Introduction
Renal interstitial fibrosis, characterized by the
activation of interstitial fibroblasts, accumulation of excess
extracellular matrix components and tubular atrophy, is a common
pathology associated with the progression of chronic kidney disease
(CKD) to end-stage renal disease caused by various etiologies
(1–3). The aforementioned interstitial and
tubular alterations in the progression of CKD may lead to
irreversible renal damage and impaired renal function, eventually
leading to end-stage renal failure (4,5).
Epidemiological studies have demonstrated that the prevalence of
CKD is increasing annually worldwide with a high incidence and
mortality rate, and has become a major global public health concern
(6–8). Therefore, understanding the
pathogenesis of the molecular mechanisms underlying
tubulointerstitial fibrosis is of great importance to identify
novel targets for the effective treatment, prevention and delay of
CKD.
Epithelial-mesenchymal transition (EMT) has been
reported as the major event characterizing the pathogenesis of
renal interstitial fibrosis (9).
In the process of EMT, renal tubular epithelial cells lose their
unique phenotype and acquire the mesenchymal cell phenotype to
differentiate into mesenchymal fibroblasts (10). Increasing evidence has indicated
that transforming growth factor- β1 (TGF-β1) is the key factor in
the regulation of renal interstitial fibrosis (11–13),
and is the most potent inducer in initiating and completing the
entire EMT course.
Bone morphogenetic protein-7 (BMP-7) is a
homodimeric protein that belongs to the TGF-β superfamily, and has
a wide range of biological activities including the regulation of
cell proliferation, and osteogenic and anti-inflammatory activity
(14,15). Previous studies have revealed that
BMP-7-knockout mice succumbed to mortality shortly after birth due
to diffuse renal dysplasia, revealing that BMP-7 is indispensable
for normal kidney development (16,17).
It has also been reported that the expression of BMP-7 in the
kidney is downregulated and gradually becomes more serious with
disease progression in the pathogenesis of ischemia-reperfusion
injury (18), diabetic nephropathy
(19), hypertensive
nephrosclerosis (20) and
unilateral ureteral obstruction (UUO) (21). In addition, numerous studies have
demonstrated that exogenous BMP-7 could inhibit or reverse
TGF-β1-induced EMT in vitro, prevent or delay the
development of fibrosis, and improve renal function in a variety of
kidney disease models (22–24);
however, the exact molecular mechanism of BMP-7 in improving renal
interstitial fibrosis has not been fully elucidated.
Uterine sensitization-associated gene-1 (USAG-1) is
a specific antagonist of BMP-7 and is mainly expressed in distal
convoluted renal tubular epithelial cells (25). In adult kidneys, it has been
reported that USAG-1 is the major negative regulator of BMP
function, and binds to BMP-7, thereby inhibiting the interaction
between BMP-7 and its receptor, ultimately weakening the renal
protective effects of BMP-7 (26).
Mice with USAG-1 knockout could promote the expansion of
mesenchymal stem cells and accelerate fracture healing, and inhibit
the development of renal interstitial fibrosis (27,28).
Tanaka et al (29) revealed
that USAG-1 gene-deficient mice with Alport syndrome had mild renal
disease and delayed disease progression. It has been reported that
USAG-1 gene defects were more capable of promoting acute kidney
injury caused by cisplatin and chronic kidney injury compared with
in wild-type mice, while renal protection was suppressed when BMP-7
neutralizing antibodies were administered. Thus, USAG-1 may affect
the protective properties of endogenous BMP-7 in the kidney
(25,30); however, whether USAG-1 affects the
process of renal interstitial fibrosis by altering the occurrence
of EMT remains unclear.
Febuxostat, a novel non-selective xanthine oxidase
inhibitor, potently reduces uric acid synthesis in the body,
decreasing the concentration (31). Febuxostat is mainly used for the
treatment of gout in clinical practice (32,33).
A recent study demonstrated that febuxostat can also reduce serum
uric acid levels, delay the progression of renal dysfunction in
patients with chronic kidney disease and reduce the risk of
cardiovascular disease, in addition to the treatment of gout
(34). It was reported that
febuxostat exhibited renal protective effects on cisplatin-induced
acute and chronic kidney injury, and streptozotocin-induced
diabetic rats in the experimental animal model, and thus could
reduce renal tissue damage (35,36).
Our previous study also revealed that febuxostat could ameliorate
renal interstitial fibrosis caused by UUO (37).
Based on previous studies, the present study aimed
to investigate the following: i) The role of USAG-1 in
TGF-β1-induced EMT in Madin-Darby canine kidney (MDCK) cells; and
ii) whether febuxostat could exert its inhibitory effect on EMT,
and if so, whether this inhibitory effect was associated with the
downregulation of USAG-1 expression, thereby activating the
BMP-7/Smad1/5/8 signaling pathway.
Materials and methods
Cell lines
MDCK cells from Otwo Biotech (Shenzhen) Inc.,
Guangzhou, China were cultured in RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), supplemented with 10%
fetal bovine serum (FBS; Hangzhou Sijiqing Bioengineering Material
Co., Ltd. Hangzhou, China) and cultured in a 5% CO2
incubator at 37°C until 60–70% confluence was attained. Cells were
treated with 10 ng/ml human recombinant TGF-β1 (PeproTech, Inc.,
Rocky Hill, NJ, USA) for 48 h at 37°C to induce EMT. For
experimental use, MDCK cells were treated with RPMI-1640 medium at
37°C, containing 1% FBS (Hangzhou Sijiqing Bioengineering Material
Co., Ltd.) for 12 h, and then treated with 10 ng/ml TGF-β1 combined
with or without low (0.1 µM), middle (1 µM) or high (10 µM)
concentrations of febuxostat for 48 h at 37°C. All experiments were
performed three times. Following treatment for 48 h, the cells were
harvested for detection.
MTT assay for cell viability
analysis
MDCK cells were seeded into 96-well culture plates
and incubated with 1% FBS for 12 h when the cells were attached,
followed by treatment with different concentrations of febuxostat
for 48 h. Cell viability was determined using an MTT assay.
Subsequently, cells were incubated with MTT solution (0.5 mg/ml)
for 4 h at 37°C. The purple formazan crystals derived from the
addition of MTT were dissolved in dimethyl sulfoxide and agitated
for 10 min. The absorbance at 490 nm was measured using a
microplate reader (BioTek Instruments, Inc., Winooski, VT,
USA).
RNA interference
The sequences of small interfering (si)RNA-control
(si-ctrl) and siRNA-USAG-1 interference plasmids flanked by EcoRI
and AgeI restriction site were constructed into the H1 promoter of
lentivirus infectious virions pGLV3-H1-GFP-Puro (Public
Protein/Plasmid Library Biotechnology Co., Ltd. Nanjing, China).
The negative control siRNA was designed as: Forward,
5′-TTCTCCGAACGTGTCACGTAA-3′; the siRNA-USAG-1 was designed as:
Forward, 5′-CCTCCTGCCATTCATTTCTT-3′. Cells (1.0×106/ml)
were plated into 6-mm wide petri dish and cultured in RPMI-1640
medium. A total of 5 µg siRNA-expressing plasmids and 10 µl
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
were used for transfection when cell confluency reached 70–80%.
MDCK cells were divided into the blank group, si-ctrl, TGF-β1 (10
ng/ml) stimulation (TGF-β1), TGF-β1 (10 ng/ml) plus siRNA-control
(TGF-β1 + si-ctrl), and TGF-β1 (10 ng/ml) plus siRNA-USAG-1 group
(TGF-β1 + si-USAG-1). Following transfection for 6 h, the medium
was removed and cells were cultured in serum-free medium containing
10% FBS without antibiotics. Following transfection for 22 h, the
cells were treated with 10 ng/ml TGF-β1, containing 1% FBS for 48 h
at 37°C, and after 70 h of transfection, the cells were collected
for experimentation.
RNA isolation and reverse
transcription (RT-) semi-quantitative polymerase chain reaction
(sqPCR) analysis
Total RNA was extracted from MDCK cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to manufacturer's protocols. RT of total RNA was
performed using the TIANScript RT kit (Tiangen Biotech Co., Ltd.,
Beijing, China) with the following temperature protocol: 42°C for
50 min, 95°C for 5 min, then held at 4°C. The primers of α-smooth
muscle actin (α-SMA) and E-cadherin were synthesized by Sangon
Biotech Co., Ltd. (Shanghai, China). The primer sequences were as
follows: α-SMA forward, 5′-GTGATGGTGGGGATGGGACAA-3′ and reverse,
5′-CCAGAGGCGTAGAGGGAAAGC-3′ (311 bp); E-cadherin forward,
5′-CAGCATCCTCACACAAGACC-3′ and reverse, 5′-TCAGCATCCGTCACTTTGAG-3′
(300 bp); GAPDH forward, 5′-CTTGAAAGGCGGGGCCAAGAGG-3′ and reverse,
5′-ACTGATACATTGGGGGTGGGGACA-3′ (393 bp). sqPCR was performed using
the 2XTaq PCR MasterMix (Tiangen Biotech Co., Ltd. Beijing, China)
with initial denaturation at 94°C for 5 min, followed by 30
consecutive cycles of denaturation at 94°C for 30 sec, annealing at
58–62°C for 30 sec, extension at 72°C for 1 min, and then a final
extension at 72°C for 7 min using a thermal cycler system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The amplified products
were analyzed by electrophoresis on a 1.5% (w/v) agarose gel which
was stained with fluorescence staining dye Goldview (Beijing
Solarbio Science and Technology Co., Ltd. Beijing, China) at 60°C.
