Introduction
Myocardial cell death caused by ischemia-reperfusion
(I/R) is one of the main causes of high morbidity and mortality
worldwide (1). It is generally
believed that the use of thrombolysis agents or direct percutaneous
coronary intervention treatment of myocardial reperfusion could
increase the survival rate of patients. However, myocardial
reperfusion may induce oxidative stress and lead to inflammation
and apoptosis; therefore, the morbidity and mortality of ischemic
cardiomyopathy patients remain at a high level (2,3). To
the best of our knowledge, no effective therapeutic agents for I/R
injury have been discovered in clinical practice (4,5).
Researchers have indicated that severe endoplasmic
reticulum (ER) stress leads to apoptosis of cardiomyocytes in
vitro and in vivo (6,7).
Inhibitors of ER stress protect the heart by inhibiting
pathological changes and apoptosis (8). C/EBP homologous protein (CHOP) plays
a key role in ER stress-induced apoptosis; the ablation of CHOP
attenuates ER-mediated apoptosis (9). The development of ER stress is caused
by dissociating abundant molecular chaperone BiP/78-kDa
glucose-regulated protein (GRP78) signaling molecules in the ER
cavity (10). ER stress signals
can eventually trigger apoptotic CHOP expression (11,12).
Cells respond to exogenous stimuli by regulating
intracellular signaling pathways. The mitogen-activated protein
kinase (MAPK) signaling pathway, which is widely distributed in the
cell, contains extracellular signal regulating kinase 1/2 (ERK1/2),
p38 and c-Jun NH2-terminal kinase (JNK). These two signaling
pathways are known to play vital roles in cell differentiation,
proliferation and apoptosis as well as in cell apoptosis induced by
ER stress (11,13,14).
Specifically, in vivo animal studies have shown that
inhibition of sustained phosphorylation of MAPK (ERK1/2, JNK, p-38)
not only reduces myocardial damage (15,16),
but also enhances cardiac function (17). The MAPK pathway has attracted much
attention due to its critical involvement in the functions of the
heart (15,18).
Curcumin (Cur) is a polyphenol from Curcuma
longa (turmeric plant). Curcumin is an alcohol-based molecule
that exists in an organic solvent (19). Studies have shown that curcumin is
an effective molecule which exerts a variety of positive
pharmacological effects including anti-inflammatory (20,21),
antioxidant (22) and
anti-apoptotic effects (23).
However, the functional roles of Cur in H/R injury still remain
largely unexplored. Therefore, the present study aimed to determine
whether Cur relieves H/R injury and whether Cur can be used as an
effective therapeutic agent for clinical cardiac I/R injury.
Materials and methods
Reagents and cell line
Curcumin was obtained from Sigma-Aldrich; Merck KGaA
(cat. no. 08511; HPLC >98%; powder). The primary antibodies for
GRP78, CHOP, p-p38, p-JNK and p-ERK1/2 were purchased from Cell
Signaling Technology (CST), and the primary antibody for GAPDH was
purchased from Santa Cruz Biotechnology. The Cell Counting Kit-8
(CCK-8) (Dojindo, Kumamoto, Japan), and lactate dehydrogenase
(LDH), malondialdehyde (MAD) and superoxide dismutase (SOD) assay
kits were all purchased from Nanjing Jiancheng Bioengineering
Institute (Nanjing, China). The H9c2 cell line was obtained from
the American Type Culture Collection (ATCC, Manassas, VA, USA).
Establishment of a
hypoxia/reoxygenation cell model
It has been reported that H9c2 cells are used as a
cell model of cardiac ischemia-reperfusion injury in vitro
(24). H9c2 cardiomyocytes were
incubated in an incubator at 37°C with 95% N2 and 5%
CO2 and used for experiments when the cell confluency
reached ~90%. In brief, the cells were cultured with
phosphate-buffered saline (PBS) which was then replaced with 10%
fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.) and
Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher Scientific,
Inc.), and then placed in the hypoxic chamber (Stem Cell
Technologies) with 95% N2 in an incubator for 10 min at
37°C. Four hours later, 10% FBS in DMEM medium was added to the
cells and the cells were incubated under a normoxic condition (20%
O2, 5% CO2) without the chamber for another
4, 8 and 12 h at 37°C. The cultured cardiomyocytes in the control
group were then cultured in an incubator without any treatment.
Three complexes were set in each group.
Evaluation of cell morphology and
determination of cell viability
H9c2 cells were seeded in a 96-well plate at a
density of 5×103 cells/well. After being pre-treated
with Cur, the cells were exposed to 4 h of hypoxia and then 12 h of
reoxygenation. Then the H9c2 cells were placed under an inverted
light microscope (magnification, ×200) to observe changes in cell
morphology. After adding 10 µl CCK-8 solution, the cells were
incubated for 2 h at 37°C. Optical density (OD) at 450 nm was
detected by a microplate reader (Thermo Fisher Scientific, Inc.).
Cell viability was detected using the CCK-8 assay kit (Dojindo,
Kumamoto, Japan), according to the manufacturer's protocol.
LDH concentration
H9c2 cells were seeded in a 96-well plate at a
density of 5×103 cells/well. After being pre-treated
with Cur, the cells were exposed to 4 h of hypoxia and then 12 h of
reoxygenation, and an LDH assay kit was prepared to detect the LDH
activity in the culture medium, according to the manufacturer's
protocol. Briefly, the culture medium was centrifuged at 400 × g
for 10 min at room temperature to collect 20 µl of the supernatant,
which was then mixed with 2,4-dinitrophenylhydrazine and incubated
for 15 min at 37°C. NaOH (0.4 M) was added into the mixture and
incubated for another 15 min at 37°C. The mixture was maintained at
room temperature for 5 min, and subsequently the OD (450 nm) was
measured on a microplate reader (Thermo Fisher Scientific,
Inc.).
Measurement of MDA activity
H9c2 cells were seeded in a 96-well plate at a
density of 5×103 cells/well. After being pre-treated
with Cur, the cells were exposed to 4 h of hypoxia and then 12 h of
reoxygenation. Next, the H9c2 cells were collected in a centrifuge
tube and the supernatant was discarded after being centrifuged at
12,000 × g at 4°C for 10 min. RIPA lysis buffer was added to fully
isolate the cell protein lysate. The cells were centrifuged at
12,000 × g for 30 min at 4°C and then the supernatant was extracted
and placed on ice for testing. A mixture of MDA test working
solution and crude enzyme solution was used at a ratio of 3:1, and
the mixture was placed in a water bath at 100°C for 30 min and then
cooled on ice. According to the manufacturer's protocol, the OD at
532 nm was read using a microplate reader (Thermo Fisher
Scientific, Inc.).
Measurement of SOD activity
H9c2 cells were seeded in a 96-well plate at a
density of 5×103 cells/well. After being pre-treated
with Cur, the cells were exposed to 4 h of hypoxia and then 12 h of
reoxygenation. The H9c2 cells were then collected. Next, an
enhanced bicinchoninic acid (BCA) protein assay kit (Beyotime
Institute of Biotechnology, Shanghai, China) was used to measure
the protein concentration after the protein was isolated using RIPA
lysis buffer. To detect SOD levels, the protein was incubated with
an SOD kit for 20 min at 37°C, and the SOD value was examined at
450 nm using a microplate reader (Thermo Fisher Scientific,
Inc.).
