Introduction
Hyperoxia-induced acute lung injury (HALI) is a
typical complication of oxygen therapy, and can result in acute
respiratory distress syndrome (ARDS) (1). In HALI, the hyperoxia can cause
apoptosis and necrosis of alveolar epithelial cells (AEC) (2). In addition, apoptosis was associated
with hyperoxia toxicity, lung injury and a decrease in survival
rate in rats (3). Type II AEC (AEC
II) is the cell origin of AEC. AEC II are essential for maintaining
homeostatic pulmonary function under normal physiological
conditions. Hyperoxia, however, results in loss of epithelial
integrity in AEC II, by increasing apoptosis and possibly by
enhancing epithelial transdifferentiation (4,5).
Loss of AEC integrity then contributes to cytokine release and
enhanced immune cell recruitment under HALI conditions (6). HALI (7), ARDS (8), pulmonary fibrosis (9) and other oxidative stress-related lung
diseases are the predominant reasons for AEC-II apoptosis. AEC II
are the primary target cells of high oxygen (4,7), and
AEC II apoptosis is considered to be the underlying mechanism for
the pathogenesis of HALI (10,11).
The survival or apoptosis of AEC II directly affect the degree of
lung injury and repair. For example, transforming growth factor-β
receptor interacting protein-1 can reduce AEC apoptosis and
increase the resistance to HALI (2). It has previously been reported that
inhibiting apoptosis of AEC II effectively decreased HALI in rats
(12). Therefore, AEC II apoptosis
is a key contributor of HALI, inhibition of which may be an
effective way to decrease the incidence of HALI, and ultimately
provide a cure.
MicroRNAs (miRNAs) are a type of small non-coding
RNAs of 18–22 nucleotides that are widely expressed in tissues and
organs throughout the body. miRNAs are involved in cell growth,
proliferation, differentiation, apoptosis, metabolism and other
biological processes. miRNA-21 has been reported to be associated
with multiple pathological conditions, including tumor development
(13,14), cardiovascular (15–17),
liver (18), lung (19,20)
and renal (21) diseases, as well
as diabetes (22). Using
microarray methods, it was revealed that miR-21-5p was
differentially expressed in AEC II cells during hyperoxia, and in
specific in the H2O2-induced HALI model
(23). miR-21-5p overexpression in
AEC II significantly decreased apoptosis (23,24),
which may be attributed to inhibition of its target gene PTEN
(24). miR-21-5p overexpression in
the HALI lung effectively decreased HALI severity in rats (23,25).
These results suggest that miR-21-5p could alleviate AEC II
apoptosis in HALI and decrease lung damage, but its anti-apoptotic
mechanisms are not yet fully understood.
Previous studies (24,26)
have suggested that PTEN is one of the target genes of miR-21-5p.
PTEN/AKT signaling has been reported to inhibit cell cycle
progression, and promote apoptosis (27,28).
In addition, it has been widely reported that miR-21 reduces
apoptosis by targeting PTEN and inhibiting PTEN/AKT signaling
(17,29). The present study investigated the
association between miR-21-5p, PTEN, AKT and AEC II apoptosis, and
speculated that miR-21-5p may decrease AEC II apoptosis via PTEN
and AKT-associated mechanisms.
Materials and methods
Materials
AAV-6-miR-21-5p-GFP [miR-21-5p mimic
(UGUCGGGUAGCUUAUCAGACUGAUGUUGACUGUUGAAUCUCAUGGCAACACCAGUCGAUGGGCUGUCUGACA)
and green fluorescent protein (GP) encoded in an adeno-associated
virus (AAV) type 6 vector] and AAV-6-miR-21-5p negative control
(only GFP encoded in the AAV-6 vector) were from Synthgene
Biotechnology Co., Ltd. Primers used for reverse
transcription-quantitative PCR (RT-qPCR) experiments were from
Thermo Fisher Scientific, Inc. Fetal bovine serum (FBS), DMEM/F12
and DMEM-low glucose medium (Gibco; Thermo Fisher Scientific,
Inc.), Annexin V-FITC/propidium iodide (PI) cell apoptosis
detection kit (BD Biosciences), QuantiTect SYBR Green PCR kit
(Takara Bio, Inc.), HiScript® II 1st Strand cDNA
Synthesis kit (Vazyme), bicinchoninic acid (BCA) protein
quantification kit and SDS-PAGE gel preparation kit (Beyotime
Institute of Biotechnology) were utilized in the present study.
Antibodies targeting PTEN (cat. no. ab32199), AKT (cat. no.
ab8805), phosphorylated (p) AKT (cat. no. ab38449), and GAPDH (cat.
no. ab181602), and the secondary horseradish peroxidase
(HRP)-labeled antibody, were from Abcam. The unlisted reagents were
of analytical grade.
Instruments
The present study utilized a FACSCalibur flow
cytometer (BD Biosciences), an inverted fluorescence microscope
(Nikon Corporation), a light microscope (Leica Microsystems GmbH),
a fluorescence quantitative PCR instrument (Bio-Rad Laboratories,
Inc.), a full-wavelength fluorescent microplate reader (Thermo
Fisher Scientific, Inc.) and a transmission electron microscope
(TEM; Hitachi, Ltd.).
Animals and grouping
All experimental procedures were performed according
to the ‘Guide for the Care and Use of Laboratory Animals’ in China,
and approved by the Experimental Animal Care and Use Committee of
Zunyi Medical University. Sprague-Dawley rats (male:female, 1:1 in
each experiment; age, 7–8 weeks; weight, 200–220 g) were purchased
from the Third Military Medical University (Chongqing, China), and
housed under controlled conditions: Food and water supplied ad
libitum; a 12 h light/dark cycle; constant temperature (22°C)
and humidity (45–55%). The rats were randomly assigned into four
groups: i) Control group (n=20), exposed to room air; ii) HALI
group (n=20), where rats were exposed to 5.0 l/min pure oxygen for
48 h, performed in an air-tight plastic chamber to maintain the
concentration of oxygen (>95%), as previously described
(30); iii) miR-21-5p + HALI group
(n=20), where rats were sedated with 5% isoflurane and
AAV-6-miR-21-5p was endotracheally administered at an MOI of
106 to the lung through an endotracheal tube inserted
into the trachea, then 3 weeks later, the rats were exposed to pure
oxygen to induce HALI; and iv) miR-21-5p control + HALI group
(n=20), where a control AAV-6 was endotracheally administrated to
the lung through an endotracheal tube, then 3 weeks, the rats were
exposed to pure oxygen in order to induce HALI. No rats showed any
signs of adverse weight loss in the four groups, and therefore no
rats were excluded from the study.
Sample collection, oxygenation index
(OI) and respiratory index (RI) calculation
At 0, 24, 48 and 60 h after hyperoxia exposure, rats
from the four groups were sedated with 5% isoflurane. Blood (2 ml)
was collected from the carotid artery and subjected to arterial
blood gas analysis, and the OI and RI were calculated. The rats
were euthanized by exsanguination, and the left lung was collected
for the wet/dry weight analysis, then the rats were transcardially
perfused with 4% paraformaldehyde prior to collection of the right
lung for hematoxylin and eosin (H&E) staining and pathological
score calculation.
Detection of lung wet/dry weight
ratio
As previously reported (30), after euthanasia of the rats, the
lungs were surgically dissected away from the heart, trachea and
main bronchi, then placed into a bottle, weighed, and dried to a
constant weight in an oven at 70°C for 48 h. The wet/dry weight
ratio was calculated to assess lung edema and to quantify the lung
water content. Lung water content was calculated as: Lung water
content = [(wet weight-dry weight)/wet weight] ×100%.
