Introduction
Lung cancer is one of the most common solid tumors
and has very high global incidence and mortality rates (1). Although therapeutic treatment of lung
cancer has made great progress, the prognosis of lung cancer
patients remains unsatisfactory and treatment causes a number of
side effects (2). Natural dietary
molecules are promising candidates for novel therapeutic agents in
cancer. Green tea is one of the most widely consumed beverages,
worldwide (3). Furthermore,
(−)-epigallocatechin-3-gallate (EGCG), the main component of green
tea polyphenols, was demonstrated to have various biological
activities and demonstrates potential as a chemical and therapeutic
agent in several diseases, including various cancers (3–8) and
atherosclerosis (9).
Long non-coding RNAs (lncRNAs) are defined as a
class of transcripts longer than 200 nucleotides (nt) in the
cytoplasm and nucleus (10–13).
LncRNAs, characterized by the complexity and diversity of their
sequences, have been implicated in a wide spectrum of cellular
activities and diseases (14,15).
Emerging studies have demonstrated that certain lncRNAs are
frequently abnormally regulated in several types of cancer and
serve important roles in the occurrence and development of tumors
(16–19). Accumulating evidence has confirmed
the activity of EGCG in lung cancer therapy and many lncRNAs and
mRNAs are involved in regulating tumorigenesis (10,20–22).
However, microarray analysis of the differential expression
profiles of lncRNAs and mRNAs in lung cancer cells treated with
EGCG has not been reported.
In the present study, the EGCG-regulated lncRNAs and
mRNAs were investigated in lung cancer cells by bioinformatics
analysis. Based on the microarray analysis, a set of lncRNAs and
mRNAs was identified whose expression levels were significantly
modulated by EGCG in lung cancer cells, some of which are novel.
However, the extent to which these genes are associated with lung
cancer cells through EGCG remains unclear and needs to be addressed
in the near future.
Materials and methods
Cell culture and cell proliferation
assay
The human lung cancer cell lines A549 (lung
adenocarcinoma) and NCI-H460 (large cell lung carcinoma) were
cultured in Dulbecco's modified Eagle's medium containing 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Cells were plated in 96-well plates (5,000 cells/well) and
were incubated overnight (37°C, 5% CO2), followed by
treatment with EGCG (20, 40, 80, 160, 320 µM) and 0 µM EGCG
(control group) for 24 or 48 h respectively. The MTT-based assay
(Sangon Biotech Co., Ltd., Shanghai, China) was performed to
determine viable cell numbers at 37°C and DMSO was used to dissolve
the formazan. Absorbance at 570 nm was measured and the 50% maximal
inhibitory concentration (IC50) of EGCG for A549 and
NCI-H460 cells were calculated by Graphpad Prism version 6.0
(GraphPad Software, Inc., CA, USA).
RNA extraction and quality
control
Total RNA was extracted from 1×106 A549
or NCI-H460 cells treated with different concentration of EGCG and
control group respectively, using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), purified with a mirVana miRNA
Isolation kit (Ambion; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol, and was quantified using a
spectrophotometer (NanoDrop ND-1000; NanoDrop, Wilmington, DE,
USA). The RNA integrity of each sample was assessed by capillary
electrophoresis using the RNA 6000 Nano Lab-on-a-Chip kit and
Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). Only
RNA extracts with RNA integrity values >6 were subjected to
further analysis.
RNA amplification, labeling, and
hybridization
complementary (c)DNA labeled with a fluorescent dye
(Cy5 or Cy3-deoxycytidine triphosphate; CapitalBio Technology Co.,
Ltd., Beijing, China) was produced by Eberwine's linear RNA
amplification method as previously described (23), and subsequently RNase H enzymatic
reaction (37°C for 45 min, and followed by 95°C for 5 min). The
labeled cDNAs were purified using a Capital Bioc RNA Amplification
and Labeling kit (CapitalBio Corporation, Beijing, China) and then
hybridized with specific probes (CapitalBio Technology Co., Ltd.)
in a hybridization oven (Xinghua Analytical Instrument Factory,
Jiangsu, China) overnight at 45°C.
Microarray analysis
The expression levels of lncRNAs and mRNAs were
determined using Gene Spring software V13.0 (Agilent Technologies).
The differentially expressed lncRNAs and mRNAs between the
EGCG-treated and control groups were identified based on the
threshold values of ≥2 and ≤-2 fold-change and paired Student's
t-test P<0.05. Data were log2-transformed and median-centered
using genes and Adjust Data function of CLUSTER 3.0 software
(http://bonsai.hgc.jp/~mdehoon/software/cluster/software.htm)
and analyzed by hierarchical clustering with average linkages. Tree
visualization was performed using the Java TreeView (Stanford
University School of Medicine, Stanford, CA, USA; http://sourceforge.net/projects/jtreeview/files).
Gene-lung cancer associations were investigated by using the
DisGeNET database (http://www.disgenet.org/web/DisGeNET/menu), which
records disease-associated genes and provides literature support,
as in the previous reference (24).
Gene Ontology (GO), pathway and
disease analysis
GO analysis was performed using an online database
(www geneontology.org). The GO database
provides a network of three structured definition terms that
describe the properties of a gene product (25). Pathway analysis of the
differentially expressed genes was performed according to the Kyoto
Encyclopedia of Genes and Genomes (KEGG) database (www.genome.jp/kegg/). Specifically, Fisher's exact
test and χ2 tests were performed to classify the GO
category and select the significant pathway. The false discovery
rate (FDR) was calculated and the threshold of significance was
defined as P<0.05. FunDO database (http://fundo.nubic.northwestern.edu/) was used for
disease analysis.
Network analysis of EGCG target
genes
Search Tool for Interactions of Chemicals (STITCH;
http://stitch.embl.de/) is a database of
interactions and correlations between compounds and genes. The EGCG
direct target genes were searched and analyzed by using STITCH
according to the reference (26).
Search Tool for the Retrieval of Interacting Genes (STRING,
http://string-db.org/) is a database of
protein-protein interactions (27). The interaction of EGCG direct
target gene and differential expression oncogene affected by EGCG
was determined in the STRING database, and only the interactions
with a combined score > 0.4 were considered as significant.
Finally, the network of compound and gene interactions was
mapped.
Analysis of lncRNA binding
transcription factors
We downloaded the transcriptional factors for each
of the 20 differential expression oncogenes most affected by EGCG
treatment from the UCSC Genome Browser (http://genome.ucsc.edu/cgi-bin/hgTracks?hgsid=698152999_7PLacukNAMsjS2GSMIFtIxAic9jW).
Then, the lncRNA-transcriptional factor-oncogene regulatory network
was constructed by using the LncPro database (http://bioinfo.bjmu.edu.cn/lncpro/) as previously
described (28).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The Total RNA of 1×106 A549 cells treated
with 80 µM EGCG for 48 h and the control group cells was extracted
as described above respectively. The PrimeScript™ RT
Reagent kit (Takara Bio, Inc., Otsu, Japan) was used for cDNA
synthesis and genomic DNA removal. qPCR was performed using
SYBR® Premix Ex Taq™ (Takara Bio, Inc., Otsu, Japan) and
in an ABI 7000 instrument (Applied Biosystems; Thermo Fisher
Scientific, Inc.) for 40 cycles (95°C for 15 sec, 60°C for 1 min)
after an initial 3 min degeneration at 95°C, and β-actin was used
as an internal control (primers for qPCR listed in Table I). The relative gene expression
data was analyzed by 2−ΔΔCq method as previously
described (29).
 | Table I.The primers for RT-qPCR. |
Table I.
