Introduction
Bacterial pneumonia, including Streptococcus
pneumoniae, Staphylococcus aureus, Klebsiella pneumoniae,
Haemophilus influenzae and Pseudomonas aeruginosa, is
the most common form of pneumonia and one of the most common
infectious diseases, and poses a threat to the health of children
and the elderly (1). Among all
bacteria, S. pneumoniae is the most common cause of
pneumonia (2). Although
antibiotics are the main treatment for pneumonia, the antimicrobial
resistance of S. pneumoniae pneumonia has increased
significantly over the past few decades (3). Therefore, there is concern regarding
S. pneumoniae pneumonia (4,5). The
diseases caused by S. pneumoniae can be partially attributed
to the inflammatory response during infection; however, research
into the inflammatory injury caused by S. pneumoniae
pneumonia is required.
According to a previous study, in the early stage of
pneumonia, the phenotype of macrophages transforms and they release
a large number of proinflammatory cytokines, including tumor
necrosis factor-α (TNF-α), interleukin (IL)-6 and IL-8, the
pathogen invades the alveoli, leading to an imbalance of the
inflammatory response and to acute lung injury (6). The occurrence and development of
pneumonia is related to the imbalance of the inflammatory response
(7–9). Follistatin-like protein 1 (FSTL-1) is
a secretory glycoprotein produced mainly by cells of the
mesenchymal lineage, including cardiomyocytes, chondrocytes,
fibroblasts and endotheliocytes, which regulate various biological
functions, such as cell proliferation, differentiation, apoptosis
and metabolism (10). A previous
study showed that FSTL-1 serves as a proinflammatory protein that
plays important roles in the inflammatory response (11). Increased levels of FSTL-1 in
macrophages lead to the upregulation of proinflammatory factors,
including IL-1β, TNF-α and IL-6 (12), supporting a role for FSTL-1 in
inflammation. Furthermore, Chaly et al (13) found that FSTL-1 modulated the
proinflammatory factor IL-1β by activating nucleotide
oligomerization domain-like receptor protein 3 (NLRP3) in monocytes
and macrophages. The activation of NLRP3 recruits
apoptosis-associated spot-like protein (ASC) and pro-caspase-1,
converting inactive pro-caspase-1 into active caspase-1, promoting
the maturation of pro-IL-1β and pro-IL-18, which in turn activate
caspase-1 through autocrine or paracrine signaling to release more
mature IL-1β (14,15). The persistence of IL-1β amplifies
the inflammatory response, leading to inflammation damage in normal
tissues (16). Therefore, the
present study was designed to understand the molecular mechanism of
FSTL-1, and determine the relationship between FSTL-1 and NLRP3 in
inflammation injury during S. pneumoniae infection.
Toll-like receptors (TLRs) serve important roles in
preventing pathogen invasion by detecting pathogen-associated
molecular patterns (PAMPs) (17).
In innate immune cells, TLRs initiate complex signaling pathways by
recognizing PAMPs, leading to the activation of several
transcription factors, including NF-κB and activator protein-1,
which are involved in the production of proinflammatory cytokines,
chemokines and the induction of various regulatory genes (18). In the present study, whether the
TLR4/NF-κB signaling pathway is involved in the inflammatory
response regulated by FSTL-1 during S. pneumoniae infection
was investigated.
In this study, the functional role of FSTL-1 in
vitro and in vivo after S. pneumoniae infection
was examined. The results indicated that downregulation of FSTL-1
suppressed inflammation injury in bone marrow-derived macrophages
(BMDMs) after S. pneumoniae infection through the NLRP3 and
TLR4/NF-κB signaling pathways.
Materials and methods
Bacterial culture
S. pneumoniae (ATCC 6305; Bena Culture
Collection) was cultured in brain heart infusion broth (BD
Biosciences) under anaerobic conditions (95% N2/5%
CO2) overnight at 37°C. Dilution of samples in brain
heart infusion broth was used to quantify colony forming units
(CFU), and colony counts × dilution was used to determine
titers.
Animals
A total of 25 (11 male and 14 female) wild type (WT)
and 25 (11 male and 14 female) FSTL-1 knockout (KO) C57BL/6 mice
(8–12 weeks old, 22–25 g) were provided by CasGene Biotech Co., Ltd
(Beijing, China). The mice were fed on standard pellet chow and
water ad libitum and housed in a controlled SPF-class
environment with 12-h light/dark cycle at an ambient temperature of
24±1°C and a humidity of 55±5%. The present study was conducted
according to Xian Yang Central Hospital medical guidelines and the
experiment was approved by Xian Yang Central Hospital Ethics
Committee.
S. pneumoniae infection
The mice were anesthetized with 2% isoflurane in
O2 using a face mask for 5 min and were then
intranasally inoculated with 1×103 CFU of S.
pneumoniae (50 µl in PBS) once when they were fully sedated.
The uninfected wild-type (WT) mice and FSTL-1 KO mice were treated
with only 50 µl of PBS. The animals were euthanized and samples
were collected for further analysis 7 days after infection.
Cell culture
BMDMs were obtained from the bone marrow of the
femur and tibia of the C57BL/6 mice as previously described
(19). Bone marrow cells were
added to L929 media containing DMEM (Life Technology; Thermo Fisher
Scientific, Inc.), containing 40 ng/ml macrophage
colony-stimulating factor (Life Technology; Thermo Fisher
Scientific, Inc.), 10% FBS (Life Technology; Thermo Fisher
Scientific, Inc.), 1% non-essential amino acids (Life Technology;
Thermo Fisher Scientific, Inc.) and 1% cyanomycin (Life Technology;
Thermo Fisher Scientific, Inc.). The cells were cultured for 2 days
at 37°C with 5% CO2 and the bone marrow cells
differentiated into macrophages. The BMDMs were washed three times
with PBS and cultured in DMEM containing 10% FBS. BMDMs were
collected 0, 4, 12, 24 and 36 h after infection with S.
pneumoniae (50 CFU) or media alone for RT-qPCR, western
blotting and ELISA analysis.
Cell transfection
Lentiviral-based scrambled-short hairpin (sh)RNA
vectors, FSTL-1-shRNA vectors, pcDNA3.1-FSTL-1-overexpression
(FSTL-1-OE) vectors, FSTL-1-control vectors and NLRP3-shRNA vectors
were synthesized by Shanghai GeneChem Co., Ltd. FSTL-1-shRNA and
FSTL-1-OE vectors were used to knockdown or overexpress FSTL-1,
respectively. NLRP3-shRNA vectors were used to knockdown NLRP3.
Full-length FSTL-1 cDNA was cloned into pcDNA3.1 plasmid and the
shRNAs targeting FSTL-1 or NLRP3 was inserted into pLKO.1 plasmid.