Histological examination
As previously reported (30), the lungs collected for histological
analyses were fixed with 4% paraformaldehyde at room temperature
for 24 h, followed by embedment in paraffin to dehydration, then
sectioned (5 µm thickness). The sections were stained with
hematoxylin and 0.5% eosin for 5 and 3 min, respectively, in room
temperature (~22°C). Tissue sections were observed under a light
microscope. Lung injury was scored as previously described by
Matute-Bello et al (31).
The parameters used to analyze HALI severity were: i) Neutrophils
in alveoli; ii) neutrophils in pulmonary interstitium; iii)
transparent membrane; iv) areas filled with protein debris; and v)
thickness of alveolar septum.
AEC II isolation and culture
The isolation of AEC II was performed based on a
previously described method (24,32).
Following anesthesia of the rats, lungs were removed and blood and
leukocytes were replaced with PBS. Lungs were minced and digested
by 0.25% trypsin for 25 min at 37°C. The effects of trypsin were
neutralized with DMEM/F12 containing 10% FBS. The cell suspension
was sequentially filtered through 70 µm cell strainers and
centrifuged at 110 × g at room temperature for 5 min. The
supernatant was then removed, and the cell pellets were resuspended
in collagenase and incubated for 15 min at 37°C. Collagenase
activity was neutralized by the addition of the same medium
containing FBS and the cells were centrifuged again at 110 × g at
room temperature for 5 min. Cell pellets were resuspended and
cultured in flasks containing DMEM/F12 medium with 10% FBS in a
37°C, 95% air humidity, and 5% CO2 incubator.
AEC II cell identification by TEM
AEC II were identified by TEM as previously
described (23). AEC II were
incubated for 48 h and digested with 0.125% trypsin. The cell
suspension was collected and centrifuged at 100 × g for 10 min at
4°C. The supernatant was removed and the cells were fixed with 4%
glutaraldehyde at room temperature for 24 h. The cell pellet was
rinsed three times for 10 min at 4°C in PBS and fixed at 4°C for 30
min in 1% osmium tetroxide, then rinsed three times with PBS and
observed by TEM.
miR-21-5p and PTEN mRNA expression
level detection by RT-qPCR
At 0, 24, 48 and 60 h after isolation and culture,
cells from each group were washed twice with PBS, extracted with
500 µl TRIzol® (Thermo Fished Scientific, Inc.) at room
temperature for 5 min, and centrifuged for 5 min (15,000 × g, 4°C).
The supernatant was collected, 0.1 ml chloroform was added and
mixed at room temperature for 5 min, and the sample was centrifuged
again for 15 min (15,000 × g, 4°C). An equal volume of isopropanol
was added to the supernatant at room temperature for 10 min, then
centrifuged for 10 min (15,000 × g, 4°C). DEPC water was used to
dissolve the RNA pellet. The absorbance of the RNA solution was
measured at 260 and 280 nm (OD260 and OD280),
and the concentration of the RNA was calculated. As previously
described (24), the settings for
reverse transcription were as follows: 37°C for 15 min and 85°C for
5 sec. The cDNA solution was stored at −80°C. The qPCR settings for
miR-21-5p quantification were: 95°C for 5 min; 39 cycles of 95°C
for 45 sec, 60°C for 30 sec, 72°C for 45 sec; and 72°C for 10 min.
The Cq values of miR-21-5p and U6 (as an internal reference
control) were determined. The qPCR settings for PTEN quantification
were: 94°C for 5 min; 39 cycles of 94°C for 45 sec, 51°C for 30
sec, 72°C for 44 sec; and 72°C for 10 min. The qPCR settings for
β-actin quantification were: 94°C for 5 min; 39 cycles of 94°C for
45 sec, 60°C for 30 sec, 72°C for 44 sec; and 72°C for 10 min.
Relative expression levels were calculated using the
2−ΔΔCq method (33).
The primer sequences were as follows: miR-21-5p, forward
5′-GTCAATAGCTTATCAGACTGA-3′ and reverse
5′-GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCA-3′; U6, forward
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse
5′-CGCTTCACGAATTTGCGTGTCAT-3′; PTEN, forward
5′-TTTGAAGACCATAACCCACCAC-3′ and reverse
5′-ATTACACCAGTTCGTCCCTTTC-3′; and β-actin, forward
5′-TTCCTCCGCAAGGATGACACGC-3′ and reverse
5′-GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCACCAGAGCCAACAAAAATAT-3′.
PTEN, pAKT and AKT protein expression
detection via western blotting
Western blotting experiments were performed
according to previously described standard procedures (34,35).
AEC II were lysed with 400 µl lysis buffer (P0013, Beyotime
Institute of Biotechnology) containing 100 mmol/l PMSF (ST506,
Beyotime Institute of Biotechnology) for 30 min. Proteins were
obtained and stored at −80°C. The protein concentration was
determined using the BCA method and separated via SDS-PAGE (10%
gel). Proteins were transferred to PVDF membrane and blocked with
5% skimmed milk at room temperature for 1 h. The membranes were
then incubated with rabbit anti-PTEN (1:2,000), rabbit anti-pan-AKT
(1:500) and rabbit anti-pAKT (T308; 1:1,000) primary antibodies
overnight at 4°C, followed by goat anti-rabbit secondary antibody
(1:5,000) at room temperature for 1 h. Enhanced chemiluminescence
was used to detect the results. The strip optical density values
were analyzed using a gel image analysis software (Image-Pro Plus;
version 6.0; Meyer Instruments, Inc.).
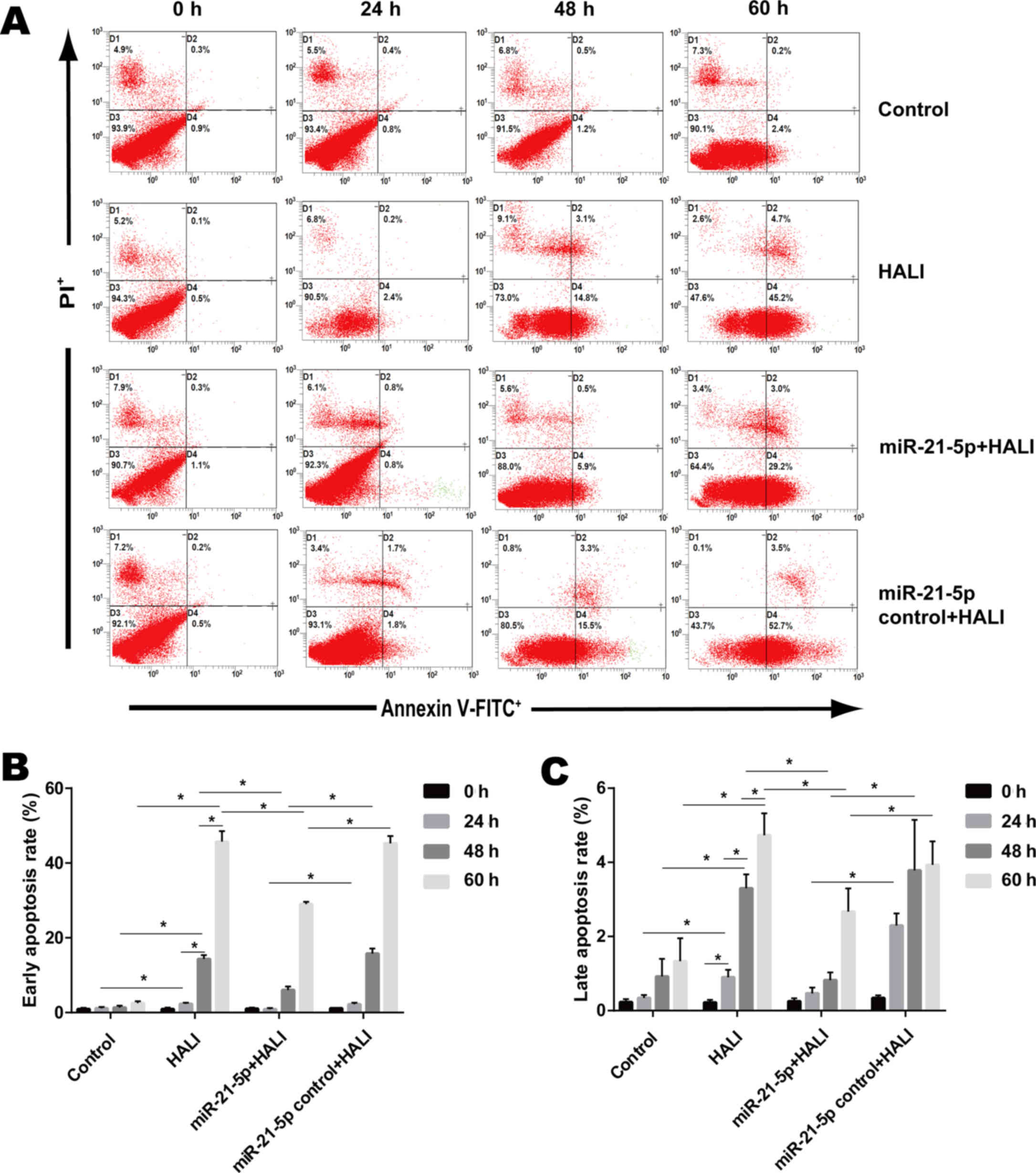
Apoptosis detection with flow
cytometry
Flow cytometry with Annexin V-FITC/PI staining was
used to evaluate early and late apoptosis of AEC II cells (17). Using a FACSCalibur flow cytometer,
the apoptosis rates of AEC II cells were detected.