Cell apoptosis
H9c2 cells were seeded in a 6-well plate at a
density of 1.3×105/well. After being pre-treated with
Cur, the cells were exposed to 4 h of hypoxia and then 12 h of
reoxygenation. Next, the supernatant was collected into a 15-ml
centrifuge tube, and the culture flask was gently washed once by
adding 2 ml of PBS. The cells were digested with trypsin (1 ml)
without ehylenediaminetetraacetic acid (EDTA) and shaken gently.
The supernatant was aspirated after the well became wet. The
mixture was maintained at room temperature for 1 min, and DMEM
containing 10% FBS was added to terminate the digestion. The cells
were centrifuged at 1,000 × g for 3 min and the supernatant was
removed. Next, the cells were washed twice with pre-cooled PBS and
resuspended in 1X Annexin V binding buffer. According to the
instructions included in the Annexin V-FITC cell apoptosis
detection kit (cat. no. K201-100; BioVision, Milpitas, CA, USA),
the H9c2 cells were collected and stained with Annexin V-FITC and
propidium iodide (PI) for 15 min and counted by flow cytometry
(version 10.0, FlowJo, FACSCalibur™; BD Biosciences) at room
temperature. Based on the flow cytometry scatter diagrams, the
lower left quadrant represented living cells (Annexin
V−/PI−), the lower right quadrant indicated
early apoptotic cells (Annexin V+/PI−), and
the upper right quadrant indicated late apoptotic cells (Annexin
V+/PI+) and the upper left quadrant
represented necrotic cells (Annexin V−/PI+).
The total apoptosis was calculated as the sum of the right upper
quadrant and right lower quadrant (25).
ROS measurement by flow cytometry
H9c2 cells were seeded in a 6-well plate at a
density of 1.3×105/well and were exposed to
hypoxia/reoxygenation (4/8 h) with or without pretreated with Cur
for 2 h. Next, the cells were incubated with 10 µmol/l carboxylated
2′,7′-dichlorodihydrofluorescein diacetate ester
(c-H2DCFDA, Beyotime Biotechnology, Shanghai, China, 5
µM) for 30 min to determine the oxidation of hydrogen peroxide
(H2O2) mediated into fluorescent compound DCF
at 37°C. Fluorescent compound DCF model was used for measuring the
fluorescence. The excitation wavelength of the flow cytometer
(version 10.0; FlowJo) was 480 nm and the emission wavelength was
525 nm (26), and the x-axis
indicated the fluorescence intensity.
Western blot analysis
H9c2 cells were seeded in a 6-well plate at a
density of 1.3×105/well and exposed to
hypoxia/reoxygenation (4/8 h) with or without pretreatment with 10
µM Cur for 2 h. After drug treatment, the H9c2 cells were flushed
by cold PBS for 3 times and put on ice with protein lysis buffer
(Radio Immunoprecipitation Assay, RIPA, cat. #9806; Cell Signaling
Technology, Inc.) for 2 h. The cells were centrifuged at 13,500 × g
for 30 min at 4°C and then the supernatant was extracted. The
concentration of protein was determined using a BCA protein assay
kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). SDS-PAGE with
10% running gels was used to separate the proteins (at least 40
µg), which were then transferred onto polyvinylidene fluoride
(PVDF) membranes (Bio-Rad Laboratories). To block the non-specific
signals, the membranes were incubated with 5% non-fat milk for at
least 2 h at room temperature. Protein strips were then incubated
with a primary antibody overnight at 4°C and then washed with 5%
bovine serum albumin (BSA, Gibco; Thermo Fisher Scientific, Inc.)
in PBS/0.1% Tween-20 (PBS/T) and incubated with the secondary
antibody for 1 h at room temperature. The protein strip was
developed with a developer (EZ-ECL kit; Biological Industries; BI)
and the protein quantity was analyzed using ImageJ software
(version 5.0; National Institutes of Health, Bethesda, MD, USA).
The antibodies used in the study were as follows: Anti-GAPDH
(mouse; dilution 1:1.000; cat. no. sc-47724; Santa Cruz
Biotechnology), anti-GRP78 (rabbit; dilution 1:500; cat. no. 3477;
CST), anti-CHOP (mouse; dilution 1:1,000; cat. no. 2895; CST),
anti-p-ERK1/2 (mouse; dilution 1:1,000; cat. no. 4307; CST),
anti-ERK1/2 (mouse; dilution 1:1,000; cat. no. 4695; CST),
anti-p-p38 (rabbit; dilution 1:1,000; cat. no. 4511; CST), anti-p38
(rabbit; dilution 1:1,000; cat. no. 9212; CST) anti-p-JNK (mouse;
dilution 1:1,000; cat. no. 9255; CST) and anti-JNK (mouse; dilution
1:1,000; cat. no. 9252; CST) as well as the secondary antibody
[horseradish peroxidase-labeled goat anti-rabbit or -mouse IgG
(1:5,000, cat. nos. sc-516102/sc-2357; Santa Cruz Biotechnology,
Inc.)].
RNA isolation and quantitative
real-time PCR
H9c2 cells were seeded in a 6-well plate at a
density of 1.3×105/well and exposed to
hypoxia/reoxygenation (4/8 h) with or without pretreatment with 10
µM Cur for 2 h. According to the instructions, total RNA of H9c2
cells was extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Chloroform (Sigma Aldrich; Merck KGaA) was added
to the tube and incubation was carried out at room temperature for
5 min and then the cells were centrifuged at 14,000 × g for 20 min
at 4°C. Next, the supernatant was transferred into a new tube and
isopropanol was added. The aqueous phase was centrifuged at 14,000
× g for 20 min at 4°C. The precipitate was washed with 70% ethanol
and suspended again in water treated with diethyl carbamate (DEPC).
The purity and concentration of RNA was tested by the NanoDrop
ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE,
USA), and the absorbance was read at 260 and 280 nm. According to
the program provided by the manufacturer (Thermo Fisher Scientific,
Inc.), reverse transcription cDNA kit was used to reversely
transcribe 1 µg total RNA for synthesis of cDNA (at 42°C for 60
min, at 70°C for 5 min, at 4°C preservation). SYBR-Green PCR Master
Mix (Roche, Basle, Switzerland) was used to perform quantitative
real-time polymerase chain reaction (qPCR) experiment using Opticon
Real-Time PCR Detection System (ABI 7500; Life Technology, USA).
The PCR cycle was as follows: pretreatment at 95°C for 10 min;
followed by 40 cycles of 94°C for 15 sec, 60°C for 1 min, finally
at 60°C for 1 min and at 4°C for preservation. The relative mRNA
quantity was determined using the comparative quantification cycle
(ΔΔCq) method (27). GAPDH
expression was used for normalization. The primer sequences were
used for RT-qPCR analysis as follows: GRP78s:
5′-GAACCAACTCACGTCCAACC-3′ (F) and 5′-AACCACCTTGAATGGCAAGA-3′ (R),
CHOP: 5′-GAAATCGAGCGCCTGACCAG-3′ (F) and
5′-GGAGGTGATGCCAACAGTTCA-3′ (R), GAPDH:
5′-CTCAACTACATGGTCTACATGTTCCA-3′ (F) and 5′-CCATTCTCGGCCTTGACTGT-3′
(R).
