Introduction
Thymic stromal lymphopoietin (TSLP) is a cytokine
secreted by airway epithelium that was first identified from thymic
stromal cells by Friend et al in 1994 (1). TSLP is known to be involved in the
initiation of an inflammatory cascade and has been shown to be
upregulated in a variety of clinical conditions, including atopic
dermatitis, allergic asthma, inflammatory bowel disease, breast
cancer, cervical cancer and lung cancer (2,3).
Recently, Kuan and Ziegler (4)
demonstrated that the antibody blockade of TSLP significantly
inhibited the growth, and lung metastasis of breast cancer in a
model system. These observations suggest a potential role for TSLP
in human disease and further suggest that antibody disruption of
TSLP and the TSLP receptor (TSLPR) signalling pathways may have
clinical benefit for diseases that lack effective treatment.
However, a fully human antibody reactive with TSLP is currently
unavailable for clinical use.
Engineered antibodies have many advantages over
murine monoclonal antibodies, including that they are not murine,
and can be modified at the genetic and molecular levels to meet
requirements for immunogenicity, stability and specificity in
vivo (5). However, engineered
antibodies obtained by antibody library screening have not
undergone affinity maturation in vivo and their affinity is
generally not acceptable for clinical use (6). Therefore, affinity maturation of
recombinant antibodies in vitro is required. Traditional
strategies for the modification of engineered antibodies are
random, labour intensive and time consuming. In the present study,
Discovery Studio (DS) 4.5 software was used for homology modelling
of antibodies, antigens and predicted molecular anomalies in amino
acid residues. With this software, molecular dynamic simulation and
site-directed mutagenesis can be performed for in vitro
affinity maturation of a recombinant anti-TSLP single-chain
antibody variable fragment (anti-TSLP-scFv). The software is simple
to operate, easy to use, low in cost and is not time or labour
intensive. The fully human recombinant immunoglobulin
(anti-TSLP-scFv-Fc-M4) obtained using the DS 4.5 software was ~50
kDa and its affinity was increased 10-fold.
Materials and methods
Virtual mutation of amino acids of
anti-TSLP-scFv-84
TSLP homology modelling
The TSLP amino acid sequence was uploaded into the
DS 4.5 platform (NeoTrident Technology Ltd.). The template (NCBI
Gene ID: 85450) was selected from gene sequences with the highest
homology, performed using the Basic Local Alignment Search Tool
(BLAST). Energy minimizations were performed as described by the
TSLP antigen homology modelling method (7,8). The
rationality of the model was evaluated using a Ramachandran plot
and the Profile-3D program using DS software.
Anti-TSLP-scFv-84 homology
modelling
A diverse and natural full-human scFvs antibody
library, with a capacity of 2.5×108, was constructed
using mRNA of PBMC from healthy volunteer in a previous study
(9). By phage display,
biotinylated TSLP protein was used as an antigen to select human
anti-TSLP-scFv from the constructed human scFv library following
the protocol previously described (9). The selected anti-TSLP-scFv-84 amino
acid sequence was uploaded into the DS 4.5 system. Antibody domain
and complementarity determining region (CDR) sequences were
identified and annotated using the Annotate Antibody Sequence
protocol (Discovery studio visualizer version 16.1.0.15350)
(10). The sequences of variable
heavy chain (VH) and variable light chain (VL) were set according
to similarity, surface, light and heavy chain templates, resulting
in the greatest degree of sequence similarity. The optimal model
V_H_V_L_M0004_M was selected using probability density function
(PDF) total energy, PDF physical energy and discrete optimized
protein energy (DOPE) Score. Finally, a three-dimensional model of
anti-TSLP-scFv-84 was constructed and the rationality of the model
evaluated using Profile-3D and a Ramachandran plot.
Antibody-antigen docking
Optimized anti-TSLP-scFv-84 as a receptor and TSLP
as a ligand, with docked poses and clusters, were obtained using
the ZDOCK operation (Discovery studio visualizer v16.1.0.15350).
Scanning alanine and saturation mutations in the docking region
were used to predict amino acid mutation sites for increased
affinity. Based on mutational energy, amino acid sites were
selected to increase the affinity of the antibody after
mutation.
Screening and verification of
anti-TSLP-scFv mutations
Overlap extension PCR
Based on DS system predictive mutation sites
(Table SI), single-point mutation
primers were designed (Table SII)
and synthetized by Invitrogen, Thermo Fisher Scientific, Inc. Using
the method of overlap extension PCR, the recombinant plasmid
anti-TSLP-scFv-84 (pre-mutated) served as the PCR template in the
two first stage reactions. The first and second PCR amplifications
were performed using 32 cycles at 98°C for 10 sec, 60°C for 30 sec
and 72°C for 1 min; The first and second PCR products were mixed as
templates for the third PCR. The third PCR amplification was
performed 10 cycles at 98°C for 10 sec, 60°C for 30 sec and 72°C
for 1 min; Then, Forward primer scFv84F-TSLP and reverse primer
KM168 were added for 20 cycles. Finally, single point mutations
products of anti-TSLP-scFv were obtained (M1, M2, M3, M4 and M5).
The amplified PCR products was electrophoresed on a 1.5% agarose
gel in 1X Tris borate EDTA buffer with pH 8.3 and 0.03 µl ethidium
bromide. The gel was visualized under ultraviolet light.
Screening of mutated anti-TSLP-scFvs
in the prokaryotic expression vector pLZ16
A TSLP-pLZ16 plasmid was constructed in our previous
study based on the pUC vector (11). The mutated anti-scFv fragment and
the pLZ16 vector were digested with NotI and NocI
restriction enzymes at 37°C for 4 h. The products were ligated
using 1 µl T4 ligase (Takara Bio, Inc.) at 16°C for 12 h and
transformed into DH5αF′ competent cells (Tiangen Biotech
Co., Ltd.). Transformed positive scFvs were identified by PCR using
30 cycles at 98°C for 10 sec, 55°C for 30 sec and 72°C for 1 min.
High-fidelity thermostable ExTaq enzyme (Takara Biotechnology Co.,
Ltd.) and forward primer scfv84F-TSLP:
5′-CATGCCATGGCCGGCCCAGCCGGCCC-3′, reverse primer KM168:
5′-CTGAGTAGAAGAACTCAAACTA-3′ were employed. The positive scFvs
clones were cultured on a shaker at 37°C, 200 rpm for overnight,
and the plasmids were extracted with QuickPure Plasmid Mini Kit (CW
Biotech) and Sanger sequenced by Shanghai Sangon Biotech Co., Ltd.