The gel was congealed at room temperature for 30 min. The
electrophoresis continued for 50 min at 110 V. The relative
abundance of mRNAs was measured using GAPDH mRNA expression levels
as the internal reference. The signal intensity of the images was
analyzed using ImageJ version 1.48 software (National Institutes of
Health, Bethesda, MD, USA).
Western blot analysis
Proteins were extracted from MDCK cells using
radioimmunoprecipitation assay lysis buffer containing RIPA and the
protease inhibitor PMSF (both Beyotime Institute of Biotechnology,
Shanghai, China). The compounds were placed at 4°C for at least 60
min and then centrifuged at 4°C and 10,000 × g for 15 min. The
supernatant was collected for western blotting. Protein
concentration in the supernatant was determined using the BCA
protein assay kit (Beyotime Institute of Biotechnology). Equal
amounts of protein (60 µg) were separated using SDS-PAGE (8–10%)
and transferred onto nitrocellulose membranes. The membranes were
then blocked in PBS containing 3% bovine serum albumin (BSA; Vicmed
Biotech Co. Ltd., Xuzhou, China) and incubated with primary
antibodies, including anti-E-cadherin (1:1,000; cat. no. ab1416),
anti-α-SMA (1:1,000; cat. no. ab7817), anti-USAG-1 (1:1,000; cat.
no. ab99340; all Abcam, Cambridge, UK), anti-p-Smad1/5/8 (1:1,000;
cat. no. 13820; Cell Signaling Technology, Inc., Danvers, MA, USA)
and anti-Smad1/5/8 (1:1,000; cat. no. sc-6031-R; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) overnight at 4°C.
Near-infrared fluorescence-conjugated goat anti-rabbit (1:1,000;
cat. no. V926-32211) or anti-mouse (1:1,000; cat. no. V926-32210;
both Vicmed Biotech Co. Ltd.) secondary antibodies were used to
incubate the membranes for 1 h at room temperature, which were
developed colorimetrically via the Odyssey biocolor infrared
fluorescence imaging system (LI-COR Biosciences, Lincoln, NE, USA).
Quantification was performed by measuring the intensity of the
signals using ImageJ software version 1.48 by quantifying the
relative expression of target protein vs. GAPDH.
Immunofluorescence staining
analysis
MDCK cells (1.0×104/ml) were seeded into
12-well plates with pre-placed slides. Following the adherence of
cells, the cells were allowed to stand for 12 h with 1% FBS, prior
to being washed with PBS twice and treated with TGF-β1 with or
without different concentrations of febuxostat (0.1, 1 or 10 µM)
for 48 h. After 48 h, the cells were washed with PBS for three
times for 5 min per wash and fixed at room temperature for 30 min
with 4% paraformaldehyde; 0.5% TritonX-100 was added to
permeabilize cells for 10 min. Cells were subsequently washed with
PBS three times for 5 min each time and covered with 2% BSA (Vicmed
Biotech Co. Ltd.) for 30 min at 37°C. Next, cells were incubated
with USAG-1 (1:200; cat. no. ab99340), α-SMA (1:200; cat. no.
ab7817; both Abcam) or E-cadherin (1:200; cat. no. 14472; Cell
Signaling Technology, Inc.) antibodies, which had been diluted in
PBS overnight at 4°C. Subsequently, the cells were incubated with
goat anti-rabbit (1:500; cat. no. A-11034; Alexa Fluor 488) or
anti-mouse (1:500; cat. no. A-11004; Alexa Fluor 568; both Thermo
Fisher Scientific, Inc.) secondary antibodies for 2 h at 37°C and
were then stained with the nuclear-specific stain DAPI (Beyotime
Institute of Biotechnology) for 3 min at room temperature.
Following three washes with PBS for 5 min per wash,
anti-fluorescence quenching liquid (Beyotime Institute of
Biotechnology) was added to the clean glass slides and cells were
analyzed under an Olympus BX43F fluorescence microscope (Olympus
Corporation, Tokyo, Japan). The excitation light of goat
anti-rabbit secondary antibody is 488 nm, and the excitation light
of goat anti-mouse secondary antibody is 568 nm. Digital images
were captured using an inverted fluorescent microscope
(magnification, ×400).
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Statistical analysis was performed using SPSS 16.0
statistical software (SPSS, Inc., Chicago, IL, USA). Comparison
between groups was conducted using one-way analysis of variance,
followed by the Least Significant Difference test with homogeneity
of variances, or the Dunnett's T3 test with heterogeneity of
variances. P<0.05 was considered to indicate a statistically
significant difference. All experiments were performed three
times.
Results
TGF-β1 upregulates the expression of
USAG-1 and induces EMT in MDCK cells, whereas USAG-1 gene silencing
prevents TGF-β1-induced EMT
According to the western blotting results,
transfection with siRNA-USAG-1 significantly decreased the protein
expression of USAG-1 compared with the siRNA-control group
(Fig. 1A). Following treatment
with TGF-β1, the protein expression levels of USAG-1 (Fig. 1B) and α-SMA (Fig. 1C) were significantly increased,
while that of E-cadherin (Fig. 1D)
was significantly decreased in MDCK cells compared with the blank
group (P<0.01). Additionally, it was also demonstrated that
transfection with USAG-1-specific siRNA for 70 h and TGF-β1
treatment significantly decreased the protein expression of α-SMA
(Fig. 1C), while that of
E-cadherin was increased compared with the TGF-β1 + siRNA-control
group (P<0.01; Fig. 1D).
Immunofluorescence revealed that the findings were consistent with
those of western blotting. MDCK cells treatment with TGF-β1
exhibited an increased intensity of anti-α-SMA labeling (Fig. 1E, red staining) and a decreased
intensity of anti-E-cadherin labeling (Fig. 1F, red staining); however,
siRNA-USAG-1 reversed the these alterations induced by TGF-β1
(Fig. 1E and F). Notable
differences were observed in protein expression between the blank
and the siRNA-control groups, or the TGF-β1 and TGF-β1 +
siRNA-control groups. Collectively, these findings indicated that
USAG-1 may promote the occurrence of EMT induced by TGF-β1 in MDCK
cells.
