Introduction
Cerebral infarction is the most common type of
ischemic cerebrovascular disease, accounting for approximately 80%
of all stroke cases (1,2). It is associated with high rates of
morbidity, disability, and mortality. Following cerebral
infarction, the blood flow to the brain is interrupted, and the
intake of oxygen and glucose in the ischemic area is hindered
(3,4). Therefore, timely recovery of blood
supply to the brain tissue is crucial to reduce subsequent neuronal
death. However, one study also revealed that reperfusion may lead
to cerebral ischemia-reperfusion injury (CIRI) (5). The pathological mechanisms of CIRI
involve inflammatory response, excitatory toxicity, calcium
overloading, production of free radicals, mitochondrial
dysfunction, and apoptosis (6–9).
Therefore, the reduction of apoptosis is an important step in
improving cerebral infarction (10). In recent years, there has been
increasing research on the endoplasmic reticulum stress
(ERS)-dependent apoptotic pathway, as an intervention for the
reduction of I/R injury in cerebral infarction (11).
Thymosin β4 (Tβ4) is a polypeptide consisting of 43
amino acid residues, and belongs to the thymosin β family (12). Recent studies have confirmed that
Tβ4 has various biological functions, such as promoting stem cell
differentiation and angiogenesis, and enhancing cell proliferation,
migration, and anti-apoptosis. Moreover, it is closely related to
tissue regeneration, angiogenesis, and wound healing (13–15).
Several studies have revealed that Tβ4 exerts a neuroprotective
effect. For example, Tβ4 can reduce the death of motor neurons
caused by staurosporine (16),
reduce the damage caused by excitatory amino acids to cortical
neurons, and improve symptoms of nerve injury in rats with cerebral
ischemia (17). However, the
specific neuroprotective mechanism of Tβ4 remains unclear. In the
present study, a rat model of focal cerebral ischemia and
reperfusion was established to evaluate the neuronal protection of
thymosin β4. In conclusion, the present study provides an
experimental basis for the clinical treatment of ischemic
cerebrovascular disease using Tβ4.
Materials and methods
Reagents
Recombinant Tβ4 was purchased from Cloud-Clone Corp.
The terminal dexynucleotidyl transferase (TdT)-mediated dUTP nick
end labeling (TUNEL) apoptosis detection kit was obtained from
Beyotime Institute of Biotechnology. Rabbit anti-glucose-regulated
protein 78 (GRP78), rabbit anti-caspase-12 (CASP12), and rabbit
anti-C/EBP homologous protein (CHOP) monoclonal antibodies were
obtained from Abclonal Biotech Co., Ltd.
Experimental animals
In total, 48 male Sprague-Dawley rats (clean grade)
(male; body weight: 250–300 g; age: 7–8 weeks) were used to
establish the focal cerebral I/R model. All animals were provided
by the Experimental Animal Co., Ltd., and housed in a specific
pathogen-free environment that was automatically maintained at a
temperature of 23±2°C, a relative humidity of 45–65%, and with a
controlled 12 h light/dark cycle. The animals had free to access
food and water. All animal procedures were approved by the Ethics
Committee of Guangzhou Medical University.
According to the random number table method, 48
Sprague-Dawley rats were divided into three groups (n=16 per
group): A sham operation group (sham group), an
ischemia/reperfusion group (I/R group), and a Tβ4 group. In the
sham group, the internal carotid artery was isolated but not tied,
and the incision was subsequently sutured. Rats in the I/R and Tβ4
groups underwent a 2 h occlusion of the right middle cerebral
artery (MCAO). Additionally, rats in the Tβ4 group were
intraperitoneally injected with Tβ4 (8 mg/kg) 1 h after MCAO,
whereas rats in the I/R group were intraperitoneally injected with
normal saline.
Preparation of the cerebral I/R
model
The Zea-Longa method was used to establish the rat
MCAO-I/R model (18). All rats
were anesthetized (4% isoflurane for induction and 1% for
maintenance) and disinfected. An incision was performed in the
middle of the neck to expose the external carotid, internal
carotid, and common carotid arteries. The right MCA was blocked
according to the method described by Longa (18). The length of the line inserted into
the internal carotid artery was ~18 mm. The muscle and skin were
sutured layer by layer, and the plug was removed to initiate
reperfusion for 2 h after ischemia.
Neurological evaluation
Approximately 1.5 h after anesthesia, the rats had
regained consciousness, and neurological scores were evaluated
according to the Longa method as follows: 0 points, no symptoms of
neurological deficits; 1 point, unable to fully extend the
contralateral forepaw; 2 points, circle to the left (paralyzed)
side when walking; 3 points, fall to the left (paralyzed) side when
walking; and 4 points, unable to walk on their own, losing
consciousness. Higher scores indicated more severe neurological
impairments. Rats with a Zea-Longa score of 1–3 were included in
the subsequent experiments.
2,3,5-Triphenyltetrazolium chloride
(TTC) and hematoxylin and eosin (H&E) staining
The rats were subsequently anesthetized using
isoflurane (4% isoflurane) and decapitated 3 days after the
surgery. The brain tissue was washed with saline, and subsequently
frozen at −20°C for 20 min. Coronal slices (thickness, 2 mm) were
collected, immediately placed in phosphate-buffered saline
containing 2% TTC, and incubated for 20 min at 37°C in the dark.
The brain slices were analyzed using the ImageJ software version
1.48 (National Institutes of Health), and the percentage of infarct
volume to total volume was calculated. The infarct volume was
calculated based upon the formula: Infarct volume (%)=ischemic
area/whole area of brain ×100, using the ImageJ software as
previously described (19). Brain
sections were also used for histopathological analysis through
H&E staining.
Assessment of apoptosis
The hippocampus was isolated on ice and fixed in 4%
paraformaldehyde at 4°C overnight and embedded in paraffin. The 3
µm sections were continuously sliced. Following dewaxing by xylene,
the tissues were dehydrated in 70, 75, 80, 85 and 95% alcohol. To
retrieve the antigen, 3% hydrogen peroxide was applied at 100°C.
