Introduction
Non-alcoholic fatty liver disease (NAFLD) is one of
the metabolic diseases affecting 20–30% of the population in
Western countries, which is characterised by an excess accumulation
of fat in hepatocytes (1). In
10–25% of subjects, NAFLD may progress to non-alcoholic
steatohepatitis (NASH), which is characterised by histological
features of hepatocyte ballooning and lobular inflammation
(2,3). As skeletal muscle accounts for
>80% of insulin-stimulated glucose from the blood stream and
stored in the muscle as glycogen, it is a key tissue type
contributing to the development of systemic insulin resistance
(4). In recent years, skeletal
muscle insulin resistance has been posited to contribute to the
development of NAFLD by driving hepatic de novo lipogenesis
of the ingested carbohydrates, as compared with skeletal muscle
glycogen synthesis, leading to hypertriglyceridemia and increased
hepatic triglyceride synthesis (5,6). The
hypothesis is further supported by data from experimental and
epidemiological studies, suggesting the importance of skeletal
muscle insulin resistance as a potential and promising therapeutic
target for treating NAFLD (7,8).
At present, the precise molecular mechanisms of
skeletal muscle insulin resistance remain unclear. Nevertheless,
emerging evidence has demonstrated the roles of inflammation and
oxidative stress in predisposition to insulin resistance: It has
been indicated that hyperlipidaemia results in the increases in
fatty acids uptake and production of fatty acids metabolites in the
skeletal muscle, which promotes the inflammatory responses
(9), and the pro-inflammatory
cytokines generated in the skeletal muscle lead to insulin
resistance by inhibiting insulin signal transduction with increased
macrophage infiltration (10).
Oxidative stress is the by-product of insufficient clearance of the
cellular production of oxidants, including reactive oxygen species
(ROS) and reactive nitrogen species (RNS), by the antioxidant
defence system within the cell (11). Emerging data indicate that
oxidative stress due to increased ROS and RNS generation and/or
compromised antioxidant systems may serve a fundamental role in the
aetiology of skeletal muscle insulin resistance (12,13).
Therefore, improving the understanding of the processes by which
inflammation, oxidative stress and insulin resistance develop and
interact in the pathogenesis of NAFLD may provide important
breakthroughs for the prevention of and interventions for this
disease.
Chronic aerobic exercise is considered as an
effective treatment strategy for NAFLD. While a number of studies
emphasise the benefits of exercise in the liver (14–16),
few studies have considered the role of skeletal muscle in the
amelioration of fatty liver as a result of chronic exercise.
Skeletal muscle insulin resistance has recently been proposed to be
a key factor in the progression of NAFLD; therefore, it is
reasonable to assume that the therapeutic effect of exercise
training on NAFLD may be partly dependent on the improvement of
insulin sensitivity with an decreased inflammatory response and
levels of oxidative stress in the skeletal muscle (5).
Of the multiple previously described NASH animal
models (17,18), the model described in the present
study, which did not require a very high level of fat in the diet
(30% fish oil), is more relevant and closer to the clinical
condition (19). Although at low
dosage (≤10% of total kcal), fish oil rich in ω-3 polyunsaturated
fatty acids has been widely demonstrated to be beneficial for
NAFLD, for example improving hepatic lipid metabolism and
regulating bile components (20,21),
30% fish oil has been observed to cause dysfunctional lipid
metabolism, particularly in female rats that are more susceptible
to liver damage, according to our previous data (19). The present study investigated the
molecular effects of a 4-week aerobic exercise regimen in the rat
skeletal muscle, involving insulin resistance, inflammation and
oxidative stress in the pathogenesis of NASH.
Materials and methods
Animal protocol
A total of 24–36 Female Sprague-Dawley rats (8–10
weeks old; 180–200 g; n=6–9 per group) were obtained from the
Laboratory Animal Unit of The University of Hong Kong. The
experimental groups maintained were in a controlled environment
(21°C; 12:12 light: Dark cycle) and divided into four groups: i)
Rats fed with normal chow (PicoLabH Rodent Diet 20; LabDiet); ii)
exercise rats fed with normal chow; iii) rats fed with a high-fat
diet (HFD); iv) rats fed an HFD + exercise. All HFD rats were fed
with 30% fat from fish oil (Sigma-Aldrich; Merck KGaA) throughout
the study for 12 weeks, while the calories of normal chow contain
13% calories from fat. The preparation of HFD and the development
of NASH animal model were performed as described previously
(19). Rats in the exercise group
were trained to run on a rotarod from 9–12 weeks (30 min/day, 6
days/week). The exercise protocol consisted of 5 m/min for 3 min,
10 m/min for 3 min, and 20 m/min for 24 min. After the 12th week,
rats underwent terminal anaesthesia by an intraperitoneal injection
of sodium pentobarbital (150 mg/kg) after 12 h fasting. All
procedures were approved by the Committee of Animal Use for
Research and Teaching at The University of Hong Kong (approval no.
CULATR No. 2787–12).
Quantitative nuclear magnetic
resonance (qNMR)
Measurements of the rats' body fat mass and
composition by qNMR were calculated individually using a Brucker
model mq10 NMR analyser (Brucker, Milton, Ontario, Canada). All
data were recorded within a 1 min period.
Glucose tolerance test (GTT)
In the 12th week, GTTs were performed in rats by
injecting D-glucose (0.5 g/kg; Sigma-Aldrich; Merck KGaA) after 12
h fasting. The ACCU-CHEK blood glucose monitoring system (Roche
Diagnostics) was used to examine the levels of glucose in the tail
vein blood samples from each rat at 0, 20, 40, 60, 80, 100 and 120
min following glucose injection.
Insulin tolerance test (ITT)
In the 12th week, rats were subjected to ITT after
12 h fasting and were injected with recombinant insulin (0.17 IU;
cat# 7544-MR-050, R&D Systems, Inc.). Tail vein blood samples
were collected for glucose level test at 0, 20, 40, 60, 80, 100 min
following insulin injection using the ACCU-CHEK blood glucose
monitoring system. Serum insulin levels were quantified by using
High Sensitive Rat Insulin kit from Antibody and Immunoassay
Services of The University of Hong Kong (cat# 33270).
Histological studies
Liver and skeletal muscle tissues (hamstrings and
gastrocnemius) were fixed for 72 h with 10% phosphate-buffered
formalin and then embedded in paraffin blocks at ambient
temperature. Following slicing of the tissues into 5-µm thick
sections, they were stained with 0.1% haematoxylin and 0.1% eosin
(H&E) and 0.1% picro-Sirius Red (Polysciences Inc.) at ambient
temperature as previously described (22). NAFLD activity scores were
calculated to evaluate the hepatic histopathological injury based
on the levels of steatosis, lobular inflammation, hepatocyte
ballooning and fibrosis as previously described (23).
Immunohistochemistry (IHC)
IHC staining was performed as described previously
(24). Briefly, paraffin sections
of skeletal muscle were processed and immunostained with glucose
transporter 4 (GLUT4; Santa Cruz Biotechnology, Inc.) and inducible
nitric oxide synthase (iNOS; BD Biosciences), using an ABC kit
(Vector Laboratories Ltd.). The peroxidase was visualized using a
DAB kit (Dako Denmark A/S; Agilent Technologies, Inc.). Negative
control sections were incubated with secondary antibodies from Dako
anti-mouse (cat# K4000) or anti-rabbit (cat# K4002). The presence
of a brown chromogen indicated a positive staining result.
Semi-quantitative assessment method was used in all IHC slides with
1+ to 3+ scoring (10–30% staining=1, 31–60%=2, 61–100%=3).