The primers for RT-qPCR.
| Gene | Name | Sequence |
|---|
|
ENSG00000272796.1 | Sense |
CATTGGACTGGGTGAGGC |
|
| Antisense |
TTGAGCACGGTAGAGGAGAC |
|
ENSG00000254054.2 | Sense |
GATTCCACTGATATTTACTGAA |
|
| Antisense |
GGACCGCTCCTTTGATGC |
|
ENSG00000260630.2 | Sense |
GCCGTGAGTGAAGGGCAGAG |
|
| Antisense |
TTCCAGAGCCGCCAGTAGGG |
|
ENSG00000235142.2 | Sense |
AGCTGGAATGCAGATGGG |
|
| Antisense |
AGCACAGGCTCAAGGGAC |
|
ENSG00000224063.1 | Sense |
AGAACTGATTTTAGAATGCCA |
|
| Antisense |
CCAATGTATTTGCCAAGA |
|
ENSG00000251018.2 | Sense |
AGACCTATTGGAACTGACT |
|
| Antisense |
AGAACCTGAGTGCCTTGT |
|
ENSG00000226403.1 | Sense |
CTGGTCCTCGCAGTCCGC |
|
| Antisense |
CTCTTTCCCAAAGGGCAC |
| PSMC3IP | Sense |
GCGGATCAGGACCAGTTT |
|
| Antisense |
TTTGACCAGGACTAGGCG |
|
ENSG00000230109.1 | Sense |
AAAACTATGAGAAAACTGGGTC |
|
| Antisense |
ATGTAAGTTTCTGATTGGTCC |
|
ENSG00000130600.10 | Sense |
GAGGAGCTGAGTGGGACC |
|
| Antisense |
TTGATGTTGGGCTGATGAG |
| ALDOC | Sense |
GGCATCAAGGTTGACAAGGG |
|
| Antisense |
GCTGGCAGATACTGGCATAA |
| HABP2 | Sense |
AATAAGTGTCAGAAAGTGCAAAA |
|
| Antisense |
CAGCGGTAGTAGGGAGGA |
| CFB | Sense |
TATGAAGACCACAAGTTGAAGT |
|
| Antisense |
GTATAGCAAGTCCCGGATC |
| IL11 | Sense |
TGCACAGCTGAGGGACAA |
|
| Antisense |
CCGCAGGTAGGACAGTAGG |
| SMOC1 | Sense |
AGATGACGGGTCTAAGCC |
|
| Antisense |
ATCACCAAGTGTTTAATCCATA |
| AREG | Sense |
CTCGGCTCAGGCCATTAT |
|
| Antisense |
AGCCAGGTATTTGTGGTTCG |
| EREG | Sense |
CTGGGTTTCCATCTTCTA |
|
| Antisense |
TGTTATTGACACTTGAGCC |
| MTRNR2L6 | Sense |
TAGGGACTTGTATGAATGAC |
|
| Antisense |
ATAGGTTGCTCGGAGGTT |
| POLR3G | Sense |
CACCTGAAGAAAGACAAG |
|
| Antisense |
TCAGTATTAGTGAGTGGTGT |
| LYAR | Sense |
TTTCTGGGGCGATGACTA |
|
| Antisense |
TTGGGGCTGACATTGGGT |
| β-actin | Sense |
AGGGGCCGGACTCGTCATACT |
|
| Antisense |
GGCGGCACCACCATGTACCCT |
Statistical analysis
The experiments were performed three times. All
results are expressed as the mean ± standard deviation. A paired
Student's t test was performed to compare the two groups in the
microarray analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Analysis of cell proliferation
Treatment with EGCG inhibited lung cancer cell
proliferation in dose-dependent manner (Fig. 1A). EGCG inhibited the growth of
A549 cells, with IC50 values of 122.4 and 108.2 µM at 24
and 48 h, respectively. Additionally, the IC50 values of
EGCG in NCI-H460 cells were 693.2 and 485.1 µM at 24 and 48 h,
respectively. These results demonstrated that A549 cells are more
sensitive to EGCG than NCI-H460 cells. Therefore, A549 cells
treated with 80 µM (≈IC30) for 48 h were used for
microarray analysis.
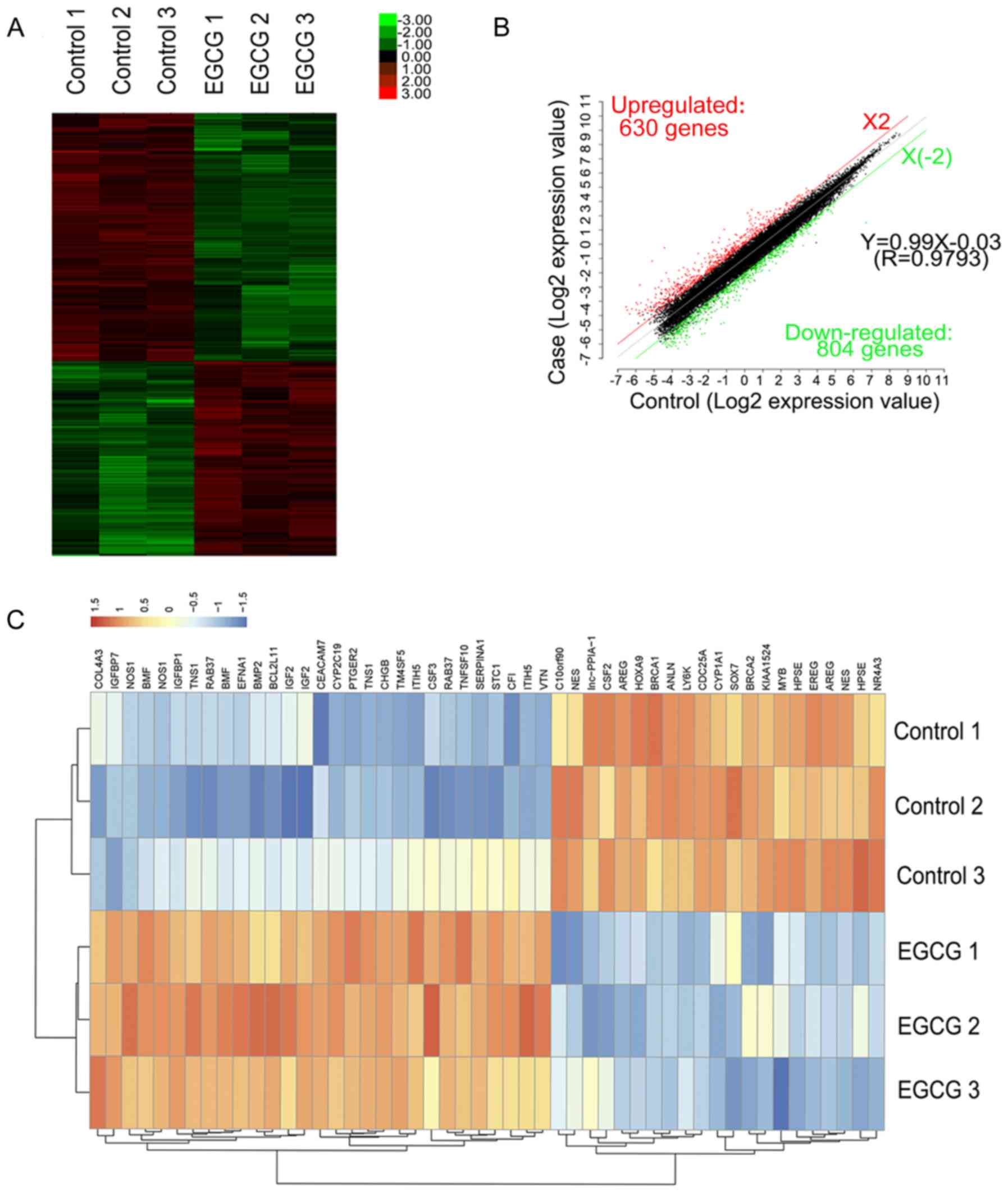
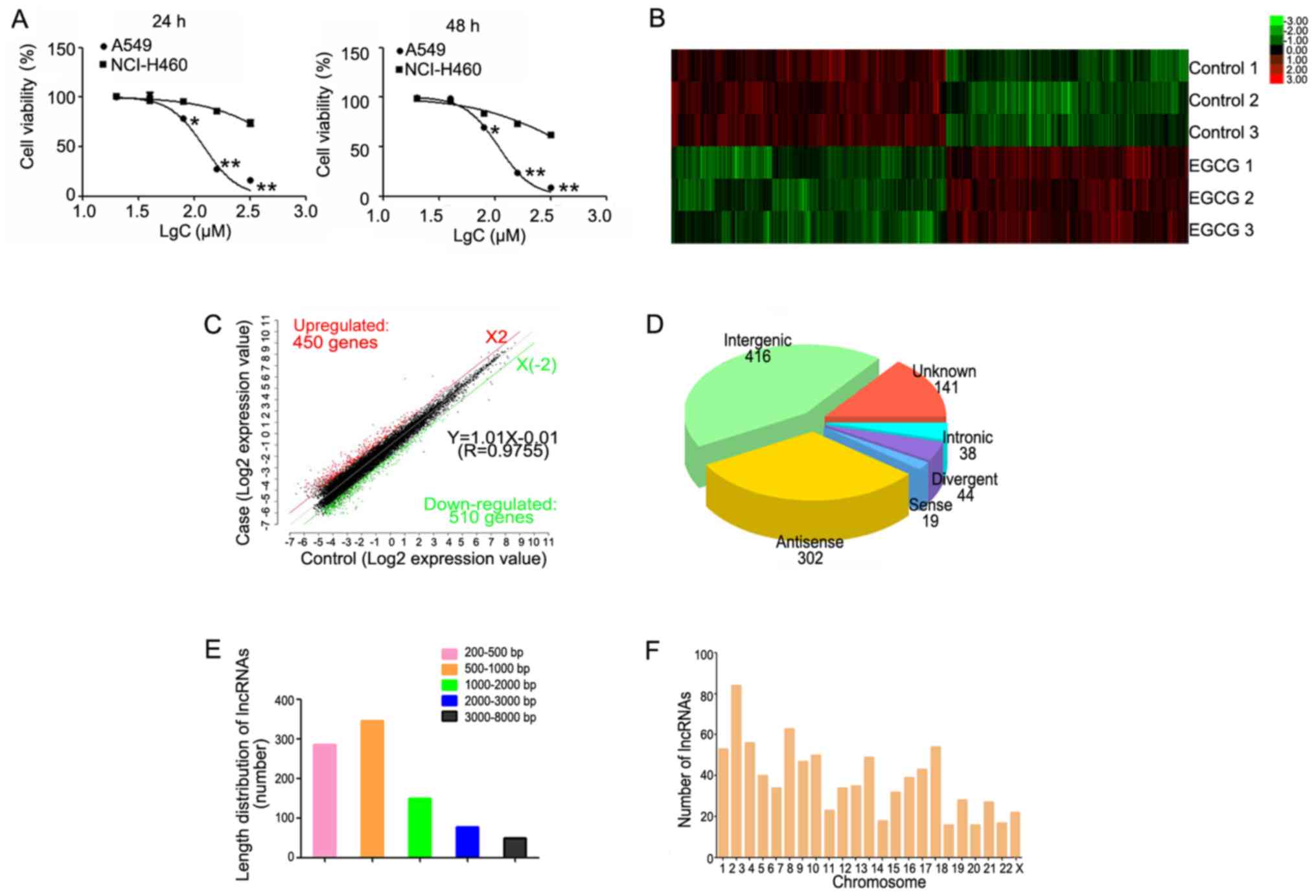 | Figure 1.Hierarchical clustering of
differentially expressed lncRNAs. (A) The survival rate of lung
cancer cells following treatment with EGCG detected by MTT assay.