The sequences were aligned in BLAST (Fig. S1). The pre-mutation scFv84 and
mutant scFvs (M1, M2, M4 and M5) were expressed at a constant
temperature of 32°C for 5 h with 1 mmol/l lisopropyl
β-D-1-thiogalactopyranoside induction. The expressed scFvs were
assessed for binding to human TSLP using indirect ELISA with 200
ng/ml HRP-labelled anti-FLAG monoclonal antibody (Thermo Fisher
Scientific, Inc.) as secondary antibody.
Construction of anti-TSLP-scFv-Fc
eukaryotic expression vectors
Construction of pcDNA3.1-scFv-Fc
expression vectors
The scFv-84 (pre-mutated) and scFv-M4 (mutated)
sequences in pLZ16 were amplified by PCR with scFv84-KpnI-F
(5′-CGGGGTACCATGGCCGGCCCAGCCGGCCCA-3′) and scFv84-BamHI-R-C
(5′-CGCGGATCCACGTTTGATCTCCAGCTTGGTCC-3′) (12). PCR amplifications were performed
with ExTaq enzyme, using 30 cycles at 98°C for 10 sec, 55°C for 30
sec and 72°C for 1 min. The target DNA fragment was digested with
KpnI at 37°C for 4 h and BamHI at 30°C for 4 h, and
then ligated with T4 ligase at 16°C for 12 h into pcDNA3.1-sp-Fc
[constructed in previous work (12), sp and IgG1 Fc were insert into
pcDNA3.1] to generate the pcDNA3.1-sp-scFv-Fc recombinant vectors.
There combinant vectors were transformed into E. coli TOPO10
competent cells (Takara Bio, Inc.). Positive clones with the
correct sp-scFv-Fc were identified by PCR using 30 cycles at 98°C
for 10 sec, 55°C for 30 sec and 72°C for 90 sec with ExTaq enzyme
(Takara Bio, Inc.). The constructed plasmids were sequenced by
Shanghai Sangon Biotech Co., Ltd.
Construction of
PMH3EN-scFv-Fc expression vectors
The sp-scFv-Fc-84 (Pre-mutated) and sp-scFv-Fc-M4
(mutated) in pcDNA3.1 were amplified by PCR using 30 cycles at 98°C
for 10 sec, 55°C for 30 sec and 72°C for 90 sec with ExTaq enzyme
(Takara Bio, Inc.) (11). Forward
primers pcDNA3.1-F (5′-CTAGAGAACCCACTGCTTAC-3′) and pcDNA3.1-R
(5′-TAGAAGGCACAGTCGAGG-3′) were employed. The target DNA fragment
was digested with HindIII and NotI at 37°C for 4 h,
and then ligated with T4 ligase (16°C, 12 h) to PMH3EN
(Hangzhou Amprotein BioEngineering Co., Ltd.) to generate the
PMH3EN-sp-scFv-Fc recombinant vectors. There combinant
vectors were transformed and positive clones were identified as
afore mentioned.
Verification of anti-TSLP-scFv-Fc
affinity
Cell preparation and transient
transfection
Transformed 293F cells were obtained from Abace
Biotechnology Co., Ltd. and maintained in serum-free FreeStyle™ 293
expression medium (Gibco; Thermo Fisher Scientific, Inc.). Cells
were cultured as described previously (13,14).
Filter-sterilized plasmid DNA (PMH3EN,
PMH3EN-sp-scFv-Fc-84/M4 vector) was prepared for
transfection as described previously (14). Briefly, 293F cells were cultured at
1×106 cells/ml to a final volume of 90 ml with
Serum-free Freestyle™ 293 expression medium in roller bottle
(Corning). Filter-sterilized DNA (100 µg) was added to 10 ml of
1XPBS and vortexed for 5 sec. Then, 313 µl of 1 mg/ml
filter-sterilized polyethylenimine PEI (Polysciences, Inc.)
solution was pipetted into the DNA/PBS mixture. The DNA/PEI/PBS
mixture was added to the cells, which were incubated in an orbital
shaker incubator at 37°C, 120 rpm and 8% CO2 for 72 h.
After centrifuging at 4°C, 350 g for 10 min, the cells pellet was
collected and stored at −80°C for ELISA and western blotting.
Direct ELISA
After centrifuging, cells were resuspended in 1XPBS
buffer and sonicated. The sonicated cell solutions were centrifuged
and the supernatant containing the expressed protein was used to
ELISA. scFv-Fc-84/M4 protein solutions were dilute with 1XPBS (at
various concentrations 1:1, 1:10, 1:100, 1:1,000 and 1:10,000)
derived from the PMH3EN vector were used to coat 96-well
plates overnight at 4°C in 50 mM
NaHCO3/Na2CO3 (pH 9.6). 200 ng/ml
goat pAb to Hu IgG (HRP) (Abcam, ab97225) was used to detect
PMH3EN-scFv-Fc as described previously (15). The absorbance of each well was
detected at 450 nm.
Indirect ELISA
As described previously (9), 10 µg/ml TSLP (Sino Biological Inc.)
was coated in 96-well plates overnight at 4°C. Following this, a
scFv-Fc-84/M4 protein solution (1:1, 1:10, 1:100, 1:1,000,
1:10,000) expressed in PMH3EN was added to the plates,
then 200 ng/ml goat pAb to Hu IgG (HRP, 1:5,000; Abcam, ab97225)
was added to measure binding between antigen TSLP (Sino Biological)
and antibody PMH3EN-scFv-Fc-84/M4. The absorbance of
each well was detected at 450 nm.
Western blotting
The scFv-Fc-84/M4 protein solutions were prepared by
sonicating the transfected 293F cells in 1×PBS as afore mentioned
and the BCA method was used to determine the protein concentration.
Total cellular proteins [30 µg: Control (cells only),
PMH3EN, scFv-Fc-84/M4] expressed in the
PMH3EN were subjected to SDS-PAGE with 10% gels and
transferred to nitrocellulose membranes. After blocking with 5%
non-fat milk for 1 h at room temperature, the membranes were
incubated with 200 ng/ml goat pAb to Hu IgG (HRP, 1:5,000; Abcam,
ab97225) for overnight at 4°C. A chemiluminescence detection kit
(GE Healthcare) was used to detect the expression of scFv-Fc-84/M4.
According to previously described methods (16), 5 µg TSLP (16135-H08H; Sino
Biological, Inc.) was subjected to electrophoresis and membranes
were subsequently incubated with scFv-Fc-84/M4 (400 ng/ml)
antibodies overnight at 4°C, a secondary anti-human IgG Fc
HRP-conjugated antibody was used to detect the binding ability of
antigen (TSLP) and antibody (scFv-Fc-84/M4).