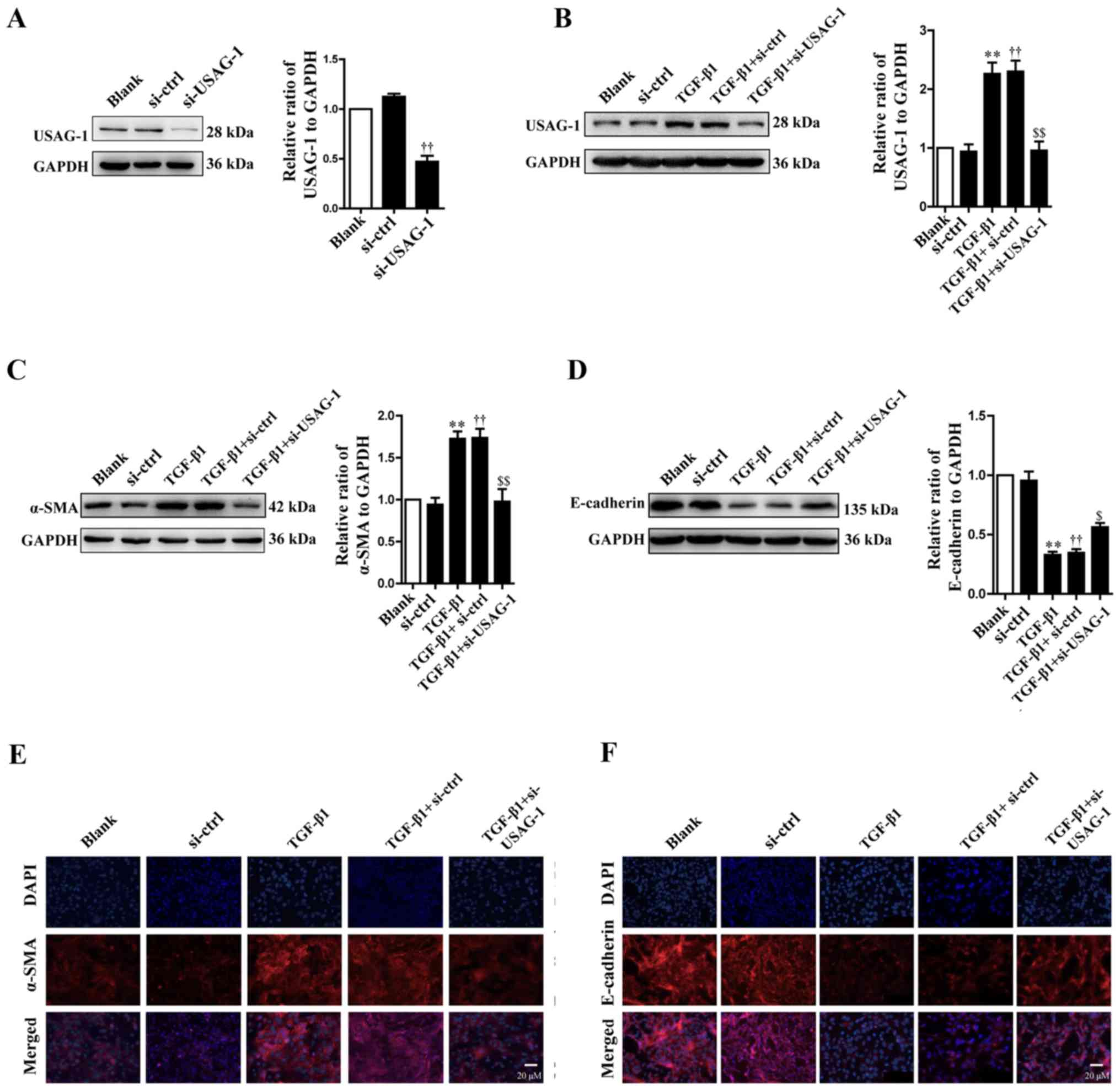 | Figure 1.Effects of USAG-1 on
epithelial-mesenchymal transition induced by TGF-β1 in MDCK cells.
MDCK cells were transfected with a negative control siRNA or a
siRNA-USAG-1 plasmid for 70 h. The protein expression levels of (A
and B) USAG-1, (C) α-SMA and (D) E-cadherin were evaluated using
western blotting. All the data are presented as the mean ± standard
error of the mean, n=3. **P<0.01 vs. blank group;
††P<0.01 vs. siRNA-ctrl; $P<0.05 and
$$P<0.01 vs. TGF-β1 + siRNA-ctrl group.
Immunofluorescence staining demonstrated the labeling intensity of
(E) α-SMA (red staining) and (F) E-cadherin (red staining).
Magnification, ×400. α-SMA, α-smooth muscle actin; MDCK,
Madin-Darby canine kidney; si, small interfering RNA; si-ctrl,
siRNA-control; si-USAG-1, siRNA-USAG-1; TGF-β1, transforming growth
factor-β1; USAG-1, uterine sensitization-associated gene-1. |
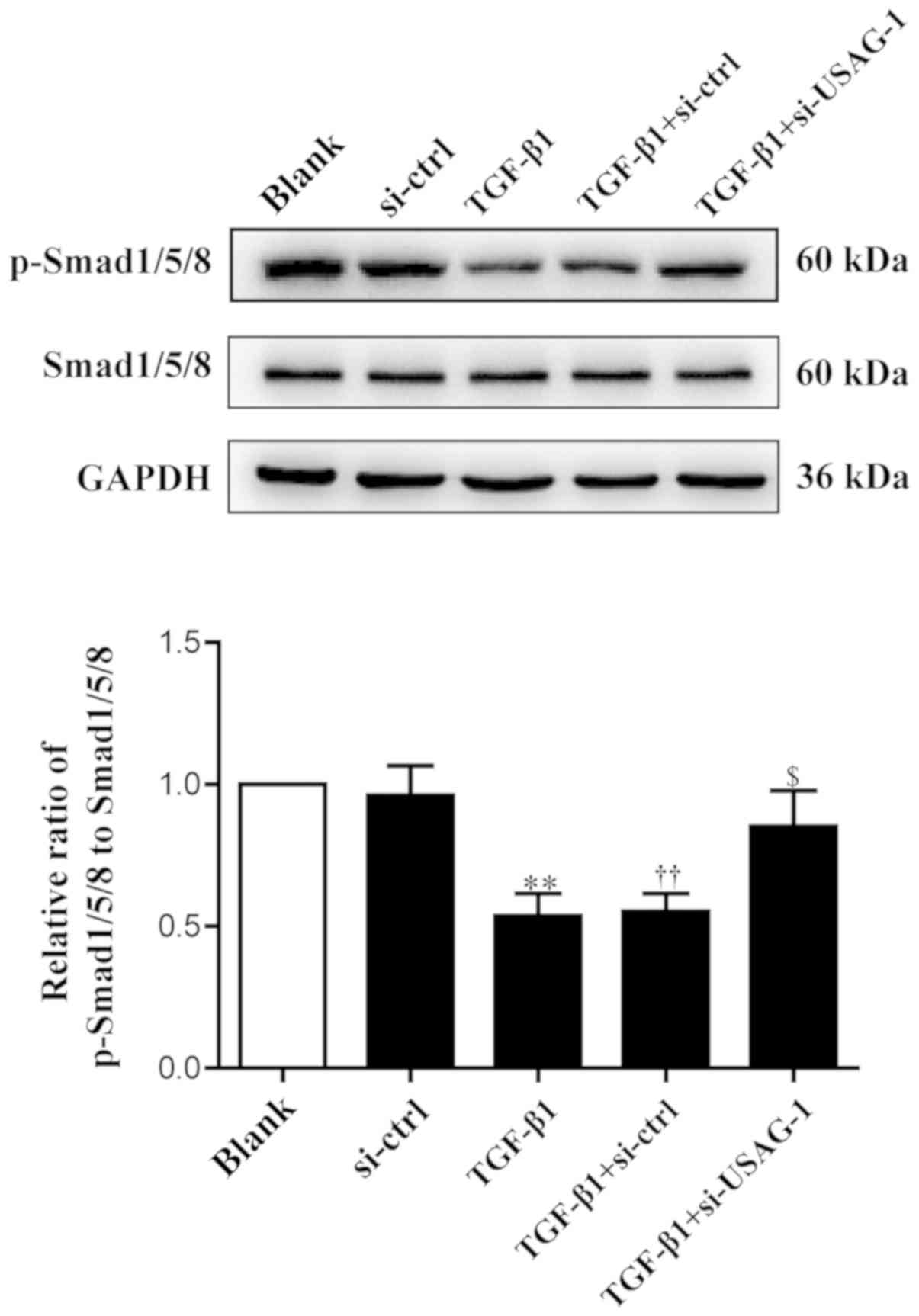
USAG-1 inhibits the activity of the
Smad1/5/8 signaling pathway in TGF-β1-induced EMT
USAG-1 is a specific antagonist of BMP-7, and
Smad1/5/8 are the key intracellular signaling proteins of BMP-7;
the phosphorylation of Smad1/5/8 is a central downstream element of
BMP signal transduction (30,38).
Therefore, the present study examined the levels of phosphorylated
and total Smad1/5/8 by western blot analysis. Compared with the
blank group, the phosphorylation levels of Smad1/5/8 were
significantly decreased in MDCK cells following treatment with
TGF-β1; the phosphorylation levels of Smad1/5/8 were significantly
upregulated compared with the TGF-β1 + siRNA-control group, almost
to basal levels of the blank group following si-USAG-1 transfection
and TGF-β1 treatment (Fig. 2). No
significant difference was observed in the protein expression
levels between the blank and the siRNA-control groups, or the
TGF-β1 and TGF-β1 + siRNA-control groups. These results indicated
that USAG-1 may promote the occurrence of EMT by, at least partly,
inhibiting the activity of the Smad1/5/8 signaling pathway.