Statistical analysis
Data are shown as the mean ± SEM. Differences
between the experimental groups were assessed by ANOVA, followed by
Dunnett-t test, and analyzed by GraphPad Prism 6 (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
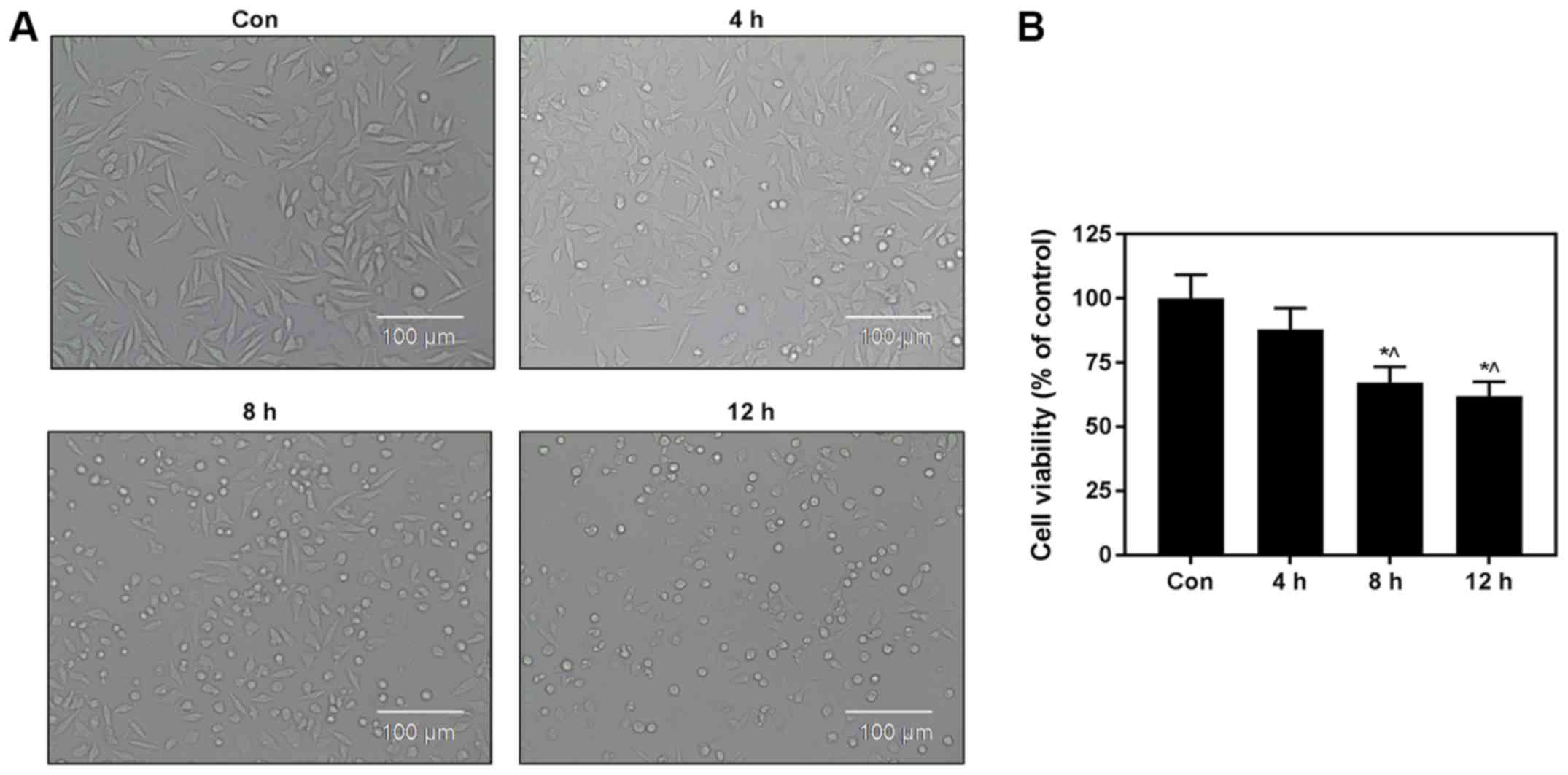
Hypoxia/reoxygenation injury model was
established in H9c2 cells
A model of hypoxia/reoxygenation (H/R) injury in
H9c2 cardiomyocytes was established in this study. Then, the cells
were exposed to 4 h of hypoxia and then 12 h of reoxygenation, and
the differences in cell morphology between the control group and
H/R groups were observed. Cell morphology was typical and regular
in the Con group, while a large number of cells exhibited a dot
state in the 4 h hypoxia/8 h reoxygenation and 4 h hypoxia/12 h
reoxygenation injury group, indicating that these cells may have
died in the H/R injury group (Fig.
1A). H9c2 cell viability was reduced in the 4 h hypoxia/8 h
reoxygenation and 4 h hypoxia/12 h reoxygenation injury groups,
compared with control group (Fig.
1B). These experimental results demonstrated that an H/R injury
cell model was successfully established.
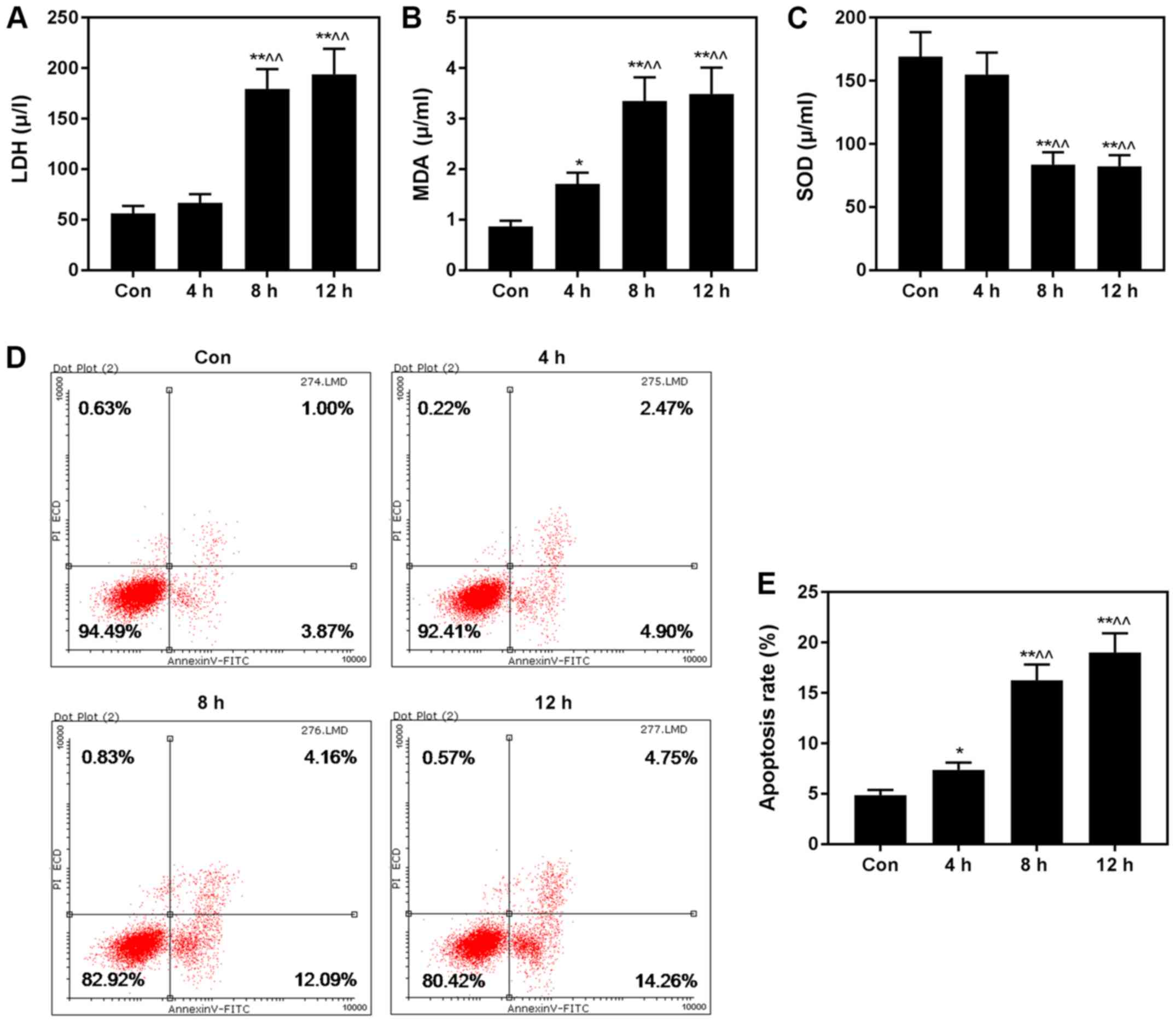
H/R injury induces apoptosis and
biomarkers
Furthermore, we assessed the levels of LDH, MDA and
SOD, and found that the levels of LDH (Fig. 2A) and MDA (Fig. 2B) were increased, while SOD
(Fig. 2C) was decreased in the H/R
groups, compared with control group. The rate of apoptosis was
significantly higher in the H/R injury groups than that in the
control group (Fig. 2D and E).
