzymes. World J Gastrointest Pharmacol Ther.
7:370–386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang AL, Vadhavkar S, Singh G and Omary
MB: Epidemiology of alcohol-related liver and pancreatic disease in
the United States. Arch Intern Med. 168:649–656. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Robles-Diaz G, Vargas F, Uscanga L and
Fernández-del Castillo C: Chronic pancreatitis in Mexico City.
Pancreas. 5:479–483. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hirota M, Shimosegawa T, Masamune A,
Kikuta K, Kume K, Hamada S, Kihara Y, Satoh A, Kimura K, Tsuji I,
et al: The sixth nationwide epidemiological survey of chronic
pancreatitis in Japan. Pancreatology. 12:79–84. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miyake H, Harada H, Ochi K, Kunichika K,
Tanaka J and Kimura I: Prognosis and prognostic factors in chronic
pancreatitis. Dig Dis Sci. 34:449–455. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ammann RW, Akovbiantz A, Largiader F and
Schueler G: Course and outcome of chronic pancreatitis.
Longitudinal study of a mixed medical-surgical series of 245
patients. Gastroenterology. 86:820–828. 1984.PubMed/NCBI
|
|
8
|
Lowenfels AB, Maisonneuve P, Cavallini G,
Ammann RW, Lankisch PG, Andersen JR, Dimagno EP, Andrén-Sandberg A
and Domellöf L: Pancreatitis and the risk of pancreatic cancer.
International pancreatitis study group. N Engl J Med.
328:1433–1437. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Haber PS, Keogh GW, Apte MV, Moran CS,
Stewart NL, Crawford DH, Pirola RC, McCaughan GW, Ramm GA and
Wilson JS: Activation of pancreatic stellate cells in human and
experimental pancreatic fibrosis. Am J Pathol. 155:1087–1095. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aghdassi AA, Mayerle J, Christochowitz S,
Weiss FU, Sendler M and Lerch MM: Animal models for investigating
chronic pancreatitis. Fibrogenesis Tissue Repair. 4:262011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Turner R: Acute pancreatitis is a chronic
disease. Pancreatic Dis Ther. 3:1182013. View Article : Google Scholar
|
|
12
|
Wehr AY, Furth EE, Sangar V, Blair IA and
Yu KH: Analysis of the human pancreatic stellate cell secreted
proteome. Pancreas. 40:557–566. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schneider A, Löhr JM and Singer MV: The
M-ANNHEIM classification of chronic pancreatitis: Introduction of a
unifying classification system based on a review of previous
classifications of the disease. J Gastroenterol. 42:101–119. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lankisch PG, Löhr-Happe A, Otto J and
Creutzfeldt W: Natural course in chronic pancreatitis. Pain,
exocrine and endocrine pancreatic insufficiency and prognosis of
the disease. Digestion. 54:148–155. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Singh YN: Kava: An overview. J
Ethnopharmacol. 37:13–45. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Srinivasan K: Black pepper and its pungent
principle-piperine: A review of diverse physiological effects. Crit
Rev Food Sci Nutr. 47:735–748. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bae GS, Kim MS, Jeong J, Lee HY, Park KC,
Koo BS, Kim BJ, Kim TH, Lee SH, Hwang SY, et al: Piperine
ameliorates the severity of cerulein-induced acute pancreatitis by
inhibiting the activation of mitogen activated protein kinases.
Biochem Biophys Res Commun. 410:382–390. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bae GS, Kim MS, Jung WS, Seo SW, Yun SW,
Kim SG, Park RK, Kim EC, Song HJ and Park SJ: Inhibition of
lipopolysaccharide-induced inflammatory responses by piperine. Eur
J Pharmacol. 642:154–162. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Van Laethem JL, Robberecht P, Résibois A
and Deviére J: Transforming growth factor beta promotes development
of fibrosis after repeated courses of acute pancreatitis in mice.
Gastroenterology. 110:576–582. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Masamune A, Watanabe T, Kikuta K and
Shimosegawa T: Roles of pancreatic stellate cells in pancreatic
inflammation and fibrosis. Clin Gastroenterol Hepatol 7 (11 Suppl).
S48–S54. 2009. View Article : Google Scholar
|
|
22
|
Vinokurova LV, Zhivaeva NS,
Tsaregorodtseva TM and Serova TI: A cytokine status in chronic
alcoholic and biliary pancreatitis. Ter Arkh. 78:57–60. 2006.(In
Russian). PubMed/NCBI
|
|
23
|
Zhang H, Neuhöfer P, Song L, Rabe B,
Lesina M, Kurkowski MU, Treiber M, Wartmann T, Regnér S, Thorlacius
H, et al: IL-6 trans-signaling promotes pancreatitis-associated
lung injury and lethality. J Clin Invest. 123:1019–1031. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shek FW, Benyon RC, Walker FM, McCrudden
PR, Pender SL, Williams EJ, Johnson PA, Johnson CD, Bateman AC,
Fine DR and Iredale JP: Expression of transforming growth
factor-beta 1 by pancreatic stellate cells and its implications for
matrix secretion and turnover in chronic pancreatitis. Am J Pathol.