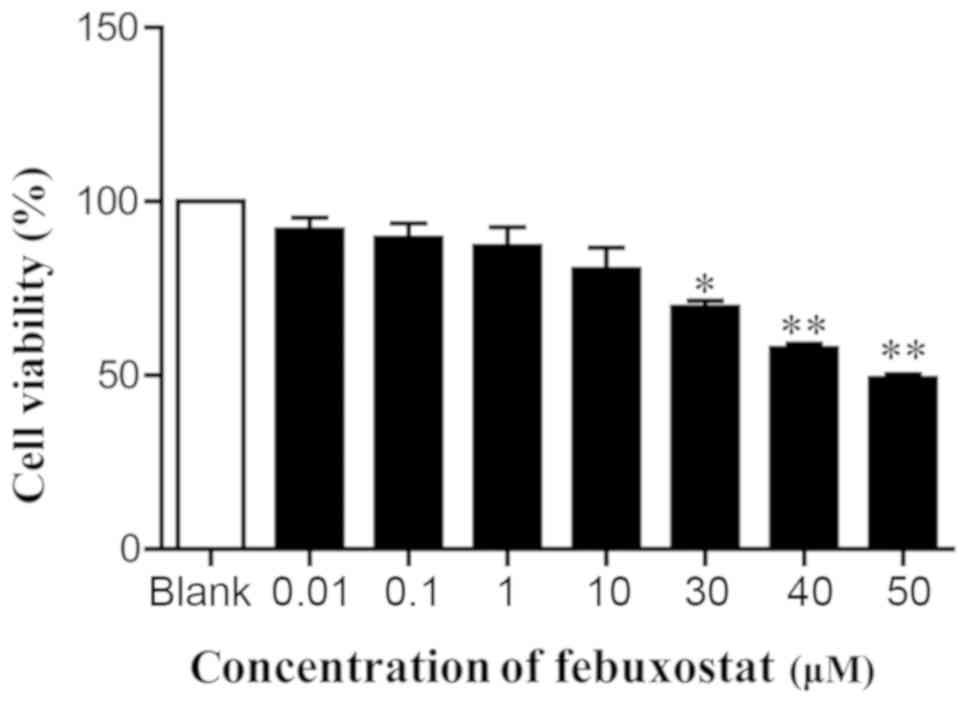
Effects of febuxostat on cell
viability
In order to examine the effects of febuxostat on the
viability of MDCK cells, cells were treated with various
concentrations of febuxostat for 48 h. The results revealed that
concentrations of febuxostat at 30, 40 and 50 µM significantly
reduced cell viability compared with the blank group. Conversely,
there was no or little effects on cell viability at doses of 0.01,
0.1, 1 and 10 µM febuxostat compared with the blank group (Fig. 3). Considering the effects of
febuxostat on cell viability and the results of MTT assay,
concentrations of 0.1, 1 and 10 µM febuxostat were selected for
subsequent experiments.
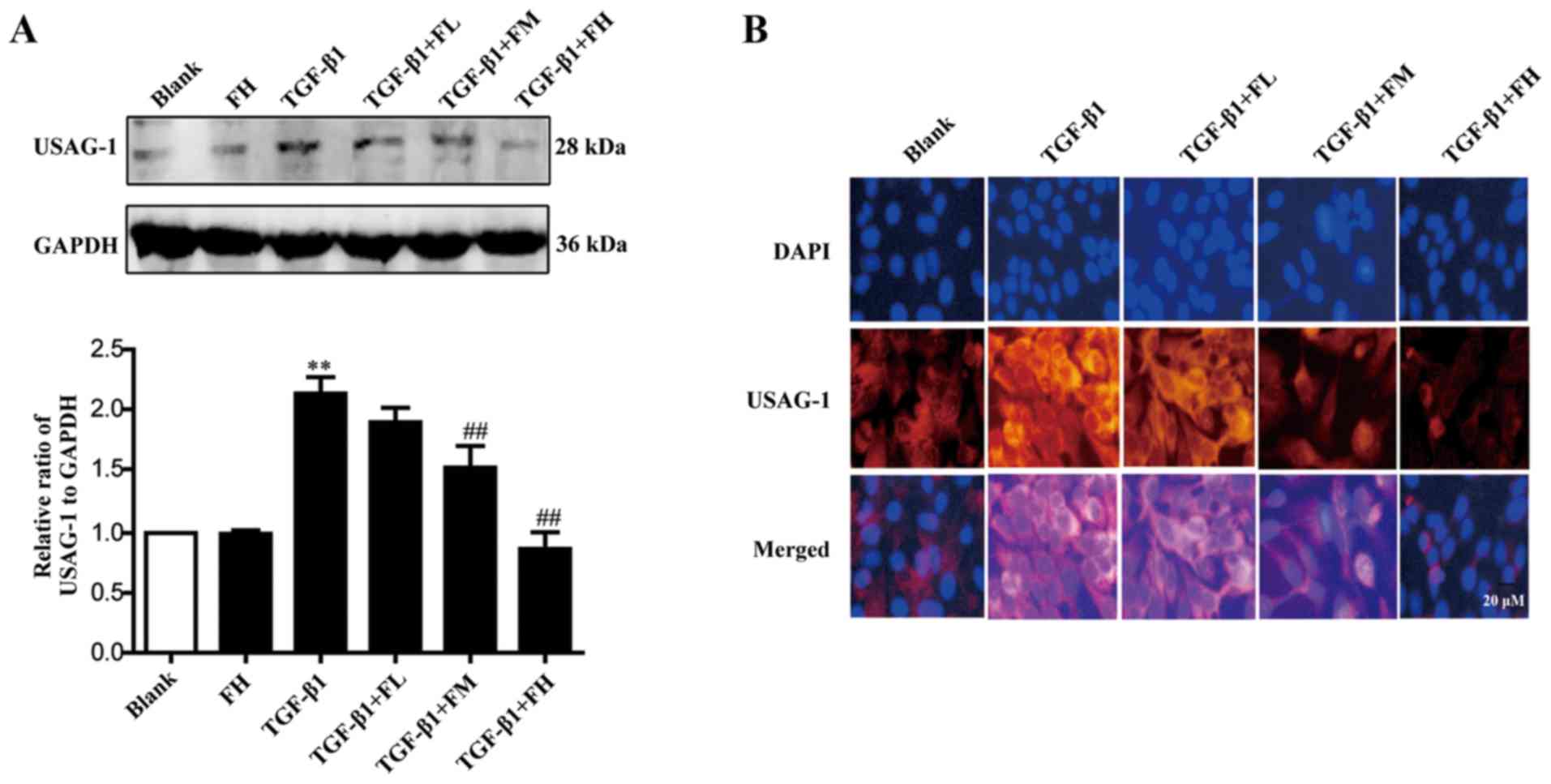
Febuxostat inhibits the expression of
USAG-1 in MDCK cells
The expression of USAG-1 in MDCK cells was detected
by western blotting and immunofluorescence staining. Immunoblotting
demonstrated that middle and high concentrations of febuxostat
significantly decreased the expression of USAG-1, while a low
concentration had no significant effect on expression compared with
the TGF-β1 group (Fig. 4A).
Immunofluorescence staining further revealed that the labeling
intensity of USAG-1 (red staining) notably increased following
TGF-β1 treatment; however, low, middle and high concentrations of
febuxostat weakened the increased labeling intensity of USAG-1
induced by TGF-β1, particularly when middle and high concentrations
of febuxostat were applied (Fig.
4B).
Febuxostat inhibits the EMT of MDCK
cells induced by TGF-β1
EMT is an important mechanism that induces renal
interstitial fibrosis, and it is essential for the development of
chronic kidney disease (39). To
investigate the role of febuxostat in TGF-β1-induced EMT, the
expression levels of the epithelial marker, E-cadherin, and the
mesenchymal marker, α-SMA, were determined by semi-qPCR and western
blotting. MDCK cells exposed to TGF-β1 for up to 48 h exhibited a
significant increase in α-SMA at the mRNA (Fig. 5A) and protein (Fig. 5B) expression levels (P<0.01),
while the expression levels of E-cadherin at the mRNA (Fig. 5C) and protein (Fig. 5D) expression levels were
significantly decreased, compared with in the blank group
(P<0.01). Conversely, middle and high concentrations of
febuxostat significantly reversed the downregulation of E-cadherin
mRNA and protein expression (Fig. 5C
and D) stimulated by TGF-β1; a low concentration of febuxostat
significantly affected the mRNA expression of E-cadherin compared
with TGF-β1 treatment alone (Fig.
5C). In addition, there were no notable effects on the
expression of α-SMA at the mRNA and protein expression levels
following treatment with a high concentration of febuxostat in
normal cells.