The target sequence of FSTL-1 shRNA was CTTCCTCGGAGCGTGGTGA, NLRP3
shRNA was CTCGGTCCAACATCTAGCA. Then, 5×105 cells/well
BMDMs were infected with a mixture of 40 nmol/ml shRNA or 50
nmol/ml pcDNA3.1 and the transfection was performed using
Lipofectamine 2000® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) following manufacturer's instructions for 3
days.
Sample collection and analysis
The bronchoalveolar lavage fluids (BALFs) and lung
tissues of the mice were collected at 7 days post-infection
(n=6/group). BALF was harvested using 1.5 ml PBS supplemented with
complete protease inhibitor cocktail (Roche Diagnostics). BALFs
were isolated via centrifugation at 1,000 × g for 10 min at 4°C and
then stored at −80°C for further analysis. The fresh lung tissues
collected from mice with different treatments were immediately
snap-frozen in liquid nitrogen and stored at −80°C until use.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from BMDMs using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. Total RNA was quantified using a
NanoDrop™ 2000 (NanoDrop Technologies; Thermo Fisher Scientific,
Inc.). Complementary DNA was synthesized using the M-MLV Reverse
Transcriptase kit (Invitrogen; Thermo Fisher scientific, Inc.). The
RT conditions were the following: 5 min at 25°C, 30 min at 42°C and
5 min at 85°C. The quantification of all gene transcripts was
performed using an SYBR Green II real-time PCR kit (Takara Bio,
Inc.). The thermocycling conditions: Initial denaturation at 95°C
for 3 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 60
sec. β-actin was used as an internal control. Each experiment was
run in triplicate. The 2−∆∆Cq method (20) was used calculate the relative
expression of genes. The following primers were used: FSTL-1,
forward 5′-TTATGATGGGCACGCAAAGAA-3′, reverse
5′-ACTGCCTTTAGAGAACCAGCC-3′; NLRP3, forward
5′-CCAGACACTCATGTTGCCTGTTC-3′, reverse 5′-GAGGCTCCGGTTGGTGCTTA-3′;
and β-actin, forward 5′-CCTCATGAAGATCCTGACCG-3′ and reverse
5′-ACCGCTCATTGCCGATAGTG-3′.
ELISAs
TNF-α (cat. no. PT512), IL-6 (cat. no. PI326) and
IL-1β (cat. no. PI301) ELISA kits were purchased from Beyotime
Institute of Biotechnology. The production of TNF-α, IL-6 and IL-1β
in the supernatants of BMDMs and BALFs of mice following different
treatments was detected using the ELISA kits according to the
manufacturer's protocols. A microplate reader was used to measure
the optical density of each group at 450 nm.
Western blotting
The proteins were extracted from BMDMs and lung
tissues with the use of radioimmunoprecipitation assay buffer
(Beijing Solarbio Science & Technology Co., Ltd.), and the
protein concentration was measured using a BCA Protein assay kit
(Thermo Fisher Scientific, Inc.). Subsequently, an equal amount of
protein (50 µg) was separated via 10–12% SDS-PAGE and transferred
to nitrocellulose membranes. After blocking with 5% skim milk for 2
h at room temperature, the membranes were incubated with primary
rabbit anti-FSTL-1 (1:500; Abcam; cat. no. ab71548), anti-TLR4
(1:500; Abcam; cat. no. ab13867), rabbit anti-NLRP3 (1:1,000;
Abcam; cat. no. ab214185), rabbit anti-ASC (1:1,000; Cell Signaling
Technology, Inc.; cat. no. 67824), rabbit anti-cleaved-caspase-1
(1:1,000; Cell Signaling Technology, Inc.; cat. no. 89332), rabbit
anti-phosphorylated (p)-NF-κB p65 (1:1,000; Cell Signaling
Technology, Inc.; cat. no. 3033), rabbit anti-NF-κB p65 (1:1,000;
Cell Signaling Technology, Inc.; cat. no. 242), rabbit anti-p-NF-κB
inhibitor α (p-IκBα; 1:1,000; Cell Signaling Technology, Inc.; cat.
no. 5209), rabbit anti-IκBα(1:1,000; Cell Signaling Technology,
Inc.; cat. no. 4812) and mouse anti-β-actin (1:1,000; Abcam; cat.
no. ab8226) overnight at 4°C. Subsequently, the membranes were
incubated with horseradish peroxidase (HRP)-conjugated secondary
antibodies (goat anti-rabbit IgG-HRP; 1:2,000; cat. no. sc-2004;
goat anti-mouse IgG-HRP; 1:2,000; cat. no. sc-2005; Santa Cruz
Biotechnology, Inc.) for 2 h at temperature. An ECL kit (GE
Healthcare) was then applied to detect the signals. The gray value
analysis of target band was analyzed by Image J software 1.48u
(National Institutes of Health). β-actin was used as a loading
control.
Statistical analysis
Data are presented as the mean ± SD from at least
three independent experiments. Student's t-test was used for
comparisons between two groups and ANOVA followed by Tukey's
post-hoc test was used when there were more than two groups to
calculate significance among the different groups. Statistical
analyses were performed using SPSS 17.0 Software (SPSS, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
FSTL-1 is upregulated in BMDMs after
S. pneumoniae injection
To investigate the functional role of FSTL-1 in
S. pneumoniae infection, the mRNA expression of FSTL1 in
BMDMs was determined following S. pneumoniae infection for
0, 4, 12, 24 and 36 h via RT-qPCR analysis. As shown in Fig. 1A, FSTL-1 mRNA expression was
significantly elevated in BMDMs after S. pneumoniae
injection. The production of the inflammatory cytokines IL-1β,
TNF-α and IL-6 was also increased after S. pneumoniae
injection as detected using ELISAs (Fig. 1B). FSTL-1, IL-1β, TNF-α and IL-6
levels were increased in a dose-dependent manner and exhibited a
significant difference at 24 h post-infection compared with 0 h.
Therefore, cells were collected 24 h after infection for further
study. The protein level of FSTL-1 in BMDMs was determined
following S. pneumoniae infection for 24 h. The results
indicated that FSTL-1 was significantly increased (Fig. 1C). These findings suggested that
FSTL-1 may play an important role in the inflammatory response
during S. pneumoniae infection.