(B) Hierarchical clustering of 3 EGCG-treated samples (EGCG1-3)
versus 3 untreated samples (CON1-3) was based on the 960
dysregulated lncRNAs. Red represents upregulated lncRNAs, while
green represents downregulated lncRNAs. (C) Red, black, and green
dots represent lncRNAs with expression that was increased,
equivalent, or decreased between the EGCG-treated and control
groups. (D) Length distribution of 960 differentially expressed
lncRNAs. (E) Class distribution of dysregulated lncRNAs, including
416 intergenic, 302 antisense, 141 unknown, 44 divergent, 38
intronic, and 19 sense lncRNAs. (F) Chromosome enrichment analysis
of 960 differentially expressed lncRNAs on chromosomes (DNA).
*P<0.05, **P<0.01 vs. NCI-H460. EGCG,
(−)-epigallocatechin-3-gallate; lncRNA, long non-coding RNA. |
Analysis of differentially expressed
lncRNAs
A total of 19,446 lncRNAs were detected by Gene
Spring software V13.0. Based on the results of the lncRNA
expression profile, differentially expressed lncRNAs between the
EGCG-treated and control groups were identified. According to their
expression levels in the samples, the clusters were hierarchically
clustered (Fig. 1B). The threshold
was set to a fold-change ≥2, P<0.05, and FDR <0.05 and a
total of 960 differential expressed lncRNAs, including 450 and 510
up- and downregulated lncRNAs, respectively were identified
(Fig. 1C).
Among the differentially expressed lncRNAs, there
were 416 intergenic, 302 antisense, 141 unknown, 44 divergent, 38
intronic, and 19 sense lncRNAs (Fig.
1D). The length of the differentially expressed lncRNAs ranged
from 200 base pairs (bp) to 8 kb, with the lengths of 660 lncRNAs
(68.75%) ranging between 200–1,000 bp (Fig. 1E). Additionally, these
differentially expressed lncRNAs were distributed in nearly all
human chromosomes (Fig. 1F).
Analysis of differentially expressed
mRNAs
A total of 1,434 mRNAs were detected whose
expression significantly differed between lung cancer cells treated
with EGCG and the control group (fold-change ≥2.0, P<0.05, and
FDR <0.05). Of these, 804 mRNAs were downregulated and 630 were
upregulated. Through hierarchical clustering analysis, different
expression patterns were predicted (Fig. 2A and B). The top 10 upregulated and
10 downregulated mRNAs between the two groups are listed in
Table II. The top 5 upregulated
and 5 downregulated mRNAs associated with lung cancer are listed in
Table III. Genes associated with
lung cancer in the DisGeNET database were investigated. By
comparing the results of the database with the results obtained
upon EGCG treatment, 168 known lung cancer genes in the
EGCG-treated groups were identified, and the top 50- changed genes
were plotted as a heat map (Fig.
2C).
| Table II.Top 10 up and downregulated mRNAs
following EGCG treatment. |
Table II.