Thus, 4 h of hypoxia, followed by 8 h of reoxygenation was selected
to construct the H/R cell model for subsequent experiments.
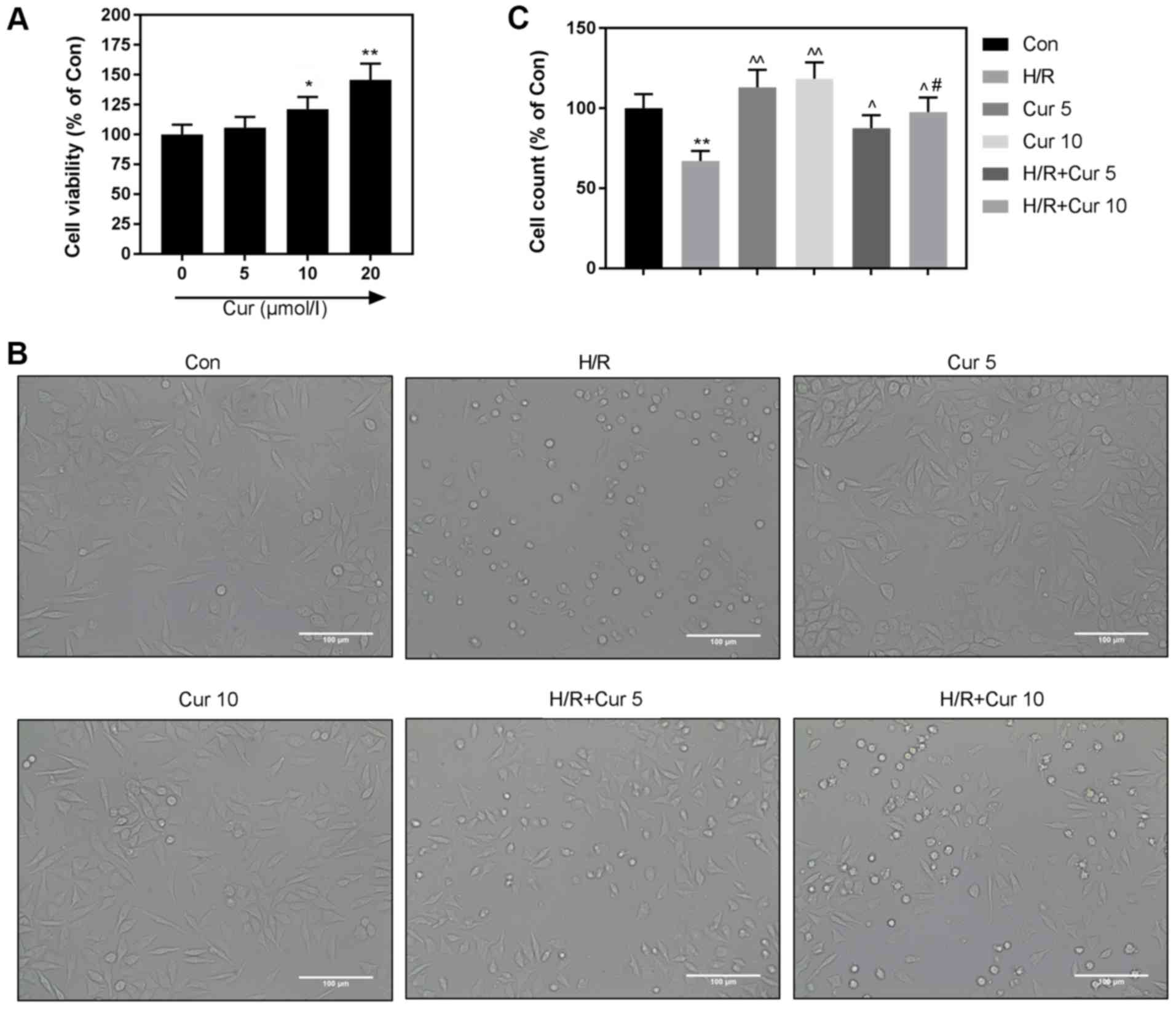
Cur exhibits no cytotoxicity on H9c2
cardiomyocytes
For the purpose of determining the cytotoxicity of
Cur on H9c2 cardiomyocytes, CCK-8 assay was used to determine the
viability of H9c2 cardiomyocytes following treatment of Cur at
different doses. The results revealed that no negative effect was
observed (Fig. 3A). Furthermore,
we found that 10 µM Cur had a better protective effect under H/R
(Fig. 3B and C). Thus, 10 µM Cur
was selected for subsequent experiments.
Cur inhibits ROS accumulation and
apoptosis caused by H/R injury
It has been reported that Cur protects heart cells
(28,29); however, its potential protective
mechanism still remains unclear. Thus, we determined whether Cur
protects H9c2 cardiomyocytes against H/R injury. We found that the
cell viability was higher in the H/R+Cur group than that in the H/R
group (Fig. 4A), and that the
levels of LDH, MDA and SOD were reversed by Cur, compared with
those in the H/R group (Fig.
4B-D). Similarly, H/R injury-induced apoptosis (Fig. 4E and G) and ROS (Fig. 4F and H) response were inhibited by
Cur. The results suggested that Cur protected the damage caused by
H/R injury.