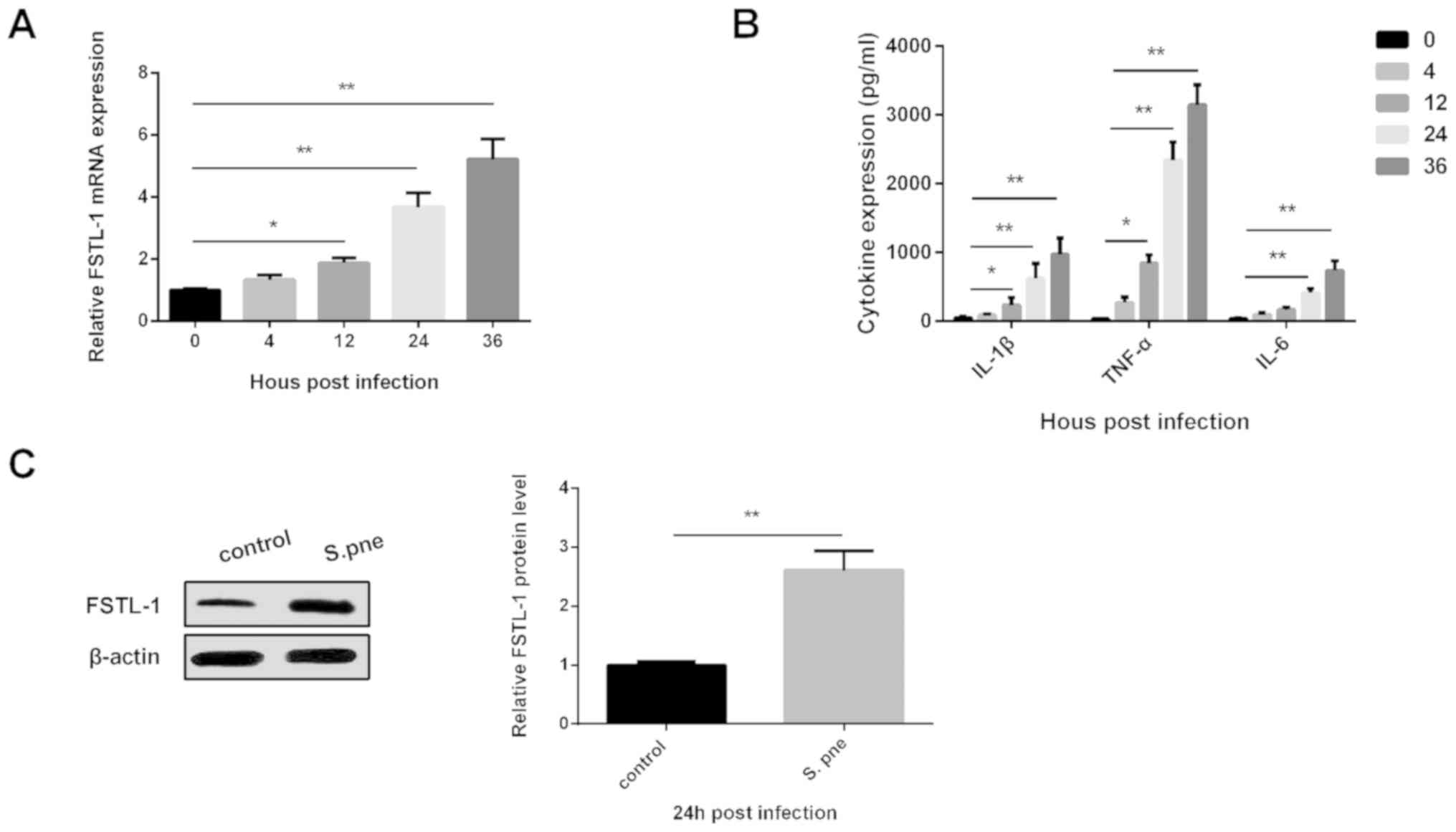 | Figure 1.Increased expression of FSTL-1 in
BMDMs infected with S. pneumoniae. (A) FSTL-1 mRNA
expression was determined in BMDMs following S. pneumoniae
infection for 0, 4, 12, 24 and 36 h via reverse
transcription-quantitative PCR analysis. (B) IL-1β, TNF-α and IL-6
expression levels were determined in BMDMs following S.
pneumoniae infection for 0, 4, 12, 24 and 36 h using ELISAs.
(C) FSTL-1 protein levels were determined in BMDMs following S.
pneumoniae infection for 24 h via western blotting. Similar
results were obtained from at least three independent experiments.
Data are expressed as the mean ± SD. *P<0.05, **P<0.01 vs. 0
h or control. BMDMs, bone marrow-derived macrophages; FSTL-1,
Follistatin-like protein 1; IL, interleukin; S.pne,
Streptococcus pneumoniae; TNF-α, tumor necrosis
factor-α. |
FSTL-1 modulates inflammatory injury
during S. pneumoniae infection via NLRP3 in vitro
To investigate the potential mechanism of FSTL-1
against inflammatory injury during S. pneumoniae infection
in BMDMs, FSTL-1 expression was increased or decreased using
FSTL-1-OE or FSTL-1-shRNA vectors, respectively, in BMDMs. FSTL-1
expression was significantly increased in the FSTL-1-OE group,
whereas it was decreased in the FSTL-1-shRNA group (Fig. 2A). Western blotting results
indicated that NLRP3 and cleaved caspase-1 levels were increased
significantly following the overexpression of FSTL-1, while they
were decreased after the depletion of FSTL-1 in BMDMs infected with
S. pneumoniae. However, ASC expression was not significantly
affected by changes in FSTL-1 expression (Fig. 2B). ELISA results showed that
increasing FSTL-1 levels enhanced, whereas decreasing FSTL-1
inhibited, the production of IL-1β, TNF-α and IL-6 (Fig. 2C-E). These data indicated that the
downregulation of FSTL-1 may attenuate inflammatory injury by
inhibiting NLRP3.
