Introduction
Breast cancer is the most common type of cancer in
women worldwide, and it is estimated that 1.4 million women a year
receive a diagnosis of breast cancer, while 458,000 succumb to the
disease (1). Breast cancer
involves a multi-step process associated with the abnormal
expression of several oncogenes and tumor suppressor genes
(2). Numerous types of malignant
tumors are associated with the abnormal expression of Ras family
genes (3). A more comprehensive
understanding of the Ras oncogene pathways could lead to improved
strategies for therapy and prevention.
SHOC2 leucine rich repeat scaffold protein (SHOC2)
was first isolated in Caenorhabditis elegans (4) and was identified as a positive
regulator of the Ras pathway (5).
SHOC2 is a scaffold protein that binds components of the Ras
pathway and modulates their functions (6,7). Ras
binds directly to SHOC2 to form a complex that binds to the
catalytic subunit of protein phosphatase 1 (PP1c); this
Ras/SHOC2/PP1c complex activates the Ras pathway by
dephosphorylating Raf-1 proto-oncogene, serine/threonine kinase
(Raf-1) at S259 (8). SHOC2 has
been reported to regulate the Ras signaling cascade in a number of
ways; in particular, it has essential roles in embryogenesis and
normal biological processes (9,10).
Furthermore, the role of SHOC2 in various types of malignant cells
has been examined, which has highlighted its potential role in
tumorigenesis (11). Lee et
al (12) demonstrated that
SHOC2 promoted colorectal tumorigenesis and metastasis via ERK and
PI3K-AKT signaling pathways. Kaduwal et al (13) suggested that SHOC2 also served an
essential role in the process of metastasis via the
Ras-PI3K-Rac-matrix metalloproteinase signaling pathway in human
melanoma. In addition, Kaplan et al (14) demonstrated that SHOC2 was
associated with the mechanisms of acquired resistance to the Raf
inhibitor vemurafenib. In this previous study, SHOC2 could alter
signaling connections and re-route oncogenic Ras signals to Raf-1
in order to mediate reactivation of the ERK1/2 pathway and
facilitate drug resistance in cells (14). SHOC2 has also been shown to be a
positive regulator that contributes to the malignant properties of
different tumor cells by modulating the growth, transformation,
migration and invasion of cancer cells (10,15,16).
Our preliminary experiments revealed that SHOC2 was highly
expressed in MCF-7 and MDA-MB-231 breast cancer cells (data not
shown). However, the function of SHOC2 in breast cancer has rarely
been explored. In this study, the association between SHOC2
upregulation and the clinicopathological features of breast cancer
was investigated, and the prognostic value of SHOC2 was evaluated.
Furthermore, the effects of SHOC2 on growth, apoptosis and cell
cycle progression in breast cancer were elucidated.
Materials and methods
Patients and tissues
A total of 120 patients with breast cancer who
underwent surgical treatment and had complete tumor tissue blocks
preserved at the Department of Breast Surgery, Qilu Hospital of
Shandong University (Jinan, China) between January 2004 and May
2016 were recruited. All 120 patients were female, and their ages
ranged between 26 and 72 years old, with an average age of 46.3±9.9
years old. The inclusion criteria were as follows: Diagnosed with
breast cancer based on the post-operative pathological section;
completed clinical follow-up survey; >18 years old; and never
received pre-operative chemotherapy or radiotherapy. The exclusion
criteria were as follows: <18 years old; had incomplete clinical
follow-up data or other malignant tumors at the same time; or
received pre-operative chemotherapy or radiotherapy. Patients all
underwent surgery: 88 had modified radical mastectomy (73%) and 32
had breast-conserving therapy (27%). After surgery, the patients
were treated with chemotherapy or radiotherapy. Indications for
post-operative chemotherapy, radiotherapy or chemoradiotherapy were
patients with various pathological features, such as advanced
cancer stage, lymph node positivity, perineural invasion,
lymphovascular invasion and receptor status. Patients positive for
hormone receptors received adjuvant endocrine therapy for 5 years
and patients positive for human epidermal growth factor receptor 2
(HER2) received trastuzumab therapy for 1 year. After finishing
treatment, patients were regularly followed-up with clinical
examinations and imaging. Patients were scheduled for clinical
visits every 4–6 months during the first and second years, every 6
months during the third, fourth and fifth years, and annually
thereafter.
Immunohistochemistry (IHC)
Formalin-fixed (10% neutral buffered formalin at 4°C
for 12–24 h at room temperature), paraffin-embedded sections (size,
4 µm) from breast cancer and normal breast tissue samples were
obtained. The normal tissue samples were obtained from the adjacent
tissue of the same patients, and the distance between tumor and
normal tissues was ≥0.5 cm (17).
The sections were deparaffinized, rehydrated in a descending
alcohol series and heated at 120°C for 10 min in 10 mM sodium
citrate (pH 6.0) to retrieve antigens. Endogenous peroxidase
activity was quenched with 3% hydrogen peroxide at room temperature
for 10 min. The sections were then incubated with an anti-SHOC2
antibody (1:400; cat. no. Fnab06912; Wuhan Fine Biotech Co., Ltd.)
at room temperature for 1 h, followed by a horseradish peroxidase
(HRP)-conjugated secondary antibody (cat. no. 8114; Cell Signaling
Technology, Inc.) at room temperature for 15 min. The sections were
developed in diaminobenzidine for 5 min and counterstained with
hematoxylin for 3 min at room temperature. The primary antibody was
omitted for the negative controls. Sections were visualized and
photographed using an optical microscope (Leica Microsystems GmbH;
magnification, ×50 and ×200). The expression levels of SHOC2 were
semi-quantified using a semi-quantitative IHC scoring system, as
previously described (18,19). The percentage of positive tumor
cells was graded on a scale between 0 and 4, as follows: 0, none;
1, 1–10%; 2, 11–50%; 3, 51–80%; 4, >80%. The intensity of
staining was graded on a scale between 0 and 3, as follows: 0,
none; 1, weak staining; 2, moderate staining; 3, strong staining.