Affinity test
The scFv-Fc-84/M4 protein solutions were prepared as
afore mentioned. In total, 4 µl protein supernatant of the
single-chain antibody scFv-Fc-84/M4 was coupled to a protein A
biosensor (Pall ForteBio). Subsequently, 4 µl standard recombinant
human (rHu) TSLP protein (5, 10 and 20 nM; Sino Biological Inc.)
was used to assess sensor interaction. By systematically analysing
antigen-antibody binding and dissociation, affinity dissociation
constant (KD) values were calculated. The interaction between
antigen (TSLP) and antibody (anti-TSLP-scFv-Fc) were analyzed by
Octet RED96 (Pall ForteBio). Acquired data were analyzed by the
custom ForteBio software Data Acquisition 9.0.
Statistical analysis
All experiments were repeated three times. Data are
expressed as the mean ± SEM. One-way ANOVA and Tukey's Multiple
Comparison Test were used to determine significance with GraphPad
Prism 5 software (GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Virtual mutation of amino acids of
anti-TSLP-scFv
Establishment and evaluation of the
TSLP homologous model
After aligning the TSLP protein sequence (NCBI Gene
ID: 85450) with the NCBI crystal library sequence, the DS 4.5
software package was used to detect the template gene sequence with
the highest homology. With homology constraints, 20 optimized
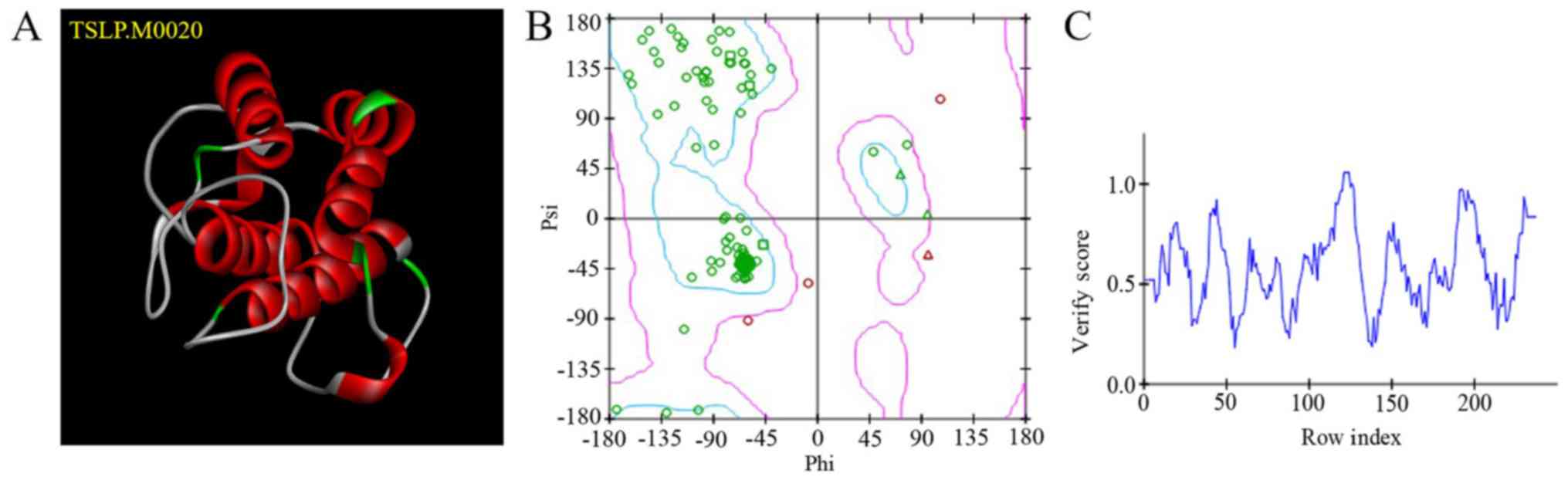
models were obtained. TSLP.M0020 (Fig.
1A) exhibited the lowest PDF total energy; this model was
evaluated using a Ramachandran plot (Fig. 1B). The blue region indicates the
optimal zone. The more amino acids (indicated by green circles) in
the optimal region, the more reliable the structure. The purple
region indicates the permitted zone. The pink circles are amino
acids with ψ-φ conformations that are unreasonable; these regions
require optimization. A Profile-3D score >0.0 indicated that the
amino acids were compatible (Fig.
1C).
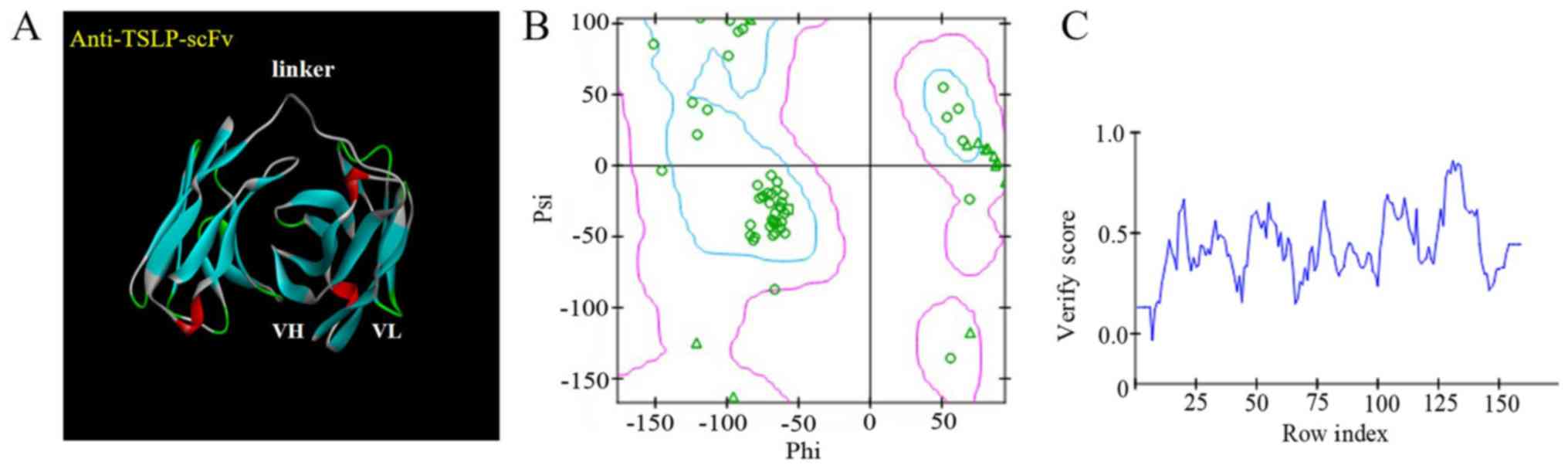
Establishment and evaluation of the
anti-TSLP-scFv-84 homologous model
The DS system predicted domains and CDRs of the
anti-TSLP-scFv-84 antibody. In total, 25 templates were obtained as
‘identify framework templates’. 4P59 LH was selected as the overall
template, 4HS6 A as the light chain template and 4KFZ C as the
heavy chain template. The model antibody framework protocol and the
model antibody loops protocol simulated models of the scFv
framework region and CDR. Finally, V_H_V_L_M0004_M. M0002 was
selected as the best 3D model through PDF total energy, PDF
physical energy and DOPE Score (Fig.