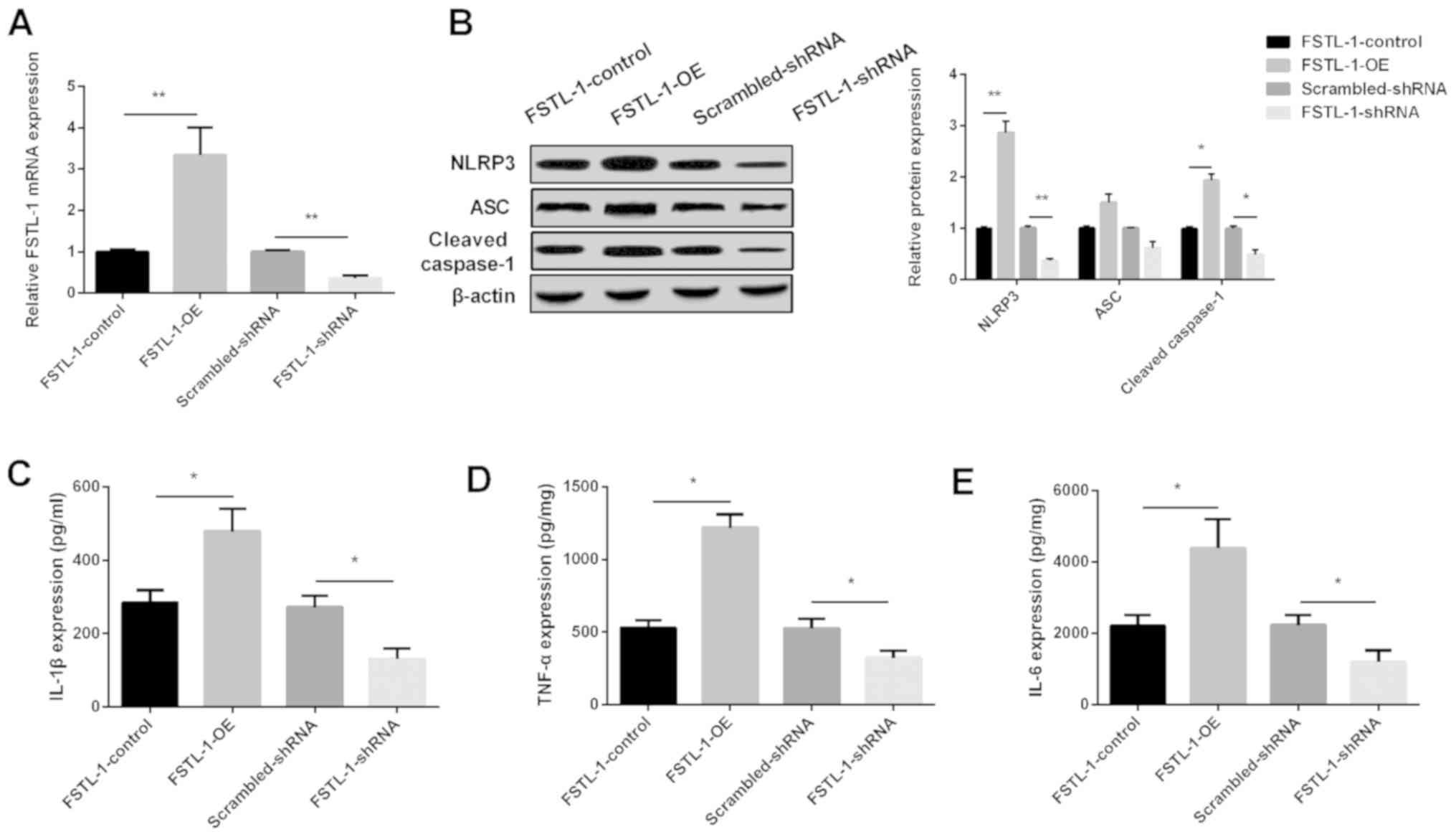 | Figure 2.FSTL-1 promotes the production of
inflammatory cytokines during S. pneumoniae infection in
BMDMs by regulating NLRP3. (A) Relative expression of FSTL-1
determined via reverse transcription-quantitative PCR analysis in
BMDMs after FSTL-1 OE or shRNA. (B) NLRP3, ASC and cleaved
caspase-1 protein levels were determined in BMDMs using western
blotting after FSTL-1 OE or shRNA. (C) IL-1β, (D) TNF-α and (E)
IL-6 expression levels were determined in BMDMs using ELISA after
FSTL-1 OE or shRNA. Similar results were obtained from at least
three independent experiments. Data are expressed as the mean ± SD.
*P<0.05, **P<0.01 vs. respective control. BMDMs, bone
marrow-derived macrophages; FSTL-1, Follistatin-like protein 1;
NLRP3, nucleotide oligomerization domain-like receptor protein 3;
OE, overexpression; shRNA, short hairpin RNA; ASC,
apoptosis-associated spot-like protein; IL, interleukin; S.
pneumoniae, Streptococcus pneumoniae; TNF-α, tumor necrosis
factor-α. |
Downregulation of NLRP3 inhibits
inflammatory injury during S. pneumoniae infection in vitro
To investigate the role of NLRP3 in inflammatory
injury during S. pneumoniae infection, NLRP3 levels were
determined in BMDMs. As shown in Fig.
3A, NLRP3 was significantly increased in BMDMs after S.
pneumoniae infection. NLRP3 mRNA and protein expression levels
were decreased significantly by NLRP3-shRNA, as shown in Fig. 3B and C. ELISA results showed that
decreasing NLRP3 expression inhibited the expression of IL-1β,
TNF-α and IL-6 in BMDMs after S. pneumoniae infection
(Fig. 3D-F). These data
demonstrated that the downregulation of NLRP3 inhibited
inflammatory injury during S. pneumoniae infection.
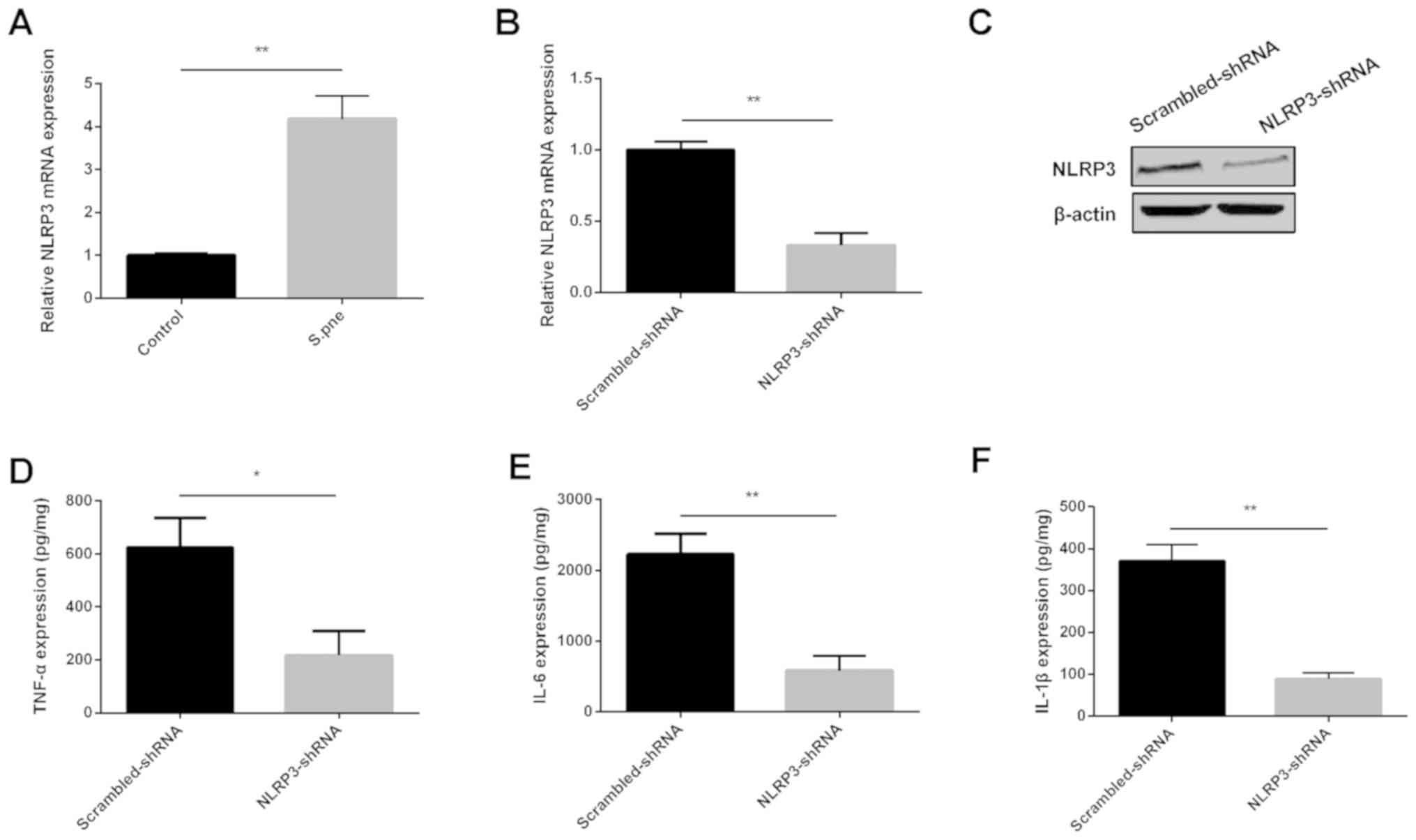 | Figure 3.Downregulation of NLRP3 inhibits the
production of inflammatory cytokines during S. pneumoniae
infection in BMDMs. (A) NLRP3 mRNA expression was determined in
BMDMs infected with S. pneumoniae via RT-qPCR analysis.