The TUNEL assay was used to detect apoptosis in the hippocampus
according to the instructions provided by the manufacturer (cat.
no. C1088; Beyotime Institute of Biotechnology). Five different
fields were randomly selected from each slice. The ImageJ software
version 1.48 was used to calculate TUNEL-positive cells,
4′,6-diamidino-2-phenylindole (DAPI)-positive cells, and apoptosis
in the corresponding regions was calculated based on the formula:
Apoptosis index=the number of TUNEL-positive cells/the number of
DAPI-positive cells as previously described (20).
Immunohistochemical staining
The hippocampus tissues were fixed in 4%
paraformaldehyde at 4°C overnight. After dehydration in 30%
sucrose, the hippocampi were sectioned into coronal slices.
Following blocking in goat serum (2 h at room temperature; Hyclone;
GE Healthcare Life Sciences), sections were incubated with rabbit
anti-mouse monoclonal primary antibodies against GRP78 (1:200; cat.
no. A11366; ABclonal Biotech Co., Ltd.), CASP12 (1:200; cat. no.
A0217; ABclonal Biotech Co., Ltd.) and CHOP (1:200; cat. no. A0221;
ABclonal Biotech Co., Ltd.) for 60 min at room temperature,
followed by incubation with anti-rabbit IgG horseradish
peroxidase-linked antibody (1:200; cat. no. 7074; ZB-2301,
ZSGB-BIO; OriGene Technologies, Inc.) at 37°C for 15 min. The
number of GRP78-, CHOP-, and CASP12-positive cells in the
corresponding region of the slice was calculated using the
streptavidin-peroxidase method and imaged using the Image-Pro Plus
software (National Institutes of Health). Immunohistochemical
staining was visualized with 3,3′-diaminobenzidine chromogen for 3
min at room temperature. The nucleus was counterstained by
hematoxylin at room temperature for 3 min.
Western blotting
After removal of the brain, brain tissues around the
infarcted region were removed and cut into pieces. The brain tissue
was subsequently homogenized, lysed using a protein isolation kit
(GE Healthcare Life Sciences) and centrifuged (11,000 × g at 4°C).
The bicinchoninic acid (BCA) method was used to determine the total
protein concentration. Western blotting was performed according to
standard procedures (21).
Briefly, protein samples (25 µg) obtained from each group were run
on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (12%)
and transferred onto nitrocellulose membranes for western blot
analysis. Subsequently, the membranes were blocked in 5% skim milk
for 2 h in room temperature. The membranes were subsequently
incubated with the following primary antibodies: Rabbit anti-GRP78
(1:1,000), rabbit anti-CASP12 (1:1,000), and rabbit anti-CHOP
(1:1,000) for 60 min at room temperature. The nitrocellulose
membranes were washed thrice, and incubated with secondary antibody
(HRP-labeled goat anti-rabbit IgG; cat. no. A16104; Thermo Fisher
Scientific, Inc.) at 4°C for 2 h. The staining of the blots was
enhanced using an electrochemiluminescence kit (Thermo Fisher
Scientific, Inc.). The densities of the blots were quantified using
the Quantity One software (v4.62; Bio-Rad Laboratories, Inc.).
Statistical data analysis
Data are presented as the mean and standard
deviation. One-way analysis of variance with Newman-Keuls as the
post-hoc test was performed using SPSS 17.0 (SPSS, Inc.)
statistical software to assess the differences between groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Tβ4 ameliorates the neurological
deficit caused by I/R injury
Animal health and behaviors were monitored every
day. The neurological behavior of the rats prior to the I/R injury
was normal and the Zea-Longa score was 0, thereby indicating that
the three groups exhibited similar baseline characteristics.
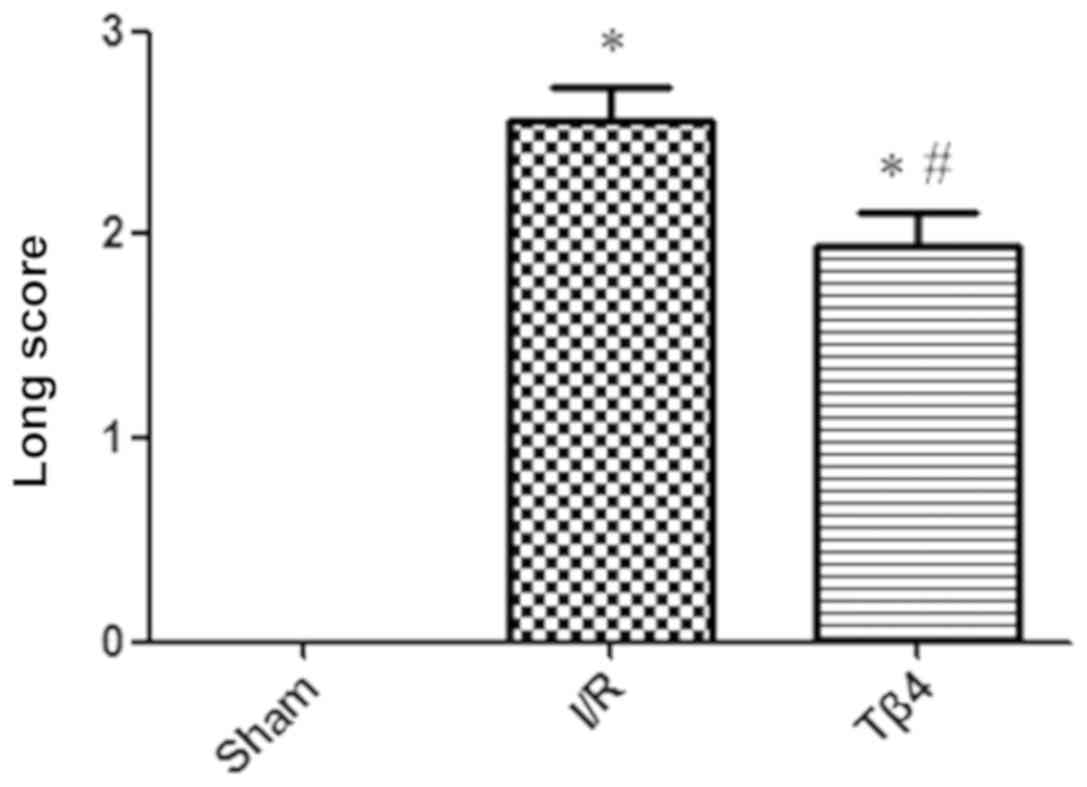
Twenty-four hours after reperfusion, the Zea-Longa score was 0 in
the sham group, 2.56±0.63 in the I/R group, and 1.94±0.68 in the
Tβ4 group. The Longa score of the I/R group was significantly
higher than that of the sham group (P<0.01), indicating that the
rat model of focal cerebral I/R was successfully established. In
contrast, the Longa score of the Tβ4 group was significantly lower
than that of the I/R group (P<0.05) (Fig. 1).