2A). This reasonable model was evaluated using a Ramachandran
plot; nearly all amino acids were within the optimal zone (blue
area). Only a few amino acids required optimization within the
permitted region (pink area; Fig.
2B).A Profile-3D score >0.0 indicated that the amino acids
were compatible (Fig. 2C).
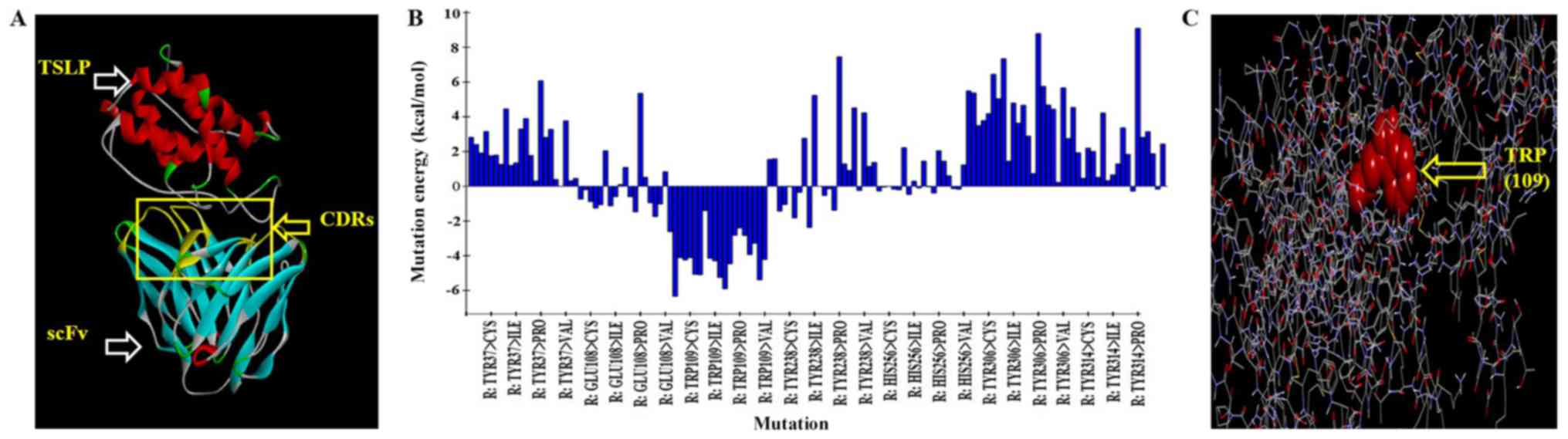
Antibody-antigen docking and virtual
mutation
Anti-TSLP-scFv-84 as the receptor and TSLP as the
ligand were docked with docking proteins. Refine docked protein
optimization was performed based on CHARMm force field energy. The
range of root mean square deviation of POSE50 was 0.2–1.0A as
measured by molecular dynamics. This result indicated that the
dynamic change of the model in solution was reasonable (Fig. 3A). The binding regions of TSLP and
anti-TSLP-scFv-84 were analyzed using the DS system and the CDRs
were mutated. In total, five key amino acid residues that could
increase antibody affinity after mutation were predicted by alanine
scanning. The five amino acid combinations were as follows: TRP
(109)-ARG, TRP (109)-LYS, TRP (109)-TYR, TRP (109)-LEU, TRP
(109)-GLU (Table SI; Fig. 3B; 109 is the number of amino acid
in the variable region), with the largest negative energy value
(<-4) identified by saturation mutagenesis and mutation energy
(Fig. 3C).
Screening and verification of
anti-TSLP-scFv mutations
Single point mutations of
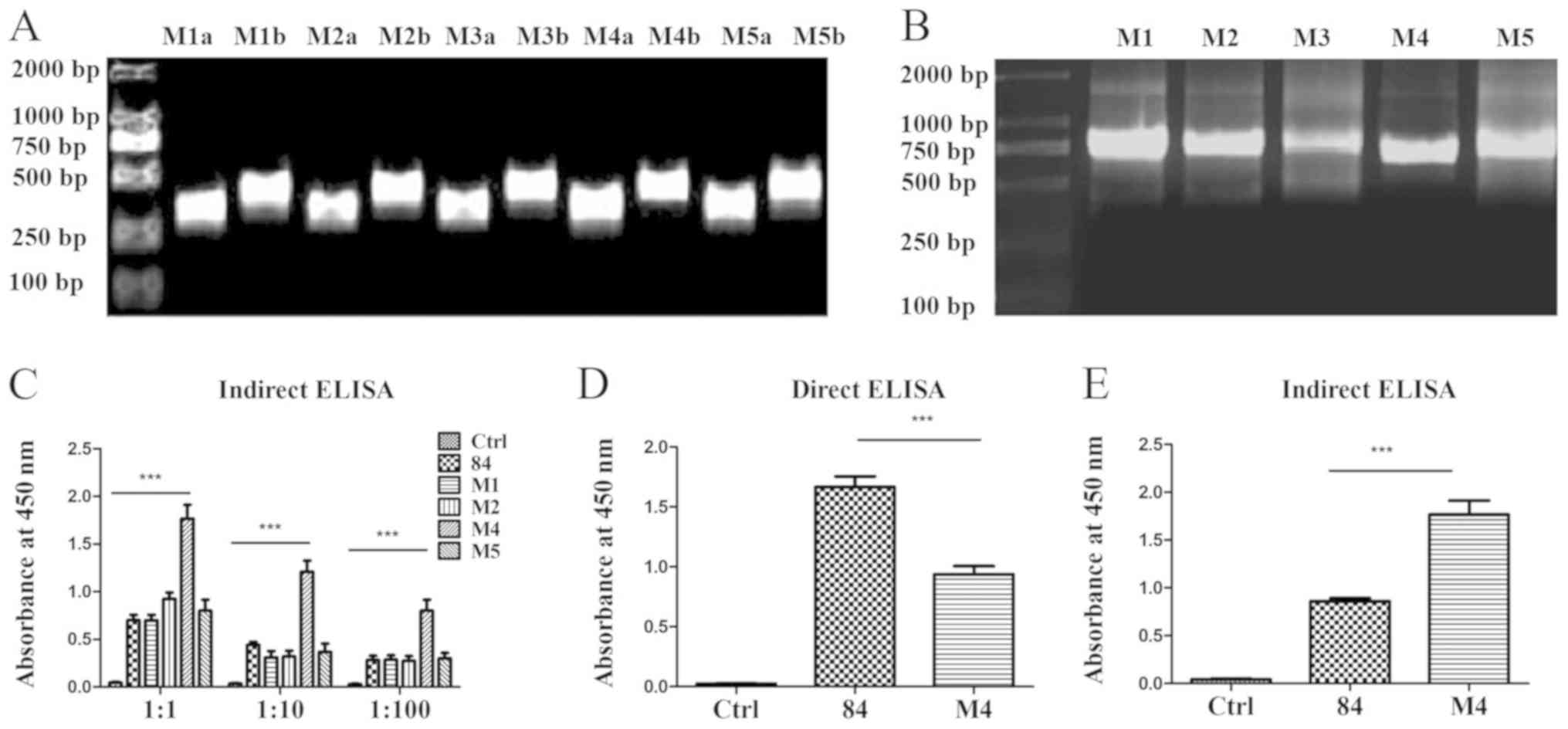
anti-TSLP-scFv assessed by PCR
The pre-mutated and DS system-predicted mutated
amino acids of scFv M1, M2, M3, M4 and M5 are presented in Table SI. Primers were designed based on
the predicted mutation sites (Table
SII) and paired to amplify the pre- and post-fragments of the
mutation sites. The fragments were connected by overlap extension
PCR. The products were 750 bp, which demonstrated that the
fragments were connected successfully (Fig. 4A and B).