Top 10 up and downregulated mRNAs
following EGCG treatment.
|
| Upregulated
mRNAs | Downregulated
mRNAs |
|---|
|
|
|
|
|---|
| mRNA | FC(abs) | mRNA | FC(abs) |
|---|
| CEACAM7 | 36.10 | MTRNR2L6 | 18.14 |
| AGT | 30.47 | CYP1A1 | 10.22 |
| TF | 15.28 | ID4 | 9.65 |
| TNS1 | 10.05 | CORO1A | 8.27 |
| SMOC1 | 9.04 | HMGCS2 | 7.21 |
| IL11 | 8.17 | MTRNR2L8 | 6.22 |
| ALDOC | 8.15 | SP5 | 6.14 |
| NRAP | 8.06 | HPDL | 5.72 |
| KCNJ16 | 7.61 | GREB1 | 5.58 |
| BMF | 7.59 | CEMIP | 5.53 |
| Table III.Top 5 up- and downregulated mRNAs
associated with lung cancer. |
Table III.
Top 5 up- and downregulated mRNAs
associated with lung cancer.
| Author, year | Genesymbol | FC(abs) | Regulation | Reported function
in lung cancer | (Refs.) |
|---|
| Pan et al,
2016 | TTR | 6.91 | Up | Reduced
transthyretin expression in sera of lung cancer | (21) |
| Liu et al,
2007 | SLC5A10 | 5.21 | Up | DNA methylation in
SLC5A8 expression in lung cancer cell lines, Reduced or lost
expression of SLC5A8 was observed in tumor tissues | (50) |
| Park et al,
2013 and Tsai et al, 2014 | RAB37 | 4.73 | Up | Lung cancer
patients with metastasis and poor survival show low hRAB37 protein
expression | (51) |
|
|
|
|
| Knockdown of hRAB37
in low metastasis cell line led to a significant increase in cell
migration | (52) |
| Wu et al,
2009 | ITIH5 | 3.46 | Up | ITIH5 mRNA
expression was significantly decreased in NSCLC compared to normal
lung tissue, ITIH5 may be a novel putative tumor suppressor gene in
NSCLC | (53) |
| Dötsch et
al, 2015 | IGFBP7 | 3.08 | Up | (IGFBP7) was
considered a tumor suppressor gene in lung cancer. | (54) |
| Chen et al,
2011 | GREB1 | 5.58 | Down | The expression of
GREB1 in lung cancers is significantly higher than that in normal
lung tissues, and closely related to smoking | (55) |
| Zhang et al,
2012 and McLemore et al, 1990 | CYP1A1 | 10.22 | Down | A biomarker for
lung cancer risk, and closely related to smoking | (56,57) |
| Taioli et
al, 1998 | CLSPN | 5.24 | Down | Claspin contribute
to the radioresistance of lung cancer brain metastases | (58) |
| Choi et al,
2014 and Taniguchi et al, 2016 | AREG | 4.72 | Down | AREG can cause lung
tumorigenesis and resistance to crizotinib | (59,60) |
| Taron et al,
2004 | BRCA1 | 3.08 | Down | BRCA1 as an
indicator of chemoresistance in lung cancer | (61) |
RT-qPCR verification of differentially
expressed lncRNAs and mRNAs
A total of 10 differentially expressed lncRNAs were
randomly selected, including 5 upregulated (ENSG00000272796.1,
ENSG00000254054.2, ENSG00000260630.2, ENSG00000235142.2 and