Tβ4 ameliorates cerebral infarction
caused by I/R injury
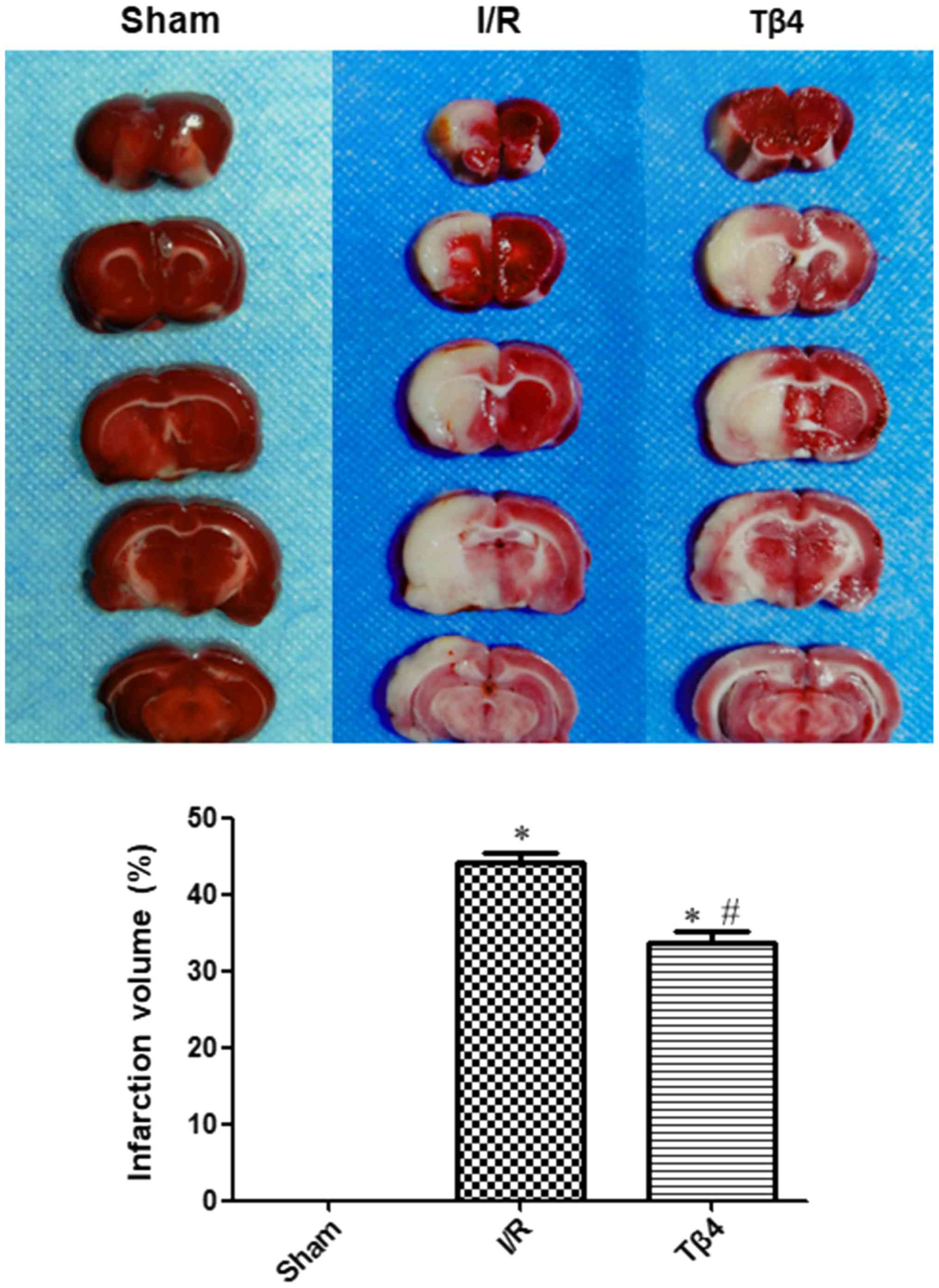
Twenty-four hours after cerebral I/R, TTC staining
was performed in brain tissues obtained from each group. In the
sham group, the brain slices were stained red and there was no
infarct observed. However, areas of white infarct regions were
observed in the I/R and Tβ4 groups, and were consistent with the
range of arterial embolization. The infarction volume in the I/R
and Tβ4 groups was significantly increased (P<0.05) compared
with that measured in the sham group. The cerebral infarction
volume was 0 in the sham group, 44.05±3.54 in the I/R group, and
33.75±3.44 in the Tβ4 group. Moreover, the volume of cerebral
infarction in the Tβ4 group was significantly lower than that
measured in the I/R group (P<0.05) (Fig. 2).