Screening of mutated anti-TSLP-scFv by
expression in pLZ16, a prokaryotic expression vector
scFv M1, M2, M3, M4 and M5 were inserted into the
pLZ16 prokaryotic expression vector. The sequencing results
demonstrated that the amino acids in M1, M2, M4 and M5 were mutated
successfully; however, scFv-M3 had a stop codon (data not shown).
Therefore, scFv M1, M2, M4 and M5 were expressed in the pLZ16
prokaryotic expression vector. The target protein was detected
using HRP-labelled anti-FLAG monoclonal antibody. Indirect ELISA
results demonstrated that mutated scFv-M4 exhibited significantly
increased binding to antigen compared with the pre-mutant scFv-84
(Fig. 4C). The expression of
scFv-84/M4 was detected by coating the wells of a 96-well plate
with the expressed single-chain antibody. The results revealed that
the expression of scFv-84 was significantly higher than M4
(Fig. 4D). However, when combined
with the antigen, scFv-84 binding was significantly lower than that
of M4 (Fig. 4E), indicating
enhanced affinity for mutated M4.
Construction of the anti-TSLP-scFv-Fc
eukaryotic expression vector
Construction of the
pcDNA3.1-sp-scFv-Fc expression vector
In general, scFv have a low stability, low affinity
and a short half-life in vivo (5). Therefore, the pre-mutated scFv-84 and
affinity enhanced scFv-M4 genes were inserted into the eukaryotic
expression vector pcDNA3.1-sp-Fc. The vectors were transfected into
293F cells to express the IgG-like anti-TSLP-scFv-Fc fusion
protein. Based on the cloning site for the pcDNA3.1-sp-Fc vector,
specific primers were designed and the scFv gene fragment of the
single-chain antibody was amplified using the ExTaq enzyme. A
single band of 750 bp was detected by agarose gel electrophoresis
of the PCR product that contained scFv (Fig. S2A). After digestion with
KpnI and BamHI, anti-TSLP-scFv was ligated into the
pcDNA3.1-sp-Fc vector with T4 ligase. The results demonstrated that
scFv-84 and scFv-M4 were recombined with pcDNA3.1-sp-Fc
successfully (Fig. S2B).
Construction of the PMH3
EN-sp-scFv-Fc expression vector
As the expression level of scFv-Fc in pcDNA3.1 was
low (Fig. 5A and B), the
recombinant plasmid PMH3EN was used to improve
expression. The pcDNA3.1 vector was digested with HindIII
and NotI. Expression of sp-scFv-Fc-84/M4 was detected by
agarose gel electrophoresis (Fig.
S3A). Specific primers were designed based on the cloning site
of the PMH3EN vector and the anti-TSLP-scFv-Fc sequence.
Empty plasmid PMH3EN was cleaved with HindIII and
NotI. Then sp-scFv-Fc was ligated into the PMH3EN
vector (Fig. S3B). Sequencing
results demonstrated that the full-length sp-scFv-Fc had been
inserted into all clones.
Verification of anti-TSLP-scFv-Fc
affinity
Expression of anti-TSLP-scFv-Fc
antibody
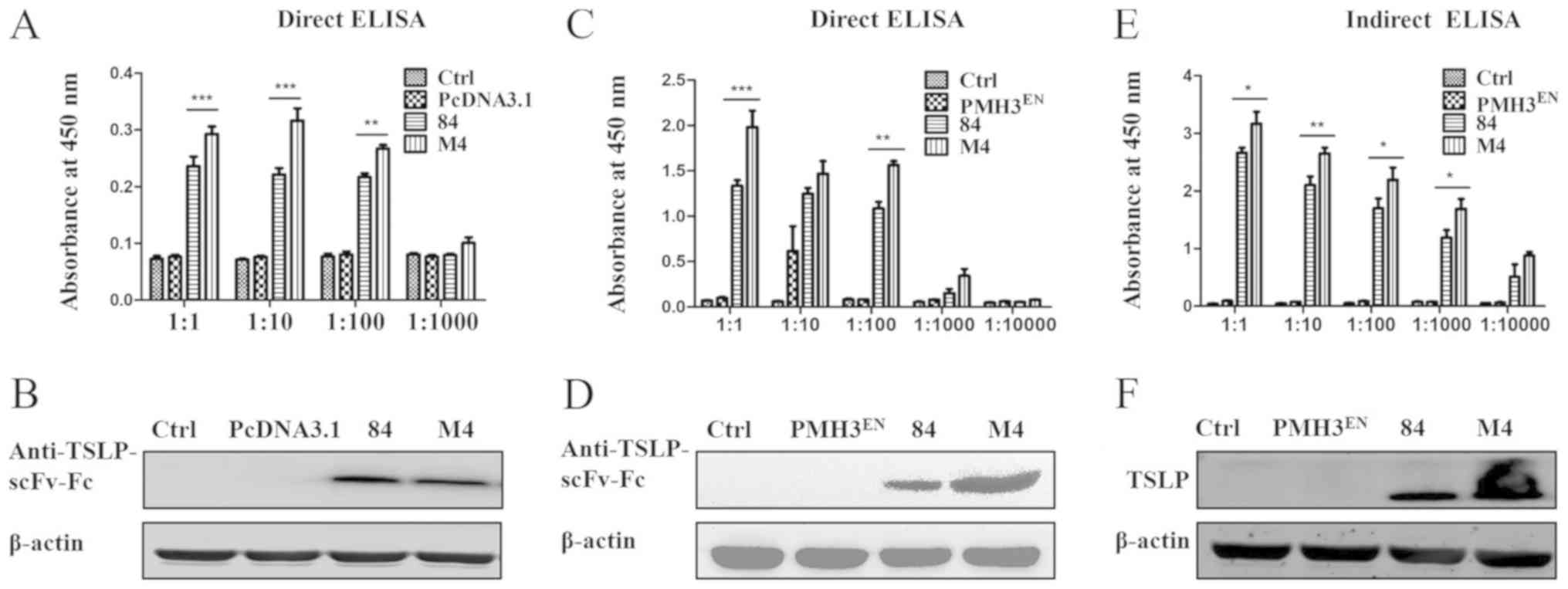
The expression of PMH3EN-scFv-Fc-84/M4
was detected by direct ELISA, which was performed using an
anti-TSLP HRP-conjugated antibody. The expression of
anti-TSLP-scFv-Fc was not detected from the control and empty
plasmid PMH3EN; however, PMH3EN-scFv-Fc-84/M4
expression was obvious. Furthermore, expression of
PMH3EN-scFv-Fc-M4 was significantly higher than
PMH3EN-scFv-Fc-84 (P<0.05; Fig. 5C). In addition, the expressed
anti-TSLP-scFv-Fc protein was identified by western blotting. Using
anti-human IgG Fc HRP-conjugated antibody, a single band of 50 kDa
was identified (Fig. 5D).