Oil red O staining
Frozen muscle tissues were cut into 8–10 µm
sections, fixed in 10% phosphate-buffered formalin for 72 h and
stained with freshly prepared Oil Red O working solution for 15 min
at ambient temperature. Nuclei were lightly stained with
haematoxylin solution 5 dips as previously described (25).
Nuclear and cytoplasmic protein
extraction
Nuclear and cytoplasmic proteins were extracted from
skeletal muscle tissue using NE-PER Nuclear and Cytoplasmic
Extraction Reagents kit (Pierce; Thermo Fisher Scientific, Inc.)
with protease inhibitors aprotinin, leupeptin and pepstatin A
(Sigma-Aldrich; Merck KGaA) according to the manufacturers'
protocol.
Serum alanine aminotransferase (ALT)
assay
Rat serum ALT levels were evaluated by using ALT
(SGPT) reagent kits (Teco Diagnostics cat# A524150) according to
the manufacturer's protocol, to assess the overall hepatic injury
caused by an HFD.
Malondialdehyde (MDA) assay
The levels of MDA were examined by Bioxytech
LPO-586™ kit (OXIS Health Products, Inc. cat# 21012) according to
the manufacturers' protocol. Skeletal muscle tissues were
homogenised, followed by the determination of total protein
concentration using protein assay kits (Bio-Rad Laboratories, Inc.
cat# 5000002) and an ELISA-based assay, with
1,1,3,3-tetraethoxypropane used to generate the standard curve.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from each liver sample was extracted using
an illustra™ RNAspin mini kit (GE Healthcare Life Sciences),
followed by RT processing using the SuperScript™ First-Strand
Synthesis System (Invitrogen; Thermo Fisher Scientific, Inc.). The
mRNA expression levels of lipogenic genes, fat-specific protein 27
(Fsp27), carbohydrate-responsive element-binding protein (ChREBP)
and liver X receptor (LXR), were evaluated by Takara SYBR premix
Taq qPCR system (Takara Bio, Inc) using a StepOnePlus™ Real-Time
PCR instrument (Thermo Fisher Scientific, Inc.). GAPDH expression
was used as a reference gene for normalisation. The thermocycle
programs comprises initial denaturation 95°C for 30 sec, 40 cycles
of denaturation at 95°C for 5 sec, annealing and elongation at 60°C
for 30 sec, and final extension at 60°C for 30 sec. The efficiency
and specificity of all primers were examined prior to the
experiment. The expression levels of the target genes were
calculated using the 2−ΔΔCq method (26) and then compared with the control
group. The primer sequences for peroxisome proliferator-activated
receptor α (PPARα), Fsp27, ChREBP and LXR are listed in Table I.
 | Table I.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Target gene | Direction | Sequence |
|---|
| PPARα | Forward |
TACCACTATGGAGTCCACGCATG |
|
| Reverse |
TTGCAGCTTCGATCACACTTGTCG |
| ChREBP | Forward |
GAAACCTGAGGCTGTTGTCTTG |
|
| Reverse |
TGTGGTATTCACGCATCA |
| Fsp27 | Forward |
AGCTATCCCTTTCCCAGAAG |
|
| Reverse |
CCCTGTAGCAGTGCAGGTCA |
| LXR | Forward |
CTGCAGGACAAAAAACTTCC |
|
| Reverse |
CCCTCCTCAGTCTGCTCCAC |
Determination of serum free fatty acid
(FFA) and triglyceride (TG)
The levels of rat serum FFA were determined by an
FFA fluorometric assay (cat# 700310, Cayman Chemical Company) using
a coupled enzymatic reaction, according to the manufacturers'
protocol. Oleic acid was used for the preparation of the standard
curve. Serum TG levels were determined by Serum Triglyceride
Determination Kit according to manufacturers' protocol (cat#
TR0100, Sigma-Aldrich; Merck KGaA).
Western blot analysis
Western blot analyses were conducted as previously
described (19). The dilution of
all primary antibodies was 1:1,000. The following proteins were
assessed: NF-κB (cat# sc-372-G, Santa Cruz Biotechnology, Inc.);
NF-κB inhibitor α (IκBα; cat# sc-1643, Santa Cruz Biotechnology,
Inc.); interleukin (IL)-6 (cat# sc-57315, Santa Cruz Biotechnology,
Inc.); nuclear factor erythroid 2-related factor 2 (Nrf2; cat#
ab92946, Abcam); kelch-like ECH-associated protein 1 (Keap1; cat#
ab139729, Abcam); superoxide dismutase [Cu-Zn] (SOD-1; cat#
sc-271014, Santa Cruz Biotechnology, Inc.); GAPDH (cat# 2118, Cell
Signaling Technology, Inc.); tripartite motif-containing family
protein 72 (TRIM72; cat# ab118651, Abcam); insulin receptor
substrate 1 (IRS1, cat# 3407, Cell Signaling Technology, Inc.);
phosphorylated (Ser307) IRS1 (p-IRS1; cat# 2381, Cell Signaling
Technology, Inc.); PI3K (cat# ab191606, Abcam); Akt (cat# 4685,
Cell Signaling Technology, Inc.); phosphorylated (Ser473) Akt
(p-Akt; cat# 4058, Cell Signaling Technology, Inc.); mTOR (cat#
2983, Cell Signaling Technology, Inc.); phosphorylated (Ser307)
mTOR (cat# 2971, p-mTOR; Cell Signaling Technology, Inc.); and
GLUT4 (cat# sc-53566, Santa Cruz Biotechnology, Inc.). The dilution
of anti-mouse IgG (H+L)-peroxidase (cat# NA931) and anti-rabbit IgG
(H+L)-peroxidase (cat# NA934) antibodies was 1:2,000 (GE
Healthcare). The optical density was read by Image J (Version
1.51j8; National Institutes of Health).
ELISA
The protein level of rat tumor necrosis factor-α
(TNF-α) was determined using a DuoSet ELISA kit (cat# DY510,
R&D Systems, Inc.).
Statistical analysis
Data from each group (n=6-9) are presented as mean ±
standard deviation. One-way analysis of variance (ANOVA) followed
by a post-hoc Tukey's multiple comparisons test was performed to
detect statistical differences between groups. For data from the
body weight, GTT and ITT assays, a two-way ANOVA followed by a
post-hoc Tukey's multiple comparisons test was performed to
evaluate the statistical differences among groups at different time
points. Kolmogorov-Smirnov test for normal distribution of data
were performed. P<0.05 was considered to indicate a
statistically significant difference. All analyses were performed
using IBM SPSS v.18 (SPSS, Inc.) software.