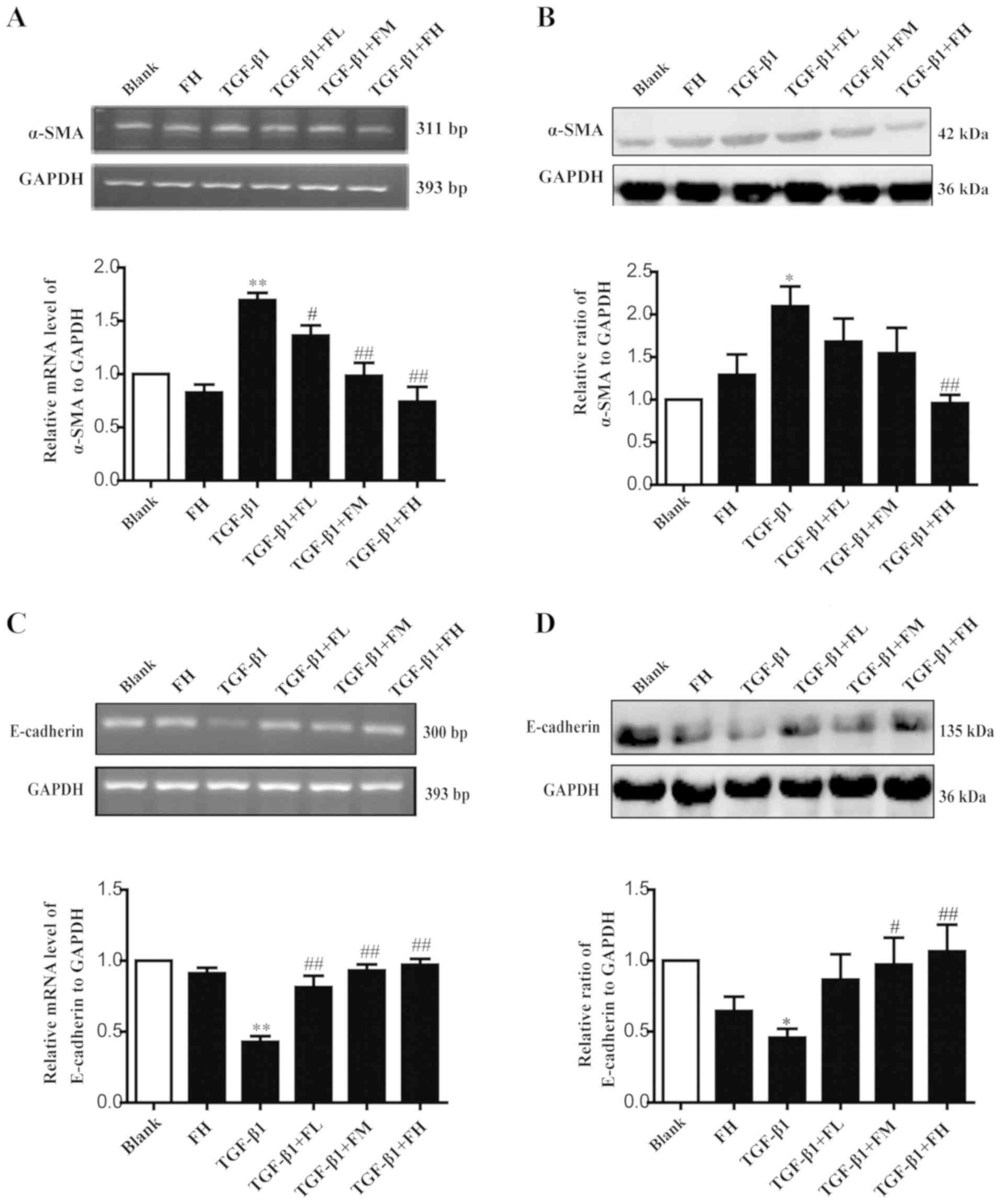 | Figure 5.Effects of febuxostat on the
expression of α-SMA and E-cadherin at the mRNA and protein levels
in MDCK cells. Incubation of MDCK cells with various concentrations
of febuxostat in the presence or absence of TGF-β1 for 48 h. (A)
mRNA and (B) protein expression levels of α-SMA were determined by
semi-qPCR and western blotting, respectively. (C) mRNA and (D)
protein expression levels of E-cadherin were determined by
semi-qPCR and western blotting, respectively. All the data are
presented as the mean ± standard error of the mean, n=3. *P<0.05
and **P<0.01 vs. blank group; #P<0.05 and
##P<0.01 vs. TGF-β1 group. α-SMA, α-smooth muscle
actin; FH, high-concentration febuxostat; FL, low-concentration
febuxostat; FM, middle-concentration febuxostat; MDCK, Madin-Darby
canine kidney; qPCR, quantitative polymerase chain reaction;
TGF-β1, transforming growth factor-β1. |
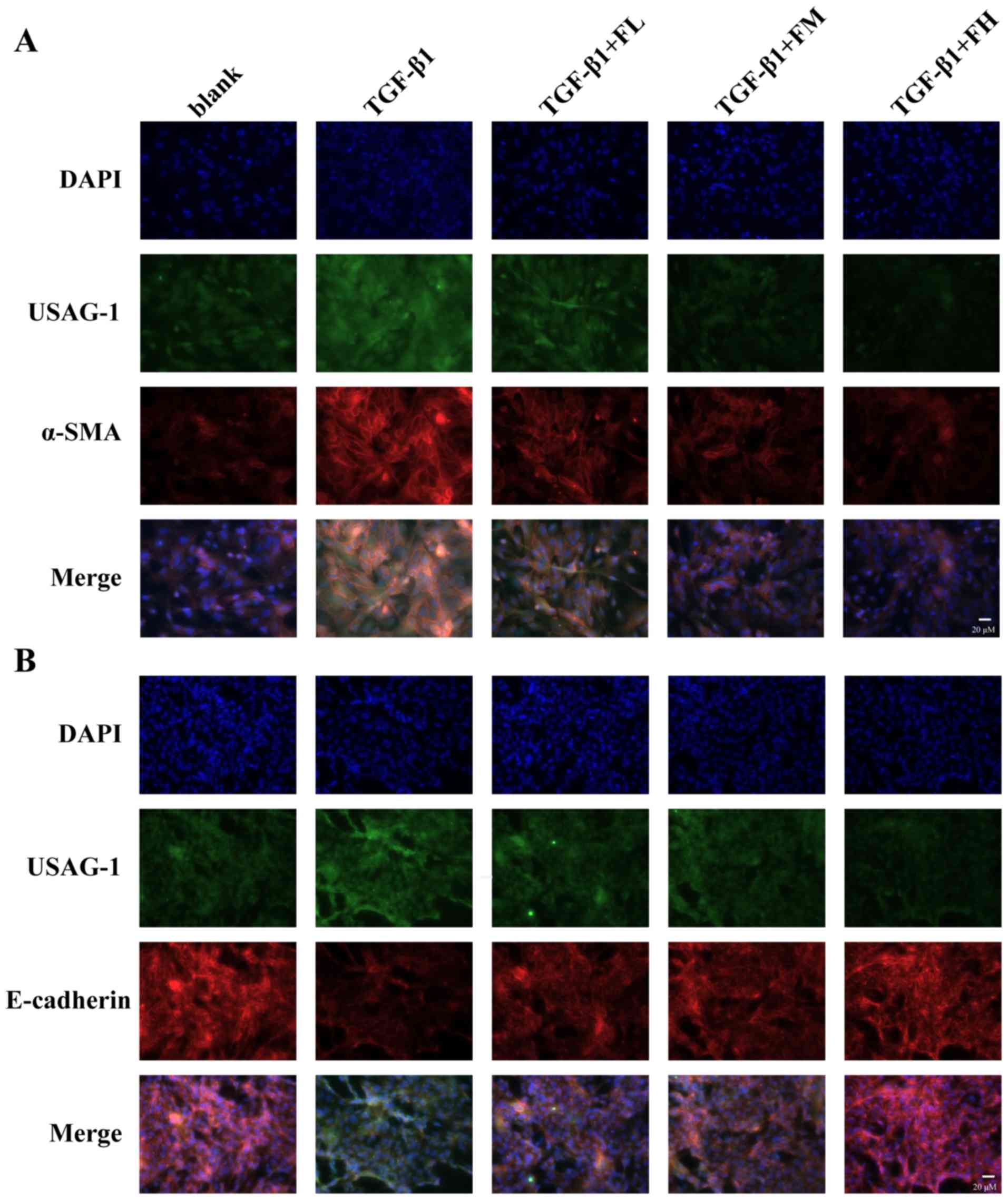
The fluorescence intensity of USAG-1 (green
staining) co-stained with α-SMA (red staining) or E-cadherin (red
staining) was further investigated by double immunofluorescence
staining (Fig. 6). MDCK cells
treated with TGF-β1 exhibited a strong fluorescence intensity for
USAG-1 and α-SMA, but the fluorescence intensity for E-cadherin was
weak compared with the blank group. Treatment with febuxostat
reversed these effects. Furthermore, double immunolabeling
indicated that USAG-1 co-expressed with α-SMA (Fig. 6A) or E-cadherin (Fig. 6B) in the same MDCK cells; the
notably increased labeling intensity of USAG-1 in MDCK cells tended
to be accompanied by strong α-SMA labeling intensity or a weak
labeling of E-cadherin, and vice versa.