The combination of the extent (E) and intensity (I) of staining was
obtained as the product of E and I (EI), which varied between 0 and
12 for each sample. Using the X-tile software program (version
3.6.1; The Rimm Lab, Yale University; http://medicine.yale.edu/lab/rimm/research/software.aspx),
a significant cutoff point for SHOC2 was identified in terms of
overall survival (OS) in patients with breast cancer, as previously
described (20). A cutoff score of
8 was selected; 0–8 was considered low expression, whereas >8
was considered high expression.
Cell culture and infection
MCF-7 and MDA-MB-231 breast cancer cell lines and
the 293T cell line were purchased from The Cell Bank of Type
Culture Collection of the Chinese Academy of Sciences and
maintained by the Center Lab of Shandong University. Cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 100 U/ml penicillin-streptomycin, and grown at 37°C in 5%
CO2. Stable knockdown of SHOC2 in the breast cancer cell
line was performed using lentiviruses. Briefly, the target sequence
(5′-TGCTTGATTTACGGCATAA-3′) for short hairpin RNA (shRNA)-SHOC2 and
the nonspecific control sequence (5′-TTCTCCGAACGTGTCACGT-3′)
(21) were inserted into the GV493
shRNA vector (Shanghai GeneChem Co., Ltd.) linearized with
AgeI and EcoRI. For GFP expression in this vector,
GFP-IRES was cloned in front of the puromycin-resistant marker
gene. Virus production was carried out using 293T cells. Briefly,
cells were transfected with lentiviral DNA constructs alongside the
lentiviral packaging plasmids pHelper 1.0 and pHelper 2.0 (Shanghai
GeneChem Co., Ltd.) at a ratio of 4:3:2. The viral supernatants
were harvested 48 h post-transfection, filtered using a 0.45-µm
pore filter, and used for infection. To establish stable cell
lines, cells were seeded in 6-well plates (2×105/well)
until they reached 60–70% confluence before being infected with
either SHOC2 knockdown or the control lentivirus (MOI=10), and
selected with puromycin (2 µg/ml) to obtain single-cell clones. The
single-cell clones obtained were cultured for 2 weeks, and
amplified for further experiments in culture media containing
puromycin (1 µg/ml).
Reverse transcription-quantitative
(RT-q)PCR
RT-qPCR was performed to detect the mRNA expression
levels of SHOC2 in 11 pairs of fresh breast cancer and adjacent
noncancerous human breast tissues. Total RNA was isolated from
breast cancer and adjacent noncancerous tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Random-primed cDNA synthesis was performed using a
QuantiTect® RT kit (Qiagen GmbH). The RT reaction was
performed as follows: Initial incubation at 42°C for 2 min,
followed by sequential steps at 42°C for 15 min, and at 95°C for 3
min. RT-qPCR was performed using a PTC-100® Thermal
cycler (MJ Research, Inc.) with a PrimeScript™ RT Reagent kit
(Takara Biotechnology Co., Ltd.), according to the manufacturer's
protocols. For qPCR, the thermocycling conditions were:
Pre-denaturation at 95°C for 30 sec, followed by 40 cycles of
denaturation at 95°C for 5 sec and of annealing at 60°C for 30–60
sec. For amplification, the following primers were designed: SHOC2,
forward 5′-TCAGTGGTGTATAGGCTGGATTCT-3′, reverse
5′-GCTACATCCAGCGTAATGAGGT-3′; GAPDH, forward
5′-CCTCCGGGAAACTGTGGCGTGATGG-3′ and reverse
5′-AGACGGCAGGTCAGGTCCACCACTG-3′. Each sample was examined three
times, and the levels of PCR product were adjusted to GAPDH, which
served as an internal control. The fold change of mRNA expression
was calculated using the formula: 2−ΔΔCq (22).
Western blot analysis
Confluent cells were washed twice with ice-cold PBS
and lysed in RIPA buffer with 1 mM PMSF (Beyotime Institute of
Biotechnology). The concentrations of the total protein extracts
were determined by the bicinchoninic acid (BCA) method using a
commercial kit from Pierce; Thermo Fisher Scientific, Inc. Equal
amounts of protein samples (40 µg) were separated by SDS-PAGE on
10% gels and transferred onto PVDF membranes. After blocking with
5% non-fat milk at room temperature for 1 h, the membranes were
incubated overnight at 4°C with primary antibodies against SHOC2
(1:1,000; cat. no. Fnab06912; Wuhan Fine Biotech Co., Ltd.), Bcl-2
(1:1,000; cat. no. 15071; Cell Signaling Technology, Inc.), Bax
(1:1,000; cat. no. 5023; Cell Signaling Technology, Inc.), poly
(ADP-ribose) polymerase (PARP) (1:1,000; cat. no. 9532; Cell
Signaling Technology, Inc.), phosphorylated (p)-AKT (1:1,000; cat.
no.4060; Cell Signaling Technology, Inc.), AKT (1:1,000; cat. no.
4691; Cell Signaling Technology, Inc.), p-ERK (1:1,000; cat. no.
4376; Cell Signaling Technology, Inc.), ERK (1:1,000; cat. no.
4696; Cell Signaling Technology, Inc.) and GAPDH (1:1,000; cat. no.
AP7873a; Abgent, Inc.), followed by incubation with either
HRP-conjugated Anti-Rabbit secondary antibodies (1:5,000; cat. no.
R4880; Sigma-Aldrich; Merck KGaA) or Anti-Mouse secondary
antibodies (1:5,000; cat. no. 7077; Cell Signaling Technology,
Inc.) at room temperature for 1 h. Protein bands were detected by
ECL western blot substrate (Thermo Fisher Scientific, Inc.).
Immunoreactive bands were visualized by ImageQuant LAS 4000 series
(GE Healthcare). Image J v.1.43 (National Institutes of Health)
software was used to quantify relative protein expression.