Results
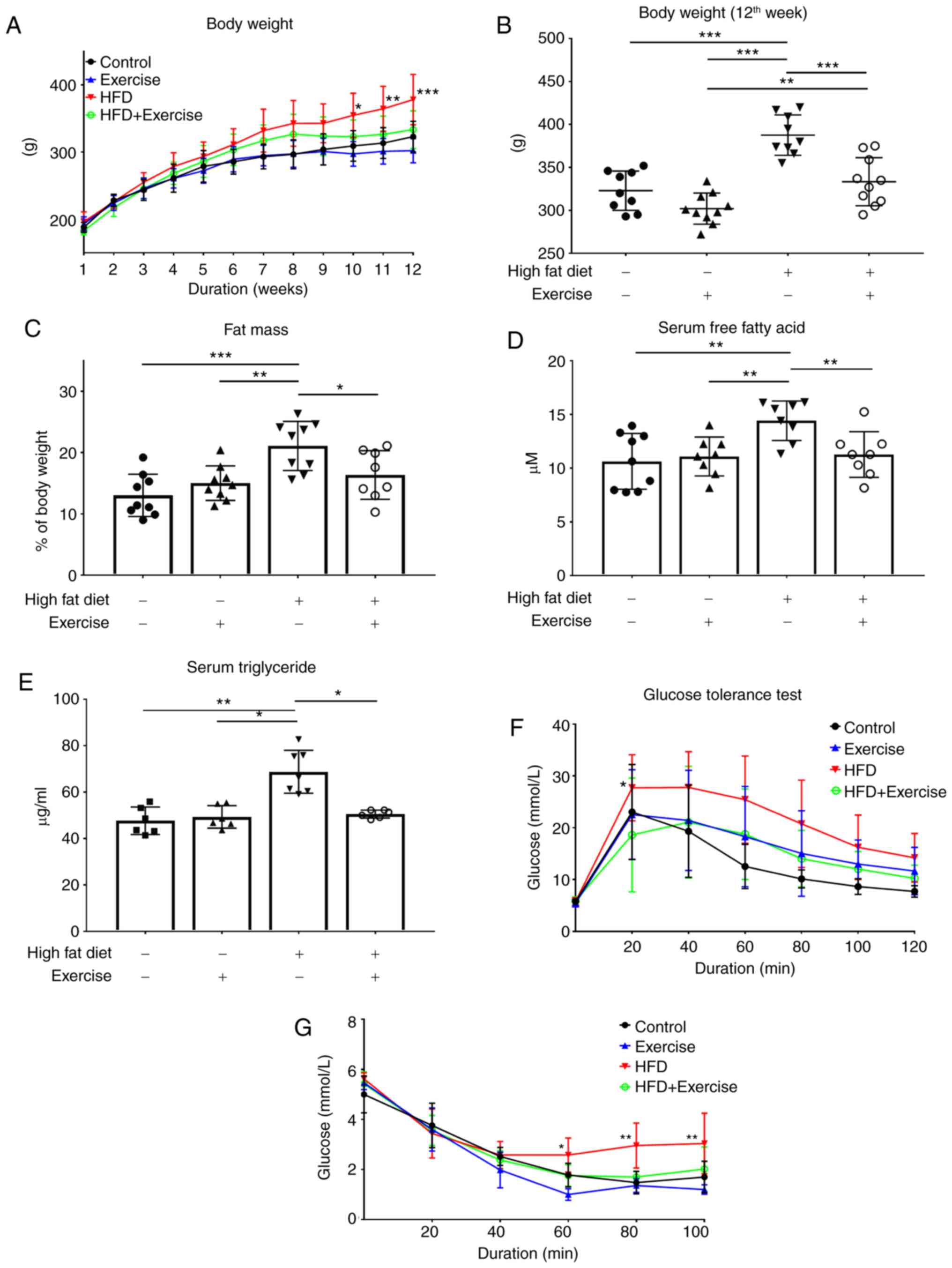
A 4-week treadmill exercise regimen
decreases body weight, fat mass and serum FFA levels, and rescues
HFD-induced glucose and insulin intolerance
In the 9th week, rats began aerobic exercise, and in
the following 4 weeks the body weight of rats in HFD + Exercise
group decreased as compared with those in the HFD group (Fig. 1A). In the 12th week, body weight
and fat mass were increased in the HFD diet only group, while the
4-week exercise treatment significantly decreased both body weight
(P<0.001) and fat mass (P<0.01) in the HFD + exercise group
(Fig. 1B and C). The elevated
levels of serum FFA in HFD rats were also downregulated following
exercise (P<0.01; Fig. 1D).
Increases in glucose and insulin intolerance were observed in the
HFD rats, but these effects were significantly decreased following
the exercise regimen (Fig. 1E and
F). No difference in the ratio of liver weight and body weight
in the rats in the present study was observed (data not shown).
Chronic exercise decreases hepatic
lipid accumulation and lipogenesis, serum ALT and fat accumulation
in muscle
There were severe liver injuries observed in the HFD
rats, characterised by foci of inflammation and fat accumulation,
as demonstrated by H&E staining of liver tissue, increases in
serum ALT levels (65.54 IU/l in HFD group) (P<0.001; Fig. 2A-C) and increases in fat
accumulation in the skeletal muscle (Fig. 2G), which were remarkably reversed
by exercise training. The elevated lipogenic markers PPARα
(P<0.001), ChREBP (P<0.001), Fsp27 (P<0.01) and LXR
(P<0.001) in HFD rats were suppressed after 4 weeks of exercise
(Fig. 2D-F). No difference in
muscle histology among the four groups was observed (data not
shown).
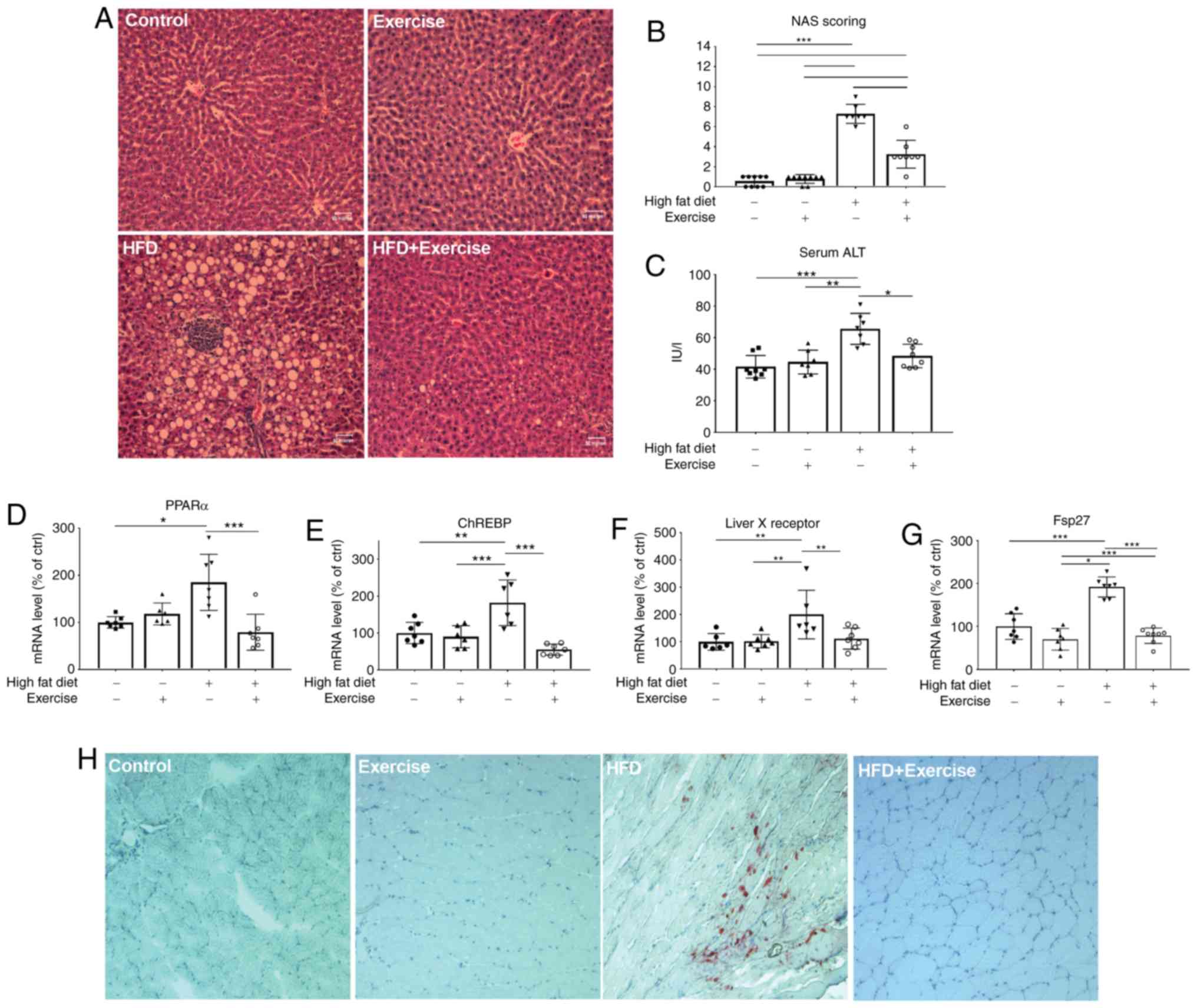 | Figure 2.Effects of a 4-week aerobic exercise
regimen. (A) Liver haematoxylin and eosin staining in the control,
exercise, HFD and HFD + exercise groups. (B) Corresponding NAS
score. The levels of (C) liver injury marker serum ALT (IU/l), and
mRNA expression levels of (D) PPARα, (E) ChREBP, (F) liver X
receptor and (G) Fsp27 were measured. The mRNA expression data are
presented as relative to the control group, which was arbitrarily
assigned a value of 100. Data are presented as mean ± standard
deviation. *P<0.05, **P<0.01 and ***P<0.001. (H) Muscle
oil red O staining (magnification, ×400). HFD, high-fat diet; NAS,