Binding activity of the
anti-TSLP-scFv-Fc antibody
Indirect ELISA was used to assess the affinity of
the antibody for the antigen. ELISA plates were coated with TSLP
and incubated with anti-TSLP-scFv-Fc. An anti-TSLP HRP-conjugated
antibody was used to assess the binding of
PMH3EN-scFv-Fc-84/M4 to TSLP (Fig. 5E). ELISA results indicated that the
antibodies bound TSLP and that the binding of
PMH3EN-scFv-Fc-M4 was higher than that of
PMH3EN-scFv-Fc-84. Western blotting revealed that the
TSLP protein bands specific binding with recombinant
PMH3EN-scFv-Fc-84/M4 was ~25 kDa (Fig. 5F). A positive association between
ELISA and western blotting data confirmed the validity of the
results.
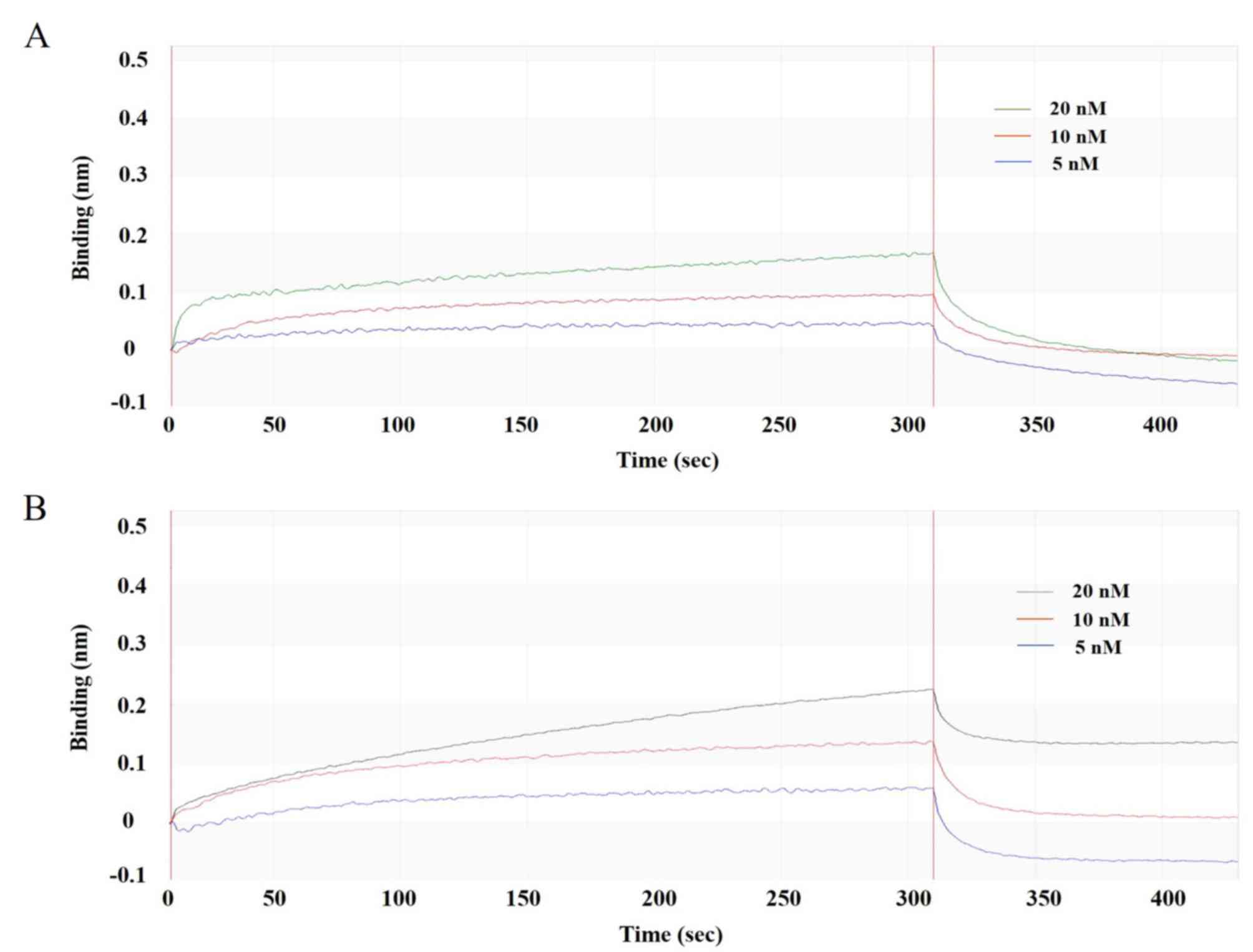
Affinity of the anti-TSLP-scFv-Fc
antibody
To verify that the affinity of mutated scFv-Fc-M4
was higher after mutation, PMH3EN-scFv-Fc-84/M4 was
captured with an anti-Fc biosensor and evaluated with different
concentrations of standard rHuTSLP protein. By analysing the
process of binding and dissociation (Fig. 6A and B), the affinity KD of
PMH3EN-scFv-Fc-M4 was calculated to be
3.21×10−9, which was ~10-fold higher than before the
mutation. The product was biologically active and retained high
binding affinity to the antigen, as detailed by BIAcore real-time
interaction analysis.