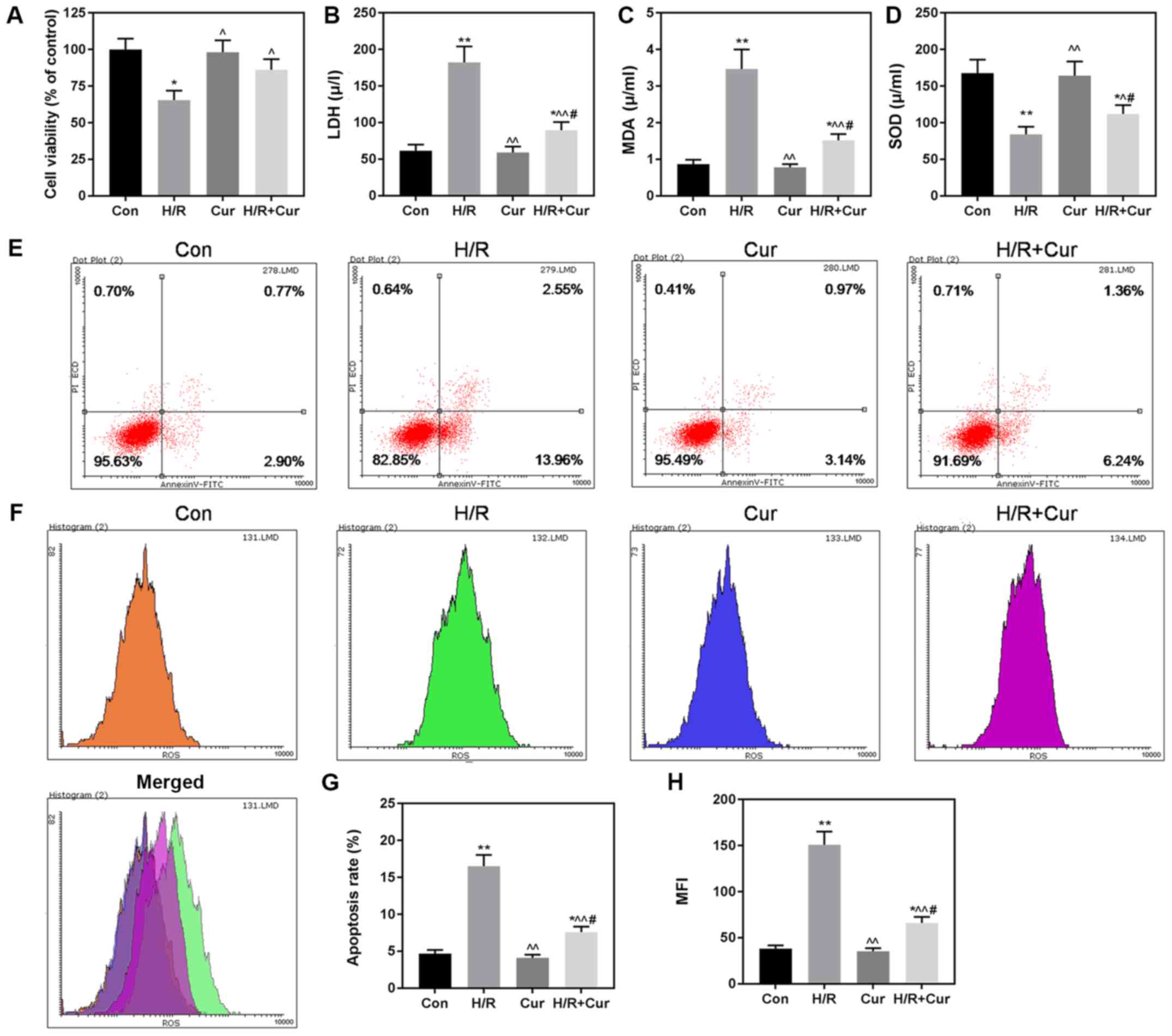 | Figure 4.Cur protects H9c2 cardiomyocytes
against H/R-induced cell death. (A) Cell viability was determined
by CCK-8 assay. The release level of LDH (B), and levels of MDA (C)
and SOD (D) in H9c2 cells were evaluated by relevant kits in line
with the manufacturer's instructions. (E and G) Apoptosis of H9c2
cardiomyocytes was analyzed and quantified by flow cytometry. (F
and H) ROS level in H9c2 cardiomyocytes was determined and
quantified by flow cytometry. n=3 wells/group. All data are
expressed by means ± SEM. *P<0.05, **P<0.01 vs. Con;
^P<0.05, ^^P<0.01 vs. H/R;
#P<0.05 vs. Cur. H/R, hypoxia/reoxygenation. LDH,
lactate dehydrogenase; MDA, malondialdehyde; SOD, superoxide
dismutase; ROS, reactive oxygen species; MFI, mean fluorescence
intensity; Con, control; Cur, Curcumin. |
ER stress and the MAPK signaling
pathway may be involved in the protective effect of Cur in H9c2
cells
In the present study, the expression levels of GRP78
and CHOP were found to be significantly increased by H/R at the
mRNA and protein levels, indicating that the activation of ER
stress and ER stress-related apoptotic signals were caused by H/R
injury (Fig. 5A-D). Pretreatment
with Cur (10 µM) for 2 h before exposure to H/R injury effectively
ameliorated these changes (Fig.
5A-D). We then further determined the effect of Cur on MAPK,
and found that MAPK-related protein expression levels of p-ERK1/2,
p-38 and p-JNK were suppressed by Cur, when compared to these
levels in the H/R group (Fig. 5E and
F).
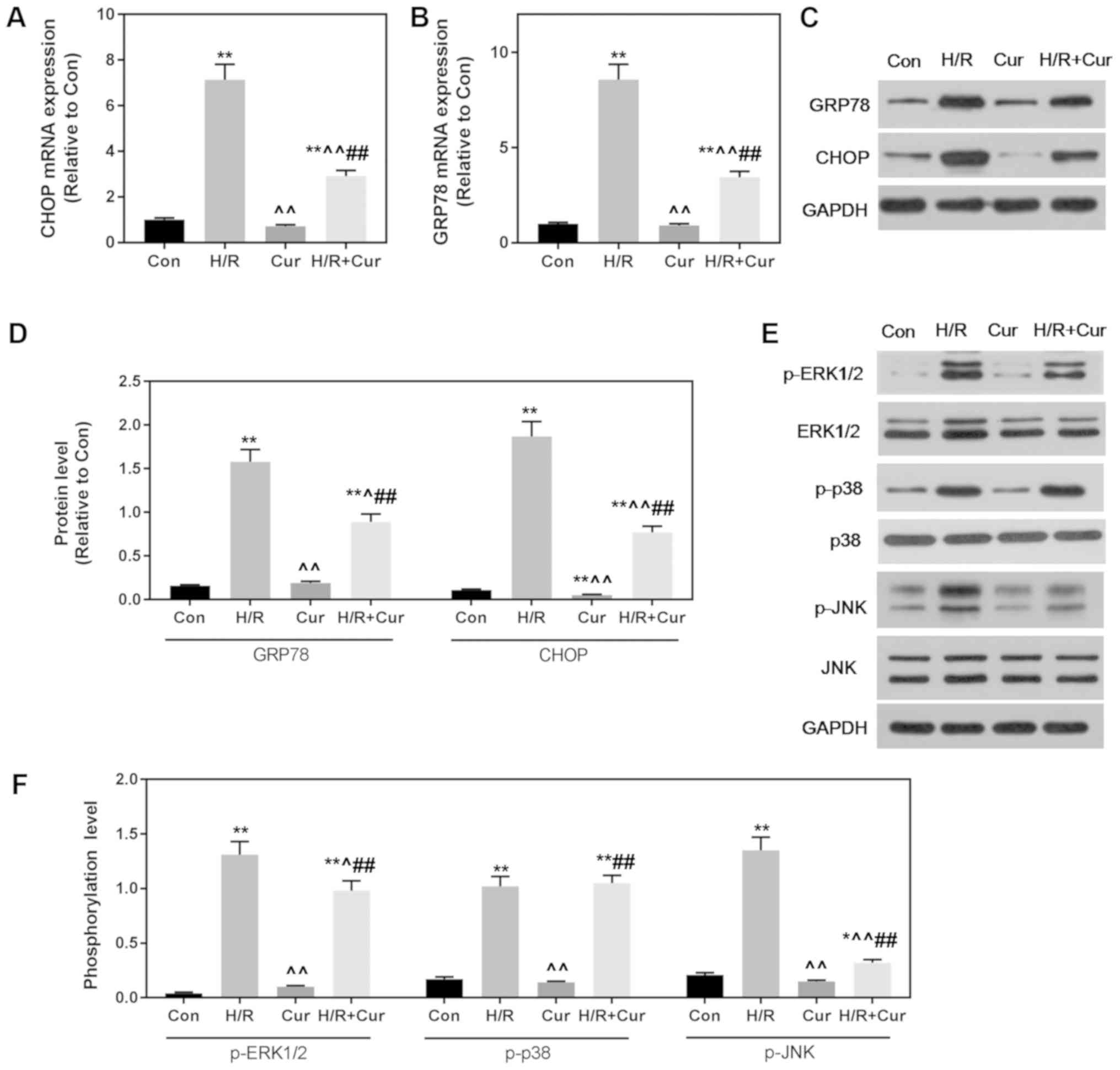 | Figure 5.Cur-mediated protection of H9c2
cardiomyocytes from H/R-induced cell death may involve ER stress
and the MAPK signal pathway. (A and B) ER stress-related mRNA
expression levels of CHOP and GRP78 were determined by reverse
transcription-quantitative PCR. (C and D) The protein expression
levels of GRP78 and CHOP were determined by western blot analysis
and quantified. GAPDH was used as a loading control. (E and F)
MAPK-related protein expression levels of p-ERK1/2, ERK1/2, p-p38,
p38, p-JNK and JNK were determined by western blot analysis and
quantified. GAPDH was used as a loading control. n=3 wells/group.