non-alcoholic fatty liver disease activity scores; ALT, alanine
aminotransferase; PPARα, peroxisome proliferator-activated receptor
α; ChREBP, carbohydrate-responsive element-binding protein; Fsp27,
fat-specific protein 27. |
Chronic aerobic exercise increases
skeletal muscle insulin sensitivity partly through regulation of
TRIM72 expression and PI3K/Akt/mTOR pathway with upregulation of
GLUT4
The molecular mechanisms of the insulin signaling
pathway in the skeletal muscle, which may contribute to whole-body
insulin resistance, were then investigated. The skeletal muscles
examined were hamstrings and gastrocnemius muscles, primarily made
up of fast twitch muscle fibres and that no differences were
observed between the muscles. TRIM72 is a key component in insulin
resistance, and it may inhibit insulin signaling by targeting IRS
and IRS1 for degradation in the skeletal muscle and heart (27–29).
Skeletal muscle is a key tissue responsible for insulin-stimulated
glucose consumption, so the present study measured the expression
levels of TRIM72, IRS1, PI3K, Akt, mTOR and GLUT4 proteins. In the
HFD rats, TRIM72 was significantly increased (P<0.01) while
p-IRS (P<0.05), PI3K (P<0.05), p-Akt (P<0.05), and GLUT4
(P<0.01) were decreased compared with rats in the control group,
suggesting that there is an impairment of insulin signaling at the
molecular level (Figs 3A-F). In
addition, IHC staining also indicated decreased GLUT4 expression in
the HFD group (Fig. 3H).
Conversely, exercise alleviated the HFD-induced elevation of
TRIM72, increased the expression levels of p-IRS1 (P<0.001) and
GLUT4 (P<0.01), and restored the PI3K-Akt pathway, as evidenced
by increases in PI3K (P<0.05), p-Akt (P<0.01) and p-mTOR
(P<0.01) expression levels (Fig.
3A-H), suggesting that exercise training may contribute to the
improvement of glucose tolerance and systemic insulin sensitivity
by stimulating glucose disposal and increasing insulin sensitivity
in the skeletal muscle. However, no changes in IRS1 expression were
observed (Fig. 3G).
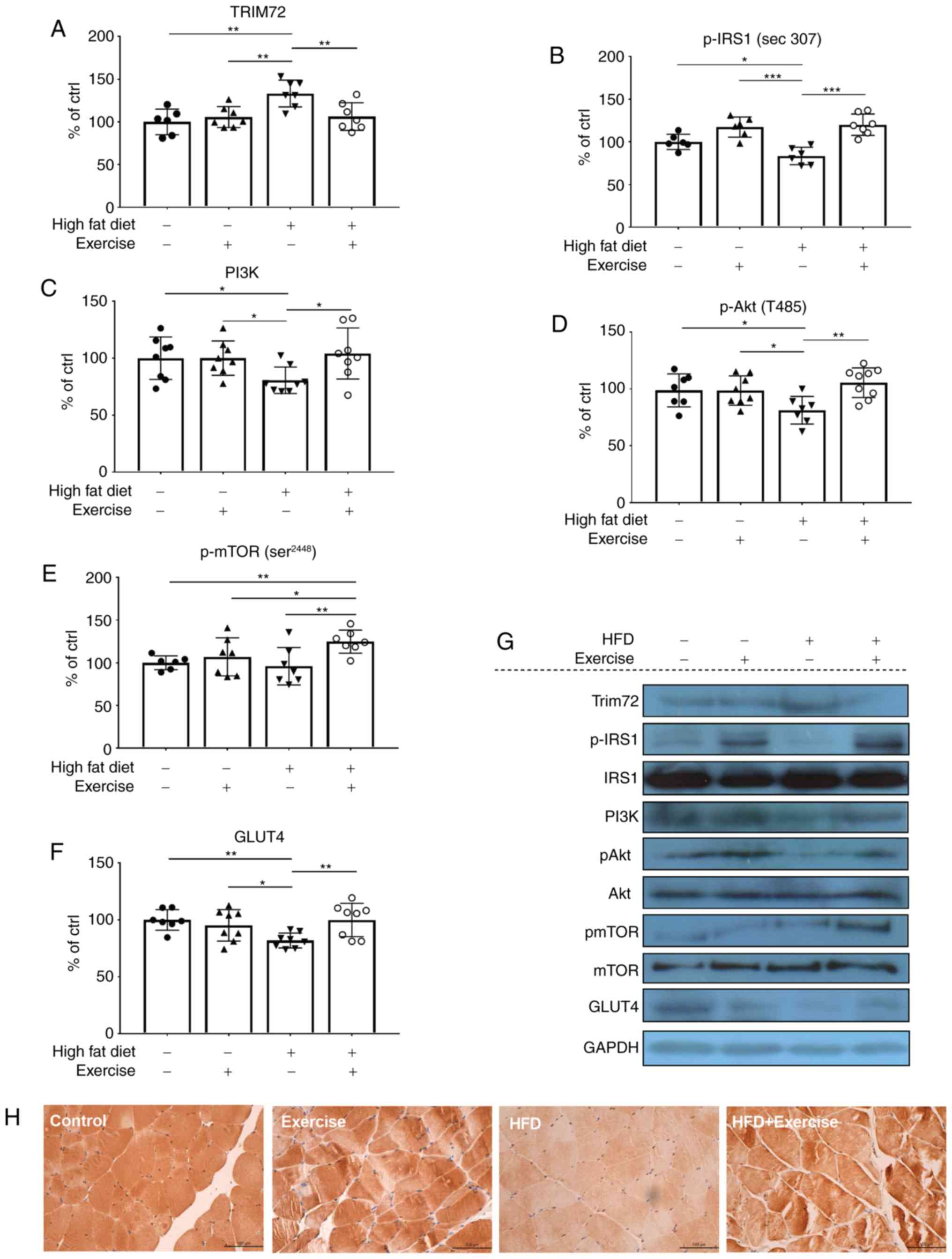 | Figure 3.Protein expression levels of (A)
TRIM72, (B) p-IRS1, (C) PI3K, (D) p-Akt, (E) p-mTOR and (F) GLUT4
were examined using western blot analysis. Data are presented as
relative to the control group, which was arbitrarily assigned a
value of 100. Data are presented as mean ± standard deviation.