Discussion
A number of previous studies have demonstrated an
important role for TSLP in different diseases (17,18).
Blocking the binding of TSLP to TSLPR may inhibit TSLP signalling,
which may provide a therapeutic strategy to treat various diseases
(19). The present study
successfully constructed an anti-TSLP-scFv-Fc recombinant antibody.
Furthermore, the affinity of the antibody was improved in
vitro by homology modelling, molecular docking, virtual
mutation and selection of mutation sites. Notably, the affinity of
post-mutation anti-TSLP-scFv-Fc increased 10-fold.
With the development of hybridoma technology, mass
spectrometry engineering, phage display and transgenic animal
technology, the preparation of antibodies has successively
transitioned from polyclonal to monoclonal antibodies (20,21).
A priority has been the humanization of murine antibodies by
genetic engineering, in order to obtain a fully humanized
monoclonal antibody with high affinity, stability and biological
activity (22,23). Using phage display technology,
human ethical issues have been avoided and large numbers of human
monoclonal antibodies can be prepared (24). However, these antibodies typically
have a low affinity; therefore, in vitro genetic engineering
is required to enhance their affinity.
In our previous study, a fully human display library
was screened for anti-TSLP-scFv-Fc-84 that had high specificity and
the capacity to block the TSLPR in vitro (25). However, single-chain antibodies
screened from natural antibody libraries have low affinity and
require affinity improvement (5).
Random mutation of gene fragments encoding variable regions and
directed introduction of mutations are the primary methods for
increasing the affinity of recombinant antibodies (26). With the former, mutations are
random, unpredictable and time-consuming. However, Lewis et
al (27) successfully used
alanine scanning to select targeted and random mutations. In the
present study, antibody affinity was increased ~10-fold by
replacing one amino acid. Alanine scanning mutagenesis was used to
identify CDR sites that increased affinity, specificity and
stability (28). In the present
study, three-dimensional models of TSLP and anti-TSLP-scFv were
obtained by homology modelling using the DS 4.5 software. CDR amino
acids were then alanine scanned and saturated mutations made to key
amino acids. The mutated gene sequence was recombined into the
prokaryotic expression vector pLZ16. Mutated M4 was screened using
ELISA and found to have an increased affinity for the antigen.
Anti-TSLP-scFv has a short half-life, is easily
dimer inactivated, and without an Fc fragment cannot function in
antibody-dependent cell-mediated cytotoxicity (29,30).
These drawbacks limit applications of anti-TSLP-scFv as a drug for
clinical treatment. Moreover, natively folded proteins can be
produced by mammalian expression systems; however, they cannot be
produced by bacterial expression systems (13). However, 293F cells can be cultured
in suspensions and used as a mammalian expression system (31). The polymeric reagent PEI is an
inexpensive and highly efficient transfection reagent. PEI can be
used to efficiently and quickly transfect 293F cells, yielding high
levels of recombinant protein (14). Proteins produced by 293F cells fold
correctly and are biologically active (14). Therefore, the gene sequences of
anti-TSLP-scFv (pre-mutated 84 and mutated M4) were cloned into the
eukaryotic expression vectors pcDNA3.1-sp-Fc and
PMH3EN-sp-Fc. 293F cells were transfected with these
vectors and IgG-like recombinant antibodies of anti-TSLP-scFv-Fc
were obtained. The size of anti-TSLP-scFv-Fc, as determined by
western blotting, was ~50 kDa. The affinity of the antibodies was
determined using the BIAcore technique, the affinity of mutated M4
was found to be ~10-fold higher than the pre-mutated antibody.
At present, there is no anti-TSLP antibody approved
by the US Food and Drug Administration (32). As a therapeutic antibody, affinity
is the most important characteristic (33). In the present study, an
anti-TSLP-scFv-Fc recombinant antibody was successfully constructed
and its affinity was increased using DS 4.5 software. This method
for improving antibody affinity is stable, efficient and
inexpensive.
The present study has some limitations. In our
previous study, anti-TSLP-scFvs were derived from a naïve fully
human scFv library. In vitro experiments demonstrated that
scFv-84 neutralized TSLP, however, with low affinity. In the
present study, the affinity of scFv-84 was enhanced and an scFv-Fc
was constructed. In future studies, anti-TSLP-scFv-Fc antibody will
be expressed and purified for use in animal models of asthma as a
strategy for disease therapy.
In summary, the present study successfully
constructed a fully human anti-TSLP-scFv-Fc recombinant antibody of
50 kDa. The affinity of mutated anti-TSLP-scFv-Fc-M4 was enhanced
10-fold compared with the pre-mutated antibody.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Department of Luzhou City (grant nos. 2017LZXNYDT06,
LY84 and 2018LZXNYD-ZK26), the Innovation Team Project of the
Sichuan Education Department (grant no. 16TD0022) and Health
Commission of Sichuan Province (grant no. 19PJ291).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QC performed data analysis and wrote the manuscript.
DX and WX performed Discovery Studio software analysis. SN, HY and
YW contributed significantly in the interpretation and analysis of
the data. QY conceived and designed the study, and revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Friend SL, Hosier S, Nelson A, Foxworthe
D, Williams DE and Farr A: A thymic stromal cell line supports in
vitro development of surface IgM+ B cells and produces a novel
growth factor affecting B and T lineage cells. Exp Hematol.
22:321–328. 1994.PubMed/NCBI
|
|
2
|
Tatsuno K, Fujiyama T, Yamaguchi H, Waki M
and Tokura Y: TSLP directly interacts with skin-homing Th2 cells
highly expressing its receptor to enhance IL-4 production in atopic
dermatitis. J Invest Dermatol. 135:3017–3024. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Park JH, Jeong DY, Peyrin-Biroulet L,
Eisenhut M and Shin JI: Insight into the role of TSLP in
inflammatory bowel diseases. Autoimmun Rev. 16:55–63. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kuan EL and Ziegler SF: A tumor-myeloid
cell axis, mediated via the cytokines IL-1α and TSLP, promotes the
progression of breast cancer. Nat Immunol. 19:366–374. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Manoutcharian K, Perez-Garmendia R and
Gevorkian G: Recombinant antibody fragments for neurodegenerative
diseases. Curr Neuropharmacol. 15:779–788. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shehata L, Maurer DP, Wec AZ, Lilov A,
Champney E, Sun T, Archambault K, Burnina I, Lynaugh H, Zhi X, et
al: Affinity maturation enhances antibody specificity but
compromises conformational stability. Cell Rep. 28:3300–3308.e4.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakamura T, Motoyama T, Hirokawa T, Hirono
S and Yamaguchi I: Computer-aided modeling of pentachlorophenol
4-monooxygenase and site-directed mutagenesis of its active site.