Relative (B) mRNA and (C) protein expression of NLRP3 as determined
using RT-qPCR or western blot analyses, respectively, in BMDMs
after depletion of NLRP3. (D) TNF-α, (E) IL-6, and (F) IL-1β
expression was determined in BMDMs using ELISAs after the depletion
of NLRP3. Similar results were obtained from at least three
independent experiments. Data are expressed as the mean ± SD.
*P<0.05, **P<0.01. BMDMs, bone marrow-derived macrophages;
NLRP3, nucleotide oligomerization domain-like receptor protein 3;
shRNA, short hairpin RNA; IL, interleukin; S.pne, Streptococcus
pneumoniae; TNF-α, tumor necrosis factor-α; RT-qPCR, reverse
transcription-quantitative PCR. |
TLR4/NF-κB signaling pathway is
involved in inflammatory injury during S. pneumoniae infection and
is regulated by FSTL-1 in vitro
Western blotting was used to determine the protein
level of TLR4 and the downstream targets of the NF-κB pathway. The
results showed that TLR4 expression was increased by FSTL-1-OE,
while it was decreased by FSTL-1-shRNA (Fig. 4A and B). The phosphorylation of
NF-κB p65 and IκBα was activated by FSTL-1-OE and was inactivated
by FSTL-1-shRNA. These data suggested that TLR4/NF-κB signaling was
involved in inflammatory injury during S. pneumoniae
infection regulated by FSTL-1 in vitro.
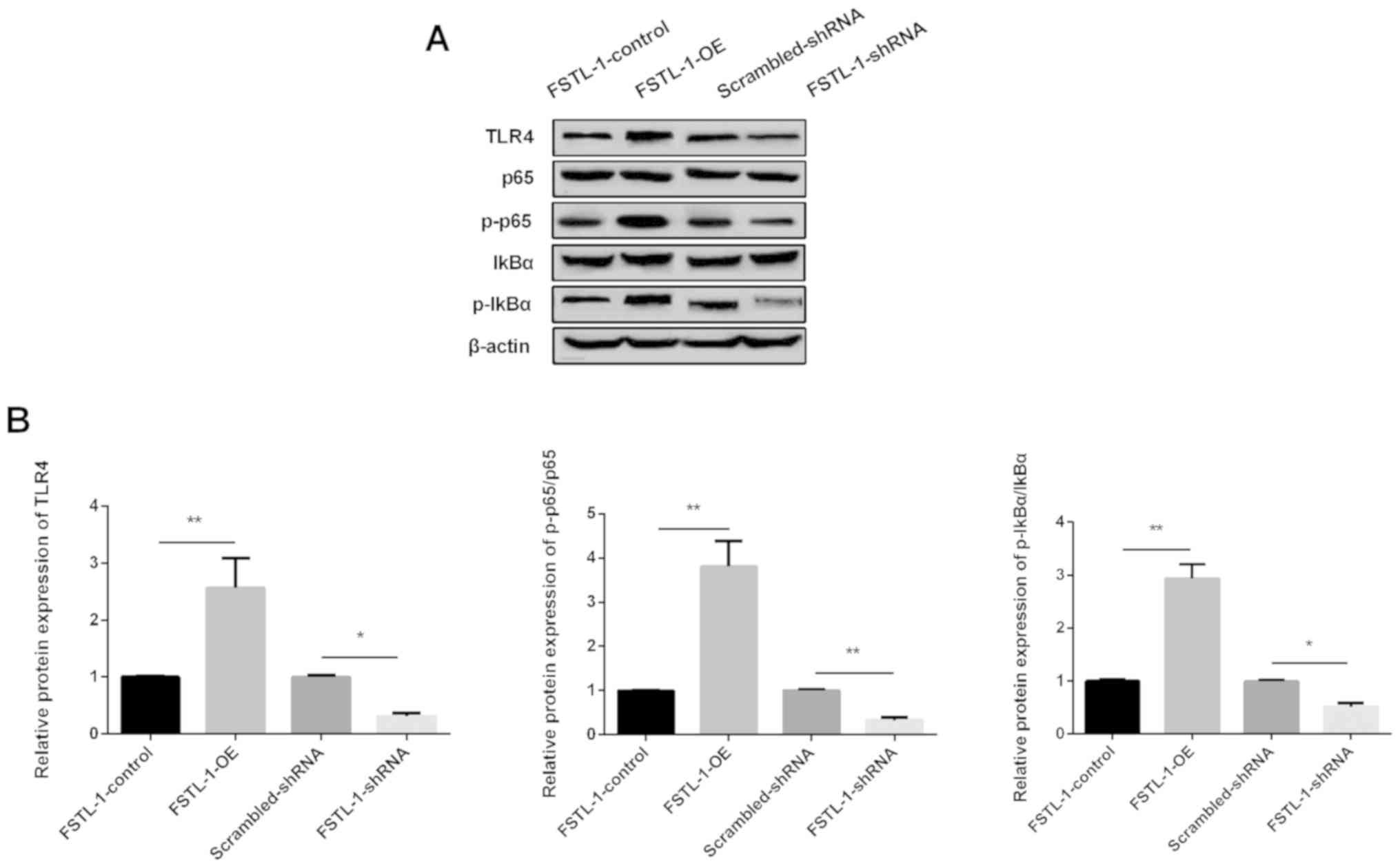 | Figure 4.TLR4/NF-κB signaling is involved in
inflammatory injury during S. pneumoniae infection and is
regulated by FSTL-1. (A) Levels of TLR4, NF-κB p65, p-p65, IκBα,
p-IκBα protein levels determined using western blotting in BMDMs
after transfection with FSTL-1-OE or FSTL-1-shRNA. (B) Relative
protein levels of TLR4, p-p65/p65, p-IκBα/IκBα were quantified in
BMDMs after transfection with FSTL-1-OE or FSTL-1-shRNA. Similar
results were obtained from at least three independent experiments.