AEC II cells were isolated from the lungs of rats in
each of the four groups. Excluding the 0 h time point groups, AEC
II cells were seeded in plates and cultured from each time point.
On the day of detection, cells were digested with 1 ml 0.125%
trypsin/EDTA for 2 min. Digested cells were centrifuged and the
supernatant was removed. Cell density was adjusted to
2×105 cells/ml, and 100 µl cell suspensions were
cultured for 24 h. A cell suspension for analysis was then prepared
at 0, 24, 48 and 60 h after isolation and culture. The cells were
collected via centrifugation (100 × g, 5 min), then washed twice
with PBS (100 × g, 5 min), and 500 µl binding buffer was added. A
total of 5 µl Annexin V-FITC and 5 µl PI were added and mixed at
room temperature in the dark for 5–15 min. Apoptosis was detected
via flow cytometry 1 h later.
Statistical analysis
SPSS software (version 23.0; IBM, Corp.) was used
for the statistical analysis. Data were expressed as the mean ±
standard deviation. One-way ANOVA and post hoc Dunnett's T3 test
were performed in order to compare the differences among and
between groups, respectively. P<0.05 was considered to indicate
a statistically significant difference.
Results
AAV-6-miR-21-5p administration in lung
decreases HALI in rats
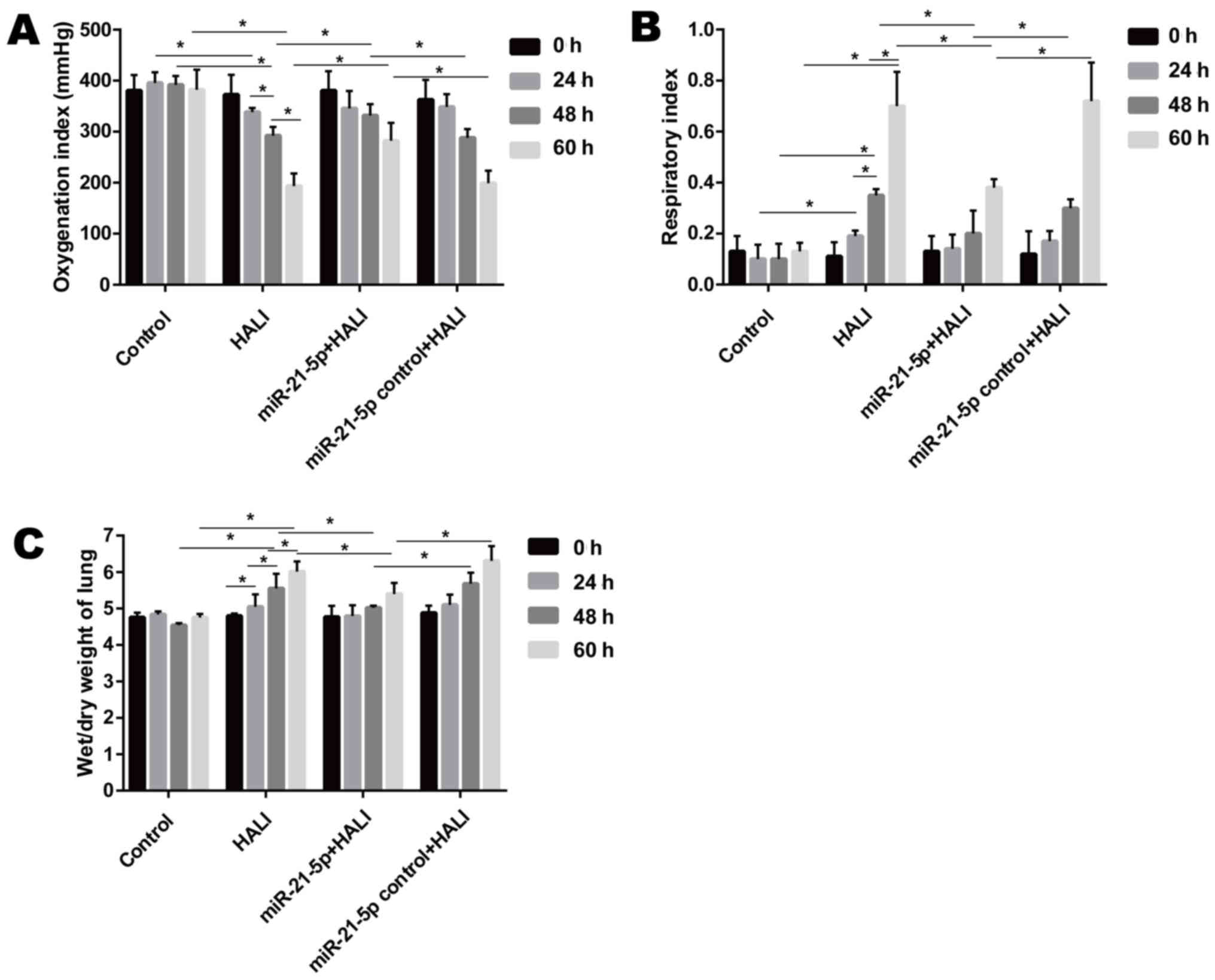
When rats were exposed to >95% oxygen, the OI
decreased significantly after hyperoxia exposure, while the RI and
wet/dry weight ratio of the lung increased significantly from 24 to
60 h after hyperoxia exposure (Fig.
1). Endotracheally administered AAV-6-miR-21-5p to the lung
reversed these changes induced by HALI, as evidenced by increased
OI and decreased RI and wet/dry weight ratio, compared with that of
the miR-21-5p control + HALI group (Fig. 1).