ENSG00000224063.1) and 5 downregulated lncRNAs (ENSG00000251018.2,
ENSG00000226403.1, PSMC3IP, ENSG00000230109.1 and
SG00000130600.10), for verification in these lung cancer cells. The
RT-qPCR results of these 10 selected lncRNAs were consistent with
those from the microarray analysis (Fig. 3A). Additionally, 10 differentially
expressed mRNAs were also randomly selected, including 5
upregulated (aldolase C, hyaluronan-binding protein 2, complement
factor B, interleukin (IL) 11, and secreted modular calcium-binding
protein 1) and 5 downregulated mRNAs (amphiregulin, epiregulin,
humanin-like protein 6, polymerase (RNA) III (DNA Directed)
Polypeptide G, and Ly1 antibody reactive). The results for all 10
selected mRNAs were consistent with those from the microarray
analysis (Fig. 3B).
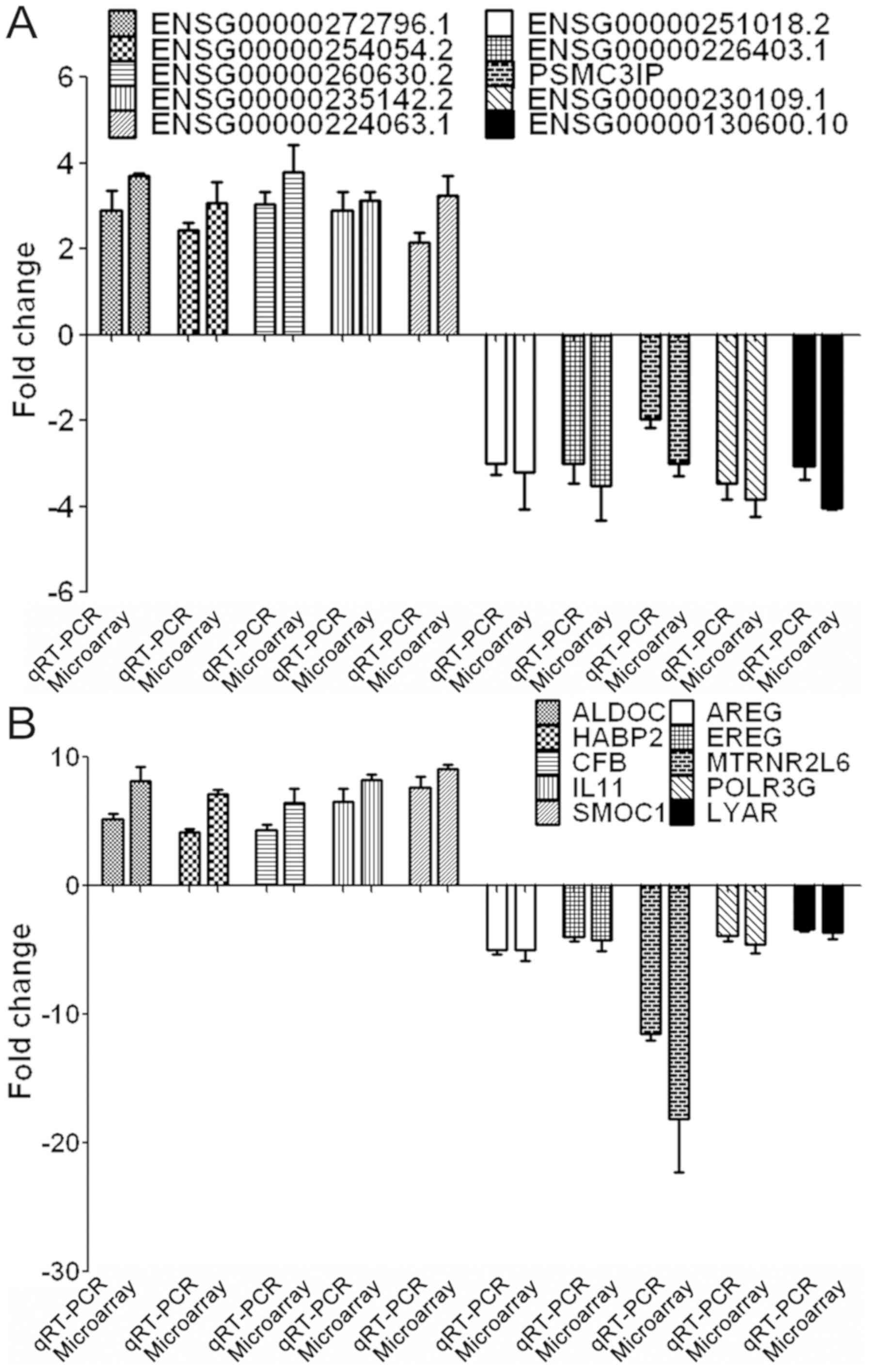 | Figure 3.RT-qPCR validation of the microarray
analysis results. (A) RT-qPCR analysis of 10 dysregulated lncRNAs,
including 5 upregulated and 5 downregulated lncRNAs. (B) RT-qPCR
analysis of 10 dysregulated mRNAs, consisting of 5 upregulated and
5 downregulated mRNAs. ALDOC, Aldolase C; AREG, Amphiregulin; CFB,
Complement factor B; EREG, Epiregulin; HABP, Hyaluronan-binding
protein; IL, Interleukin; lncRNA, long non-coding RNA; LYAR, Ly1
antibody reactive; MTRNR2L6, Humanin-like protein; POLR3G,
Polymerase (RNA) III (DNA Directed) Polypeptide G; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; SMOC,
Secreted modular calcium-binding protein. |
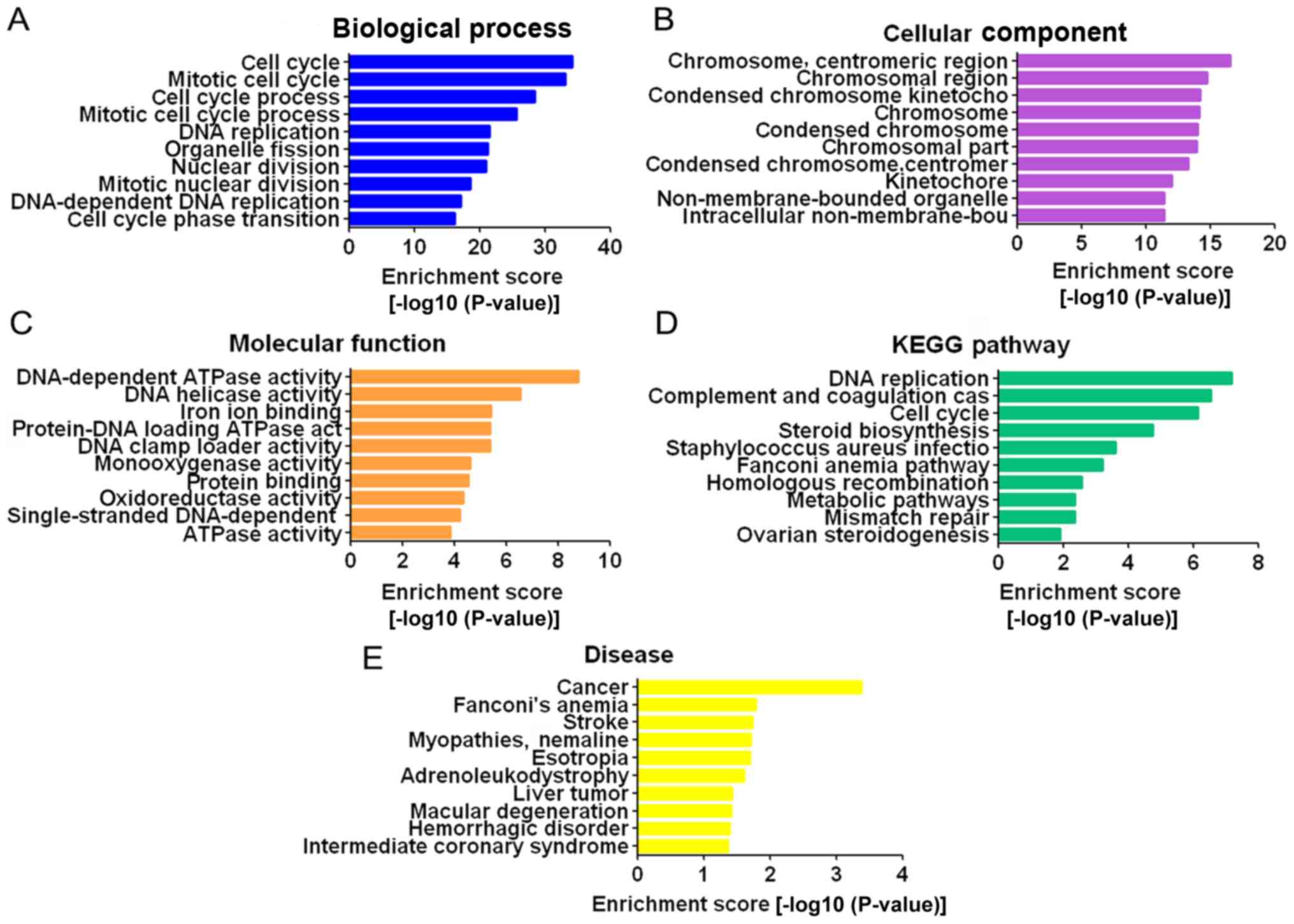
GO and pathway analysis for
differentially expressed mRNAs
GO analysis consists of three parts: Biological
processes, cellular components, and molecular functions. Through GO
analysis, it was demonstrated that the differentially expressed
mRNAs were mainly enriched in the GO terms ‘cell cycle’, ‘mitotic
cell cycle’, ‘cell cycle process’ and ‘mitotic cell cycle process’
for biological processes (Fig.
4A); ‘chromosome’, ‘centromeric region’, ‘chromosomal region’,
‘condensed chromosome kinetochore’ and ‘chromosome’ for cellular
components (Fig. 4B); and
DNA-dependent ATPase activity’, ‘DNA helicase activity’ and ‘iron
ion binding’ for molecular functions (Fig. 4C). Pathway analysis was based on
the KEGG database. The differentially expressed mRNAs were
associated with DNA replication, complement and coagulation
cascades, cell cycle and other pathways (Fig. 4D). The dysregulated mRNAs were
associated with a number of diseases, of which cancer was the most
relevant (Fig. 4E).