MTT assay
Cells (5 ×103/well) were seeded and
incubated in 96-well plates at 37°C in a 5% CO2
environment for 1, 2, 3, 4 and 5 days. At each time point, 5 mg/ml
MTT was added to each well. After incubation at 37°C for 4 h, the
supernatants were removed. Subsequently, 100 µl DMSO was added to
each well, and the well contents were thoroughly mixed for 5 min.
The absorbance was measured at 490 nm using a microplate reader
(TECAN Infinite® 200; Tecan Group, Ltd.).
Cell cycle and apoptosis analysis
Cells were cultured to 85% confluence, collected and
then rinsed with cold D-Hanks balanced salt solution. After
centrifugation at 1,000 × g for 5 mins at room temperature, the
supernatants were removed, and the cells were fixed in 75% cold
ethanol for 1 h. The fixed cells were rinsed with D-Hanks solution
and permeabilized with 0.1% Triton X-100 and 2 mg/ml RNase A in
D-Hanks solution for 30 min at 37°C. The cells were then rinsed
with D-Hanks solution and stained with 50 mg/ml propidium iodide
(Sigma-Aldrich; Merck KGaA) at 4°C for 1 h. The stained cells were
analyzed with a Millipore Guava® easyCyte 5HT flow
cytometer (EMD Millipore). A total of 8 days after lentiviral
infection, cells (5×105) were collected and rinsed with
cold D-Hanks balanced salt solution. After centrifugation at 1,000
× g for 3 mins at room temperature, the supernatants were removed.
The cells were resuspended with 200 µl binding buffer and then
stained with 10 µl Annexin V-APC (cat. no. 88-8007; eBioscience;
Thermo Fisher Scientific, Inc.) at room temperature in the dark for
10–15 min. Subsequently, flow cytometry was performed using the
Guava easyCyte 5HT flow cytometer. Data were analyzed using FlowJo
software v.10 (FlowJo LLC).
Statistical analysis
All results were repeated three times and the data
are presented as the mean ± standard deviation. The association
among SHOC2 expression, proliferation and apoptosis was assessed
using unpaired Student's t-test. The mRNA expression was compared
between normal and tumor tissues using paired Student's t-test. IHC
data were compared between normal and tumor tissues using Wilcoxon
signed-rank test. The relationship between SHOC2 expression and
clinicopathological characteristics was analyzed by the
χ2 test. Survival curves were plotted by the
Kaplan-Meier method and compared using the log-rank test. Survival
data were evaluated using univariate and multivariate Cox
regression analyses. All statistical analyses were performed using
SPSS version 18.0 (SPSS, Inc.). P<0.05 was considered to be
statistically significant.
Results
SHOC2 knockdown impairs breast cancer
proliferation, and induces apoptosis and cell cycle arrest
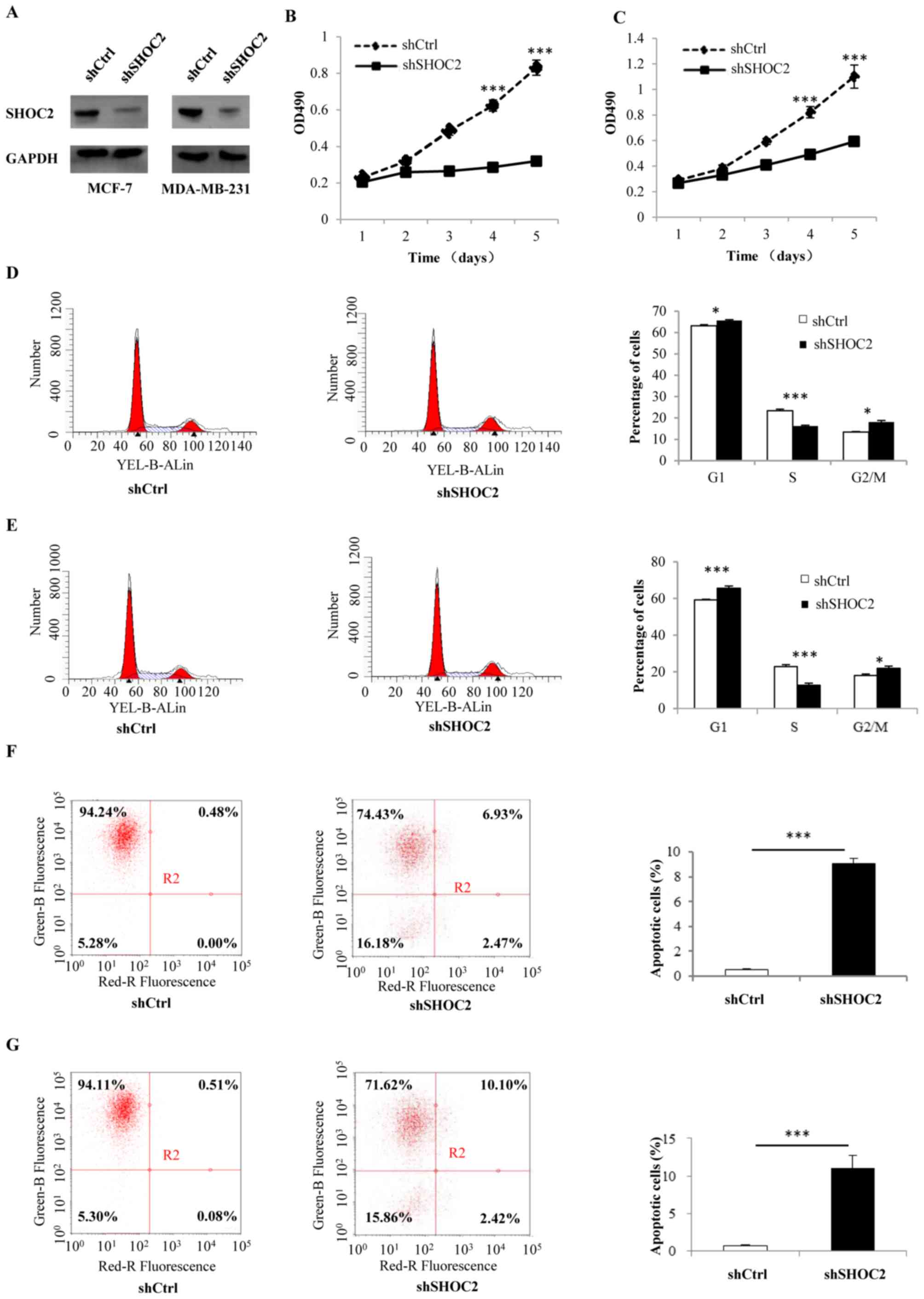
MCF-7 and MDA-MB-231 breast cancer cells were
infected with shRNA-SHOC2 to induce SHOC2 knockdown (Fig. 1A). Subsequently, MTT assays were
performed to determine the effects of SHOC2 on breast cancer cell
proliferation, which revealed that cell counts were significantly
decreased by SHOC2 knockdown. Therefore, SHOC2 knockdown may
inhibit the growth of MCF-7 and MDA-MB-231 breast cancer cells
(P<0.001; Fig. 1B and C). The
mechanism underlying the antiproliferative effects of SHOC2
knockdown was examined by analyzing apoptosis using Annexin V
staining. The number of Annexin V-positive cells was significantly
higher in shRNA-SHOC2 breast cancer cells than in control cells
(P<0.001; Fig. 1F and G).