Tβ4 ameliorates the pathological
changes caused by I/R injury
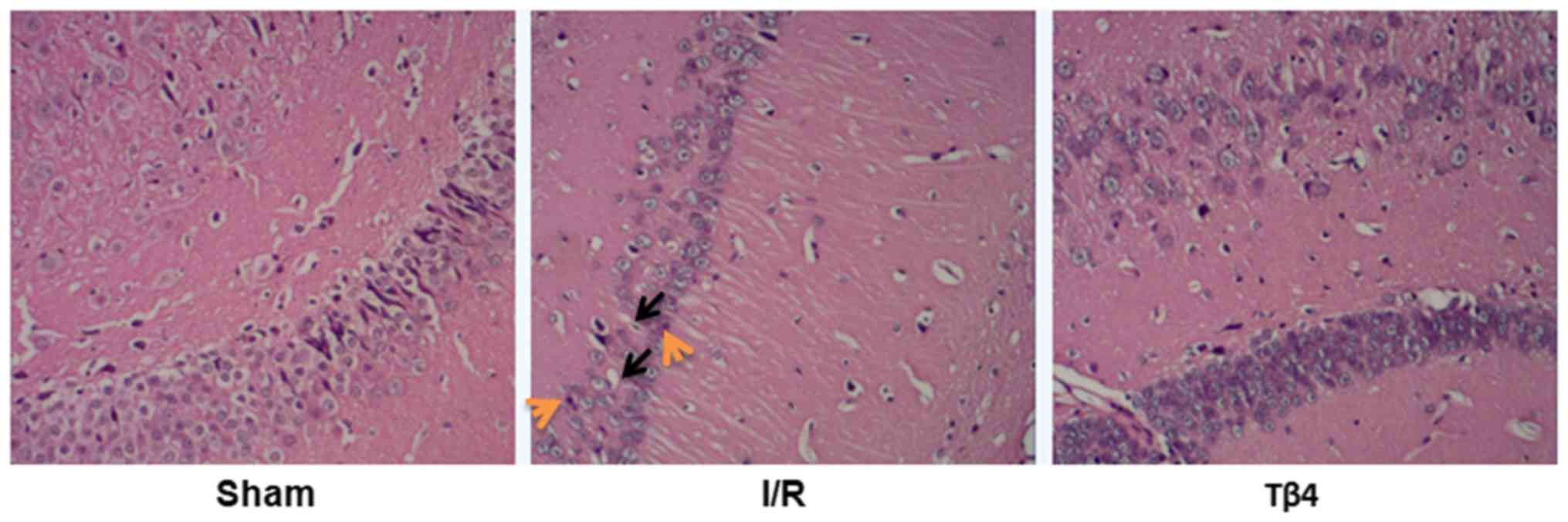
Pathological examination revealed that brain tissues
obtained from the I/R and Tβ4 groups exhibited different degrees of
damage. However, this damage was not observed in the sham group.
The pathological manifestations included interstitial edema,
vacuolation, nuclear condensation, and dissolution. The
pathological manifestations were also mild in the Tβ4 group
(Fig. 3).
Tβ4 ameliorates the neuronal apoptosis
caused by I/R injury
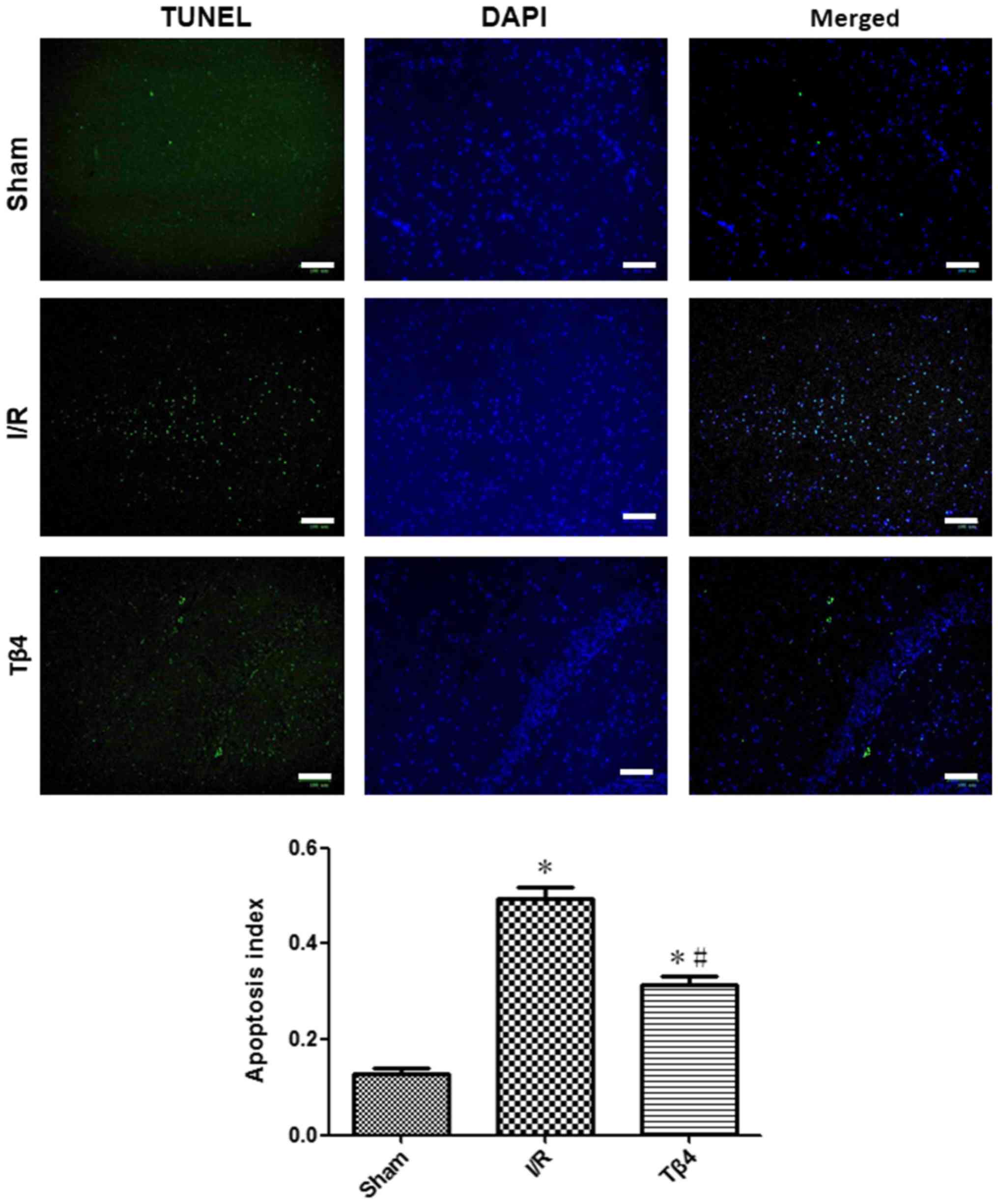
The results of the TUNEL assay revealed that the
rate of apoptosis in hippocampus was 0.13±0.03 in the sham group,
0.50±0.05 in the I/R group, and 0.32±0.04 in the Tβ4 group
(P<0.05). The rate of apoptosis in the Tβ4 group was lower
compared with that reported in the I/R group (P<0.05) (Fig. 4).