Data are expressed as the mean ± SD. *P<0.05, **P<0.01 vs.
FSTL-1 control. BMDMs, bone marrow-derived macrophages; FSTL-1,
Follistatin-like protein 1; S.pne, Streptococcus pneumoniae;
TLR4, Toll-like receptor 4; OE, overexpression; shRNA, short
hairpin RNA; IκBα, NF-κB inhibitor α; p-, phosphorylated. |
Knockdown of FSTL-1 protects against
S. pneumoniae-induced inflammatory injury by inhibiting NLRP3 and
through the TLR4/NF-κB signaling pathway in vivo
The present study found that FSTL-1 was important
for inflammation in BMDMs infected with S. pneumoniae.
Therefore, the role of FSTL-1 during S. pneumoniae infection
in vivo was investigated. As shown in Fig. 5A-C, the production of IL-1β, IL-6
and TNF-α in BALFs was increased significantly after C57BL/6 mice
were infected with S. pneumoniae, and knockout of FSTL-1
decreased the expression of these factors. In addition, the mRNA
expression of FSTL-1 detected by RT-qPCR and the protein levels of
NLRP3, TLR4, p-p65 measured by western blotting in the lung tissues
of infected FSTL-1 KO mice were decreased compared with the
infected WT group (Fig. 5D). These
results indicated that the knockout of FSTL-1 might protect against
inflammatory injury induced by S. pneumoniae infection by
inhibiting the NLRP3 and TLR4/NF-κB signaling pathway in
vivo.
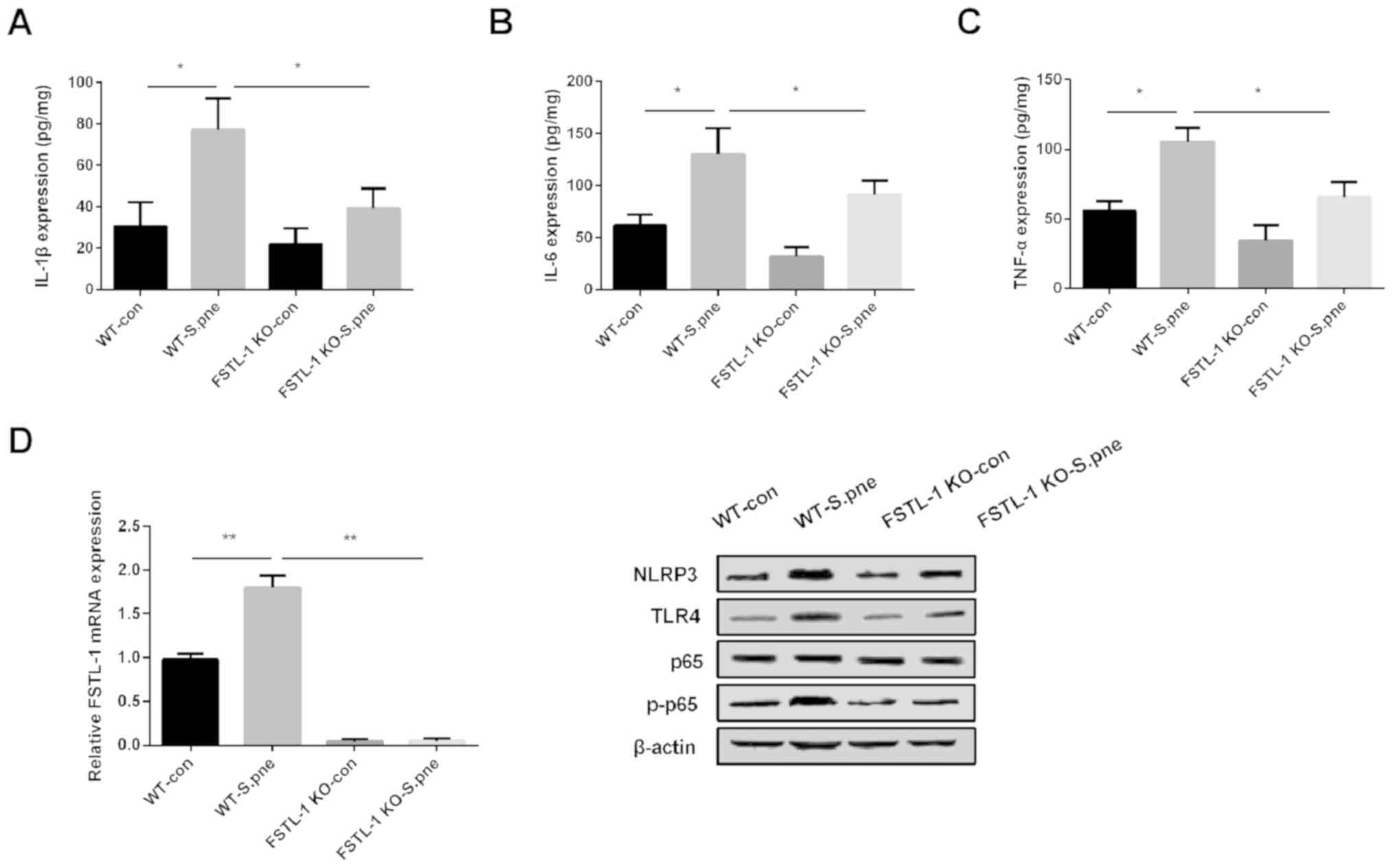 | Figure 5.Knockout of FSTL-1 inhibits the
inflammatory response against S. pneumoniae infection in
vivo by suppressing the NLRP3 and TLR4/NF-κB signaling pathway.
(A) IL-1β (B) IL-6, and (C) TNF-α expression levels were determined
in bronchoalveolar lavage fluids of FSTL-1 KO mice infected with S.
pne. (D) Protein levels of FSTL-1, NLRP3, TLR4, p-p65 were
determined in lung tissues of FSTL-1 KO mice infected with S. pne
(n=6). Similar results were obtained from at least three
independent experiments. Data are expressed as the mean ± SD.