The aforementioned results suggested that febuxostat
could reverse the occurrence of EMT in MDCK cells induced by
TGF-β1; the underlying mechanism may be at least partly associated
with the downregulation of USAG-1.
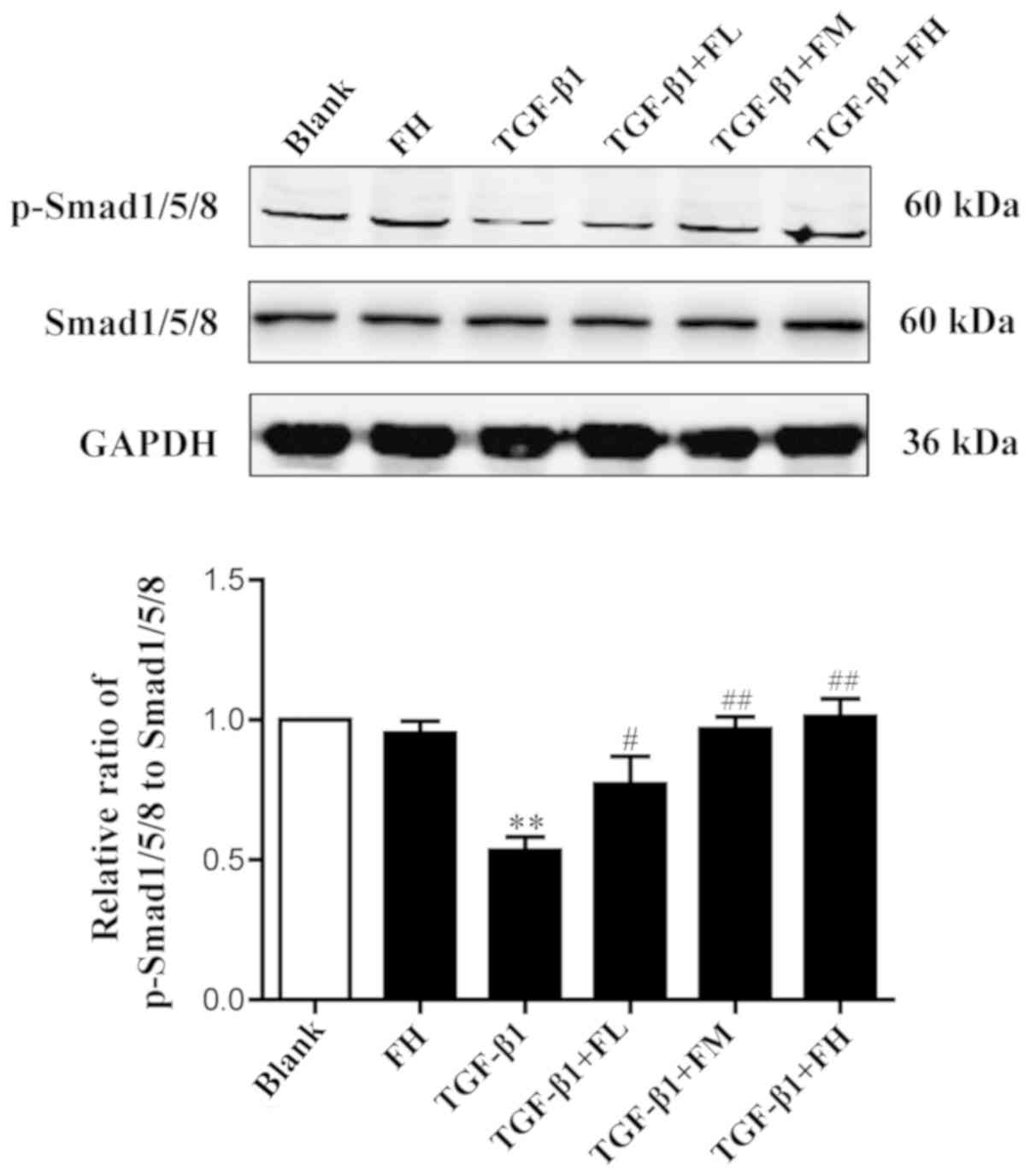
Effects of febuxostat on the activity
of the Smad1/5/8 signaling pathway in MDCK cells
To determine the effect of febuxostat on the
activity of the Smad1/5/8 signaling pathway, the levels of
p-Smad1/5/8 and total Smad1/5/8 in MDCK cells were examined.
Western blotting demonstrated that TGF-β1 significantly decreased
the expression levels of p-Smad1/5/8, indicating a reduced activity
of Smad1/5/8 signaling. Compared with the TGF-β1 group, treatment
with different concentrations of febuxostat significantly increased
the levels of p-Smad1/5/8. Febuxostat alone had no significant
effect on the expression of p-Smad1/5/8 in uninduced cells. These
results suggested that the Smad1/5/8 signaling pathway is involved
in the TGF-β1-induced EMT process, and these effects may be
inhibited by febuxostat (Fig.
7).
Discussion
Our previous study reported that febuxostat was able
to ameliorate renal tubulointerstitial fibrosis in a rat model of
UUO (37). The present study
reported that USAG-1 served an important role in the onset of
TGF-β1-induced EMT, and that febuxostat exerts its inhibitory
effect on EMT by inhibiting the expression of USAG-1 and activating
the Smad1/5/8 signaling pathway.
Renal interstitial fibrosis is the final outcome of
the development of all types of CKD, and is associated with the
progression of chronic renal failure, the main cause of end-stage
renal failure (3). Recent studies
have demonstrated that almost all types of renal injury can induce
the occurrence of EMT in renal tubular epithelial cells, which is
an important process in the pathogenesis of tubulointerstitial
fibrosis (39,40). EMT is characterized by the loss of
epithelial cell characteristics and the acquisition of mesenchymal
markers due to excessive exposure to a variety of profibrotic
cytokines (2). TGF-β1 is a
well-characterized fibrogenic cytokine, which is associated with
renal diseases and serves a key role in EMT (11). The present study revealed that
TGF-β1 could induce EMT in MDCK cells. Following 48 h of TGF-β1
treatment, EMT was readily induced in MDCK cells, as determined by
the downregulation of E-cadherin for the epithelial phenotype, and
upregulation of α-SMA for the mesenchymal phenotype. These results
demonstrated that TGF-β1 is capable of successfully inducing EMT,
which is consistent with a previous report (41).
BMP-7, formerly known as osteogenic protein-1, is a
member of the BMP-subfamily within the TGF-β superfamily, and
serves an important role in maintaining the function of renal
tubular epithelial cells (42). It
has been reported that BMP-7 could upregulate the expression of
E-cadherin in human fibroblasts induced by TGF-β1, and reduce the
secretion of proinflammatory cytokines and growth factors,
inhibiting or reversing the renal tubular EMT process; thus, the
formation of renal fibrosis is reduced (43). The Smad signaling pathway is
essential for TGF-β1-induced EMT. Increasing evidence has indicated
that BMP-7 could inhibit or reverse renal interstitial fibrosis in
animal models by regulating downstream of the Smad1/5/8 signaling
pathway (19,21,38).
BMP-7 activity in the kidney is not only determined by the
availability of BMP-7 itself, but also by the balance of
antagonists (44). USAG-1, a
specific BMP-7 antagonist that is mainly produced by renal distal
tubular epithelial cells of the adult kidney, serves a critical
role in the modulation of the renoprotective action of BMP-7
(45). Therefore, the present
study investigated the effects of USAG-1 on TGF-β1-induced EMT and
the activity of the p-Smad1/5/8 pathway in MDCK cells by
dysregulating the expression of USAG-1. The results of the present
study demonstrated that TGF-β1 upregulated the expression of USAG-1
and α-SMA, but downregulated that of E-cadherin and p-Smad1/5/8
protein, which were statistically significantly different to the
expression profiles in the control group. Conversely, siRNA-USAG-1
significantly decreased the expression of α-SMA, and increased the
protein expression levels of E-cadherin and p-Smad1/5/8. Therefore,
it may be suggested that USAG-1 is involved in promoting the
occurrence of EMT in MDCK cells, and its mechanism may be
associated with downregulation of the activity of the Smad1/5/8
signaling pathway.
Due to the important role of EMT in renal fibrosis,
any therapeutic strategy that targets the EMT may improve fibrosis.
Febuxostat, a xanthine oxidase inhibitor, achieves its therapeutic
effect by decreasing the levels of uric acid in the serum (31). The results of early animal
experiments indicated that febuxostat could alleviate kidney
dysfunction in patients, and improve renal interstitial fibrosis in
a rat model of UUO (37,46); however, whether febuxostat serves a
role in preventing renal tubular EMT induced by TGF-β1 remains
unknown. In the present study, it was observed that the protein
expression levels of USAG-1 were significantly increased in the
TGF-β1 group; however, febuxostat treatment suppressed the
expression of USAG-1 and α-SMA, while upregulating that of
E-cadherin, suggesting that the anti-EMT effect of febuxostat may
be associated with the downregulation of USAG-1 expression in MDCK
cells.