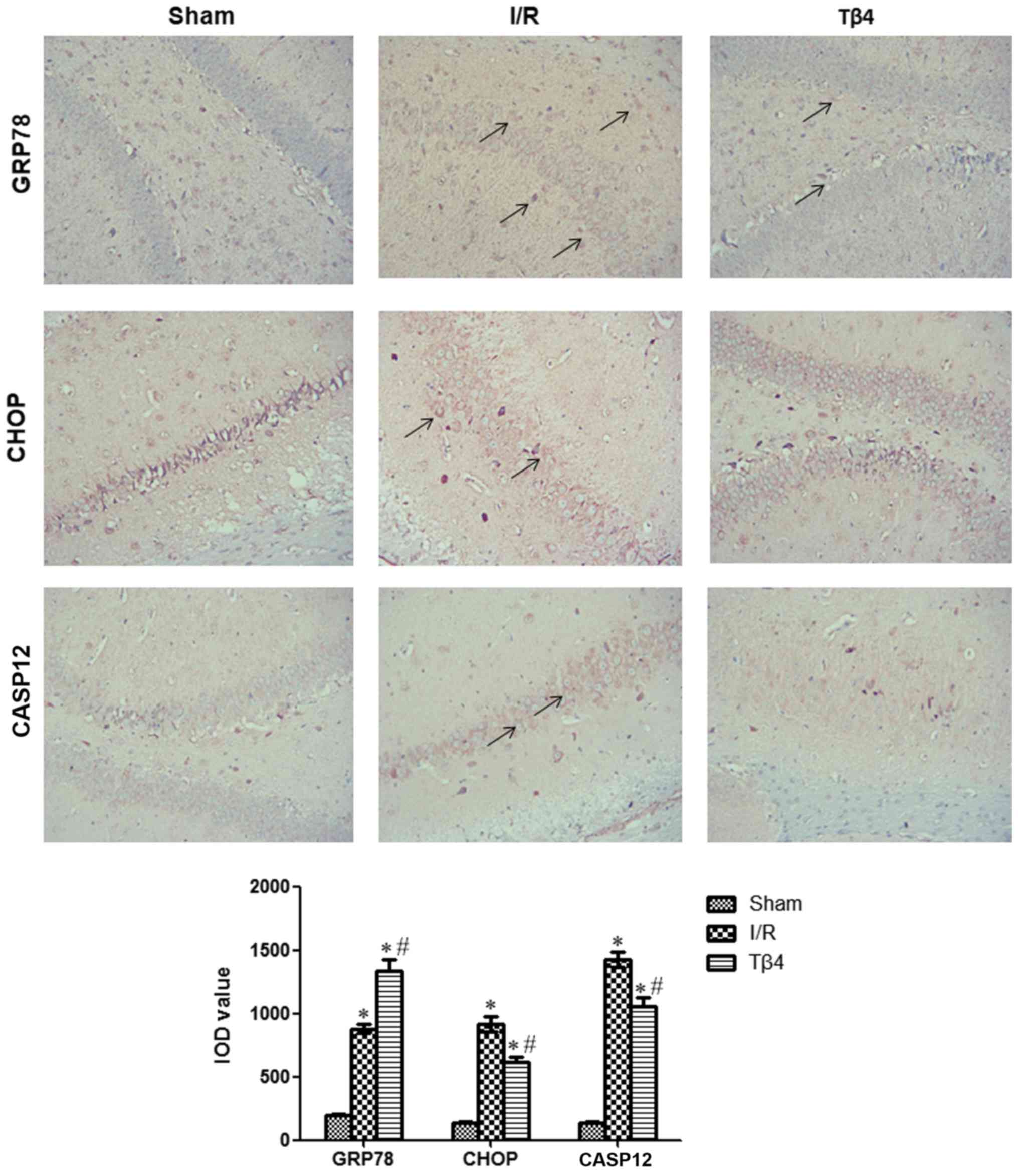
Tβ4 promotes the expression of GRP78
and reduces that of CHOP and CASP12
Immunohistochemical analysis revealed that the
number of GRP78-, CHOP-, and CASP12-positive cells in the I/R and
Tβ4 groups was significantly higher than that observed in the sham
group 24 h after reperfusion (P<0.05). The number of
GRP78-positive cells in the Tβ4 group was higher than that recorded
in the I/R group (P<0.05). In contrast, the number of CHOP- and
CASP12-positive cells in the Tβ4 group was lower than those
reported in the I/R group (P<0.05) (Fig. 5).