*P<0.05; **P<0.01. FSTL-1, Follistatin-like protein 1; WT,
wild-type; KO, knockout; IL, interleukin; TNF-α, tumor necrosis
factor-α; S.pne, Streptococcus pneumoniae; con, control;
NLRP3, nucleotide oligomerization domain-like receptor protein 3;
TLR4, Toll-like receptor 4; p-, phosphorylated. |
Discussion
In the present study, it was found that the
infection of BMDMs with S. pneumoniae stimulated the
expression of FSTL-1. Downregulation of FSTL-1 protected against
inflammatory injury during S. pneumoniae infection by
inhibiting NLRP3. In addition, it was found that the TLR4/NF-κB
signaling pathway was involved in inflammatory injury regulated by
FSTL-1.
S. pneumoniae is a gram-positive bacterium
that is a common cause of pneumonia. S. pneumoniae-mediated
disease progression is significantly affected by the
proinflammatory response during acute infection (21–23).
Previous studies have shown that inflammatory cytokines were
increased in the lung tissues during S. pneumoniae
infection. For instance, Madouri et al (21) showed that IL-20, IL-1β and IL-6
cytokines were increased significantly during S. pneumoniae
infection. Additionally, Li et al (24) showed that the expression levels of
TNF-α, IL-6 and IL-1β in severe pneumonia induced by S.
pneumoniae were increased significantly in vitro and
in vivo. In the present study, it was found that the
production of TNF-α, IL-6 and IL-1β was significantly increased in
BMDMs infected with S. pneumoniae.
FSTL-1 plays an important role in the inflammatory
response. For instance, Campfield et al (25) showed that FSTL-1 modulated
IL-17-driven inflammation in stromal cells. Chaly et al
(13) suggested that FSTL-1 was
upregulated in patients with bacterial sepsis, and promoted the
secretion of NLRP3 and IL-1β from monocytes and macrophages. The
present study found that FSTL-1 was upregulated in BMDMs after
S. pneumoniae injection. The overexpression of FSTL-1
promoted, while the knockdown of FSTL-1 suppressed the production
of inflammatory factors. Moreover, it was also found that FSTL-1
positively modulated NLRP3. The present study suggested that the
reduced expression of FSTL-1 may attenuate inflammatory injury
during S. pneumoniae infection by inhibiting NLRP3.
NLRP3 was shown to be important in the inflammatory
response (26). A previous study
reported that in a meningitis model infected with S.
pneumoniae, NLRP3 promoted an increase in host pathology
(27). Stout-Delgado et al
(28) found that macrophages
infected with S. pneumoniae led to the development of
harmful NLRP3-mediated inflammation and pulmonary fibrosis.
Furthermore, excessive NLRP3 activation induced by S.
pneumoniae infection was reported to induce an increase in the
number of macrophages, which in turn increased morbidity (29). In the present study, it was found
that NLRP3 was upregulated in BMDMs after S. pneumoniae
injection and that depleting NLRP3 suppressed the production of
IL-1β, TNF-α and IL-6.
FSTL-1 has been reported to be induced by innate
immune signals, including TLR4 agonists (30). TLR-induced FSTL-1 expression
provides further understanding of the molecular mechanism of
FSTL-1. Several previous studies have reported that TLR signaling
may lead to the activation of NF-κB (31–33).
TLR4/NF-κB signaling was shown to be important for the immune
response. For example, TLRs play an important role in IL-1β
secretion and TLR4/NF-κB signaling promoted the expression of IL-1β
(34,35). In the present study, it was found
that FSTL-1 modulated the TLR4/NF-κB signaling pathway in
vitro.
In conclusion, the present study demonstrated that
the downregulation of FSTL-1 attenuated inflammation injury during
S. pneumoniae infection by regulating NLRP3 in vitro
and in vivo. The TLR4/NF-κB signaling pathway was found to
be involved in the inflammatory response during S.
pneumoniae infection and was regulated by FSTL-1. However,
there were limitations to the present study; for example, evidence
was not provided to show the relationship between FSTL-1 and NLRP3.
In the future, hematoxylin and eosin staining should be performed
in lung tissues to investigate the injury levels modulated by
FSTL-1, which could better demonstrate the effect of FSTL-1 on
inflammatory injury.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL was a major contributor to the writing the
manuscript and contributed to the conception of the study. LC
performed the data analyses and wrote the manuscript. All authors
read and approved the final study.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Xian Yang Central Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rendon A, Rendon-Ramirez EJ and
Rosas-Taraco AG: Relevant cytokines in the management of
community-acquired pneumonia. Curr Infect Dis Rep. 18:102016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Adler A, Baraniak A, Izdebski R, Fiett J,
Gniadkowski M, Hryniewicz W, Salvia A, Rossini A, Goossens H,
Malhotra S, et al: A binational cohort study of intestinal
colonization with extended-spectrum β-lactamase-producing proteus
mirabilis in patients admitted to rehabilitation centres. Clin
Microbiol Infect. 19:E51–E58. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Domon H, Nagai K, Maekawa T, Oda M,
Yonezawa D, Takeda W, Hiyoshi T, Tamura H, Yamaguchi M, Kawabata S
and Terao Y: Neutrophil elastase subverts the immune response by
cleaving toll-like receptors and cytokines in pneumococcal
pneumonia. Front Immunol. 9:7322018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baldo V, Cocchio S, Gallo T, Furlan P,
Romor P, Bertoncello C, Buja A and Baldovin T: Pneumococcal
conjugated vaccine reduces the high mortality for
community-acquired pneumonia in the elderly: An italian regional
experience. PLoS One. 11:e01666372016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weycker D, Farkouh RA, Strutton DR,
Edelsberg J, Shea KM and Pelton SI: Rates and costs of invasive
pneumococcal disease and pneumonia in persons with underlying
medical conditions. BMC Health Serv Res. 16:1822016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maus U, Rosseau S, Knies U, Seeger W and
Lohmeyer J: Expression of pro-inflammatory cytokines by flow-sorted
alveolar macrophages in severe pneumonia. Eur Respir J. 11:534–541.
1998.PubMed/NCBI
|
|
7
|
Perny M, Roccio M, Grandgirard D, Solyga
M, Senn P and Leib SL: The severity of infection determines the
localization of damage and extent of sensorineural hearing loss in
experimental pneumococcal meningitis. J Neurosci. 36:7740–7749.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Weichelt U, Cay R, Schmitz T, Strauss E,
Sifringer M, Buhrer C and Endesfelder S: Prevention of
hyperoxia-mediated pulmonary inflammation in neonatal rats by
caffeine. Eur Respir J. 41:966–973. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song C, Li H, Zhang Y and Yu J: Effects of
Pseudomonas aeruginosa and Streptococcus mitis mixed
infection on TLR4-mediated immune response in acute pneumonia mouse
model. BMC Microbiol. 17:822017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oshima Y, Ouchi N, Sato K, Izumiya Y,
Pimentel DR and Walsh K: Follistatin-like 1 is an Akt-regulated
cardioprotective factor that is secreted by the heart. Circulation.