Moreover, cell cycle distribution was analyzed using flow
cytometry, which showed that SHOC2 knockdown led to cell arrest in
the G1 and G2/M phases (P<0.05; Fig. 1D and E). Next, a series of cellular
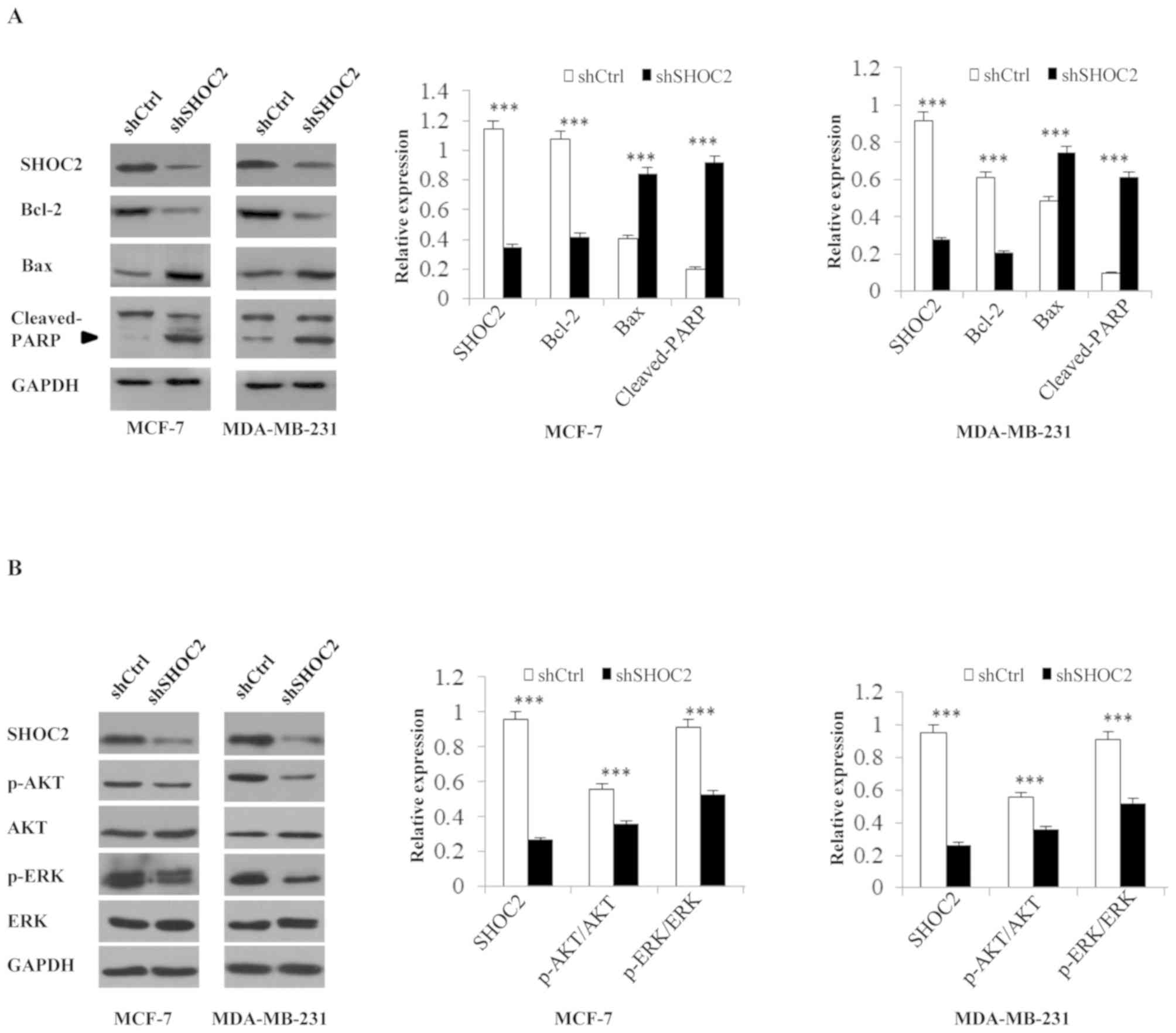
apoptosis-related proteins were analyzed; the expression levels of
the anti-apoptosis marker Bcl-2 were significantly decreased,
whereas the levels of the pro-apoptotic markers Bax and
cleaved-PARP were increased in shRNA-SHOC2 cells (P<0.001;
Fig. 2A). These results indicated
that knockdown of SHOC2 significantly inhibited proliferation, and
promoted apoptosis and cell cycle arrest in breast cancer
cells.