All data were expressed as the mean ± SEM. *P<0.05, **P<0.01
vs. Con; ^P<0.05, ^^P<0.01 vs. H/R;
##P<0.01 vs. Cur. CHOP, C/EBP homologous protein;
GRP78, 78-kDa glucose-regulated protein; MAPK, mitogen-activated
protein kinase; p-, phosphorylated; H/R, hypoxia/reoxygenation;
Cur, Curcumin. |
Discussion
In the hypoxia/reoxygenation (H/R) injury cell
model, we observed that the cellular morphology was obviously
altered and the levels of LDH and MDA and cell viability were
significantly decreased. Meanwhile, we found that the accumulation
of ROS during the development of reperfusion injury effectively
decreased SOD activity. We observed that apoptosis also occurred
following H/R injury, and that Cur strengthened the function of
cardiomyocytes under H/R injury and downregulated the activities of
LDH and MDA, therefore, it protected cardiomyocytes from injury. We
also revealed that the inhibitory effect of Cur on the accumulation
of ROS may be through the upregulation of SOD in H/R injury, and
Cur attenuated apoptosis. Cur also inhibited the expression of
GRP78 and CHOP, which are markers of ER stress and apoptosis.
Furthermore, Cur significantly decreased phosphorylation of MAPK
(ERK1/2 and JNK) in the H/R + Cur group, compared with that noted
in the H/R group. Taken together, these data showed that Cur
treatment produced a strong protective effect on H9c2
cardiomyocytes during H/R injury.
H9c2 cells exhibit a similar morphology to immature
embryo myocardial cell morphology, which retain the features of
adult cardiac muscle cells (30).
Many studies have also shown that H9c2 cells have been widely used
to study the pathological processes of enlarged heart and apoptosis
(31–33). Thus, we used H9c2 cells to set up a
cardiomyocyte ischemia-reperfusion injury model.
Cell viability is a vital indicator to cell
survival, and LDH is a marker of cardiac cell damage which leaks
into the bloodstream when the cell membrane becomes permeable or
ruptured (34). In the present
study, the cell viability and LDH levels were obviously decreased
in the H/R injury group and Cur pretreatment mitigated increases of
the LDH level and also increased H9c2 cell viability during H/R,
which suggested that Cur has protective effects on cardiomyocytes
against H/R-mediated damage.
Promoted by H/R injury, oxidative stress
accumulation is one of the causes of myocardial cell death
(35). ROS scavenger has been
shown to reduce the myocardial cell injury induced by H/R (36). Clinical studies and vector animal
experiments have shown that ischemia-reperfusion results in a
significant accumulation of oxidative stress (37,38).
Our experiment found that Cur reduced cell ROS production,
decreased cardiomyocyte MDA concentrations and increased SOD
activity, indicating that Cur relieved over-oxidation caused by
H/R. Studies have shown that apoptosis is induced by myocardial
ischemia/reperfusion in humans and rats (39,40),
and that ROS may activate apoptosis (1,41,42).
It is known that apoptosis is usually a passage to death after
cells have been damaged. Our experimental results also confirmed
that apoptosis was promoted by H/R injury and pretreatment with Cur
relieved apoptosis. These data indicate that Cur protects against
myocardial damage caused by H/R. Studies have shown that ER stress
is an important pathway in cardiac myocyte apoptosis during the
progression of H/R (43). ER is
considered to be an important organelle involved in the apoptotic
signaling pathway and a common feature of mediated destruction,
especially in apoptosis (44,45).
Activation of ER partner proteins is an early adoption of ER stress
response, and is also a sign indicating stress severity (46). In the present study, we found that
Cur downregulated ER chaperone protein GRP78, induced by H/R
injury. Transcription factor CHOP is one of the important factors
that promotes the ER stress-mediated apoptosis factor, playing an
important role in myocardial cell apoptosis (11,47).
ER stress can trigger pro-apoptotic signals including
CHOP-dependent pathways (48). We
discovered that the expression of CHOP was decreased by Cur, and
therefore, it could reasonably be assumed that the decrease in ER
chaperon proteins was attributed to ER stress inhibition. Thus,
these data suggest that ER stress relief was responsible for the
antiapoptotic effect of Cur.
The cell response to external stimuli is primarily
realized by modulating signaling pathways within the cell. Studies
have shown that MAPK plays a vital role in cardiomyocytes. The
dominant inactivation or constitutive activation of the p38 MAPK
and ERK1/2 signaling pathways indicate that activation of p38 MAPK
or the inhibition of ERK1/2 is essential for many cells (including
cardiomyocytes) to induce apoptosis (49). Studies have shown that the
inhibition of ERK1/2 could enhance apoptosis induced by
ischemia/reoxygenation and norepinephrine-induced hypertrophy
(50). In the present study, we
found that p-ERK1/2 in the H/R groups was suppressed by Cur at the
protein level. The suppression of p38 MAPK and JNK is effective at
reducing heart remodeling (51).
However, p-JNK rather than p-p38 was observed to be inhibited by
Cur in the H/R groups. Under physiological conditions, the ERK1/2
signaling pathway is a signaling pathway for survival, while p-38
and JNK are signaling pathways for promoting apoptosis and the
balance between the two is important in deciding the fate or
survival of the cell (15,49,52).
Cur significantly inhibited H/R injury-induced phosphorylation of
ERK1/2 and JNK rather than p-38, showing a better response of Cur
to ERK1/2 and JNK. Therefore, the suppression of apoptosis by Cur
may be related to inhibition of the ERK1/2 and JNK signaling
pathways.