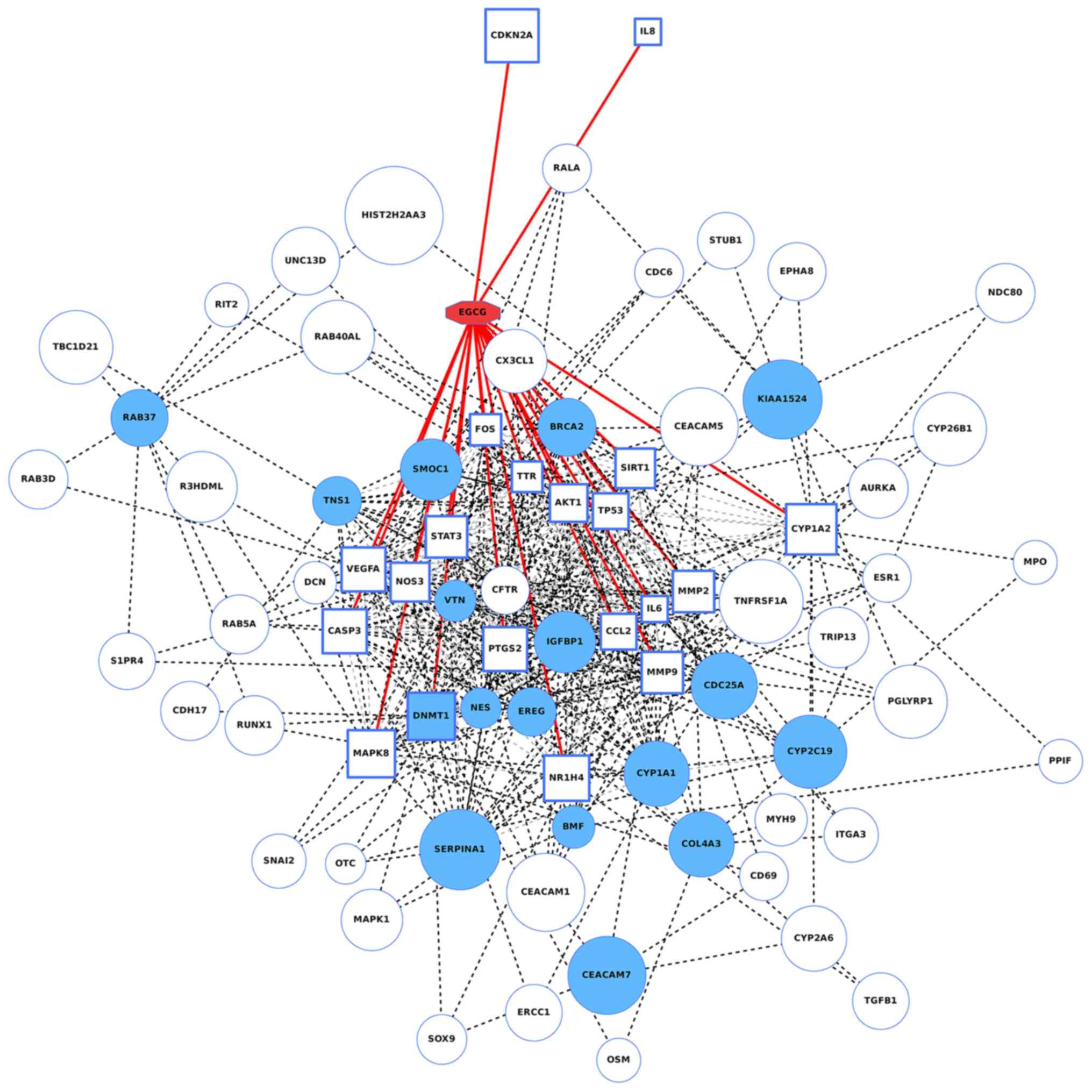
Bioinformatic analysis of potential
mechanisms of EGCG regulating lncRNAs/mRNAs
A total of 20 EGCG target genes were detected using
the STITCH database; these genes were AKT1, Caspase 3, IL-6,
prostaglandin-endoperoxide synthase 2 (PTGS2), tumor protein 53
(TP53), vascular endothelial growth factor A (VEGFA), FOS,
mitogen-activated protein kinase 8 (MAPK8), nitric oxide synthase 3
(NOS3), signal transducer and activator of transcription 3 (STAT3),
matrix metalloproteinase 9 (MMP9), C-C motif chemokine ligand 2
(CCL2), transthyretin (TTR), MMP2, DNA
(cytosine-5)-methyltransferase 1 (DNMT1), sirtuin 1 (SIRT1),
nuclear receptor subfamily 1 group H member 4 (NR1H4), cytochrome
P450 family 1 subfamily A member 2 (CYP1A2), cyclin dependent
kinase inhibitor 2A (CDKN2A), and IL-8. The genes are
annotated as being directly targeted by EGCG (26). According to the lung cancer
tissues’ gene expression levels, obtained from The Cancer Genome
Atlas database, it was demonstrated that the expression levels of
these 20 genes were closely associated to the lncRNAs regulated by
EGCG, among which DNMT1 is also a well-known lung
cancer-associated gene. All the 20 EGCG direct target genes and 20
oncogenes mostly affected by EGCG were entered into the STRING
database for network construction. The network of gene interactions
as well as compound-gene interactions were illustrated in Fig. 5. A broad linkage between EGCG
target genes and known lung cancer-associated genes suggested that
EGCG was very likely to affect NSCLC by acting on these genes.
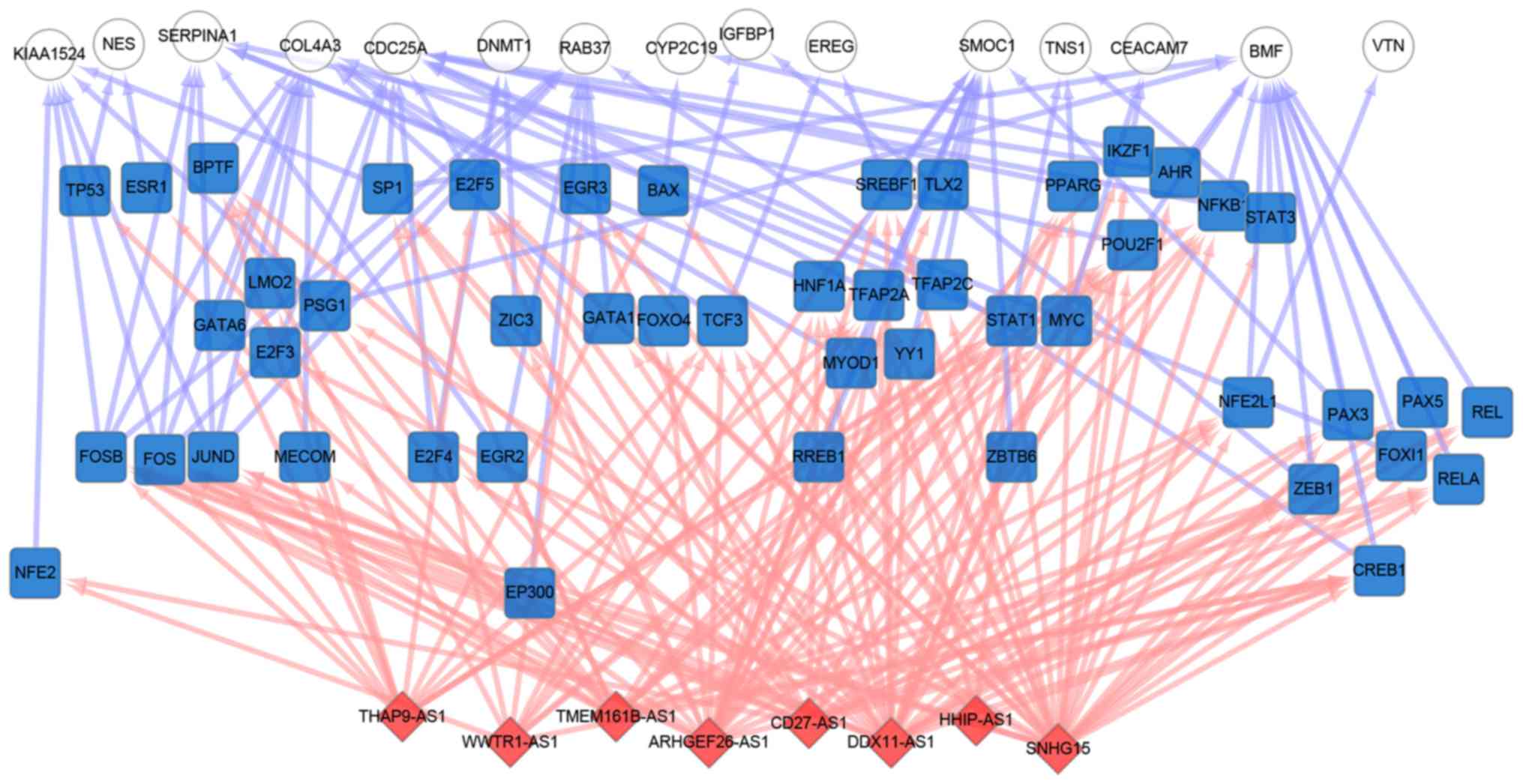
LncPro was used to predict the interaction of the
top 10 lncRNAs and top 20 oncogenes affected by EGCG. A total of 8
of the 10 lncRNAs were demonstrated to bind the transcriptional
factors (TF) of relevant oncogenes with relatively high binding
scores. The network of lncRNA-TF-oncogenes is demonstrated in
Fig. 6, where the lncRNAs can bind
to nearly all oncogene-associated TFs, including B-cell
lymphoma-2-modifying factor and collagen type IV α3 chain, which
are regulated by additional lncRNA-bound transcriptional
factors.