The RAS-MAPK/PI3K pathway is inhibited
by SHOC2 knockdown
Analysis of RAS-MAPK/PI3K pathway-associated protein
expression revealed that after SHOC2 was knocked down in MCF-7 and
MDA-MB-231 breast cancer cells, p-ERK and p-AKT expression was
decreased. These findings indicated that the Ras-ERK and PI3K-AKT
pathways were inhibited by SHOC2 knockdown, as revealed by the
decrease in positive signals for p-ERK and p-AKT (P<0.001;
Fig. 2B).
SHOC2 is overexpressed in human breast
cancer tissues
To explore the roles of SHOC2 in human breast
cancer, SHOC2 mRNA was evaluated in fresh clinical samples by
RT-qPCR. SHOC2 expression was significantly higher in human breast
cancer tissues than in paired normal breast tissues (P<0.01;
Fig. 3A). In the IHC analysis,
SHOC2 protein was predominantly found in the cytoplasm of tumor
cells, and this demonstrated that 88 of 120 (73.3%) breast cancer
patients had high SHOC2 expression. Consistent with the RT-qPCR
results, increased SHOC2 expression levels, compared with those in
normal breast tissues, were detected in the tumor tissues
(P<0.01; Fig. 3B-F).
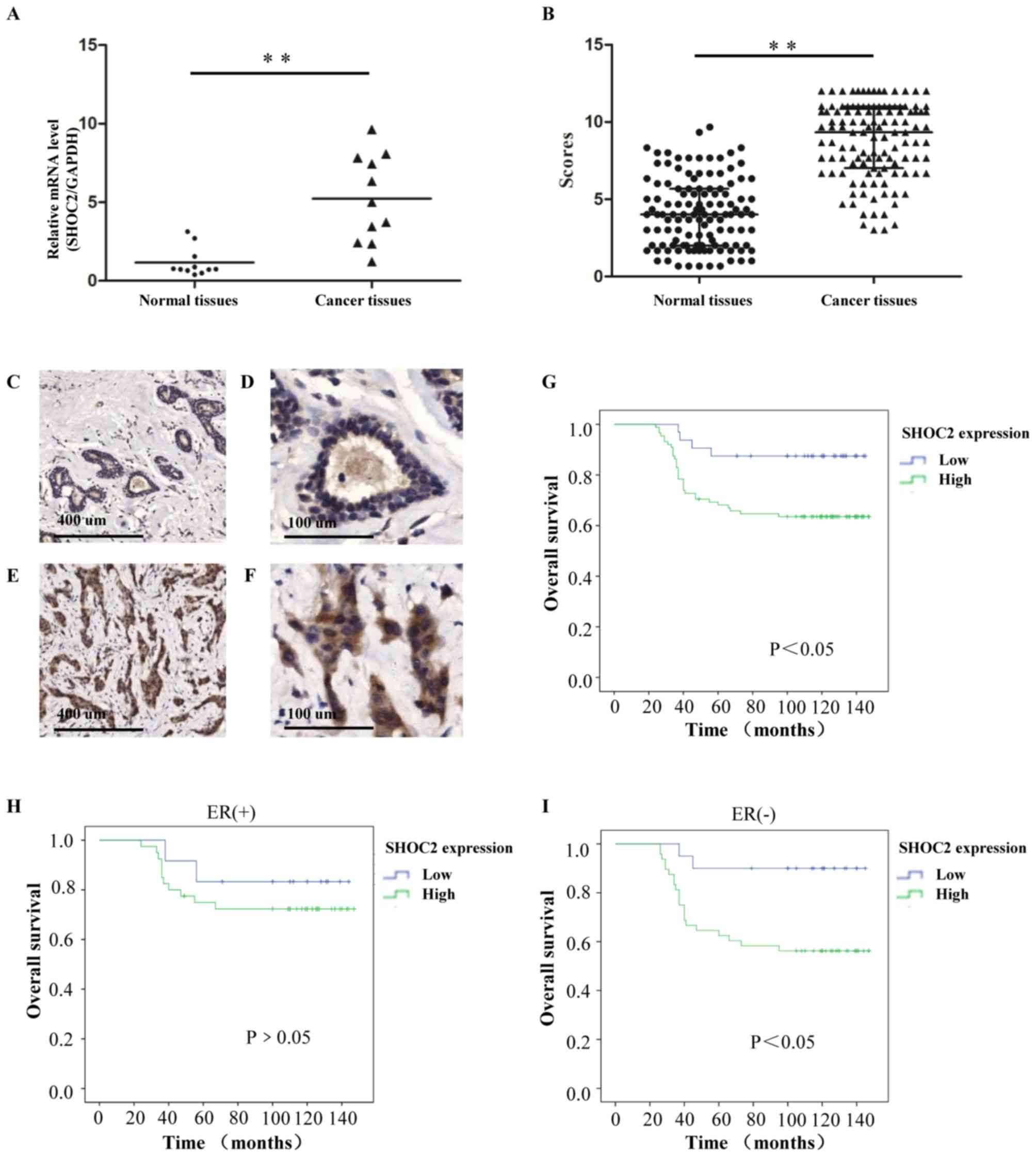 | Figure 3.Clinical association and prognostic
value of SHOC2 expression for breast cancer. (A) Relative mRNA
expression levels of SHOC2 in fresh clinical samples. (B) IHC
staining scores for the normal breast and invasive breast cancer
tissues. (C and D) IHC staining shows that SHOC2 expression was low
in normal breast tissues; (C) magnification, ×50 and (D)
magnification, ×200. (E and F) IHC staining shows that SHOC2 was
upregulated in invasive breast cancer tissues; (E) magnification,
×50 and (F) magnification, ×200. (G) Kaplan-Meier plot showing a
significant association between SHOC2 expression and OS for
patients with breast cancer. (H) In the ER (+) subgroups, OS rates
were not significantly affected by SHOC2 expression. (I) In the ER
(−) subgroups, OS rates were significantly affected by SHOC2
expression. **P<0.01. ER, estrogen receptor; IHC,
immunohistochemistry; OS, overall survival; SHOC2, SHOC2 leucine
rich repeat scaffold protein. |
Association between SHOC2 expression
and the clinicopathological features of breast cancer
The associations between SHOC2 expression and the
clinicopathological features of breast cancer are summarized in
Table I. The results showed that
significant associations were observed between high SHOC2
expression and high histopathological grades (P<0.01).