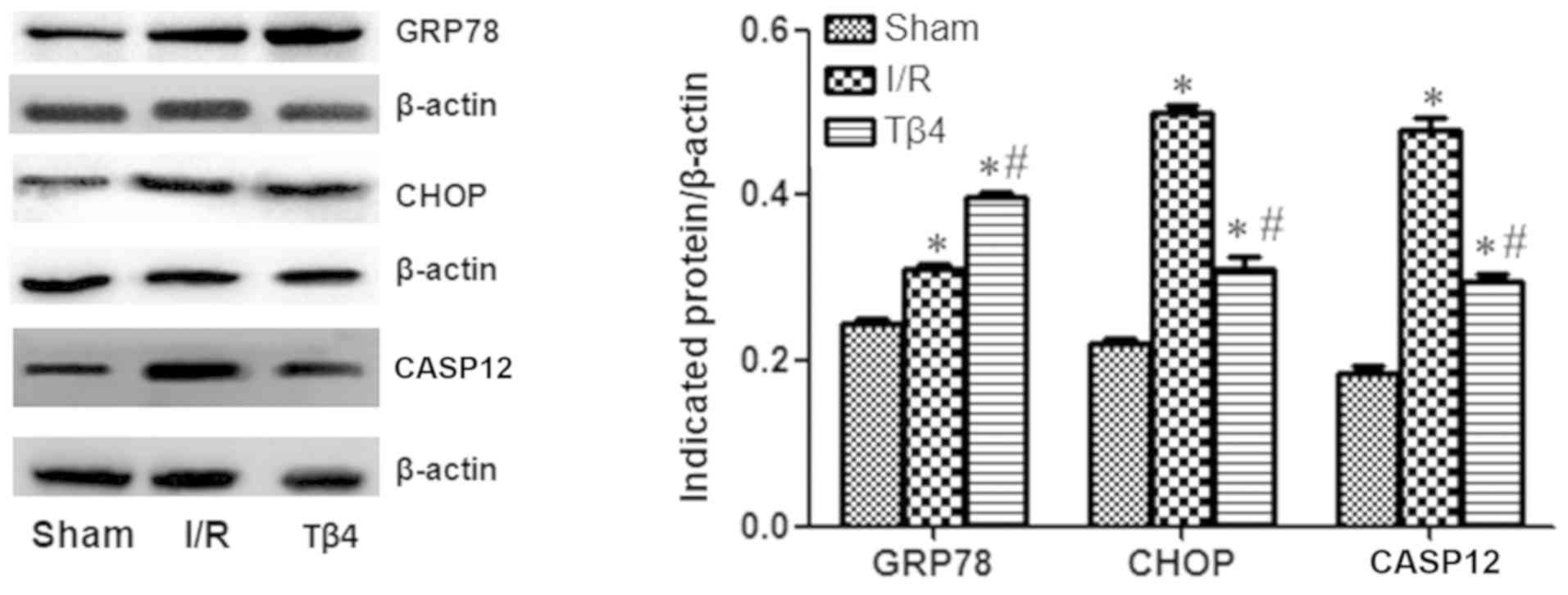
Moreover, western blotting revealed that the GRP78,
CHOP, and CASP12 proteins were highly expressed in the I/R and Tβ4
groups compared with the sham group. The expression of GRP78 was
higher in the Tβ4 group versus the I/R group. In contrast, the
expression of CHOP and CASP12 was lower in the Tβ4 group compared
to the I/R group (P<0.05) (Fig.
6).
Discussion
Considering that brain tissue has limited energy for
self-sustenance, energy metabolism in the brain is almost entirely
dependent on blood circulation (22). Therefore, brain tissue is extremely
sensitive to ischemia and hypoxia. The occurrence of cerebral
ischemia results in rapid depletion of the energy stores of the
brain, followed by initiation of neuronal programmed cell death
(23). In the present study, it
was demonstrated that intervention with Tβ4 after ischemia can
reduce the neurological deficits in rats. The mechanism involved in
this process was inhibition of neuronal apoptosis.
Tβ4 is a small molecule composed of 43 amino acid
residues. It is involved in multiple responses, such as wound
healing, tissue development, angiogenesis, and myocardial repair
(24–27). Tβ4 can inhibit apoptosis and exerts
neuroprotective effects (28). In
the present study, it was demonstrated that treatment with Tβ4
reduced the neurological deficits in rats. It also revealed
different degrees of apoptosis in the ischemic region of the rat
brain 24 h after reperfusion. This finding was consistent with the
results of previous research (29,30).
The rate of apoptosis in the Tβ4 group was significantly lower than
that observed in the I/R group, suggesting that Tβ4 can reduce
apoptosis induced by cerebral I/R. Cerebral infarction volume is
one of the most intuitive indicators for the evaluation of the
degree of brain tissue damage. In this experiment, brain infarction
in the Tβ4 group rats was significantly lower than those in the I/R
group, indicating that Tβ4 reduced the injury caused by I/R.
Compared with the Tβ4 group, the I/R group exhibited greater
infarct area and brain tissue loss, interstitial edema, neuronal
vacuolar degeneration, and nucleus pyrolysis. These results
indicated that Tβ4 exerted neuroprotective effects on neurological
behavior, cerebral infarct volume, microscopic pathology, and
apoptosis in rats with cerebral I/R injury.
The ER is one of important organelles of eukaryotic
cells. It is present in all cells, except the red blood cells.
Furthermore, ERS refers to the disruption of ER homeostasis by
harmful factors, such as ischemia, hypoxia, and glucose
deprivation. This process interferes with the function of the ER,
and can cause the accumulation of misfolded and unfolded proteins.
The unfolded protein response (UPR) induced by the accumulation of
unfolded/misfolded proteins in the ER is the most important
signaling mechanism of ERS (31,32).
GRP78 is a molecular chaperone located in the ER,
and plays an important role in maintaining the stability of the ER.
Studies have revealed that the upregulation of GRP78 is an
important marker of ERS, and can protect cells from stress.
Cerebral ischemia impairs energy metabolism in the cell, and leads
to the accumulation of unfolded or misfolded proteins. GRP78 is
rapidly upregulated to relieve ERS injury (33,34).
A previous study demonstrated that, under ERS, the upregulation of
GRP78 promoted the expression of pro-survival proteins, reduced
neuronal death, and exerted anti-apoptotic effects (35). The increase in GRP78 after ischemic
injury indicated the presence of a protective mechanism of the ER
in response to stress. Moreover, intervention with Tβ4 further
increased the expression of the GRP78 protein to reduce the
production of unfolded or misfolded proteins, maintain ER
homeostasis, and inhibit apoptosis. The mechanism of Tβ4 for the
reduction of apoptosis may involve the upregulation of GRP78 and
the UPR pathway to relieve ERS after cerebral I/R.