Discussion
EGCG, the main active and water-soluble component of
green tea polyphenols, accounting for 9–13% of the green weight of
green tea, has been demonstrated to possess anticancer activity
(30,31). The IC50 value (in A549
cells) conversion for green tea was about 0.4–2 g, while this is
less than a daily intake (15–30 g) of people who have tea drinking
habits. The golden ratio of brewing tea is 1:50, that is, 50 ml of
boiling water poured into 1 g of tea. However, there are
differences between in vitro and in vivo experiments,
which need further verification.
As a powerful cancer chemopreventive agent, the
anti-tumor effect of EGCG has been recognized in numerous studies
conducted in various countries (6,32,33);
however, it is difficult to determine the molecular mechanisms of
the anticancer effects of EGCG, particularly in vivo.
Therefore, in the present study, the expression levels of lncRNAs
and mRNAs in EGCG-treated and control groups were analyzed in
genomic studies combined with bioinformatic analysis. Based on the
bioinformatic analysis, the general characteristics, functional
comments, and pathways of the differentially expressed lncRNAs and
mRNAs were identified. The present results provide a comprehensive
understanding of the lncRNAs and mRNAs in lung cancer cells
regulated by EGCG, and may help to reveal the molecular mechanisms
of EGCG in lung cancer, but these need to be validated further.
Previous studies demonstrated that ~18% of encoded
protein genes that produce lncRNAs are associated with cancer,
while only ~9% of all encoded protein-coding genes are associated
with cancer (34). Increasing
evidence has revealed the importance of lncRNAs in lung cancer. For
example, a previous study demonstrated that Oct4 regulates the
expression of the lncRNAs nuclear paraspeckle assembly transcript 1
and metastasis associated lung adenocarcinoma transcript 1
promoting lung cancer progression (35). The lncRNA HOTAIR was demonstrated
to regulate lung cancer cell growth and metastasis (36). The lncRNA XLOC008466 was reported
as an oncogene in human non-small cell lung cancer by virtue of its
targeting miR-874 (37). However,
studies on lncRNAs regulated by EGCG in lung cancer are limited.
The present study aimed to identify significantly down- or
upregulated lncRNAs and mRNAs in lung cancer cells treated with
EGCG. First, a lung cancer cell line demonstrating sensitivity to
EGCG was selected by MTT analysis and was treated with 80 µΜ EGCG
for 48 h, followed by microarray analysis.
GO analysis revealed that these differentially
expressed mRNAs are associated with different biological processes,
including cell cycle, mitotic cell cycle, DNA replication,
organelle fission, nuclear division, mitotic nuclear division, and
cell cycle phase transition. Thus, the present results indicate
that EGCG may exert its anticancer function by regulating lung
cancer cell proliferation, including suppression of the cell cycle
and mitotic cell cycle. Furthermore, a previous mechanistic study
demonstrated that EGCG inhibits lung cancer-associated processes,
including anchorage-independent growth and the cell cycle, by
directly targeting the epidermal growth factor receptor (EGFR)
signaling pathway (38).
Additionally, a study on salivary adenoid cystic
carcinoma revealed that the EGCG anticancer effect is mediated via
the EGFR/extracellular regulated kinase (ERK) signaling
transduction and mitochondrial apoptosis pathways (34). Differential proteomic analysis
demonstrated that EGCG inhibited hepatoma-derived growth factor and
activated apoptosis to increase the chemosensitivity of non-small
cell lung cancer (39). Based on
the present results, it may be hypothesized that the various
lncRNAs, mRNAs, or proteins regulated by EGCG identified in these
studies constitute or regulate important cellular components,
including the chromosome centromeric region and chromosome region,
but this needs to be investigated further. Several studies have
confirmed that EGCG significantly inhibits the expression of
telomerase and increases the chemosensitivity of lung cancer cells
(21,40,41).
GO analysis indicated that the mRNAs induced by EGCG
are involved in various molecular functions, including
DNA-dependent ATPase activity, DNA helicase activity and iron ion
binding. A previous study reported that EGCG regulates the
cross-talk between JWA (also known as ADP Ribosylation Factor Like
GTPase 6 Interacting Protein 5) and topoisomerase Iiα regulating
lung cancer cell invasion (42).
Antioxidant supplementation is known to increase the risk of lung
cancer, and EGCG as an exemplary antioxidant reported to induce
significant cell death and DNA damage in human lung cells through a
reductive mechanism (43). KEGG
pathway enrichment analysis indicated that EGCG inhibited lung
cancer by affecting DNA replication, complement and coagulation
cascades, cell cycle, and other signaling pathways, including the
Wnt/β-catenin, unclear respirator factor2/Kelch-like
ECH-associated protein 1, mitogen-activated kinase/ERK, EGFR,
STAT3, and AKT signaling pathways (38,44–47).
However, the precise co-regulatory functions and mechanisms of
lncRNA-mRNA require further investigation.
Additionally, combined analysis of the microarray
chip sequencing and bioinformatics results obtained from databases
revealed a number of key factors in the EGCG process, including
TP53 and DNMT1, which are well-known
cancer-associated genes (48,49).
These factors also demonstrate high variability in cancer patients;
therefore, the levels of variation in these factors may affect the
anticancer effects of EGCG in patients, but additional studies are
necessary to determine the effects of these factors.
In conclusion, the present study provided the lncRNA
and RNA expression profile in vitro. In the present study,
960 lncRNAs and 1,434 mRNAs were identified to be differentially
expressed in lung cancer cells treated with EGCG compared with
those without treatment. Furthermore, through cluster analysis and
GO function and KEGG pathway analysis, it was demonstrated that
EGCG is associated with possible regulation of the expression of
several lncRNA and mRNAs, but this needs to be investigated
further.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Project of Hunan Province, China (grant no.