Similarly, regarding tumor size, SHOC2 expression scores were
higher in T2-T3 (>2 cm) tumors than in T1 (≤2 cm) tumors
(P=0.023). In addition, SHOC2 expression was associated with
estrogen receptor (ER) status and was higher in patients with
ER-negative breast cancer than in patients with ER-positive breast
cancer (P=0.028). However, the associations between SHOC2
expression and patient age, progesterone receptor, HER2 or lymph
node status were not statistically significant.
 | Table I.Association between SHOC2 expression
and clinicopathological features of patients with breast
cancer. |
Table I.
Association between SHOC2 expression
and clinicopathological features of patients with breast
cancer.
|
| Total no. | SHOC2 expression |
|
|---|
|
|
|
|
|
|---|
| Characteristics | 120 | Low | High | P-value |
|---|
| Age |
|
|
| 0.366 |
| ≤40
years | 35 | 7 | 28 |
|
| >40
years | 85 | 25 | 60 |
|
| Tumor size |
|
|
| 0.023 |
| ≤2
cm | 57 | 21 | 36 |
|
| 2–5
cm | 63 | 11 | 52 |
|
| Histopathological
grade |
|
|
| <0.001 |
| I | 34 | 18 | 16 |
|
| II | 53 | 9 | 44 |
|
| III | 33 | 5 | 28 |
|
| ER |
|
|
| 0.028 |
| + | 40 | 16 | 24 |
|
| − | 80 | 16 | 64 |
|
| PR |
|
|
| 0.533 |
| + | 68 | 20 | 48 |
|
| − | 52 | 12 | 40 |
|
| HER2 |
|
|
| 0.536 |
| + | 76 | 20 | 56 |
|
| − | 44 | 12 | 32 |
|
| Lymph node
status |
|
|
| 0.371 |
| + | 86 | 21 | 65 |
|
| − | 34 | 11 | 23 |
|
SHOC2 expression is an independent
prognostic factor for the survival of patients with breast
cancer
The Kaplan-Meier method was applied to explore the
significance of SHOC2 expression for the OS rates of the enrolled
patients. The results revealed that patients with low SHOC2
expression had improved OS than those with high SHOC2 expression
(Fig. 3G). The prognostic value of
SHOC2 expression was analyzed in different subgroups of patients.
For the patients with different SHOC2 expression statuses,
differences in prognosis were more notable in the ER-negative
subgroup than in the ER-positive subgroup. The OS was significantly
lower for patients with high SHOC2 expression than for patients
with low expression (P<0.05; Fig.
3H and I); this result implied that the expression of SHOC2 may
be more important for ER-negative patients. The univariate analysis
revealed that tumor size (P=0.023), ER status (P=0.015), lymph node
status (P<0.001) and SHOC2 expression (P=0.020) were all related
to OS (Table II). These data
suggested that SHOC2 might have prognostic value for patients with
breast cancer. Subsequently, a multivariate analysis was performed
using the Cox proportional hazards model and demonstrated that high
SHOC2 expression (P<0.001), lymph node status (P<0.001) and
negative ER expression status (P=0.010) were independent prognostic
factors for survival (Table
II).
| Table II.Univariate and multivariate survival
analyses of various factors in patients with breast cancer. |
Table II.
Univariate and multivariate survival
analyses of various factors in patients with breast cancer.
|
| Univariate | Multivariate |
|---|
|
|
|
|
|---|
|
Characteristics | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age |
|
|
|
|
|
|
| ≤40
years | 1 |
|
|
|
|
|
| >40
years | 0.765 | 0.387–1.510 | 0.439 |
|
|
|
| Tumor size |
|
|
|
|
|
|
| ≤2
cm | 1 |
|
| 1 |
|
|
| >2
cm | 2.280 | 1.121–4.635 | 0.023 | 1.077 | 0.507–2.288 | 0.847 |
| Histopathological
grade |
|
|
|
|
|
|
| I | 1 |
|
|
|
|
|
| II | 1.019 | 0.462–2.244 | 0.964 |
|
|
|
|
III | 1.102 | 0.726–1.674 | 0.648 |
|
|
|
| ER |
|
|
|
|
|
|
| − | 1 |
|
| 1 |
|
|
| + | 0.444 | 0.231–0.854 | 0.015 | 0.415 | 0.212–0.812 | 0.010 |
| PR |
|
|
|
|
|
|
| − | 1 |
|
|
|
|
|
| + | 0.703 | 0.356–1.387 | 0.309 |
|
|
|
| HER2 |
|
|
|
|
|
|
| − | 1 |
|
|
|
|
|
| + | 1.087 | 0.560–2.108 | 0.805 |
|
|
|
| Lymph node
status |
|
|
|
|
|
|
| − | 1 |
|
| 1 |
|
|
| + | 4.984 | 2.561–9.700 | <0.01 | 5.262 | 2.590–10.691 | <0.001 |
| SHOC2
expression |
|
|
|
|
|
|
|
Low |
|
|
| 1 |
|
|
|
High | 3.428 | 1.212–9.697 | 0.020 | 5.440 | 1.865–15.863 | <0.001 |
Discussion
It is commonly known that high frequency mutations
of Ras serve a crucial role in the development and progression of
various types of cancer, including 90% of pancreatic cancers. In
contrast, Ras mutations are rare in breast cancer (~5%) (23). Despite the low frequency of
mutations, considerable research has shown that abnormal activation
of the Ras pathway promotes breast cancer development, and Ras is
activated in numerous breast cancer cell lines, including MCF-7 and
MDA-MB-231 (24). Studies have
demonstrated that even in the presence of Ras mutations,
interfering with SHOC2 expression can still affect the
proliferation and apoptosis of cancer cells (8,25).