AAV-6-miR-21-5p administration in the
lung decreases pathological changes in the lung tissue of HALI
rats
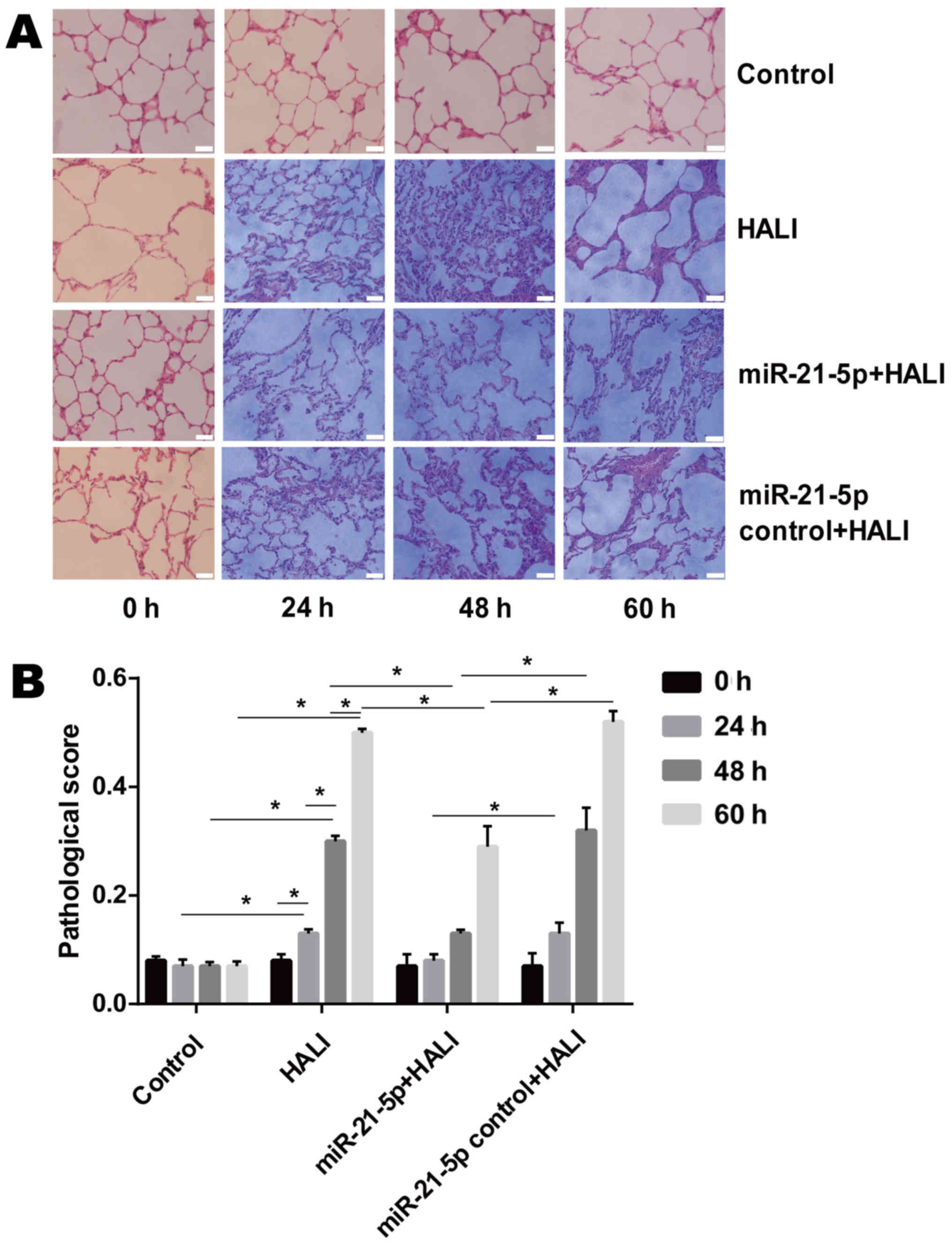
The microscopic results of H&E-stained lung
tissue sections revealed that neutrophils were rare in the alveolar
pulmonary interstitium in the group control (Fig. 2A). In the HALI and the miR-21-5p
control + HALI group, neutrophils were occasionally observed and
were gradually increased in alveolar pulmonary interstitium at 60 h
after hyperoxia (Fig. 2A). In
addition, protein fragments and occasional hyaline membrane were
observed in the alveolar interstitium at 48 and 60 h after
hyperoxia (Fig. 2A), which
indicated that HALI rats possessed severe lung injury. In the
miR-21-5p + HALI group, the frequency of neutrophil occurrence,
protein fragments and hyaline membrane decreased compared with the
HALI or miR-21-5p control + HALI groups (Fig. 2A). Endotracheally administered
AAV-6-miR-21-5p to the lung significantly decreased pathological
changes in the lung of HALI rats, as evidenced by the decreased
pathological score (Fig. 2B).
HALI and AAV-6-miR-21-5p
administration in the lung induces miR-21-5p upregulation
After 24 h culture, the AEC II cells revealed
adherent, island-like and spindle-like growth. After 48 h culture,
AEC II cells were polygonal and up to 95% adherent (Fig. 3A). The presence of osmiophilic
lamellar corpuscles and microvilli, both of which are
characteristic structures of AEC II, was confirmed by TEM (Fig. 3C). RT-qPCR results demonstrated
that both HALI and AAV-6-miR-21-5p administration in the lung
induced miR-21-5p upregulation (Fig.
3D). However, AAV-6-miR-1-5p administration significantly
increased miR-21-5p expression compared with the HALI group
(Fig. 3D).