Under severe ERS, the function of the ER is
impaired, and the ER apoptotic signaling pathway is activated to
induce apoptosis. CHOP is an ERS-specific transcription factor.
Under normal conditions, CHOP is rarely expressed; hence, a marked
increase in its expression may be indicative of ERS (36). The activation of CASP12 is another
signal transduction pathway involved in ERS-induced apoptosis.
Studies have revealed that increased expression of CASP12 is an
important marker of ERS-induced apoptosis (37,38).
The present data revealed that, in the I/R rat
model, the expression of CHOP and CASP12 was significantly
increased, along with an increased rate of apoptosis. These
findings indicated that excessive ERS may initiate the apoptosis
pathway by upregulating the expression of CHOP and CASP12.
Treatment with Tβ4 reduced the expression of CHOP and CASP12
proteins in rat ischemic brain tissue. It is speculated that the
downregulation of the expression of CHOP and CASP12 may be one of
the mechanisms involved in the anti-apoptotic effect of Tβ4.
The present study had some limitations. Firstly,
although the expression of GRP78, CHOP, and CASP12 was detected in
the present study, and GRP78 and CHOP are typical markers of ERS,
the potential function of GRP78 and CHOP in the protection induced
by Tβ4 needs to be assessed. Secondly, other UPR pathways were not
investigated in the present study.
In conclusion, treatment with Tβ4 can reduce
cerebral I/R injury in rats through inhibition of neuronal
apoptosis, promoting the recovery of the normal physiological
functions of damaged cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by the grants from
Guangdong Provincial Medical Science and Technology Research Fund
(grant no. A2016468).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZZ, SL and SH performed the experiments and analyzed
the data. ZZ designed the study and wrote the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All animal procedures were approved by the Ethics
Committee of Guangzhou Medical University (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
GBD 2013 Mortailty and Cause of Death
Collaborators, . Global, regional, and national age-sex specific
all-cause and cause-specific mortality for 240 causes of death,
1990–2013: A systematic analysis for the global burden of disease
study 2013. Lancet. 385:117–171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lai T, Li M, Zheng L, Song Y, Xu X, Guo Y,
Zhang Y, Zhang Z and Mei Y: Over-expression of VEGF in marrow
stromal cells promotes angiogenesis in rats with cerebral
infarction via the synergistic effects of VEGF and Ang-2. J
Huazhong Uni Sci Technolog Med Sci. 32:724–731. 2012. View Article : Google Scholar
|
|
3
|
Benjamin EJ, Muntner P, Alonso A,
Bittencourt MS, Callaway CW, Carson AP, Chamberlain AM, Chang AR,
Cheng S, Das SR, et al: Heart disease and stroke statistics-2019
update: A report from the american heart association. Circulation.
139:e56–e528. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liberale L, Carbone F, Montecucco F,
Gebhard C, Lüscher TF, Wegener S and Camici GG: Ischemic stroke
across sexes: What is the status quo? Front Neuroendocrinol.
50:3–17. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma XH, Gao Q, Jia Z and Zhang ZW:
Neuroprotective capabilities of TSA against cerebral
ischemia/reperfusion injury via PI3K/Akt signaling pathway in rats.
Int J Neurosci. 125:140–146. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dong S, Tong X, Li J, Huang C, Hu C, Jiao
H and Gu Y: Total flavonoid of Litsea coreana leve exerts
anti-oxidative effects and alleviates focal cerebral
ischemia/reperfusion injury. Neural Regen Res. 8:3193–3202.
2013.PubMed/NCBI
|
|
7
|
Thompson JW, Narayanan SV, Koronowski KB,
Morris- Blanco K, Dave KR and Perez-Pinzon MA: Signaling pathways
leading to ischemic mitochondrial neuroprotection. J Bioenerg
Biomembr. 47:101–110. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sadana P, Coughlin L, Burke J, Woods R and
Mdzinarishvili A: Anti-edema action of thyroid hormone in MCAO
model of ischemic brain stroke: Possible association with AQP4
modulation. J Neurol Sci. 354:37–45. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang K: Integration of ER stress,
oxidative stress and the inflammatory response in health and
disease. Int J Clin Exp Med. 3:33–40. 2010.PubMed/NCBI
|
|
10
|
Shen YQ, Guerra-Librero A, Fernandez-Gil
BI, Florido J, García-López S, Martinez-Ruiz L, Mendivil-Perez M,
Soto-Mercado V, Acuña-Castroviejo D, Ortega-Arellano H, et al:
Combination of melatonin and rapamycin for head and neck cancer
therapy: Suppression of AKT/mTOR pathway activation and activation
of mitophagy and apoptosis via mitochondrial function regulation. J
Pineal Res. 64:2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cybulsky AV: Endoplasmic reticulum stress,
the unfolded protein response and autophagy in kidney diseases. Nat
Rev Nephrol. 13:681–696. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goldstein AL, Hannappel E and Kleinman HK:
Thymosin beta4: Actin-sequestering protein moonlights to repair
injured tissues. Trends Mol Med. 11:421–429. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wong CG, Taban M, Osann K, Ross-Cisneros
FN, Bruice TC, Zahn G and You T: Subchoroidal release of VEGF and
bFGF produces choroidal neovascularization in rabbit. Curr Eye Res.
42:237–243. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim S and Kwon J: Thymosin beta 4 improves
dermal burn wound healing via downregulation of receptor of
advanced glycation end products in db/db mice. Biochim Biophys
Acta. 1840:3452–3459. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sosne G and Ousler GW: Thymosin beta 4
ophthalmic solution for dry eye: A randomized, placebo-controlled,
phase II clinical trial conducted using the controlled adverse
environment (CAE™) model. Clin Ophthalmol. 9:877–884.