Therefore, as a key scaffold protein in activation of the Ras
pathway, SHOC2 may have an important role in breast cancer. In the
present study, shRNA was generated to assess the potential role of
SHOC2 in cancer cell proliferation. The results showed that SHOC2
knockdown could significantly decrease cell numbers and inhibit
breast cancer cell proliferation. Furthermore, SHOC2 knockdown
resulted in apoptosis and cell cycle arrest in breast cancer cells.
To explore the mechanism underlying the effects of SHOC2 on breast
cancer cells, the Ras pathway was examined, for which SHOC2 is
required (26). The results showed
that both the PI3K/AKT and MAPK/ERK pathways were inhibited by
SHOC2 knockdown, which supported previous research into colorectal
carcinoma (12).
In the present study, clinical data were also
analyzed, the results revealed that SHOC2 protein was frequently
expressed in breast tumors and associated with clinicopathological
variables important for disease outcome. Additionally, to the best
of our knowledge, this study was the first to reveal that the
difference in SHOC2 expression between breast cancer and normal
breast tissue was significant (P<0.01), suggesting that SHOC2
may play an important role in breast cancer progression. To assess
the clinical significance of SHOC2 protein expression in breast
cancer, the association between a number of clinicopathological
characteristics and SHOC2 protein expression status was assessed in
the breast tumor samples. It was observed that SHOC2 expression was
more abundant in T2-T3 tumors than in T1 tumors (P=0.023).
Similarly, patients with higher SHOC2 expression had higher
histological grade tumors than patients with low SHOC2 protein
expression (P<0.01). Therefore, SHOC2 upregulation is associated
with worse clinical features.
The effect of SHOC2 expression on patient survival
was estimated by Kaplan-Meier analysis. Patients with breast cancer
and high SHOC2 protein expression had a significantly worse
prognosis than patients with low SHOC2 protein expression
(P<0.05). Notably, in the subgroup analysis, it was revealed
that patients in the ER-negative subgroup with low SHOC2 expression
had an improved prognosis than those with high SHOC2 expression.
Therefore, SHOC2 may serve a more important role in patients with
ER-negative breast cancer than in patients with ER-positive breast
cancer (P<0.05). In addition, the results demonstrated that high
SHOC2 expression, large tumor size, ER-negative status and positive
lymph node status were independent prognostic factors for this
disease. These data suggested that SHOC2 upregulation was related
to poor prognosis and could act as a therapeutic target in breast
cancer. As shown in this study, the clinical effects of SHOC2 on
patients with ER-negative breast cancer were more significant.
Previous studies have demonstrated that the Ras pathway, in which
SHOC2 has a major role, is more active in ER-negative breast tumors
(27,28).
However, there were several limitations of the
present study. Firstly, only one shRNA target sequence was used to
knockdown SHOC2; it would have been useful to use more. Although,
to avoid the off-target effects, two cell lines were used to verify
its reliability. Secondly, even though the present study included
120 patients, they were recruited from a single center so this
study suffered from selection bias. Additionally, the rate of
disease-free survival (DFS; the time to recurrence or distant
metastasis) of the patients was recorded during initial follow-ups,
but these patients all passed away in further follow-ups. This
meant that the effect of SHOC2 expression on DFS and OS are similar
in this research so only the OS results were shown, which is
another limitation to the present study. More clinical and
experimental studies are required to define the genetic mechanisms
in order to further understand the role of SHOC2 in normal mammary
gland and breast cancer tissues.
In conclusion, it was found that SHOC2 knockdown has
a significant effect on breast cancer cell proliferation, as well
as the induction of cell cycle arrest and apoptosis. Additionally,
SHOC2 was overexpressed in breast cancer and was associated with
the OS of patients with breast cancer. The biological function of
SHOC2 was examined in this study, and these results suggested that
SHOC2 could be a therapeutic target for breast cancer.
Acknowledgements
The authors would like to thank Dr Qianqian Zhao and
Dr Haiyun Song of the Pathology Department, Qilu Hospital (Qingdao)
of Shandong University for their help in the IHC experiment.
Funding
This study was funded by The NSFC (grant no.
81572587) and The Shandong Natural Science Fund (grant no.
ZR2018PH029).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HG designed the experiments. WG, KD and YL performed
the experiments. WG and QP drafted the manuscript, and QP performed
statistical analysis and figure illustration. YL provided
experimental technical guidance. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
This study has been approved by the Research Ethics
Committee of Qilu Hospital (Qingdao) of Shandong University (policy
no. KYLL-2016038). All procedures performed in the studies
involving human participants were in accordance with the ethical
standards of the Research Ethics Committee of Qilu Hospital
(Qingdao) of Shandong University and with The Declaration of
Helsinki (1964) and its later amendments or comparable ethical
standards. Informed consent was obtained from all individual
participants included in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ban KA and Godellas CV: Epidemiology of
breast cancer. Surg Oncol Clin N Am. 23:409–422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee EY and Muller WJ: Oncogenes and tumor
suppressor genes. Cold Spring Harb Perspect Biol. 2:a0032362010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Downward J: Targeting RAS signalling
pathways in cancer therapy. Nat Rev Cancer. 3:11–22. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sieburth DS, Sun Q and Han M: SUR-8, a
conserved Ras-binding protein with leucine-rich repeats, positively
regulates Ras-mediated signaling in C. elegans. Cell. 94:119–130.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jang ER, Jang H, Shi P, Popa G, Jeoung M
and Galperin E: Spatial control of Shoc2-scaffold-mediated ERK1/2
signaling requires remodeling activity of the ATPase PSMC5. J Cell
Sci. 128:4428–4441. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matsunaga-Udagawa R, Fujita Y, Yoshiki S,
Terai K, Kamioka Y, Kiyokawa E, Yugi K, Aoki K and Matsuda M: The
scaffold protein Shoc2/SUR-8 accelerates the interaction of Ras and
Raf. J Biol Chem. 285:7818–7826. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li W, Han M and Guan KL: The leucine-rich
repeat protein SUR-8 enhances MAP kinase activation and forms a
complex with Ras and Raf. Genes Dev. 14:895–900. 2000.PubMed/NCBI
|
|
8
|
Rodriguez-Viciana P, Oses-Prieto J,
Burlingame A, Fried M and McCormick F: A phosphatase holoenzyme
comprised of Shoc2/Sur8 and the catalytic subunit of PP1 functions
as an M-Ras effector to modulate Raf activity. Mol Cell.