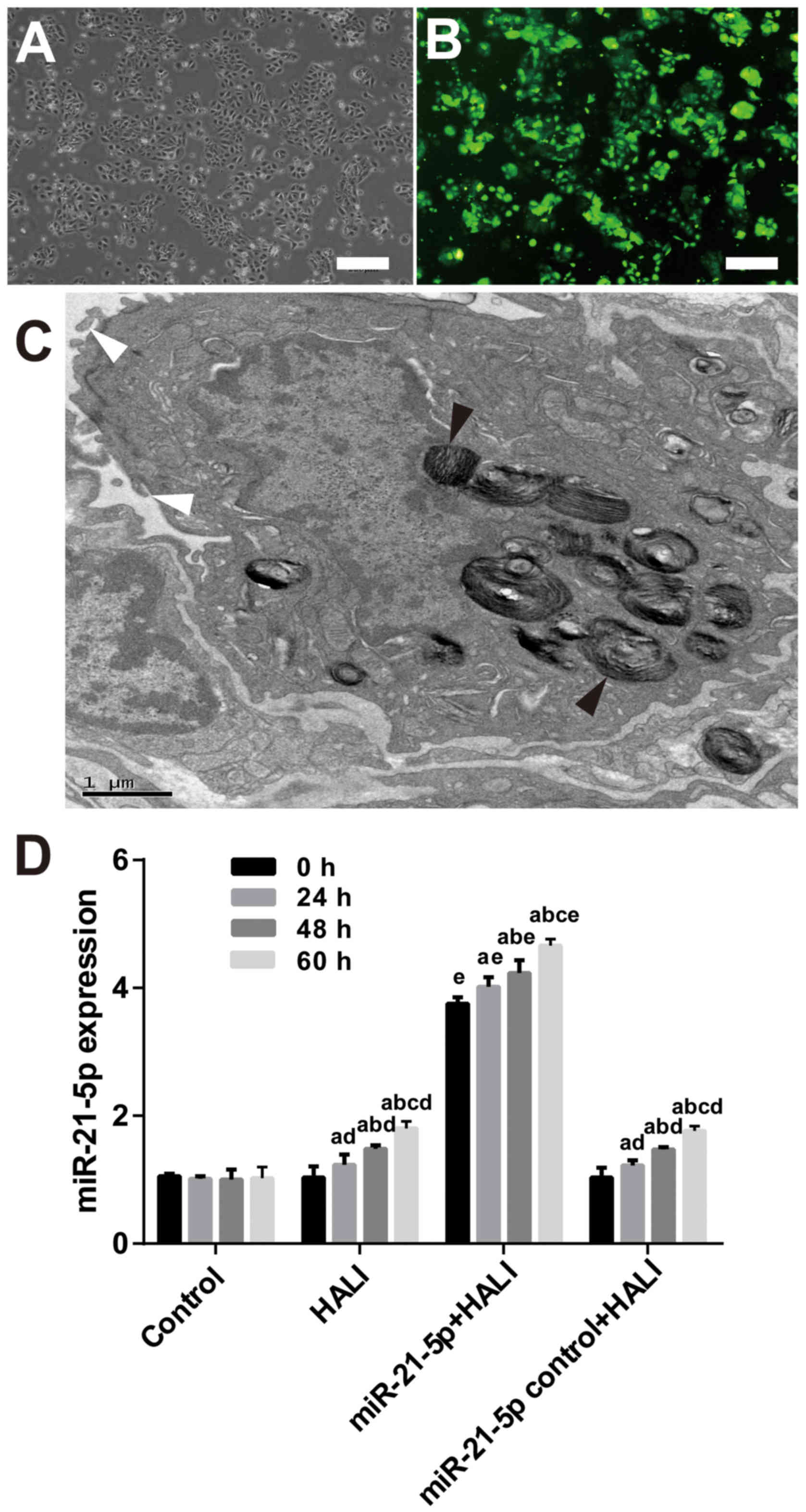 | Figure 3.AAV-6-miR-21-5p-GFP administration in
the lung induces miR-21-5p upregulation in AEC II cells. AAV-6
virus was dripped into the lungs of rats through tracheal
intubation, and 3 weeks later, HALI was induced. AEC II were
isolated from lungs at the indicated times after HALI and cultured
for 24 h. (A) Representative image of phase-contrast microscopy
showing the cell morphology. Scale bar, 250 µm. (B) Representative
image from fluorescence microscopy showing successful expression of
the GFP tag in the AAV vector. Scale bar, 250 µm. (C) The isolated
AEC II cells were confirmed by transmission electron microscopy.
The black arrowheads indicate osmiophilic lamellar body and the
white arrowheads indicate microvilli, both of which are
characteristic structures of AEC II. Scale bar, 1 µm. (D) Reverse
transcription-quantitative PCR analysis of the miR-21-5p levels in
the AECII cells isolated from the experimental rats. Data are
expressed as the mean ± standard deviation (n=6 rats per group).
aP<0.05 vs. 0 h; bP<0.05 vs. 24 h;
cP<0.05 vs. 48 h; dP<0.05 vs. the same
time point in the control group; eP<0.05 vs. the same
time point in the HALI group. AAV, adeno-associated virus; miR,
microRNA; GFP, green fluorescent protein; AEC II, type II alveolar
epithelial cells; HALI, hyperoxic acute lung injury. |
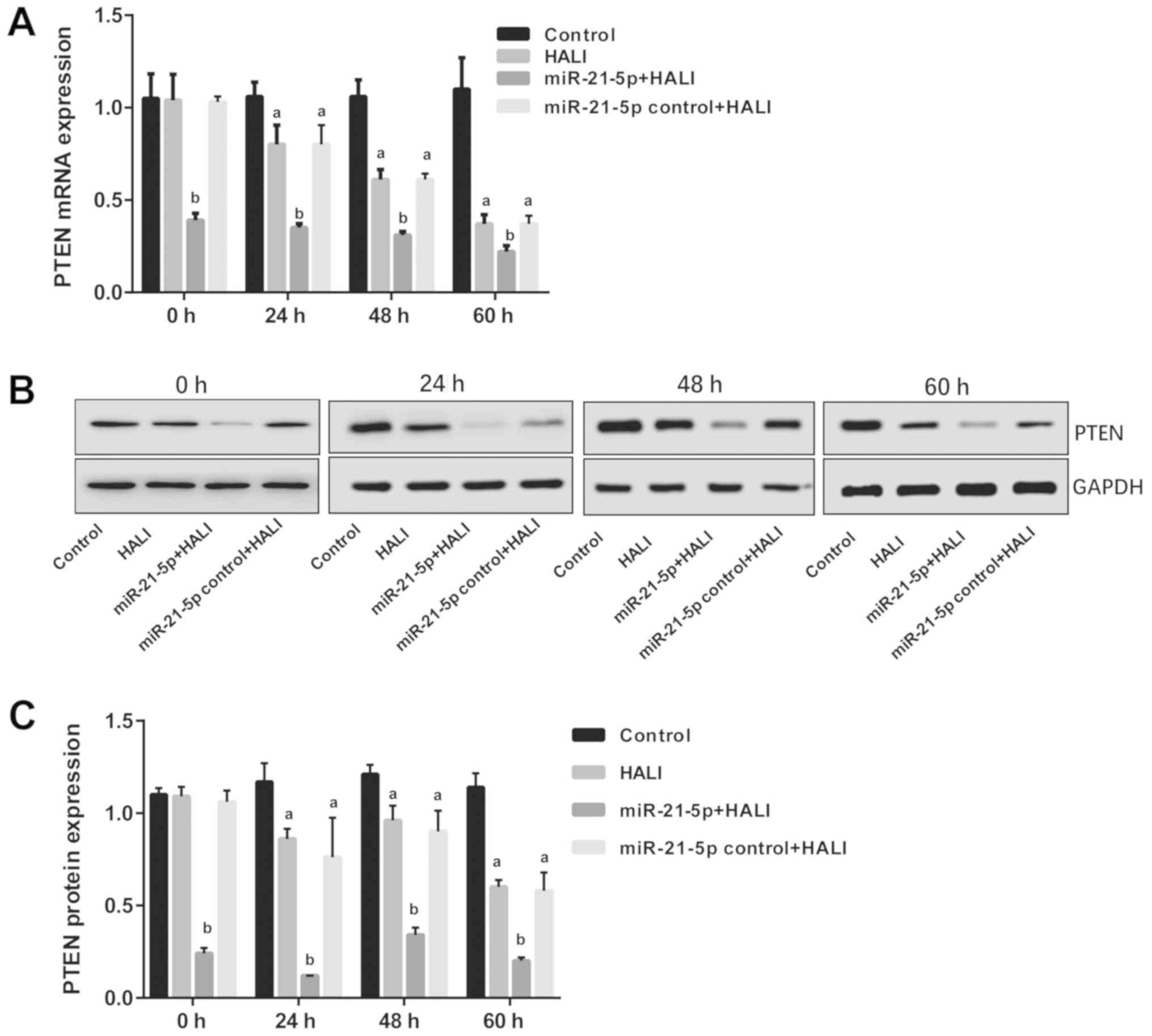
miR-21-5p overexpression decreases
PTEN mRNA and protein expression in AEC II cells isolated from the
HALI lung
The expression levels of PTEN were detected via
RT-qPCR and western blotting following isolation and culture of AEC
II from HALI rats. The results revealed that both mRNA and protein
PTEN expression were decreased in AEC II from the HALI rats
(Fig. 4). However, in the
miR-21-5p + HALI group, PTEN expression levels were significantly
decreased compared with the HALI group, while the expression of
PTEN in AEC II from rats receiving the negative control virus did
not exhibit any change compared with the HALI rats (Fig. 4).