*P<0.05, **P<0.01 and ***P<0.001. (G) Representative
western blot analysis gel using samples from in the skeletal
muscles of the control, exercise, HFD and HFD + exercise groups.
(H) Representative images of immunohistochemical staining of GLUT4
expression levels in the skeletal muscle (magnification, ×400).
HFD, high-fat diet; TRIM72, tripartite motif-containing family
protein 72; IRS1, insulin receptor substrate 1; p-, phosphorylated;
GLUT4, glucose transporter 4. |
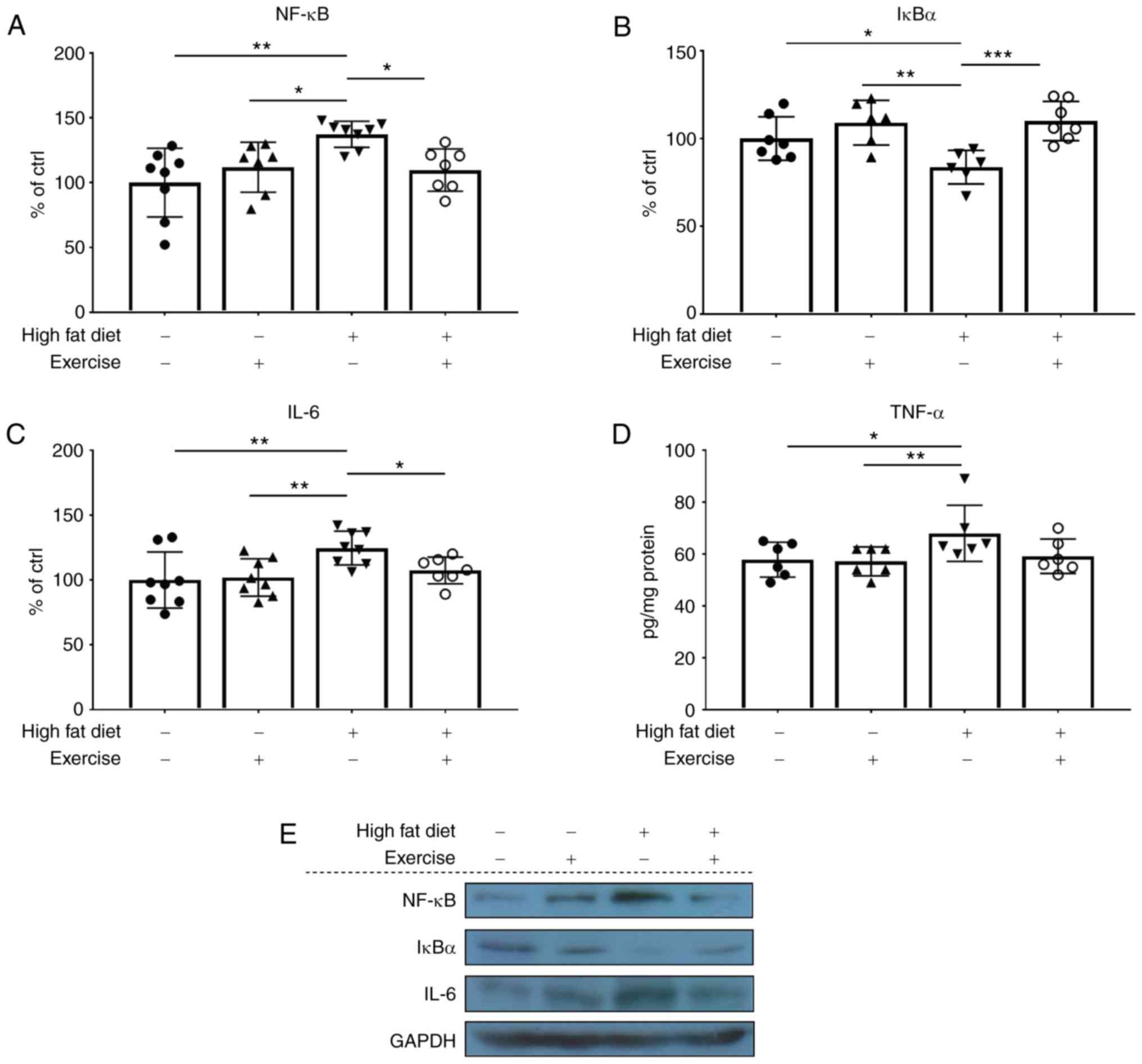
Exercise training ameliorates
HFD-induced inflammation via the NF-κB/IκBα pathway in skeletal
muscle
NF-κB is a master regulator of the inflammation
pathway; therefore, the protein expression levels of NF-κB in the
nucleus, and the downstream inflammatory markers IL-6 and TNF-α,
were measured. NF-κB (P<0.01; Fig.
4A and E), IL-6 (P<0.01; Fig.
4C and E) and TNF-α (P<0.05; Fig. 4D and E) were increased in the HFD
rats, while exercise training decreased the HFD-induced increase of
these markers to the normal level. These results were additionally
verified by corresponding levels of IκBα protein expression in the
cytoplasm (Fig. 4B and E).
Chronic aerobic exercise activates
Nrf2 and upregulates the antioxidant enzyme SOD-1 but fails to
decrease oxidative and nitrosative stress in the skeletal muscle of
NASH rats
To investigate whether exercise training was able to
exert insulin-sensitizing effects partly via the decrease in
oxidative stress, the antioxidant response and oxidative stress
level were measured in the skeletal muscle. Nrf2 is a key
transcription factor that regulates the antioxidant response.
Notably, Nrf2 expression was significantly increased in the
exercise rats with or without an HFD diet (Fig. 5A and D), suggesting that aerobic
exercise acts on Nrf2, which may trigger the antioxidant response
in the skeletal muscle. These results were additionally verified by
examinations of Keap1 protein expression in the cytoplasm (Fig. 5B and D). The level of SOD-1, an
antioxidant enzyme that is a downstream marker for Nrf-2, exhibited
a similar trend to Nrf2 (Fig. 5C and
D). However, exercise training failed to ameliorate the high
levels of oxidative stress marker MDA (wHFD group=0.152 µg/mg; HFD
+ exercise group=0.156 µg/mg) and nitrosative stress marker iNOS in
the skeletal muscle of HFD rats (Fig.