160:1787–1798. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vogelmann R, Ruf D, Wagner M, Adler G and
Menke A: Effects of fibrogenic mediators on the development of
pancreatic fibrosis in a TGF-beta1 transgenic mouse model. Am J
Physiol Gastrointest Liver Physiol. 280:G164–G172. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gressner AM, Weiskirchen R, Breitkopf K
and Dooley S: Roles of TGF-beta in hepatic fibrosis. Front Biosci.
7:d793–d807. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chung H, Ramachandran R, Hollenberg MD and
Muruve DA: Proteinase-activated receptor-2 transactivation of
epidermal growth factor receptor and transforming growth factor-β
receptor signaling pathways contributes to renal fibrosis. J Biol
Chem. 288:37319–37331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fu M, Sun ZH and Zuo HC: Neuroprotective
effect of piperine on primarily cultured hippocampal neurons. Biol
Pharm Bull. 33:598–603. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bang JS, Oh DH, Choi HM, Sur BJ, Lim SJ,
Kim JY, Yang HI, Yoo MC, Hahm DH and Kim KS: Anti-inflammatory and
antiarthritic effects of piperine in human interleukin
1beta-stimulated fibroblast-like synoviocytes and in rat arthritis
models. Arthritis Res Ther. 11:R492009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Taqvi SI, Shah AJ and Gilani AH: Blood
pressure lowering and vasomodulator effects of piperine. J
Cardiovasc Pharmacol. 52:452–458. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mehmood MH and Gilani AH: Pharmacological
basis for the medicinal use of black pepper and piperine in
gastrointestinal disorders. J Med Food. 13:1086–1096. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Diwan V, Poudyal H and Brown L: Piperine
attenuates cardiovascular, liver and metabolic changes in high
carbohydrate, high fat-fed rats. Cell Biochem Biophys. 67:297–304.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Durgaprasad S, Pai CG, Vasanthkuma r,
Alvres JF and Namitha S: A pilot study of the antioxidant effect of
curcumin in tropical pancreatitis. Indian J Med Res. 122:315–318.
2005.PubMed/NCBI
|
|
34
|
Schneider E, Schmid-Kotsas A, Zhao J,
Weidenbach H, Schmid RM, Menke A, Adler G, Waltenberger J, Grünert
A and Bachem MG: Identification of mediators stimulating
proliferation and matrix synthesis of rat pancreatic stellate
cells. Am J Physiol Cell Physiol. 281:C532–C543. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Omary MB, Lugea A, Lowe AW and Pandol SJ:
The pancreatic stellate cell: A star on the rise in pancreatic
diseases. J Clin Invest. 117:50–59. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Apte MV, Pirola RC and Wilson JS:
Pancreatic stellate cells: A starring role in normal and diseased
pancreas. Front Physiol. 3:3442012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen B, Li J, Fellows GF, Sun Z and Wang
R: Maintaining human fetal pancreatic stellate cell function and
proliferation require β1 integrin and collagen I matrix
interactions. Oncotarget. 6:14045–14059. 2015.PubMed/NCBI
|
|
38
|
Prud'homme GJ: Pathobiology of
transforming growth factor beta in cancer, fibrosis and immunologic
disease, and therapeutic considerations. Lab Invest. 87:1077–1091.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Czaja MJ, Weiner FR, Flanders KC,
Giambrone MA, Wind R, Biempica L and Zern MA: In vitro and in vivo
association of transforming growth factor-beta 1 with hepatic
fibrosis. J Cell Biol. 108:2477–2482. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ma ZG, Yuan YP, Zhang X, Xu SC, Wang SS
and Tang QZ: Piperine attenuates pathological cardiac fibrosis via
PPAR-γ/AKT pathways. EBioMedicine. 18:179–187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang L, Shen J, He S, Hu G, Shen J, Wang
F, Xu L, Dai W, Xiong J, Ni J, et al: L-cysteine administration
attenuates pancreatic fibrosis induced by TNBS in rats by
inhibiting the activation of pancreatic stellate cell. PLoS One.
7:e318072012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fabregat I, Moreno-Càceres J, Sánchez A,
Dooley S, Dewidar B, Giannelli G and Ten Dijke P; IT-LIVER
Consortium, : TGF-β signaling and liver disease. FEBS J.
283:2219–2232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wynn TA: Cellular and molecular mechanisms
of fibrosis. J Pathol. 214:199–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ohnishi H, Miyata T, Yasuda H, Satoh Y,
Hanatsuka K, Kita H, Ohashi A, Tamada K, Makita N, Iiri T, et al:
Distinct roles of Smad2-, Smad3-, and ERK-dependent pathways in
transforming growth factor-beta1 regulation of pancreatic stellate
cellular functions. J Biol Chem. 279:8873–8878. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xu L, Zheng N, He Q, Li R, Zhang K and
Liang T: Puerarin, isolated from Pueraria lobata (Willd.),
protects against hepatotoxicity via specific inhibition of the
TGF-β1/Smad signaling pathway, thereby leading to anti-fibrotic
effect. Phytomedicine. 20:1172–1179. 2013. View Article : Google Scholar : PubMed/NCBI
|