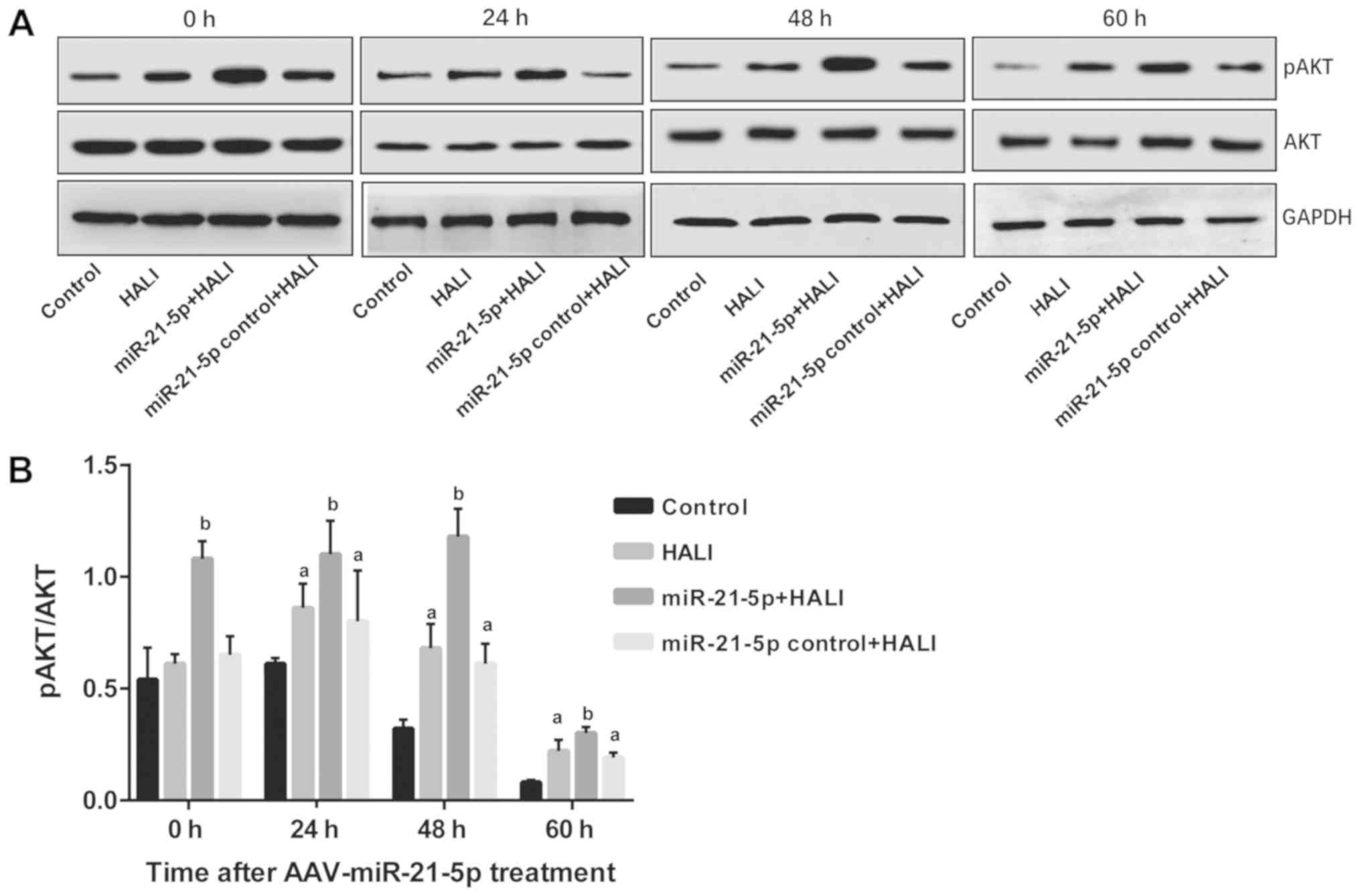
miR-21-5p overexpression increases AKT
phosphorylation in AEC II from the HALI lung
The protein expression levels of AKT and pAKT in AEC
II from HALI rats were detected via western blotting following
different culture times. pAKT/AKT ratios were calculated, and the
results revealed that in the control group, pAKT/AKT demonstrated
no difference between 0 and 24 h; however, this ratio decreased
significantly at 48 and 60 h compared with the 0 and 24 h time
points (P<0.05; Fig. 5). In
HALI and miR-21-5p control + HALI groups, pAKT/AKT ratio was
increased at 24 h of hyperoxia compared with their 0 h time points
(Fig. 5). Of note, the miR-21-5p +
HALI group exhibited the highest levels of p-AKT/AKT ratio among
the four groups, which was significantly higher compared with that
of group HALI and group miR-21-5p control + HALI (Fig. 5).
miR-21-5p overexpression decreases the
apoptosis rate of AEC II from the HALI lung
Flow cytometry (Fig.
6A) was employed in order to detect the apoptosis rate of AEC
II cells isolated from the HALI lung that were cultured for
different times. Control AEC II cells isolated from the normal lung
exhibited low early and late apoptosis rates, 0.93–2.53 and
0.23–1.33% respectively (Fig. 6B and
C). Cells isolated from the HALI lung exhibited gradually
increased early and late apoptosis after 24, 48 or 60 h culture
(Fig. 6B and C). AAV-miR-21-5p
administrated to the lung decreased the early and late apoptosis
rate of AEC II cells isolated from HALI lung compared with that of
the HALI group and the miR-21-5p control + HALI group (Fig. 6B and C).
Discussion
It has previously been reported that miR-21-5p
expression decreased in AEC II cells when cells underwent
H2O2 insult (23), but the specific underlying
molecular mechanism is not yet fully understood. In the present
study, rats were endotracheally administered AAV-6-miR-21-5p to the
lung before a HALI model was established in these rats. miR-21-5p
overexpression ameliorated HALI and decreased apoptosis of AEC II
cells isolated from HALI rats. Molecular experiments demonstrated
that miR-21-5p overexpression in the lung decreased PTEN mRNA and
protein expression levels in AEC II cells isolated from HALI rats.
In addition, the pAKT/AKT ratio increased following miR-21-5p
intervention in lung of HALI rats. These results indicate that
miR-21-5p targeted PTEN and decreased HALI via apoptosis-associated
mechanisms.
The present study isolated the AEC II cells from the
modeled rats and molecular changes in the isolated cells were
investigated ex vivo; therefore, it can be concluded that
the PTEN, AKT, pAKT expression and apoptosis results obtained from
the HALI rats were due to the effects on the AEC II cells. The data
from the present study demonstrated that miR-21-5p overexpression,
as a result of administering AAV-6 carrying an miR-21-5p-expression
vector, decreased the severity of HALI in vivo, and
decreased the apoptosis rate of ACE II cells ex vivo.
Further experiments revealed that PTEN mRNA and protein expression
levels were negatively regulated by the expression of miR-21-5p. By
using dual-luciferase reporter gene assay, it has previously been
demonstrated that PTEN is one of the target genes of miR-21-5p
(24,36,37).