5E and F)
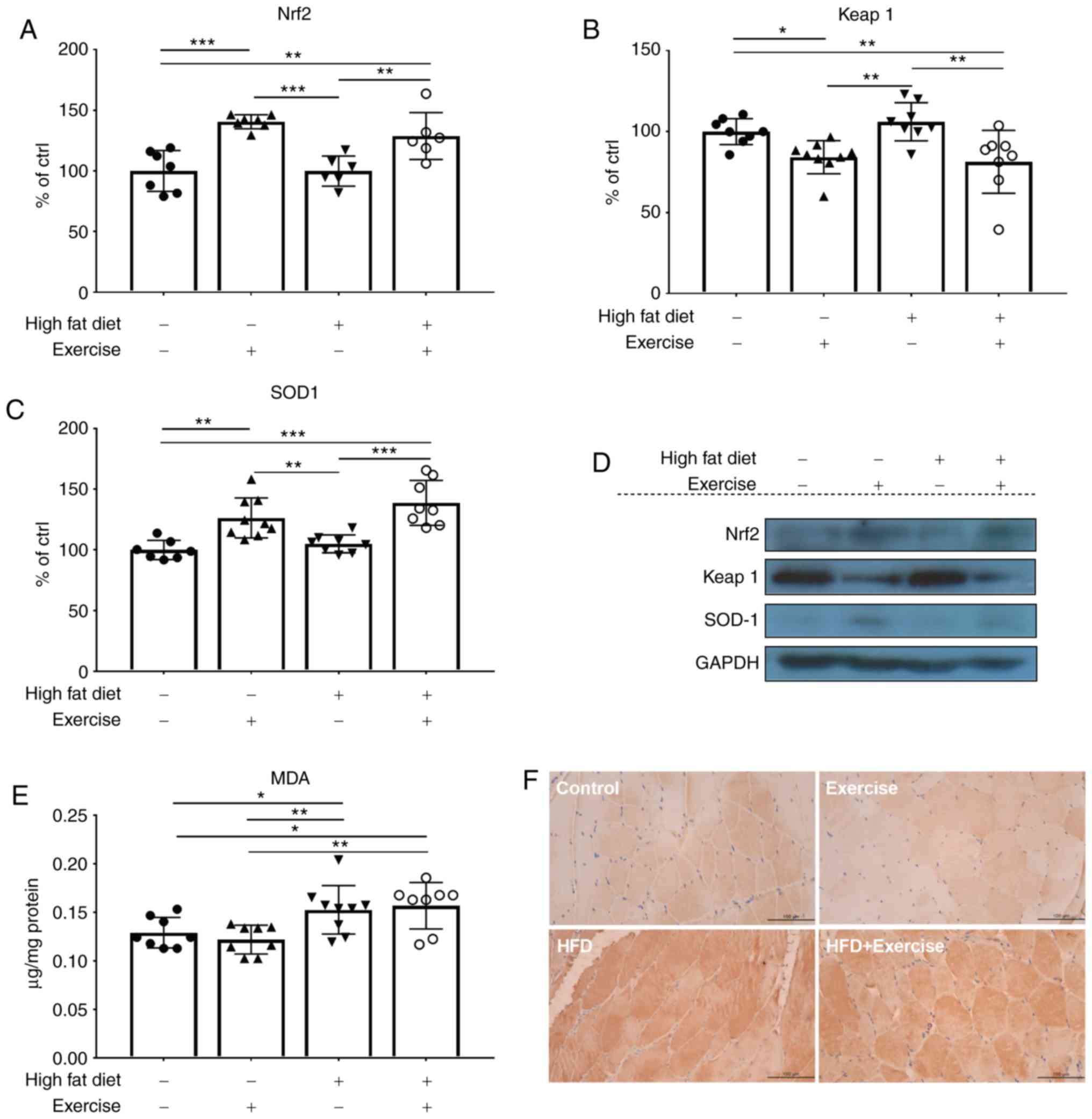 | Figure 5.Protein expression levels of (A)
Nrf2, (B) Keap1 and (C) SOD1 were examined using western blot
analysis. (D) Representative western blot analysis gel using
samples from in the skeletal muscles of the control, exercise, HFD
and HFD + exercise groups. Western blot analysis data are presented
as mean ± standard deviation. *P<0.05, **P<0.01 and
***P<0.001. The results are presented as relative to the control
group, which was arbitrarily assigned a value of 100. (E) The
content of oxidative stress marker MDA is presented as µg/mg
protein. (F) Representative images of immunohistochemical staining
of nitrosative stress marker inducible nitric oxide synthase
(magnification, ×400). Nrf2, nuclear factor erythroid 2-related
factor 2; Keap1, kelch-like ECH-associated protein 1; SOD1,
superoxide dismutase [Cu-Zn]; HFD, high-fat diet; MDA,
malondialdehyde. |
Discussion
TRIM72, also termed mitsugumin-53, was previously
identified to participate in the cell membrane repair process, with
specific expression in the skeletal muscle and heart compared with
other tissues (27). Previously,
Song et al (28)
demonstrated that this protein inhibited insulin signaling by
targeting IRS and IRS1 for degradation. To the best of our
knowledge, the present study was the first to investigate the
effects of chronic aerobic exercise on the expression level of
TRIM72 in the skeletal muscle in a NASH model. The study performed
by Qi et al (30) revealed
that an 8-week swimming regimen markedly decreased the level of
TRIM72 and increased the protein expression of p-Akt, compared with
the control. Swimming also improved the levels of total IRS1 whilst
suppressing the HFD-induced phosphorylation of IRS1 on serine 307
(p-IRS1). In the present study, chronic aerobic exercise decreased
the HFD-induced increase of TRIM72, maintained the levels of IRS1
phosphorylation, restored the PI3K/Akt/mTOR signaling pathway, and
finally resulted in the increase in GLUT4 levels, which suggested
an increase in glucose utilisation in the skeletal muscle and an
improvement in systemic insulin sensitivity. However, the results
observed for p-IRS1 and total IRS1 levels in the present study were
inconsistent with previous data: Phosphorylation of IRS1 on serine
residues has been suggested to mediate insulin resistance (31–34).
Conversely, a previous study revealed that IRS1 Ser307 is in fact a
positive regulatory site that promotes insulin sensitivity
(35). It has been hypothesized
that upregulation of Ser307 phosphorylation in mice with insulin
resistance may be an adaptive response as opposed to a pathological
change. Thus we proposed the inconsistent results observed may be
as a result of the specific type of HFD used in the present study,
which contains 30% fat from fish oil and has a pathogenic function
in the muscle. As for total IRS1, although Song et al
(28) concluded that TRIM72
targeted IR and IRS1 for degradation, a marked decrease in the HFD
group was not observed, probably as the male mice in the study by
Song et al (28) were fed
with an HFD consisting of 60% calorie from fat for 35 weeks, while
the animal model in the present study were all female rats fed with
an HFD that contained only 30% calories from fat (fish oil) for 12
weeks. It is possible that the inconsistencies between the data are
a result of the differences between the sex, diet and duration of
the animal models. However, there may be alternative explanations
for these data, and further investigations are required.
The effects of exercise on the skeletal muscle are
an integral part in the improvement of compromised systemic insulin
resistance, which manifests in decreases in the levels of oxidative
stress and inflammation. Nrf2 serves a pivotal role in the
regulation of a series of genes encoding antioxidant systems under
stress stimulus to adapt to ROS and xenobiotics (36,37).
A number of studies have demonstrated the beneficial effects of
Nrf2 on various organs and tissues, including skeletal muscle,
brain, kidney and liver (38–41).
The data from the present study also demonstrated similar results,
that a 4-week aerobic exercise regimen markedly activated the Nrf2
pathway and decreased Keap1 expression in the skeletal muscle
tissues of the rats in the exercise and HFD + exercise groups.
Therefore, consistent with previous studies, the results from the
present study suggested that aerobic exercise may exert its
beneficial effects on the improvement of insulin sensitivity partly
through the improvement of the antioxidant response. However,
chronic aerobic exercise did not decrease the level of lipid
peroxidation product MDA in the skeletal muscle of HFD + exercise
group, but instead increased MDA level by 20% compared with the
control group. Consistently, exercise training failed to decrease
the levels of nitrosative stress, as indicated by high level of
iNOS. Numerous studies have demonstrated that exercise may decrease
MDA levels in healthy individuals, however, in disease models such
as hypercholesterolemia and diabetes, it has been suggested that
exercise-induced changes in MDA were more pronounced (42,43).