In summary, we found that Cur exerted significant
effects, such as protecting cardiomyocytes against H/R injury
through the reduction of LDH and MDA activity, by decreasing ROS
production, and by increasing antioxidant activity and mitigating
ER stress and apoptosis induced by hypoxia/reoxygenation in H9c2
cells. Additionally, the anti-apoptotic effects of Cur may be
related to the reduction in the proteins levels of p-ERK1/2 and
p-JNK. Thus, Cur may be used as an effective therapeutic agent for
clinical cardiac ischemia-reperfusion injury.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
WW made substantial contributions to the conception
and design of the research study. JP and JL conducted all data
acquisition, data analysis and interpretation. WW performed the
drafting of the manuscript and critical revision for important
intellectual content. All authors approved the version to be
published. All authors agree to be accountable for all aspects of
the work in ensuring that questions related to the accuracy or
integrity of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Murphy E and Steenbergen C: Mechanisms
underlying acute protection from cardiac ischemia-reperfusion
injury. Physiol Rev. 88:581–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Frank A, Bonney M, Bonney S, Weitzel L,
Koeppen M and Eckle T: Myocardial ischemia reperfusion injury: From
basic science to clinical bedside. Semin Cardiothorac Vasc Anesth.
16:123–132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heusch G and Gersh BJ: The pathophysiology
of acute myocardial infarction and strategies of protection beyond
reperfusion: A continual challenge. Eur Heart J. 38:774–784.
2017.PubMed/NCBI
|
|
4
|
Hausenloy DJ and Yellon DM: Ischaemic
conditioning and reperfusion injury. Nat Rev Cardiol. 13:193–209.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heusch G: Critical issues for the
translation of cardioprotection. Circ Res. 120:1477–1486. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhuo XZ, Wu Y, Ni YJ, Liu JH, Gong M, Wang
XH, Wei F, Wang TZ, Yuan Z, Ma AQ and Song P: Isoproterenol
instigates cardiomyocyte apoptosis and heart failure via AMPK
inactivation-mediated endoplasmic reticulum stress. Apoptosis.
18:800–810. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dalal S, Foster CR, Das BC, Singh M and
Singh K: Β-adrenergic receptor stimulation induces endoplasmic
reticulum stress in adult cardiac myocytes: Role in apoptosis. Mol
Cell Biochem. 364:59–70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu T, Dong Z, Geng J, Sun Y, Liu G, Kang
W, Zhang Y and Ge Z: Valsartan protects against ER stress-induced
myocardial apoptosis via CHOP/Puma signaling pathway in
streptozotocin-induced diabetic rats. Eur J Pharm Sci. 42:496–502.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu HY, Okada K, Liao Y, Tsukamoto O,
Isomura T, Asai M, Sawada T, Okuda K, Asano Y, Sanada S, et al:
Ablation of C/EBP homologous protein attenuates endoplasmic
reticulum-mediated apoptosis and cardiac dysfunction induced by
pressure overload. Circulation. 122:361–369. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu XD, Zhang ZY, Sun S, Li YZ, Wang XR,
Zhu XQ, Li WH and Liu XH: Hypoxic preconditioning protects
microvascular endothelial cells against hypoxia/reoxygenation
injury by attenuating endoplasmic reticulum stress. Apoptosis.
18:85–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sari FR, Widyantoro B, Thandavarayan RA,
Harima M, Lakshmanan AP, Zhang S, Muslin AJ, Suzuki K, Kodama M and
Watanabe K: Attenuation of CHOP-mediated myocardial apoptosis in
pressure-overloaded dominant negative p38α mitogen-activated
protein kinase mice. Cell Physiol Biochem. 27:487–496. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Okada K, Minamino T, Tsukamoto Y, Liao Y,
Tsukamoto O, Takashima S, Hirata A, Fujita M, Nagamachi Y, Nakatani
T, et al: Prolonged endoplasmic reticulum stress in hypertrophic
and failing heart after aortic constriction: Possible contribution
of endoplasmic reticulum stress to cardiac myocyte apoptosis.
Circulation. 110:705–712. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arumugam S, Thandavarayan RA, Arozal W,
Sari FR, Giridharan VV, Soetikno V, Palaniyandi SS, Harima M,
Suzuki K, Nagata M, et al: Quercetin offers cardioprotection
against progression of experimental autoimmune myocarditis by
suppression of oxidative and endoplasmic reticulum stress via
endothelin-1/MAPK signalling. Free Radic Res. 46:154–163. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Muslin AJ: MAPK signalling in
cardiovascular health and disease: Molecular mechanisms and
therapeutic targets. Clin Sci (Lond). 115:203–218. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen S, Yang B, Xu Y, Rong Y and Qiu Y:
Protection of Luteolin-7-O-glucoside against apoptosis induced by
hypoxia/reoxygenation through the MAPK pathways in H9c2 cells. Mol
Med Rep. 17:7156–7162. 2018.PubMed/NCBI
|
|
16
|
Martin JL, Avkiran M, Quinlan RA, Cohen P
and Marber MS: Antiischemic effects of SB203580 are mediated
through the inhibition of p38alpha mitogen-activated protein
kinase: Evidence from ectopic expression of an inhibition-resistant
kinase. Circ Res. 89:750–752. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nagarkatti DS and Sha'afi RI: Role of p38
MAP kinase in myocardial stress. J Mol Cell Cardiol. 30:1651–1664.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maulik A, Davidson SM, Piotrowska I,
Walker M and Yellon DM: Ischaemic preconditioning protects
cardiomyocytes from anthracycline-induced toxicity via the PI3K
pathway. Cardiovasc Drugs Ther. 32:245–253. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Manolova Y, Deneva V, Antonov L, Drakalska
E, Momekova D and Lambov N: The effect of the water on the curcumin
tautomerism: A quantitative approach. Spectrochim Acta A Mol Biomol
Spectrosc. 132:815–820. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sikora E, Scapagnini G and Barbagallo M:
Curcumin, inflammation, ageing and age-related diseases. Immun
Ageing. 7:12010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Koeberle A and Werz O: Multi-target
approach for natural products in inflammation. Drug Discov Today.
19:1871–1882. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakmareong S, Kukongviriyapan U,
Pakdeechote P, Donpunha W, Kukongviriyapan V, Kongyingyoes B,
Sompamit K and Phisalaphong C: Antioxidant and vascular protective
effects of curcumin and tetrahydrocurcumin in rats with
L-NAME-induced hypertension. Naunyn Schmiedebergs Arch Pharmacol.
383:519–529. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Topcu-Tarladacalisir Y, Akpolat M, Uz YH,
Kizilay G, Sapmaz-Metin M, Cerkezkayabekir A and Omurlu IK: Effects
of curcumin on apoptosis and oxidoinflammatory regulation in a rat
model of acetic acid-induced colitis: The roles of c-Jun N-terminal
kinase and p38 mitogen-activated protein kinase. J Med Food.
16:296–305. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Anestopoulos I, Kavo A, Tentes I,
Kortsaris A, Panayiotidis M, Lazou A and Pappa A: Silibinin
protects H9c2 cardiac cells from oxidative stress and inhibits
phenylephrine-induced hypertrophy: Potential mechanisms. J Nutr
Biochem. 24:586–594. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jia WQ, Wang ZT, Zou MM, Lin JH, Li YH,
Zhang L and Xu RX: Verbascoside inhibits glioblastoma cell
proliferation, migration and invasion while promoting apoptosis
through upregulation of protein tyrosine phosphatase SHP-1 and
inhibition of STAT3 phosphorylation. Cell Physiol Biochem.