Previous studies have revealed that miR-21 regulates cell
proliferation and apoptosis in different disease models. For
instance, in human hepatocellular carcinoma cells, downregulation
of miR-21 increased the expression of PTEN, and decrease the
proliferation, invasion and metastasis of hepatocarcinoma cells
(38,39). miR-21 overexpression inhibited the
apoptosis of nasopharyngeal carcinoma cells, and dual-luciferase
reporter assay revealed that miR-21-5p targeted the 3′ untranslated
region of the PTEN mRNA (36).
miR-21-5p from mesenchymal stem cell-derived exosomes also
inhibited hypoxia/reoxygenation-induced lung microvascular
endothelial cell apoptosis in vitro, and decreased lung
ischemia/reperfusion injury in vivo (40). Although no agonist/inhibitor of
PTEN or AKT were used in the present study, our group has
previously used the PTEN inhibitor Phen in an in vitro HALI
model and found that PTEN inhibition partially offset
H2O2-induced AECII apoptosis (24). In a previous study of
H2O-induced apoptosis in cardiac stem cells, it was
found that the PTEN inhibitor Phen or the PI3K inhibitor LY294002
efficiently attenuated the antiapoptotic effect of miR-21/PTEN/AKT
signaling (17).
A number of studies have reported that the PTEN/AKT
axis is associated with apoptosis (41–43).
In cancer research, PTEN is known to inhibit the growth of cancer
cells and lead to apoptosis (44).
HALI induces a pathological condition in lung tissues, and PTEN
overexpression in HALI AEC II may accelerate cell apoptosis,
although no study has reported that miR-21-5p decreases ACE II
apoptosis through PTEN/AKT. miR-21-5p overexpression promotes
non-small cell lung cancer cell growth and mobility through the
PTEN-AKT axis (45). miR-21-5p
overexpression also enhanced the invasion and sphere forming
abilities of keratinocytes through PTEN/AKT signaling (46). Finally, it was previously
demonstrated that miR-21-5p decreased ACE II apoptosis through the
PTEN/AKT pathway in vitro using an
H2O2-induced HALI model (24). The results from the present study
confirmed that miR-21-5p overexpression ameliorated HALI in
vivo via PTEN/AKT-associated antiapoptotic pathways.
Although miRNA-21-associated therapies have not been
applied to clinical use, it is believed that gene therapy is an
effective strategy for ARDS/acute lung injury (ALI) treatment
(47). In the present study,
miR-21-5p was successfully administered to the lung through the
trachea. In fact, cells in the lung, such as alveolar epithelial
cells, can be specifically targeted through endotracheal intubation
management (48), or through
antibodies, pulmonary endothelial cell surface antigen and
selective targeting intravenous injection (49). These data indicated the operability
of miRNA-based therapy in ARDS/ALI. Although studies regarding the
protective effects of miR-21-5p in pulmonary diseases are still in
the laboratory research phase, it can be speculated that the
elucidation of the antiapoptotic mechanism of miR-21-5p may bring
opportunities for the application of miR-21-5p-associated therapies
to the clinic.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 8156010205).
Availability of data and materials
The datasets analyzed/generated during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
SQ, HW, HM and GL performed the experiments, wrote
the article and prepared figures. MC conceived and designed the
experiments, and provided the reagents, materials and analysis
tools. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental procedures were performed according
to the Guide for the Care and Use of Laboratory Animals. The
protocol of the present study was approved by the Experimental
Animal Care and Use Committee of Zunyi Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dos Santos CC: Hyperoxic acute lung injury
and ventilator-induced/associated lung injury: New insights into
intracellular signaling pathways. Crit Care. 11:1262007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nyp MF, Mabry SM, Navarro A, Menden H,
Perez RE, Sampath V and Ekekezie II: Lung epithelial-specific
TRIP-1 overexpression maintains epithelial integrity during
hyperoxia exposure. Physiol Rep. 6:e135852018. View Article : Google Scholar :
|
|
3
|
Budinger GR, Mutlu GM, Urich D, Soberanes
S, Buccellato LJ, Hawkins K, Chiarella SE, Radigan KA, Eisenbart J,
Agrawal H, et al: Epithelial cell death is an important contributor
to oxidant-mediated acute lung injury. Am J Respir Crit Care Med.
183:1043–1054. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee HS and Kim CK: Cathepsin B is
activated as an executive protease in fetal rat alveolar type II
cells exposed to hyperoxia. Exp Mol Med. 43:223–229. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kalluri R and Neilson EG:
Epithelial-mesenchymal transition and its implications for
fibrosis. J Clin Invest. 112:1776–1784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barazzone C, Horowitz S, Donati YR,
Rodriguez I and Piguet PF: Oxygen toxicity in mouse lung: Pathways
to cell death. Am J Respir Cell Mol Biol. 19:573–581. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee HS and Kim CK: Effect of recombinant
IL-10 on cultured fetal rat alveolar type II cells exposed to
65%-hyperoxia. Respir Res. 12:682011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma X, Xu D, Ai Y, Ming G and Zhao S: Fas
inhibition attenuates lipopolysaccharide-induced apoptosis and
cytokine release of rat type II alveolar epithelial cells. Mol Biol
Rep. 37:3051–3056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kamp DW, Liu G, Cheresh P, Kim SJ, Mueller
A, Lam AP, Trejo H, Williams D, Tulasiram S, Baker M, et al:
Asbestos-induced alveolar epithelial cell apoptosis. The role of
endoplasmic reticulum stress response. Am J Respir Cell Mol Biol.
49:892–901. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang X, Wei SQ, Lee SJ, Fung JK, Zhang M,
Tanaka A, Choi AM and Jin Y: P62 sequestosome 1/light Chain 3b
complex confers cytoprotection on lung epithelial cells after
hyperoxia. Am J Respir Cell Mol Biol. 48:489–496. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Husari AW, Khayat A, Awdeh H, Hatoum H,
Nasser M, Mroueh SM, Zaatari G, El-Sabban M and Dbaibo GS:
Activated protein C attenuates acute lung injury and apoptosis in a
hyperoxic animal model. Shock. 33:467–472. 2010.PubMed/NCBI
|
|
13
|
Xu G, Zhang Y, Wei J, Jia W, Ge Z, Zhang Z
and Liu X: MicroRNA-21 promotes hepatocellular carcinoma HepG2 cell
proliferation through repression of mitogen-activated protein
kinase-kinase 3. BMC Cancer. 13:4692013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Teng Y, Radde BN, Litchfield LM, Ivanova
MM, Prough RA, Clark BJ, Doll MA, Hein DW and Klinge CM:
Dehydroepiandrosterone activation of G-protein-coupled estrogen
receptor rapidly stimulates MicroRNA-21 transcription in human
hepatocellular carcinoma cells. J Biol Chem. 290:15799–15811. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong S, Ma W, Hao B, Hu F, Yan L, Yan X,
Wang Y, Chen Z and Wang Z: MicroRNA-21 promotes cardiac fibrosis
and development of heart failure with preserved left ventricular
ejection fraction by up-regulating Bcl-2. Int J Clin Exp Pathol.
7:565–574. 2014.PubMed/NCBI
|
|
16
|
Richart A, Loyer X, Neri T, Howangyin K,
Guérin CL, Ngkelo A, Bakker W, Zlatanova I, Rouanet M, Vilar J, et
al: MicroRNA-21 coordinates human multipotent cardiovascular
progenitors therapeutic potential. Stem Cells. 32:2908–2922. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng W, Wang Y, Long X, Zhao R, Wang Z,
Liu Z, Cao S and Shi B: miR-21 reduces hydrogen peroxide-induced
apoptosis in c-kit+ cardiac stem cells in vitro through
PTEN/PI3K/Akt signaling. Oxid Med Cell Longev. 2016:53891812016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Otsuka M, Kishikawa T, Yoshikawa T, Ohno
M, Takata A, Shibata C and Koike K: The role of microRNAs in
hepatocarcinogenesis: Current knowledge and future prospects. J
Gastroenterol. 49:173–184. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yehya N, Yerrapureddy A, Tobias J and
Margulies SS: MicroRNA modulate alveolar epithelial response to
cyclic stretch. BMC Genomics. 13:1542012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li X, Mei ZJ, Wang YC, Chen J, Sun SX, Yau
Y, Li Z and Xie C: Antagonism of miR-21 reverses radiation-induced
EMT in alveolar epithelial cells via PI3K/Akt pathway. Int J Clin
Exp Pathol. 9:1158–1167. 2016.
|
|
21
|
Gomez IG, MacKenna DA, Johnson BG, Kaimal
V, Roach AM, Ren S, Nakagawa N, Xin C, Newitt R, Pandya S, et al:
Anti-microRNA-21 oligonucleotides prevent Alport nephropathy
progression by stimulating metabolic pathways. J Clin Invest.