Therefore, the possibility that although a 4-week aerobic exercise
regimen increased the antioxidant response, it may not be
sufficient to compensate the oxidative stress and nitrosative
stress generated in the skeletal muscle, must not be dismissed.
Nevertheless, the level of oxidative stress in the HFD + exercise
group was only increased by 20% compared with the control group,
with a significant increase in the level of antioxidant enzyme
marker SOD1 as compared with the control group. In combination with
the other beneficial effects of exercise observed in the HFD group,
it may be concluded that the level of oxidative and nitrosative
stress in the HFD + exercise group was inconsequential and not
detrimental when compared with the HFD group.
In the animal model of the present study, rats fed
with an HFD exhibited slight inflammation in the skeletal muscle. A
number of studies have investigated the anti-inflammatory effects
of chronic exercise in the skeletal muscle (44–48).
It has been previously demonstrated that Nrf2 deletion may
exaggerate inflammatory responses by inducing the expression of
NF-κB, IL-1ß and TNF-α in Nrf2 knockout mice fed with a
methionine-choline deficient diet (49). Furthermore, in a recent study, Liu
et al (50) identified that
the hepatic insulin resistance was regulated by the NF-κB signaling
pathway in Nrf2 knockout mice with NAFLD, indicating that Nrf2
deletion may lead to hepatic insulin resistance by activation of
NF-κB pathway, Therefore, we hypothesized that the ameliorative
effect of insulin resistance of chronic exercise may be mediated
via the activation of Nrf2 and suppression of the NF-κB pathway,
which are master regulators of the oxidative stress and
inflammation pathway, respectively, and are closely associated.
In conclusion, to the best of our knowledge, the
present study was the first to investigate the role of TRIM72 in
alleviating insulin resistance during chronic aerobic exercise in a
female NASH animal model fed with 30% of unsaturated fatty acids
via fish oil. The results of the present study also supported the
recently proposed hypothesis that skeletal muscle contributes to
the improvement of systemic insulin resistance, therefore driving
less postprandial glucose into hepatic lipogenesis, which may
potentially alleviate fatty liver disease. Therefore, aerobic
exercise is proposed as an important early lifestyle intervention
to prevent fatty liver progression to steatohepatitis.
Acknowledgements
The authors would like to thank Ms. Carman Leung
(School of Biomedical Sciences, The University of Hong Kong) for
her technical help.
Funding
The present study was funded by the Seed Fund for
Basic Research of University Research Committee (grant no.
20161159263).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QY and GLT designed the experiments. EL assisted in
the design of the study. QY and ZX conducted the experiments. QY
and GLT analysed data, performed statistical tests and wrote the
manuscript. All authors reviewed the final version of the
manuscript.
Ethics approval and consent to
participate
All procedures were approved by the Committee of
Animal Use for Research and Teaching at The University of Hong Kong
(approval no. CULATR No. 2787-12).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tilg H and Kaser A: Treatment strategies
in nonalcoholic fatty liver disease. Nat Clin Pract Gastroenterol
Hepatol. 2:148–155. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Krenkel O, Hundertmark J, Abdallah AT,
Kohlhepp M, Puengel T, Roth T, Branco DPP, Mossanen JC, Luedde T,
Trautwein C, et al: Myeloid cells in liver and bone marrow acquire
a functionally distinct inflammatory phenotype during
obesity-related steatohepatitis. Gut. May 10–2019.(Epub ahead of
print). View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jennison E, Patel J, Scorletti E and Byrne
CD: Diagnosis and management of non-alcoholic fatty liver disease.
Postgrad Med J. 95:314–322. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duque-Guimarães DE and Ozanne SE:
Nutritional programming of insulin resistance: Causes and
consequences. Trends Endocrinol Metab. 24:525–535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Flannery C, Dufour S, Rabøl R, Shulman GI
and Petersen KF: Skeletal muscle insulin resistance promotes
increased hepatic de novo lipogenesis, hyperlipidemia, and hepatic
steatosis in the elderly. Diabetes. 61:2711–2717. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jornayvaz FR, Samuel VT and Shulman GI:
The role of muscle insulin resistance in the pathogenesis of
atherogenic dyslipidemia and nonalcoholic fatty liver disease
associated with the metabolic syndrome. Annu Rev Nutr. 30:273–290.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kato K, Takeshita Y, Misu H, Zen Y, Kaneko
S and Takamura T: Liver steatosis is associated with insulin
resistance in skeletal muscle rather than in the liver in Japanese
patients with non-alcoholic fatty liver disease. J Diabetes
Investig. 6:158–163. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rabøl R, Petersen KF, Dufour S, Flannery C
and Shulman GI: Reversal of muscle insulin resistance with exercise
reduces postprandial hepatic de novo lipogenesis in insulin
resistant individuals. Proc Natl Acad Sci USA. 108(8): 13705–13709.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wellen KE and Hotamisligil GS:
Inflammation, stress, and diabetes. J Clin Invest. 115:1111–1119.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
de Luca and Olefsky JM: Inflammation and
insulin resistance. FEBS Lett. 582:97–105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stump CS, Henriksen EJ, Wei Y and Sowers
JR: The metabolic syndrome: Role of skeletal muscle metabolism. Ann
Med. 38:389–402. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Henriksen EJ: Exercise training and the
antioxidant alpha-lipoic acid in the treatment of insulin
resistance and type 2 diabetes. Free Radic Biol Med. 40:3–12. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Henriksen EJ, Diamond-Stanic MK and
Marchionne EM: Oxidative stress and the etiology of insulin
resistance and type 2 diabetes. Free Radic Biol Med. 51:993–999.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Johnson NA, Keating SE and George J:
Exercise and the liver: Implications for therapy in fatty liver
disorders. Semin Liver Dis. 32:65–79. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Keating SE, Hackett DA, George J and
Johnson NA: Exercise and non-alcoholic fatty liver disease: A
systematic review and meta-analysis. J Hepatol. 57:157–166. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kistler KD, Brunt EM, Clark JM, Diehl AM,
Sallis JF and Schwimmer JB; NASH CRN Research Group, : Physical
activity recommendations, exercise intensity, and histological
severity of nonalcoholic fatty liver disease. Am J Gastroenterol.
106:460–468; quiz 469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lau JK, Zhang X and Yu J: Animal models of
non-alcoholic fatty liver disease: Current perspectives and recent
advances. J Pathol. 241:36–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takahashi Y, Soejima Y and Fukusato T:
Animal models of nonalcoholic fatty liver disease/nonalcoholic
steatohepatitis. World J Gastroenterol. 18:2300–2308. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tipoe GL, Ho CT, Liong EC, Leung TM, Lau
TY, Fung ML and Nanji AA: Voluntary oral feeding of rats not
requiring a very high fat diet is a clinically relevant animal
model of non-alcoholic fatty liver disease (NAFLD). Histol
Histopathol. 24:1161–1169. 2009.PubMed/NCBI
|
|
20
|
Liu Y, Li Q, Wang H, Zhao X, Li N, Zhang
H, Chen G and Liu Z: Fish oil alleviates circadian bile composition
dysregulation in male mice with NAFLD. J Nutr Biochem. 69:53–62.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamazaki T, Nakamori A, Sasaki E, Wada S
and Ezaki O: Fish oil prevents sucrose-induced fatty liver but
exacerbates high-safflower oil-induced fatty liver in ddy mice.