Chem Pharm Bull (Tokyo). 51:1293–1298. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen JM, Grad R, Monaco R and Pincus MR:
Prediction of the three-dimensional structure of the rap-1A protein
from its homology to the ras-gene-encoded p21 protein. J Protein
Chem. 15:11–15. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yuan Q, Huang L, Wang X, Wu Y, Gao Y, Li C
and Nian S: Construction of human nonimmune library and selection
of scFvs against IL-33. Appl Biochem Biotechnol. 167:498–509. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sircar A: Methods for the homology
modeling of antibody variable regions. Methods Mol Biol.
857:301–311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nian S, Wu T, Ye Y, Wang X, Xu W and Yuan
Q: Development and identification of fully human scFv-Fcs against
Staphylococcus aureus. BMC Immunol. 17:82016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ye Y, Nian S, Xu W, Wu T, Wang X, Gao Y
and Yuan Q: Construction and expression of human scFv-Fc against
interleukin-33. Protein Expr Purif. 114:58–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nigi I, Fairall L and Schwabe JWR:
Expression and purification of protein complexes suitable for
structural studies using mammalian HEK 293F cells. Curr Protoc
Protein Sci. 90:5.28.1–5.28.16. 2017. View
Article : Google Scholar
|
|
14
|
Portolano N, Watson PJ, Fairall L, Millard
CJ, Milano CP, Song Y, Cowley SM and Schwabe JW: Recombinant
protein expression for structural biology in HEK 293F suspension
cells: A novel and accessible approach. J Vis Exp.
e518972014.PubMed/NCBI
|
|
15
|
Molek P, Vodnik M, Strukelj B and
Bratkovič T: Screening of synthetic phage display scFv libraries
yields competitive ligands of human leptin receptor. Biochem
Biophys Res Commun. 452:479–483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan Q, Wang Z, Nian S, Yin Y, Chen G and
Xia Y: Screening of high-affinity scFvs from a ribosome displayed
library using BIAcore biosensor. Appl Biochem Biotechnol.
152:224–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kahramanoğlu Aksoy E, Akpınar MY, Pirinççi
Sapmaz F, Doğan Ö, Uzman M and Nazlıgül Y: Thymic stromal
lymphopoietin levels are increased in patients with celiac disease.
Bosn J Basic Med Sci. 19:282–287. 2019.PubMed/NCBI
|
|
18
|
Tahaghoghi-Hajghorbani S, Ajami A,
Ghorbanalipoor S, Hosseini-Khah Z, Taghiloo S, Khaje-Enayati P and
Hosseini V: Protective effect of TSLP and IL-33 cytokines in
ulcerative colitis. Auto Immun Highlights. 10:12019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Borowski A, Vetter T, Kuepper M, Wohlmann
A, Krause S, Lorenzen T, Virchow JC, Luttmann W and Friedrich K:
Expression analysis and specific blockade of the receptor for human
thymic stromal lymphopoietin (TSLP) by novel antibodies to the
human TSLPRα receptor chain. Cytokine. 61:546–555. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yazdi IK, Taghipour N, Hmaidan S, Palomba
R, Scaria S, Munoz A, Boone TB and Tasciotti E: Antibody-mediated
inhibition of Nogo-A signaling promotes neurite growth in PC-12
cells. J Tissue Eng. 7:20417314166297672016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hafeez U, Gan HK and Scott AM: Monoclonal
antibodies as immunomodulatory therapy against cancer and
autoimmune diseases. Curr Opin Pharmacol. 41:114–121. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moutel S, Bery N, Bernard V, Keller L,
Lemesre E, de Marco A, Ligat L, Rain JC, Favre G, Olichon A and
Perez F: NaLi-H1: A universal synthetic library of humanized
nanobodies providing highly functional antibodies and intrabodies.
Elife. 5:e162282016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Townsend S, Fennell BJ, Apgar JR, Lambert
M, McDonnell B, Grant J, Wade J, Franklin E, Foy N, Ní
Shúilleabháin D, et al: Augmented binary substitution: Single-pass
CDR germ-lining and stabilization of therapeutic antibodies. Proc
Natl Acad Sci USA. 112:15354–15359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rodgers KR and Chou RC: Therapeutic
monoclonal antibodies and derivatives: Historical perspectives and
future directions. Biotechnol Adv. 34:1149–1158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nian S, Zhu J, Yu H, Chen Q, Ye Y, Cao X
and Yuan Q: Development and identification of a fully human
single-chain variable fragment 29 against TSLP. Biotechnol Appl
Biochem. 66:510–516. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zahnd C, Spinelli S, Luginbühl B, Amstutz
P, Cambillau C and Plückthun A: Directed in vitro evolution and
crystallographic analysis of a peptide-binding single chain
antibody fragment (scFv) with low picomolar affinity. J Biol Chem.
279:18870–18877. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lewis CM, Hollis GF, Mark GE III, Tung JS
and Ludmerer SW: Use of a novel mutagenesis strategy, optimized
residue substitution, to decrease the off-rate of an anti-gp120
antibody. Mol Immunol. 32:1065–1072. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tiller KE, Chowdhury R, Li T, Ludwig SD,
Sen S, Maranas CD and Tessier PM: Facile affinity maturation of
antibody variable domains using natural diversity mutagenesis.
Front Immunol. 8:9862017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang YF and Ho M: Humanization of
high-affinity antibodies targeting glypican-3 in hepatocellular
carcinoma. Sci Rep. 6:338782016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahmed M, Cheng M, Zhao Q, Goldgur Y, Cheal
SM, Guo HF, Larson SM and Cheung NK: Humanized affinity-matured
monoclonal antibody 8H9 has potent antitumor activity and binds to
FG loop of tumor antigen B7-H3. J Biol Chem. 290:30018–30029. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Subedi GP, Johnson RW, Moniz HA, Moremen
KW and Barb A: High yield expression of recombinant human protein
with the transient transfection of HEK293 cells in suspension. J
Vis Exp. e535682015.PubMed/NCBI
|
|
32
|
Mitchell PD, El-Gammal AI and O'Byrne PM:
Emerging monoclonal antibodies as targeted innovative therapeutic
approaches to asthma. Clin Pharmacol Ther. 99:38–48. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cohan SL, Lucassen EB, Romba MC and Linch
SN: Daclizumab: Mechanisms of action, therapeutic efficacy, adverse
events and its uncovering the potential role of innate immune
system recruitment as a treatment strategy for relapsing multiple
sclerosis. Biomedicines. 7:E182019. View Article : Google Scholar : PubMed/NCBI
|