47:1871–1882. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Al-Obeed O, Vaali-Mohammed MA, Eldehna WM,
Al-Khayal K, Mahmood A, Abdel-Aziz HA, Zubaidi A, Alafeefy A,
Abdulla M and Ahmad R: Novel quinazoline-based sulfonamide
derivative (3D) induces apoptosis in colorectal cancer by
inhibiting JAK2-STAT3 pathway. OncoTargets Ther. 11:3313–3322.
2018. View Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Naserzadeh P, Mehr SN, Sadabadi Z, Seydi
E, Salimi A and Pourahmad J: Curcumin protects mitochondria and
cardiomyocytes from oxidative damage and apoptosis induced by
Hemiscorpius lepturus venom. Drug Res (Stuttg). 68:113–120.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lv FH, Yin HL, He YQ, Wu HM, Kong J, Chai
XY and Zhang SR: Effects of curcumin on the apoptosis of
cardiomyocytes and the expression of NF-κB, PPAR-γ and Bcl-2 in
rats with myocardial infarction injury. Exp Ther Med. 12:3877–3884.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hescheler J, Meyer R, Plant S, Krautwurst
D, Rosenthal W and Schultz G: Morphological, biochemical, and
electrophysiological characterization of a clonal cell (H9c2) line
from rat heart. Circ Res. 69:1476–1486. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dai D, Yang J, Zhao C, Wu H, Ding J, Sun X
and Hu S: Effect of geranylgeranyl pyrophosphate synthase on
hypoxia/reoxygenation-induced injury in heart-derived H9c2 cells.
Int Heart J. 59:821–828. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng J, Wu Q, Lv R, Huang L, Xu B, Wang
X, Chen A and He F: MicroRNA-449a inhibition protects H9C2 cells
against hypoxia/reoxygenation-induced injury by targeting the
Notch-1 signaling pathway. Cell Physiol Biochem. 46:2587–2600.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang HH, Chen Y, Gao CY, Cui ZT and Yao
JM: Protective effects of microRNA-126 on human cardiac
microvascular endothelial cells against
hypoxia/reoxygenation-induced injury and inflammatory response by
activating PI3K/Akt/eNOS signaling pathway. Cell Physiol Biochem.
42:506–518. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cai Y, Hu X, Yi B, Zhang T and Wen Z:
Glucagon-like peptide-1 receptor agonist protects against
hyperglycemia-induced cardiocytes injury by inhibiting high
mobility group box 1 expression. Mol Biol Rep. 39:10705–10711.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bartz RR, Suliman HB and Piantadosi CA:
Redox mechanisms of cardiomyocyte mitochondrial protection. Front
Physiol. 6:2912015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Granger DN and Kvietys PR: Reperfusion
injury and reactive oxygen species: The evolution of a concept.
Redox Biol. 6:524–551. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fan Q, Chen M, Fang X, Lau WB, Xue L, Zhao
L, Zhang H, Liang YH, Bai X, Niu HY, et al: Aging might augment
reactive oxygen species (ROS) formation and affect reactive
nitrogen species (RNS) level after myocardial ischemia/reperfusion
in both humans and rats. Age (Dordr). 35:1017–1026. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rachmat FD, Rachmat J, Sastroasmoro S and
Wanandi SI: Effect of allopurinol on oxidative stress and hypoxic
adaptation response during surgical correction of tetralogy of
fallot. Acta Med Indones. 45:94–100. 2013.PubMed/NCBI
|
|
39
|
Liu M, Zhang P, Chen M, Zhang W, Yu L,
Yang XC and Fan Q: Aging might increase myocardial
ischemia/reperfusion-induced apoptosis in humans and rats. Age
(Dordr). 34:621–632. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hu Q, Luo W, Huang L, Huang R and Chen R:
Apoptosis-related microRNA changes in the right atrium induced by
remote ischemic perconditioning during valve replacement surgery.
Sci Rep. 6:189592016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li Q, Xiang Y, Chen Y, Tang Y and Zhang Y:
Ginsenoside Rg1 protects cardiomyocytes against
hypoxia/reoxygenation injury via activation of Nrf2/HO-1 signaling
and inhibition of JNK. Cell Physiol Biochem. 44:21–37. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Molavi B and Mehta JL: Oxidative stress in
cardiovascular disease: Molecular basis of its deleterious effects,
its detection, and therapeutic considerations. Curr Opin Cardiol.
19:488–493. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang C, Tang Y, Li Y, Xie L, Zhuang W,
Liu J and Gong J: Unfolded protein response plays a critical role
in heart damage after myocardial ischemia/reperfusion in rats. PLoS
One. 12:e01790422017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu J, Hu H, Chen B, Yue R, Zhou Z, Liu Y,
Zhang S, Xu L, Wang H and Yu Z: Lycopene protects against
hypoxia/reoxygenation injury by alleviating ER stress induced
apoptosis in neonatal mouse cardiomyocytes. PLoS One.
10:e01364432015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zou XJ, Yang L and Yao SL: Endoplasmic
reticulum stress and C/EBP homologous protein-induced Bax
translocation are involved in angiotensin II-induced apoptosis in
cultured neonatal rat cardiomyocytes. Exp Biol Med (Maywood).
237:1341–1349. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gupta MK, Tahrir FG, Knezevic T, White MK,
Gordon J, Cheung JY, Khalili K and Feldman AM: GRP78 interacting
partner Bag5 responds to ER stress and protects cardiomyocytes from
ER stress-induced apoptosis. J Cell Biochem. 117:1813–1821. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Long M, Chen X, Wang N, Wang M, Pan J,
Tong J, Li P, Yang S and He J: Proanthocyanidins protect epithelial
cells from zearalenone-induced apoptosis via inhibition of
endoplasmic reticulum stress-induced apoptosis pathways in mouse
small intestines. Molecules. 23:E15082018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gorman AM, Healy SJ, Jäger R and Samali A:
Stress management at the ER: Regulators of ER stress-induced
apoptosis. Pharmacol Ther. 134:306–316. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xia Z, Dickens M, Raingeaud J, Davis RJ
and Greenberg ME: Opposing effects of ERK and JNK-p38 MAP kinases
on apoptosis. Science. 270:1326–1331. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yue TL, Wang C, Gu JL, Ma XL, Kumar S, Lee
JC, Feuerstein GZ, Thomas H, Maleeff B and Ohlstein EH: Inhibition
of extracellular signal-regulated kinase enhances
Ischemia/Reoxygenation-induced apoptosis in cultured cardiac
myocytes and exaggerates reperfusion injury in isolated perfused
heart. Circ Res. 86:692–699. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
See F, Thomas W, Way K, Tzanidis A, Kompa
A, Lewis D, Itescu S and Krum H: p38 mitogen-activated protein
kinase inhibition improves cardiac function and attenuates left
ventricular remodeling following myocardial infarction in the rat.
J Am Coll Cardiol. 44:1679–1689. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hassanzade A, Mandegary A, Sharif E,
Rasooli R, Mohammadnejad R and Masoumi-Ardekani Y: Cyclooxygenase
inhibitors combined with deuterium-enriched water augment
cytotoxicity in A549 lung cancer cell line via activation of
apoptosis and MAPK pathways. Iran J Basic Med Sci. 21:508–516.
2018.PubMed/NCBI
|