2015.PubMed/NCBI
|
|
16
|
Choi SY, Noh MR, Kim DK, Sun W and Kim H:
Neuroprotective function of thymosin-beta and its derivative
peptides on the programmed cell death of chick and rat neurons.
Biochem Biophys Res Commun. 362:587–593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Morris DC, Cui Y, Cheung WL, Lu M, Zhang
L, Zhang ZG and Chopp M: A dose-response study of thymosin β4 for
the treatment of acute stroke. J Neurol Sci. 345:61–67. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu XH, Bi HY, Cao J, Ren S and Yue SW:
Early constraint-induced movement therapy affects behavior and
neuronal plasticity in ischemia-injured rat brains. Neural Regen
Res. 14:775–782. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
O'Donnell ME, Tran L, Lam TI, Liu XB and
Anderson SE: Bumetanide inhibition of the blood-brain barrier
Na-K-Cl cotransporter reduces edema formation in the rat middle
cerebral artery occlusion model of stroke. J Cereb Blood Flow
Metab. 24:1046–1056. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li J, Yang S and Zhu G: Postnatal calpain
inhibition elicits cerebellar cell death and motor dysfunction.
Oncotarget. 8:87997–88007. 2017.PubMed/NCBI
|
|
21
|
Song ZJ, Yang SJ, Han L, Wang B and Zhu G:
Postnatal calpeptin treatment causes hippocampal neurodevelopmental
defects in neonatal rats. Neural Regen Res. 14:834–840. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang S, Li T, Ji T, Yi W, Yang Z, Wang S,
Yang Y and Gu C: AMPK: Potential therapeutic target for ischemic
stroke. Theranostics. 8:4535–4551. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lv J, Jiang S, Yang Z, Hu W, Wang Z, Li T
and Yang Y: PGC-1α sparks the fire of neuroprotection against
neurodegenerative disorders. Ageing Res Rev. 44:8–21. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuzan A: Thymosin β as an actin-binding
protein with a variety of functions. Adv Clin Exp Med.
25:1331–1336. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang YY, Zhu QS, Wang YW and Yin RF:
Thymosin beta-4 recombinant adeno-associated virus enhances human
nucleus pulposus cell proliferation and reduces cell apoptosis and
senescence. Chin Med J (Engl). 128:1529–1535. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sosne G, Rimmer D, Kleinman HK and Ousler
G: Thymosin beta 4: A potential novel therapy for neurotrophic
keratopathy, dry eye, and ocular surface diseases. Vitam Horm.
102:277–306. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stark CK, Tarkia M, Kentala R, Malmberg M,
Vähäsilta T, Savo M, Hynninen VV, Helenius M, Ruohonen S, Jalkanen
J, et al: Systemic dosing of thymosin beta 4 before and after
ischemia does not attenuate global myocardial ischemia-reperfusion
injury in pigs. Front Pharmacol. 7:1152016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deville C, Girard-Blanc C, Assrir N, Nhiri
N, Jacquet E, Bontems F, Renault L, Petres S and van Heijenoort C:
Mutations in actin used for structural studies partially disrupt
β-thymosin/WH2 domains interaction. FEBS Lett. 590:3690–3699. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang W, Liang G, Li X, Li Z, Gao X, Feng
S, Wang X, Liu M and Liu Y: Intracarotid transplantation of
autologous adipose-derived mesenchymal stem cells significantly
improves neurological deficits in rats after MCAo. J Mater Sci
Mater Med. 25:1357–1366. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu ZH, Cai M, Xiang J, Zhang ZN, Zhang JS,
Song XL, Zhang W, Bao J, Li WW and Cai DF: PI3K/Akt pathway
contributes to neuroprotective effect of Tongxinluo against focal
cerebral ischemia and reperfusion injury in rats. J Ethnopharmacol.
181:8–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brown MK and Naidoo N: The endoplasmic
reticulum stress response in aging and age-related diseases. Front
Physiol. 3:2632012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hetz C: The unfolded protein response:
Controlling cell fate decisions under ER stress and beyond. Nat Rev
Mol Cell Biol. 13:89–102. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li J, Ni M, Lee B, Barron E, Hinton DR and
Lee AS: The unfolded protein response regulator GRP78/BiP is
required for endoplasmic reticulum integrity and stress-induced
autophagy in mammalian cells. Cell Death Differ. 15:1460–1471.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ye Z, Wang N, Xia P, Wang E, Liao J and
Guo Q: Parecoxib suppresses CHOP and Foxo1 nuclear translocation,
but increases GRP78 levels in a rat model of focal ischemia.
Neurochemical Res. 38:686–693. 2013. View Article : Google Scholar
|
|
35
|
Chen HL, Qi H, Liu XJ and Wang MS: Effect
of electroacupuncture pretreatment on apoptotic neurons and
expression of GRP 78 and GADD 153 in the hippocampus in rats with
global cerebral ischemia/reperfusion injury. Zhen Ci Yan Jiu.
39:431–436. 2014.(In Chinese). PubMed/NCBI
|
|
36
|
Dong YF, Chen ZZ, Zhao Z, Yang DD, Yan H,
Ji J and Sun XL: Potential role of microRNA-7 in the
anti-neuroinflammation effects of nicorandil in astrocytes induced
by oxygen-glucose deprivation. J Neuroinflammation. 13:602016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tong Q, Wu L, Jiang T, Ou Z, Zhang Y and
Zhu D: Inhibition of endoplasmic reticulum stress-activated
IRE1α-TRAF2-caspase-12 apoptotic pathway is involved in the
neuroprotective effects of telmisartan in the rotenone rat model of
Parkinson's disease. Eur J Pharmacol. 776:106–115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Q, Liu J, Chen S, Liu J, Liu L, Liu
G, Wang F, Jiang W, Zhang C, Wang S and Yuan X: Caspase-12 is
involved in stretch-induced apoptosis mediated endoplasmic
reticulum stress. Apoptosis. 21:432–442. 2016. View Article : Google Scholar : PubMed/NCBI
|