Hepatology. 46:1779–1790. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tipoe GL, Leung TM, Liong EC, Lau TY, Fung
ML and Nanji AA: Epigallocatechin-3-gallate (EGCG) reduces liver
inflammation, oxidative stress and fibrosis in carbon tetrachloride
(CCl4)-induced liver injury in mice. Toxicology. 273:45–52. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kleiner DE, Brunt EM, Van Natta, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y, Tipoe GL and Fung ML: Melatonin
attenuates intermittent hypoxia-induced lipid peroxidation and
local inflammation in rat adrenal medulla. Int J Mol Sci.
15:18437–18452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen B, Ma Y, Xue X, Wei J, Hu G and Lin
Y: Tetramethylpyrazine reduces inflammation in the livers of mice
fed a high fat diet. Mol Med Rep. 19:2561–2568. 2019.PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao CM, Zhang Y, Weisleder N, Ferrante C,
Wang X, Lv F, Zhang Y, Song R, Hwang M, Jin L, et al: MG53
constitutes a primary determinant of cardiac ischemic
preconditioning. Circulation. 121:2565–2574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song R, Peng W, Zhang Y, Lv F, Wu HK, Guo
J, Cao Y, Pi Y, Zhang X, Jin L, et al: Central role of E3 ubiquitin
ligase MG53 in insulin resistance and metabolic disorders. Nature.
494:375–379. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cai C, Masumiya H, Weisleder N, Matsuda N,
Nishi M, Hwang M, Ko JK, Lin P, Thornton A, Zhao X, et al: MG53
nucleates assembly of cell membrane repair machinery. Nat Cell
Biol. 11:56–64. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qi J, Yang B, Ren C, Fu J and Zhang J:
Swimming exercise alleviated insulin resistance by regulating
tripartite motif family protein 72 expression and AKT signal
pathway in sprague-dawley rats fed with high-fat diet. J Diabetes
Res. 2016:15643862016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hirosumi J, Tuncman G, Chang L, Görgün CZ,
Uysal KT, Maeda K, Karin M and Hotamisligil GS: A central role for
JNK in obesity and insulin resistance. Nature. 420:333–336. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yi Z, Langlais P, De Filippis EA, Luo M,
Flynn CR, Schroeder S, Weintraub ST, Mapes R and Mandarino LJ:
Global assessment of regulation of phosphorylation of insulin
receptor substrate-1 by insulin in vivo in human muscle. Diabetes.
56:1508–1516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu C, Chen Y, Cline GW, Zhang D, Zong H,
Wang Y, Bergeron R, Kim JK, Cushman SW, Cooney GJ, et al: Mechanism
by which fatty acids inhibit insulin activation of insulin receptor
substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase
activity in muscle. J Biol Chem. 277:50230–50236. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kruszynska YT, Worrall DS, Ofrecio J,
Frias JP, Macaraeg G and Olefsky JM: Fatty acid-induced insulin
resistance: Decreased muscle PI3K activation but unchanged Akt
phosphorylation. J Clin Endocrinol Metab. 87:226–234. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Copps KD, Hancer NJ, Opare-Ado L, Qiu W,
Walsh C and White MF: Irs1 serine 307 promotes insulin sensitivity
in mice. Cell Metab. 11:84–92. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tebay LE, Robertson H, Durant ST, Vitale
SR, Penning TM, Dinkova-Kostova AT and Hayes JD: Mechanisms of
activation of the transcription factor Nrf2 by redox stressors,
nutrient cues, and energy status and the pathways through which it
attenuates degenerative disease. Free Radic Biol Med. 88:108–146.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chanas SA, Jiang Q, McMahon M, McWalter
GK, McLellan LI, Elcombe CR, Henderson CJ, Wolf CR, Moffat GJ, Itoh
K, et al: Loss of the Nrf2 transcription factor causes a marked
reduction in constitutive and inducible expression of the
glutathione S-transferase Gsta1, Gsta2, Gstm1, Gstm2, Gstm3 and
Gstm4 genes in the livers of male and female mice. Biochem J.
365:405–416. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Asghar M, George L and Lokhandwala MF:
Exercise decreases oxidative stress and inflammation and restores
renal dopamine D1 receptor function in old rats. Am J Physiol Renal
Physiol. 293:F914–F919. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Done AJ and Traustadóttir T: Nrf2 mediates
redox adaptations to exercise. Redox Biol. 10:191–199. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Muthusamy VR, Kannan S, Sadhaasivam K,
Gounder SS, Davidson CJ, Boeheme C, Hoidal JR, Wang L and
Rajasekaran NS: Acute exercise stress activates Nrf2/ARE signaling
and promotes antioxidant mechanisms in the myocardium. Free Radic
Biol Med. 52:366–376. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao X, Bian Y, Sun Y, Li L, Wang L, Zhao
C, Shen Y, Song Q, Qu Y, Niu S, et al: Effects of moderate exercise
over different phases on age-related physiological dysfunction in
testes of SAMP8 mice. Exp Gerontol. 48:869–880. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sen C, Packer L and Hänninen O: Handbook
of oxidants and antioxidants in exercise. Elsevier Science.
2000.PubMed/NCBI
|
|
43
|
Kim JS and Yi HK: Intermittent bout
exercise training down-regulates age-associated inflammation in
skeletal muscles. Exp Gerontol. 72:261–268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cechella JL, Leite MR, Dobrachinski F, da
Rocha JT, Carvalho NR, Duarte MM, Soares FA, Bresciani G, Royes LF
and Zeni G: Moderate swimming exercise and caffeine supplementation
reduce the levels of inflammatory cytokines without causing
oxidative stress in tissues of middle-aged rats. Amino Acids.
46:1187–1195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Isanejad A, Saraf ZH, Mahdavi M,
Gharakhanlou R, Shamsi MM and Paulsen G: The effect of endurance
training and downhill running on the expression of IL-1ß, IL-6, and
TNF-α and HSP72 in rat skeletal muscle. Cytokine. 73:302–308. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jeong JH, Park HG, Lee YR and Lee WL:
Moderate exercise training is more effective than resveratrol
supplementation for ameliorating lipid metabolic complication in
skeletal muscle of high fat diet-induced obese mice. J Exerc
Nutrition Biochem. 19:131–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tantiwong P, Shanmugasundaram K, Monroy A,
Ghosh S, Li M, DeFronzo RA, Cersosimo E, Sriwijitkamol A, Mohan S
and Musi N: NF-B activity in muscle from obese and type 2 diabetic
subjects under basal and exercise-stimulated conditions. Am J
Physiol Endocrinol Metab. 299:E794–E801. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Knudsen JG, Joensen E, Bertholdt L, Jessen
H, van Hauen L, Hidalgo J and Pilegaard H: Skeletal muscle IL-6 and
regulation of liver metabolism during high-fat diet and exercise
training. Physiol Rep. 4:e127882016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chowdhry S, Nazmy MH, Meakin PJ,
Dinkova-Kostova AT, Walsh SV, Tsujita T, Dillon JF, Ashford ML and
Hayes JD: Loss of Nrf2 markedly exacerbates nonalcoholic
steatohepatitis. Free Radic Biol Med. 48:357–371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu Z, Dou W, Ni Z, Wen Q, Zhang R, Qin M,
Wang X, Tang H, Cao Y, Wang J and Zhao S: Deletion of Nrf2 leads to
hepatic insulin resistance via the activation of NF-B in mice fed a
high-fat diet. Mol Med Rep. 14:1323–1331. 2016. View Article : Google Scholar : PubMed/NCBI
|