BMP-7 serves a key role in the phosphorylation of
Smad1/5/8 in the transcription of target genes. It has been
revealed that the BMP-7-Smad1/5/8 signaling pathway was suppressed
in a UUO rat model (37). The
signaling pathway is activated while kidney obstruction is
repaired, suggesting that the BMP-7-Smad1/5/8 signaling pathway
serves an important role in the treatment of renal interstitial
fibrosis (38). Therefore, the
phosphorylation levels of Smad1/5/8 in TGF-β1-induced EMT in MDCK
cells was measured in the present study. The phosphorylation levels
of BMP-7 downstream protein Smad1/5/8 appeared to be maintained in
normal MDCK cells, but was decreased following treatment with
TGF-β1. Nevertheless, the phosphorylation level of Smad1/5/8
protein was significantly increased following the treatment with
febuxostat in MDCK cells. Therefore, it is also possible that
febuxostat may downregulate the expression of the BMP-7 antagonist
USAG-1, which in turn enhances the activity of the Smad1/5/8
signaling pathway, ultimately inhibiting TGF-β1-induced EMT in MDCK
cells.
Homeobox protein Hox-A13 (HOXA13), a transcriptional
factor encoded by a homeobox gene, can bind to the promoter region
of USAG-1 and negatively modulating expression (47). A recent study revealed that albumin
could significantly inhibit HOXA13 expression in time- and
dose-dependent manners in HKC renal tubular epithelial cells, and
the upregulation of HOXA13 could exert a beneficial effect in
albumin-induced EMT in HKC cells (48). Additionally, it has been reported
that EMT-induced by albumin in HK-2 cells is directly mediated by
the production of reactive oxygen species (ROS) (49,50).
Therefore, the increased production of ROS may be the leading cause
of the reduced HOXA13 expression in albumin-induced EMT.
Febuxostat, a non-purine selective xanthine oxidase inhibitor, can
effectively inhibit ROS production in various diseases and
conditions such as ischemia/reperfusion injury (51,52),
atherosclerosis (53). Based the
aforementioned reports, decreased ROS production by febuxostat
could upregulate the expression of HOXA13 and subsequently inhibit
the expression of USAG-1 in MDCK cells, thereby reversing the
occurrence of TGF-β1-induced EMT; however, the exact mechanism of
action of febuxostat on USAG-1 requires further investigation.
In summary, the results of the present study
provided evidence that USAG-1 may be involved in the process of EMT
induced by TGF-β1, and febuxostat could reduce the expression of
USAG-1 and activate the BMP-7-Smad1/5/8 signaling pathway, thereby
reversing the occurrence of EMT. The present study not only
provides a theoretical basis for the treatment of renal
interstitial fibrosis using febuxostat in clinical practice, but
also proposes USAG-1 as a potential therapeutic target in the
treatment of kidney disease.
Acknowledgments
Not applicable.
Funding
The present study was funded by a grant from the
Director Fund Project of Jiangsu Key Laboratory of New Drug and
Clinical Pharmacy, Xuzhou Medical University (grant no.
ZR-XY201408).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AX, LL and JZ made substantial contributions to the
conception and design of the present study. YZ and LL performed the
experiments. AX, LL and YZ wrote the manuscript. YW and SZ
performed the statistical analysis and edited the manuscript. All
authors agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the study
are appropriately investigated and resolved. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Coca SG, Singanamala S and Parikh CR:
Chronic kidney disease after acute kidney injury: A systematic
review and meta-analysis. Kidney Int. 81:442–448. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boor P, Ostendorf T and Floege J: Renal
fibrosis: Novel insights into mechanisms and therapeutic targets.
Nat Rev Nephrol. 6:643–656. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Becker GJ and Hewitson TD: The role of
tubulointerstitial injury in chronic renal failure. Curr Opin
Nephrol Hypertens. 9:133–138. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wynn TA: Common and unique mechanisms
regulate fibrosis in various fibroproliferative diseases. J Clin
Invest. 117:524–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sulikowska B, Rutkowski B, Marszałek A and
Manitius J: The role of interstitial changes in the progression of
chronic kidney disease. Postepy Hig Med Dosw (Online). 69:830–837.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Levin A: Improving global kidney health:
International society of nephrology initiatives and the global
kidney health atlas. Ann Nutr Metab. 72 Suppl 2:S28–S32. 2018.
View Article : Google Scholar
|
|
7
|
Jager KJ and Fraser SDS: The ascending
rank of chronic kidney disease in the global burden of disease
study. Nephrol Dial Transplant. 32 Suppl_2:ii121–ii128. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jha V, Garcia-Garcia G, Iseki K, Li Z,
Naicker S, Plattner B, Saran R, Wang AY and Yang CW: Chronic kidney
disease: Global dimension and perspectives. Lancet. 382:260–272.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun YB, Qu X, Caruana G and Li J: The
origin of renal fibroblasts/myofibroblasts and the signals that
trigger fibrosis. Differentiation. 92:102–107. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cannito S, Novo E, di Bonzo LV, Busletta
C, Colombatto S and Parola M: Epithelial-mesenchymal transition:
From molecular mechanisms, redox regulation to implications in
human health and disease. Antioxid Redox Signal. 12:1383–1430.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Meng XM, Nikolic-Paterson DJ and Lan HY:
TGF-β: The master regulator of fibrosis. Nat Rev Nephrol.
12:325–338. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu JH, He L, Zou ZM, Ding ZC, Zhang X,
Wang H, Zhou P, Xie L, Xing S and Yi CZ: A novel inhibitor of
homodimerization targeting MyD88 ameliorates renal interstitial
fibrosis by counteracting TGF-β1-induced EMT in vivo and in vitro.
Kidney Blood Press Res. 43:1677–1687. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bae E, Kim SJ, Hong S, Liu F and Ooshima
A: Smad3 linker phosphorylation attenuates Smad3 transcriptional
activity and TGF-β1/Smad3-induced epithelial-mesenchymal transition
in renal epithelial cells. Biochem Biophys Res Commun. 427:593–599.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bei K, Du Z, Xiong Y, Liao J, Su B and Wu
L: BMP7 can promote osteogenic differentiation of human periosteal
cells in vitro. Mol Biol Rep. 39:8845–8851. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Singla DK, Singla R and Wang J: BMP-7
treatment increases M2 macrophage differentiation and reduces
inflammation and plaque formation in Apo E-/- Mice. PLoS One.
11:e01478972016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dudley AT, Lyons KM and Robertson EJ: A
requirement for bone morphogenetic protein-7 during development of
the mammalian kidney and eye. Genes Dev. 9:2795–2807. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo G, Hofmann C, Bronckers AL, Sohocki M,
Bradley A and Karsenty G: BMP-7 is an inducer of nephrogenesis, and
is also required for eye development and skeletal patterning. Genes
Dev. 9:2808–2820. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Almanzar MM, Frazier KS, Dube PH, Piqueras
AI, Jones WK, Charette MF and Paredes AL: Osteogenic protein-1 mRNA
expression is selectively modulated after acute ischemic renal
injury. J Am Soc Nephrol. 9:1456–1463. 1998.PubMed/NCBI
|
|
19
|
Ivanac-Janković R, Ćorić M, Furić-Čunko V,
Lovičić V, Bašić-Jukić N and Kes P: BMP-7 protein expression is
downregulated in human diabetic nephropathy. Acta Clin Croat.
54:164–168. 2015.PubMed/NCBI
|
|
20
|
Bramlage CP, Tampe B, Koziolek M, Maatouk
I, Bevanda J, Bramlage P, Ahrens K, Lange K, Schmid H, Cohen CD, et
al: Bone morphogenetic protein (BMP)-7 expression is decreased in
human hypertensive nephrosclerosis. BMC Nephrol. 11:312010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Klahr S and Morrissey J: Obstructive
nephropathy and renal fibrosis: The role of bone morphogenic
protein-7 and hepatocyte growth factor. Kidney Int Suppl.
87:S105–S112. 2003. View Article : Google Scholar
|
|
22
|
Zeisberg M and Kalluri R: Reversal of
experimental renal fibrosis by BMP7 provides insights into novel
therapeutic strategies for chronic kidney disease. Pediatr Nephrol.
23:1395–1398. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu Z, Zai-Chun X, Wun-Lun H and Yun-Yun Z:
BMP-7 attenuates TGF-β1-induced fibronectin secretion and apoptosis
of NRK-52E cells by the suppression of miRNA-21. Oncol Res.