117:3099–3108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wilson DC, Marinov AD, Blair HC, Bushnell
DS, Thompson SD, Chaly Y and Hirsch R: Follistatin-like protein 1
is a mesenchyme-derived inflammatory protein and may represent a
biomarker for systemic-onset juvenile rheumatoid arthritis.
Arthritis Rheum. 62:2510–2516. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Miyamae T, Marinov AD, Sowders D, Wilson
DC, Devlin J, Boudreau R, Robbins P and Hirsch R: Follistatin-like
protein-1 is a novel proinflammatory molecule. J Immunol.
177:4758–4762. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chaly Y, Fu Y, Marinov A, Hostager B, Yan
W, Campfield B, Kellum JA, Bushnell D, Wang Y, Vockley J and Hirsch
R: Follistatin-like protein 1 enhances NLRP3 inflammasome-mediated
IL-1β secretion from monocytes and macrophages. Eur J Immunol.
44:1467–1479. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Katsuyama E, Miyamoto H, Kobayashi T, Sato
Y, Hao W, Kanagawa H, Fujie A, Tando T, Watanabe R, Morita M, et
al: Interleukin-1 receptor-associated kinase-4 (IRAK4) promotes
inflammatory osteolysis by activating osteoclasts and inhibiting
formation of foreign body giant cells. J Biol Chem. 290:716–726.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dinarello CA: Immunological and
inflammatory functions of the interleukin-1 family. Annu Rev
Immunol. 27:519–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baroja-Mazo A, Martin-Sanchez F, Gomez AI,
Martinez CM, Amores-Iniesta J, Compan V, Barbera-Cremades M, Yague
J, Ruiz-Ortiz E, Anton J, et al: The NLRP3 inflammasome is released
as a particulate danger signal that amplifies the inflammatory
response. Nat Immunol. 15:738–748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Medzhitov R: Toll-like receptors and
innate immunity. Nat Rev Immunol. 1:135–145. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kawai T and Akira S: The role of
pattern-recognition receptors in innate immunity: Update on
Toll-like receptors. Nat Immunol. 11:373–384. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Inaba K, Inaba M, Romani N, Aya H, Deguchi
M, Ikehara S, Muramatsu S and Steinman RM: Generation of large
numbers of dendritic cells from mouse bone marrow cultures
supplemented with granulocyte/macrophage colony-stimulating factor.
J Exp Med. 176:1693–1702. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Madouri F, Barada O, Kervoaze G, Trottein
F, Pichavant M and Gosset P: Production of interleukin-20 cytokines
limits bacterial clearance and lung inflammation during infection
by Streptococcus pneumoniae. EBioMedicine. 37:417–427. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kato H, Yamagishi Y, Hagihara M, Yokoyama
Y, Suematsu H, Asai N, Koizumi Y and Mikamo H: Antimicrobial
activity of solithromycin and levofloxacin against a murine
pneumonia mixed-infection model caused by Streptococcus
pneumoniae and anaerobic bacteria. J Infect Chemother.
25:311–313. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsu CF, Hsiao CH, Tseng SF, Chen JR, Liao
YJ, Chen SJ, Lin CS, Sytwu HK and Chuang YP: PrtA immunization
fails to protect against pulmonary and invasive infection by
Streptococcus pneumoniae. Respir Res. 19:1872018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li H, Chen X and Zhou SJ: Dauricine
combined with clindamycin inhibits severe pneumonia co-infected by
influenza virus H5N1 and Streptococcus pneumoniae in vitro
and in vivo through NF-κB signaling pathway. J Pharmacol Sci.
137:12–19. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Campfield BT, Eddens T, Henkel M, Majewski
M, Horne W, Chaly Y, Gaffen SL, Hirsch R and Kolls JK:
Follistatin-like protein 1 modulates IL-17 signaling via IL-17RC
regulation in stromal cells. Immunol Cell Biol. 95:656–665. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McElvaney OJ, Zaslona Z, Becker-Flegler K,
Palsson-McDermott EM, Boland F, Gunaratnam C, Gulbins E, O'Neill
LA, Reeves EP and McElvaney NG: Specific inhibition of the NLRP3
inflammasome as an anti-inflammatory strategy in cystic fibrosis.
Am J Respir Crit Care Med. Aug 27–2019.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hoegen T, Tremel N, Klein M, Angele B,
Wagner H, Kirschning C, Pfister HW, Fontana A, Hammerschmidt S and
Koedel U: The NLRP3 inflammasome contributes to brain injury in
pneumococcal meningitis and is activated through ATP-dependent
lysosomal cathepsin B release. J Immunol. 187:5440–5451. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stout-Delgado HW, Cho SJ, Chu SG, Mitzel
DN, Villalba J, El-Chemaly S, Ryter SW, Choi AM and Rosas IO:
Age-dependent susceptibility to pulmonary fibrosis is associated
with NLRP3 inflammasome activation. Am J Respir Cell Mol Biol.
55:252–263. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moon JS, Nakahira K, Chung KP, DeNicola
GM, Koo MJ, Pabon MA, Rooney KT, Yoon JH, Ryter SW, Stout-Delgado H
and Choi AM: NOX4-dependent fatty acid oxidation promotes NLRP3
inflammasome activation in macrophages. Nat Med. 22:1002–1012.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Clutter SD, Wilson DC, Marinov AD and
Hirsch R: Follistatin-like protein 1 promotes arthritis by
up-regulating IFN-gamma. J Immunol. 182:234–239. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dong D, Zhou H, Na SY, Niedra R, Peng Y,
Wang H, Seed B and Zhou GL: GPR108, an NF-κB activator suppressed
by TIRAP, negatively regulates TLR-triggered immune responses. PLoS
One. 13:e02053032018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu L, Pang XL, Shang WJ, Xie HC, Wang JX
and Feng GW: Over-expressed microRNA-181a reduces glomerular
sclerosis and renal tubular epithelial injury in rats with chronic
kidney disease via down-regulation of the TLR/NF-κB pathway by
binding to CRY1. Mol Med. 24:492018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Niu W, Sun B, Li M, Cui J, Huang J and
Zhang L: TLR-4/microRNA-125a/NF-κB signaling modulates the immune
response to Mycobacterium tuberculosis infection. Cell Cycle.
17:1931–1945. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Becker CE and O'Neill LA: Inflammasomes in
inflammatory disorders: The role of TLRs and their interactions
with NLRs. Semin Immunopathol. 29:239–248. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bankers-Fulbright JL, Kalli KR and McKean
DJ: Interleukin-1 signal transduction. Life Sci. 59:61–83. 1996.
View Article : Google Scholar : PubMed/NCBI
|