22:217–230. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shaw AS and Filbert EL: Scaffold proteins
and immune-cell signalling. Nat Rev Immunol. 9:47–56. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rouchka EC, Jeoung M, Jang ER, Liu J, Wang
C, Li X and Galperin E: Data set for transcriptional response to
depletion of the Shoc2 scaffolding protein. Data Brief. 7:770–778.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Young LC, Hartig N, Munoz-Alegre M,
Oses-Prieto JA, Durdu S, Bender S, Vijayakumar V, Vietri Rudan M,
Gewinner C, Henderson S, et al: An MRAS, SHOC2, and SCRIB complex
coordinates ERK pathway activation with polarity and tumorigenic
growth. Mol Cell. 52:679–692. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee YM, Kaduwal S, Lee KH, Park JC, Jeong
WJ and Choi KY: Sur8 mediates tumorigenesis and metastasis in
colorectal cancer. Exp Mol Med. 48:e2492016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kaduwal S, Jeong WJ, Park JC, Lee KH, Lee
YM, Jeon SH, Lim YB, Min do S and Choi KY: Sur8/Shoc2 promotes cell
motility and metastasis through activation of Ras-PI3K signaling.
Oncotarget. 6:33091–33105. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kaplan FM, Kugel CH III, Dadpey N, Shao Y,
Abel EV and Aplin AE: SHOC2 and CRAF mediate ERK1/2 reactivation in
mutant NRAS-mediated resistance to RAF inhibitor. J Biol Chem.
287:41797–41807. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Traini S, Piccolo E, Tinari N, Rossi C, La
Sorda R, Spinella F, Bagnato A, Lattanzio R, D'Egidio M, Di Risio
A, et al: Inhibition of tumor growth and angiogenesis by SP-2, an
anti-lectin, galactoside-binding soluble 3 binding protein
(LGALS3BP) antibody. Mol Cancer Ther. 13:916–925. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jeoung M, Jang ER, Liu J, Wang C, Rouchka
EC, Li X and Galperin E: Shoc2-tranduced ERK1/2 motility
signals--Novel insights from functional genomics. Cell Signal.
28:448–459. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Aran D, Camarda R, Odegaard J, Paik H,
Oskotsky B, Krings G, Goga A, Sirota M and Butte AJ: Comprehensive
analysis of normal adjacent to tumor transcriptomes. Nat Commun.
8:10772017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li C, Cao L, Xu C, Liu F, Xiang G, Liu X,
Jiao J and Niu Y: The immunohistochemical expression and potential
prognostic value of HDAC6 and AR in invasive breast cancer. Hum
Pathol. 75:16–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ye W, Chen C, Gao Y, Zheng ZS, Xu Y, Yun
M, Weng HW, Xie D, Ye S and Zhang JX: Overexpression of SLC34A2 is
an independent prognostic indicator in bladder cancer and its
depletion suppresses tumor growth via decreasing c-Myc expression
and transcriptional activity. Cell Death Dis. 8:e25812017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen Q, Yao Q, Sun J, Feng L, Lu H, Ma Y,
Liu L, Wang F, Li J, Yue Y, et al: Downregulation of histone
deacetylase 1 by microRNA-520h contributes to the chemotherapeutic
effect of doxorubicin. FEBS Lett. 588:184–191. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin X, Yu Y, Zhao H, Zhang Y, Manela J and
Tonetti DA: Overexpression of PKCalpha is required to impart
estradiol inhibition and tamoxifen-resistance in a T47D human
breast cancer tumor model. Carcinogenesis. 27:1538–1546. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Clark GJ and Der CJ: Aberrant function of
the Ras signal transduction pathway in human breast cancer. Breast
Cancer Res Treat. 35:133–144. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Eckert LB, Repasky GA, Ulku AS, McFall A,
Zhou H, Sartor CI and Der CJ: Involvement of Ras activation in
human breast cancer cell signaling, invasion, and anoikis. Cancer
Res. 64:4585–4592. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jones GG, Del Rio IB, Sari S, Sekerim A,
Young LC, Hartig N, Areso Zubiaur I, El-Bahrawy MA, Hynds RE, Lei
W, et al: SHOC2 phosphatase-dependent RAF dimerization mediates
resistance to MEK inhibition in RAS-mutant cancers. Nat Commun.
10:25322019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jang ER and Galperin E: The function of
Shoc2: A scaffold and beyond. Commun Integr Biol. 9:e11882412016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Loboda A, Nebozhyn M, Klinghoffer R,
Frazier J, Chastain M, Arthur W, Roberts B, Zhang T, Chenard M,
Haines B, et al: A gene expression signature of RAS pathway
dependence predicts response to PI3K and RAS pathway inhibitors and
expands the population of RAS pathway activated tumors. BMC Med
Genomics. 3:262010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li W, Liang RR, Zhou C, Wu MY, Lian L,
Yuan GF, Wang MY, Xie X, Shou LM, Gong FR, et al: The association
between expressions of Ras and CD68 in the angiogenesis of breast
cancers. Cancer Cell Int. 15:172015. View Article : Google Scholar : PubMed/NCBI
|