23:147–154. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Celic T, Spanjol J, Grskovic A, Markic D,
Prebilic I, Fuckar Z and Bobinac D: Bone morphogenetic protein-7
reduces cold ischemic injury in rat kidney. Transplant Proc.
43:2505–2509. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yanagita M, Okuda T, Endo S, Tanaka M,
Takahashi K, Sugiyama F, Kunita S, Takahashi S, Fukatsu A,
Yanagisawa M, et al: Uterine sensitization-associated gene-1
(USAG-1), a novel BMP antagonist expressed in the kidney,
accelerates tubular injury. J Clin Invest. 116:70–79. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yanagita M: Balance between bone
morphogenetic proteins and their antagonists in kidney injury. Ther
Apher Dial. 11 Suppl 1:S38–S43. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Collette NM, Yee CS, Hum NR, Murugesh DK,
Christiansen BA, Xie L, Economides AN, Manilay JO, Robling AG and
Loots GG: Sostdc1 deficiency accelerates fracture healing by
promoting the expansion of periosteal mesenchymal stem cells. Bone.
88:20–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Murashima-Suginami A, Takahashi K, Sakata
T, Tsukamoto H, Sugai M, Yanagita M, Shimizu A, Sakurai T, Slavkin
HC and Bessho K: Enhanced BMP signaling results in supernumerary
tooth formation in USAG-1 deficient mouse. Biochem Biophys Res
Commun. 369:1012–1016. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tanaka M, Asada M, Higashi AY, Nakamura J,
Oguchi A, Tomita M, Yamada S, Asada N, Takase M, Okuda T, et al:
Loss of the BMP antagonist USAG-1 ameliorates disease in a mouse
model of the progressive hereditary kidney disease Alport syndrome.
J Clin Invest. 120:768–777. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tanaka M, Endo S, Okuda T, Economides AN,
Valenzuela DM, Murphy AJ, Robertson E, Sakurai T, Fukatsu A,
Yancopoulos GD, et al: Expression of BMP-7 and USAG-1 (a BMP
antagonist) in kidney development and injury. Kidney Int.
73:181–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yisireyili M, Hayashi M, Wu H, Uchida Y,
Yamamoto K, Kikuchi R, Shoaib Hamrah M, Nakayama T, Wu Cheng X,
Matsushita T, et al: Xanthine oxidase inhibition by febuxostat
attenuates stress-induced hyperuricemia, glucose dysmetabolism, and
prothrombotic state in mice. Sci Rep. 7:12662017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Day RO, Kamel B, Kannangara DR, Williams
KM and Graham GG: Xanthine oxidoreductase and its inhibitors:
Relevance for gout. Clin Sci (Lond). 130:2167–2180. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chinchilla SP, Urionaguena I and
Perez-Ruiz F: Febuxostat for the chronic management of
hyperuricemia in patients with gout. Expert Rev Clin Pharmacol.
9:665–673. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sircar D, Chatterjee S, Waikhom R, Golay
V, Raychaudhury A, Chatterjee S and Pandey R: Efficacy of
febuxostat for slowing the GFR decline in patients with CKD and
asymptomatic hyperuricemia: A 6-month, double-blind, randomized,
placebo-controlled trial. Am J Kidney Dis. 66:945–950. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fahmi AN, Shehatou GS, Shebl AM and Salem
HA: Febuxostat exerts dose-dependent renoprotection in rats with
cisplatin-induced acute renal injury. Naunyn Schmiedebergs Arch
Pharmacol. 389:819–830. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee HJ, Jeong KH, Kim YG, Moon JY, Lee SH,
Ihm CG, Sung JY and Lee TW: Febuxostat ameliorates diabetic renal
injury in a streptozotocin-induced diabetic rat model. Am J
Nephrol. 40:56–63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cao J, Li Y, Peng Y, Zhang Y, Li H, Li R
and Xia A: Febuxostat prevents renal interstitial fibrosis by the
activation of BMP-7 signaling and inhibition of USAG-1 expression
in rats. Am J Nephrol. 42:369–378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Manson SR, Niederhoff RA, Hruska KA and
Austin PF: The BMP-7-Smad1/5/8 pathway promotes kidney repair after
obstruction induced renal injury. J Urol. 185 (6
Suppl):S2523–S2530. 2011. View Article : Google Scholar
|
|
39
|
Liu H, Xiong J, He T, Xiao T, Li Y, Yu Y,
Huang Y, Xu X, Huang Y, Zhang J, et al: High uric acid-induced
epithelial-mesenchymal transition of renal tubular epithelial cells
via the TLR4/NF-κB signaling pathway. Am J Nephrol. 46:333–342.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Song S, Qiu D, Luo F, Wei J, Wu M, Wu H,
Du C, Du Y, Ren Y, Chen N, et al: Knockdown of NLRP3 alleviates
high glucose or TGFB1-induced EMT in human renal tubular cells. J
Mol Endocrinol. 61:101–113. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zeisberg M, Hanai J, Sugimoto H, Mammolo
T, Charytan D, Strutz F and Kalluri R: BMP-7 counteracts
TGF-beta1-induced epithelial-to-mesenchymal transition and reverses
chronic renal injury. Nat Med. 9:964–968. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sun J, Yin A, Zhao F, Zhang W and Lv J and
Lv J: Protection of tubular epithelial cells during renal injury
via post-transcriptional control of BMP7. Mol Cell Biochem.
435:141–148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zeisberg M, Shah AA and Kalluri R: Bone
morphogenic protein-7 induces mesenchymal to epithelial transition
in adult renal fibroblasts and facilitates regeneration of injured
kidney. J Biol Chem. 280:8094–8100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Massagué J and Chen YG: Controlling
TGF-beta signaling. Genes Dev. 14:627–644. 2000.PubMed/NCBI
|
|
45
|
Yanagita M, Oka M, Watabe T, Iguchi H,
Niida A, Takahashi S, Akiyama T, Miyazono K, Yanagisawa M and
Sakurai T: USAG-1: A bone morphogenetic protein antagonist
abundantly expressed in the kidney. Biochem Biophys Res Commun.
316:490–500. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Omori H, Kawada N, Inoue K, Ueda Y,
Yamamoto R, Matsui I, Kaimori J, Takabatake Y, Moriyama T, Isaka Y
and Rakugi H: Use of xanthine oxidase inhibitor febuxostat inhibits
renal interstitial inflammation and fibrosis in unilateral ureteral
obstructive nephropathy. Clin Exp Nephrol. 16:549–556. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hamasaki Y, Doi K, Okamoto K, Ijichi H,
Seki G, Maeda-Mamiya R, Fujita T and Noiri E:
3-Hydroxy-3-methylglutaryl-coenzyme A reductase inhibitor
simvastatin ameliorates renal fibrosis through HOXA13-USAG-1
pathway. Lab Invest. 92:1161–1170. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Peng L, He Q, Li X, Shuai L, Chen H, Li Y
and Yi Z: HOXA13 exerts a beneficial effect in albumin-induced
epithelial-mesenchymal transition via the glucocorticoid receptor
signaling pathway in human renal tubular epithelial cells. Mol Med
Rep. 14:271–276. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lee JH, Kim JH, Kim JS, Chang JW, Kim SB,
Park JS and Lee SK: AMP-activated protein kinase inhibits TGF-β-,
angiotensin II-, aldosterone-, high glucose-, and albumin-induced
epithelial-mesenchymal transition. Am J Physiol Renal Physiol.
304:F686–F697. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lee JY, Chang JW, Yang WS, Kim SB, Park
SK, Park JS and Lee SK: Albumin-induced epithelial-mesenchymal
transition and ER stress are regulated through a common ROS-c-Src
kinase-mTOR pathway: Effect of imatinib mesylate. Am J Physiol
Renal Physiol. 300:F1214–F1222. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang S, Li Y, Song X, Wang X, Zhao C, Chen
A and Yang P: Febuxostat pretreatment attenuates myocardial
ischemia/reperfusion injury via mitochondrial apoptosis. J Transl
Med. 13:2092015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tsuda H, Kawada N, Kaimori JY, Kitamura H,
Moriyama T, Rakugi H, Takahara S and Isaka Y: Febuxostat suppressed
renal ischemia-reperfusion injury via reduced oxidative stress.
Biochem Biophys Res Commun. 427:266–272. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nomura J, Busso N, Ives A, Matsui C,
Tsujimoto S, Shirakura T, Tamura M, Kobayashi T, So A and Yamanaka
Y: Xanthine oxidase inhibition by febuxostat attenuates
experimental atherosclerosis in mice. Sci Rep. 4:45542014.
View Article : Google Scholar : PubMed/NCBI
|