125:141–156. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong L, Wang X, Tan J, Li H, Qian W, Chen
J, Chen Q, Wang J, Xu W, Tao C and Wang S: Decreased expression of
microRNA-21 correlates with the imbalance of Th17 and Treg cells in
patients with rheumatoid arthritis. J Cell Mol Med. 18:2213–2224.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qin S, Chen M, Ji H, Liu GY, Mei H, Li K
and Chen T: miR-21-5p regulates type II alveolar epithelial cell
apoptosis in hyperoxic acute lung injury. Mol Med Rep.
17:5796–5804. 2018.PubMed/NCBI
|
|
24
|
Qin S, Hui Y, Liu G, Mei H and Chen M:
miR-21-5p targets PTEN and reduces
H2O2-induced apoptosis in rat AECII cells.
Int J Clin Exp Med. 12:5630–5637. 2019.
|
|
25
|
Shi L, He Y, Bai B and Chen M: Effects of
microRNA-21 inhibitor on apoptosis of type II alveolar epithelial
cells in rats with hyperoxia-induced acute lung injury. Zhonghua
Wei Zhong Bing Ji Jiu Yi Xue. 29:244–248. 2017.(In Chinese).
PubMed/NCBI
|
|
26
|
Tang C, Gu Y, Wang H, Wu H, Wang Y, Meng
Y, Han Z, Gu Y, Ma W, Jiang Z, et al: Targeting of microRNA-21-5p
protects against seizure damage in a kainic acid-induced status
epilepticus model via PTEN-mTOR. Epilepsy Res. 144:34–42. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Small EM, O'Rourke JR, Moresi V,
Sutherland LB, McAnally J, Gerard RD, Richardson JA and Olson EN:
Regulation of PI3-kinase/Akt signaling by muscle-enriched
microRNA-486. Proc Natl Acad Sci USA. 107:4218–4223. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hao XJ, Xu CZ, Wang JT, Li XJ, Wang MM, Gu
YH and Liang ZG: miR-21 promotes proliferation and inhibits
apoptosis of hepatic stellate cells through targeting PTEN/PI3K/AKT
pathway. J Recept Signal Transduct Res. 38:455–461. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu G, Mei H, Chen M, Qin S, Li K, Zhang W
and Chen T: Protective effect of agmatine against hyperoxia-induced
acute lung injury via regulating lncRNA gadd7. Biochem Biophys Res
Commun. 13:68–74. 2019. View Article : Google Scholar
|
|
31
|
Matute-Bello G, Downey G, Moore BB,
Groshong SD, Matthay MA, Slutsky AS and Kuebler WM; Acute Lung
Injury in Animals Study Group, : An official american thoracic
society workshop report: Features and measurements of experimental
acute lung injury in animals. Am J Respir Cell Mol Biol.
44:725–738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin J, Tian J, Wang L, Wu W, Li H, Wang X,
Zeng X and Zhang W: Apoptosis and surfactant protein-C expression
inhibition induced by lipopolysaccharide in AEC II cell may
associate with NF-κB pathway. J Toxicol Sci. 42:53–61. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao S, Xiao Z, Sun M and Li Y: D-serine in
the midbrain periaqueductal gray contributes to morphine tolerance
in rats. Mol Pain. 12(pii): 17448069166467862016.PubMed/NCBI
|
|
35
|
Cao S, Li Q, Hou J, Li Z, Cao X, Liu X and
Qin B: Intrathecal TRPM8 blocking attenuates cold hyperalgesia via
PKC and NF-κB signaling in the dorsal root ganglion of rats with
neuropathic pain. J Pain Res. 12:1287–1296. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ou H, Li Y and Kang M: Activation of
miR-21 by STAT3 induces proliferation and suppresses apoptosis in
nasopharyngeal carcinoma by targeting PTEN gene. PLoS One.
9:e1099292014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fang L, Wang X, Sun Q, Papakonstantinou E,
S'ng C, Tamm M, Stolz D and Roth M: IgE downregulates PTEN through
microRNA-21-5p and stimulates airway smooth muscle cell remodeling.
Int J Mol Sci. 20(pii): E8752019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Meng F, Henson R, Lang M, Wehbe H,
Maheshwari S, Mendell JT, Jiang J, Schmittgen TD and Patel T:
Involvement of human micro-RNA in growth and response to
chemotherapy in human cholangiocarcinoma cell lines.
Gastroenterology. 130:2113–2129. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li JW, Wei L, Han Z and Chen Z:
Mesenchymal stromal cells-derived exosomes alleviate
ischemia/reperfusion injury in mouse lung by transporting
anti-apoptotic miR-21-5p. Eur J Pharmacol. 852:68–76. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu M, Zhu S, Xiong S, Xue X and Zhou X:
MicroRNAs and the PTEN/PI3K/Akt pathway in gastric cancer (Review).
Oncol Rep. 41:1439–1454. 2019.PubMed/NCBI
|
|
42
|
Matsuda S, Nakagawa Y, Kitagishi Y,
Nakanishi A and Murai T: Reactive oxygen species, superoxide
dimutases, and PTEN-p53-AKT-MDM2 signaling loop network in
mesenchymal stem/stromal cells regulation. Cells. 7(pii): E362018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gao X, Qin T, Mao J, Zhang J, Fan S, Lu Y,
Sun Z, Zhang Q, Song B and Li L: PTENP1/miR-20a/PTEN axis
contributes to breast cancer progression by regulating PTEN via
PI3K/AKT pathway. J Exp Clin Cancer Res. 38:2562019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ren W, Hou J, Yang C, Wang H, Wu S, Wu Y,
Zhao X and Lu C: Extracellular vesicles secreted by hypoxia
pre-challenged mesenchymal stem cells promote non-small cell lung
cancer cell growth and mobility as well as macrophage M2
polarization via miR-21-5p delivery. J Exp Clin Cancer Res.
38:622019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yan L, Cao R, Liu Y, Wang L, Pan B, Lv X,
Jiao H, Zhuang Q, Sun X and Xiao R: miR-21-5p links
epithelial-mesenchymal transition phenotype with stem-like cell
signatures via AKT signaling in keloid keratinocytes. Sci Rep.
6:282812016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Devaney J, Contreras M and Laffey JG:
Clinical review: Gene-based therapies for ALI/ARDS: Where are we
now? Crit Care. 15:2242011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
MacLoughlin RJ, Higgins BD, Laffey JG and
O'Brien T: Optimized aerosol delivery to a mechanically ventilated
rodent. J Aerosol Med Pulm Drug Deliv. 22:323–332. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Suen CM, Mei SH, Kugathasan L and Stewart
DJ: Targeted delivery of genes to endothelial cells and cell- and
gene-based therapy in pulmonary vascular diseases. Compr Physiol.
3:1749–1779. 2013. View Article : Google Scholar : PubMed/NCBI
|