2013FJ3036).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
D-LH and GW conceived and designed the study. D-LH
and L-HZ performed the experiments. GW provided the financial aid
and approved the final version. JY acquired data. Y-FH, L-HZ and DW
analyzed the data. H-HZ contributed to the interpretation of the
results. Y-FH, DW and H-HZ drafted and revised the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
EGCG
|
(−)-epigallocatechin-3-gallate
|
|
lncRNAs
|
long non-coding RNAs
|
|
nt
|
nucleotides
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
GO
|
gene ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
FDR
|
false discovery rate
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
TF
|
transcription factor
|
References
|
1
|
Nørskov MS, Dahl M and Tybjærghansen A:
Genetic variation in GSTP1, lung function, risk of lung cancer and
mortality. J Thorac Oncol. 12:1664–1672. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang JC, Mok T, Han B, Orlando M, Puri T
and Park K: A review of regimens combining pemetrexed with an
epidermal growth factor receptor tyrosine kinase inhibitor in the
treatment of advanced nonsquamous non-small-cell lung cancer. Clin
Lung Cancer. 19:27–34. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cao J, Han J, Xiao H, Qiao J and Han M:
Effect of tea polyphenol compounds on anticancer drugs in terms of
anti-tumor activity, toxicology, and pharmacokinetics. Nutrients.
8:E7622016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oya Y, Mondal A, Rawangkan A, Umsumarng S,
Iida K, Watanabe T, Kanno M, Suzuki K, Li Z, Kagechika H, et al:
Down-regulation of histone deacetylase 4, −5 and −6 as a mechanism
of synergistic enhancement of apoptosis in human lung cancer cells
treated with the combination of a synthetic retinoid, Am80 and
green tea catechin. J Nutr Biochem. 42:7–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hong OY, Noh EM, Jang HY, Lee YR, Lee BK,
Jung SH, Kim JS and Youn HJ: Epigallocatechin gallate inhibits the
growth of MDA-MB-231 breast cancer cells via inactivation of the
β-catenin signaling pathway. Oncol Lett. 14:441–446. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li M, Li JJ, Gu QH, An J, Cao LM, Yang HP
and Hu CP: EGCG induces lung cancer A549 cell apoptosis by
regulating Ku70 acetylation. Oncol Rep. 35:2339–2347. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang L, Tao C, He A and He X:
Overexpression of miR-126 sensitizes osteosarcoma cells to
apoptosis induced by epigallocatechin-3-gallate. World J Surg
Oncol. 12:3832014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Deng PB, Hu CP, Xiong Z, Yang HP and Li
YY: Treatment with EGCG in NSCLC leads to decreasing interstitial
fluid pressure and hypoxia to improve chemotherapy efficacy through
rebalance of Ang-1 and Ang-2. Chin J Nat Med. 11:245–253. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yin J, Huang F, Yi Y, Yin L and Peng D:
EGCG attenuates atherosclerosis through the Jagged-1/Notch pathway.
Int J Mol Med. 37:398–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma S, Deng X, Yang Y, Zhang Q, Zhou T and
Liu Z: The lncRNA LINC00675 regulates cell proliferation,
migration, and invasion by affecting Wnt/beta-catenin signaling in
cervical cancer. Biomed Pharmacother. 108:1686–1693. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xi Z, Si J and Nan J: LncRNA MALAT1
potentiates autophagyassociated cisplatin resistance by regulating
the microRNA30b/autophagyrelated gene 5 axis in gastric cancer. Int
J Oncol. 54:239–248. 2019.PubMed/NCBI
|
|
12
|
Liang H, Zhang C, Guan H, Liu J and Cui Y:
LncRNA DANCR promotes cervical cancer progression by upregulating
ROCK1 via sponging miR-335-5p. J Cell Physiol. 2018 Oct 26;Doi:
10.1002/jcp.27484.
|
|
13
|
Kong X, Duan Y, Sang Y, Li Y, Zhang H,
Liang Y, Liu Y, Zhang N and Yang Q: LncRNA-CDC6 promotes breast
cancer progression and function as ceRNA to target CDC6 by sponging
microRNA-215. J Cell Physiol. 2018 Oct 26;Doi: 10.1002/jcp.27587.
View Article : Google Scholar
|
|
14
|
Delás MJ and Hannon GJ: lncRNAs in
development and disease: From functions to mechanisms. Open Biol.
7:1701212017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng L, Yuan X, Jiang B, Tang Z and Li GC:
LncRNAs: Key players and novel insights into cervical cancer.
Tumour Biol. 37:2779–2788. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang ZY, Yang F, Zhang YL, Liu B, Wang M,
Hong X, Yu Y, Zhou YH and Zeng H: LncRNA-ANCR down-regulation
suppresses invasion and migration of colorectal cancer cells by
regulating EZH2 expression. Cancer Biomark. 18:95–104. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao L, Han T, Li Y, Sun J, Zhang S, Liu
Y, Shan B, Zheng D and Shi J: The lncRNA SNHG5/miR-32 axis
regulates gastric cancer cell proliferation and migration by
targeting KLF4. FASEB J. 31:893–903. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu H, Zhao H, Zhang L, Xu J, Zhu C, Zhao
H and Lv G: Dandelion root extract suppressed gastric cancer cells
proliferation and migration through targeting lncRNA-CCAT1. Biomed
Pharmacother. 93:1010–1017. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun H, Wang G, Peng Y, Zeng Y, Zhu QN, Li
TL, Cai JQ, Zhou HH and Zhu YS: H19 lncRNA mediates
17β-estradiol-induced cell proliferation in MCF-7 breast cancer
cells. Oncol Rep. 33:3045–3052. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li N, Yu J, Luo A, Tang Y, Liu W, Wang S,
Liu Y, Song Y, Fang H, Chen B, et al: LncRNA and mRNA signatures
associated with neoadjuvant chemoradiotherapy downstaging effects
in rectal cancer. J Cell Biochem. 2018 Oct 15;Doi:
10.1002/jcb.27796.
|
|
21
|
Pan J, Wu X, Wang X, Huang W and Feng Q:
NEAT1 upregulates EGCG-induced CTR1 to enhance cisplatin
sensitivity in lung cancer cells. Oncotarget. 7:43337–43351.
2016.PubMed/NCBI
|
|
22
|
Wang H, Fang L, Jiang J, Kuang Y, Wang B,
Shang X, Han P, Li Y, Liu M, Zhang Z and Li P: The
cisplatin-induced lncRNA PANDAR dictates the chemoresistance of
ovarian cancer via regulating SFRS2-mediated p53 phosphorylation.
Cell Death Dis. 9:11032018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Phillips J and Eberwine JH: Antisense RNA
amplification: A linear amplification method for analyzing the mRNA
population from single living cells. Methods. 10:283–288. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gene Ontology Consortium: Gene Ontology
Consortium: Going forward. Nucleic Acids Res. 43:(Database Issue).
D1049–D1056. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Piñero J, Queralt-Rosinach N, Bravo À,
Deu-Pons J, Bauer-Mehren A, Baron M, Sanz F and Furlong LI:
DisGeNET: A discovery platform for the dynamical exploration of
human diseases and their genes. Database (Oxford) 2015. bav0282015.
View Article : Google Scholar
|
|
26
|
Kuhn M, Szklarczyk D, Franceschini A,
Campillos M, von Mering C, Jensen LJ, Beyer A and Bork P: STITCH 2:
An interaction network database for small molecules and proteins.
Nucleic Acids Res. 38:(Database Issue). D552–D556. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43:(Database Issue). D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu Q, Ren S, Lu M, Zhang Y, Zhu D, Zhang X
and Li T: Computational prediction of associations between long
non-coding RNAs and proteins. BMC Genomics. 14:6512013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wei R, Mao L, Xu P, Zheng X, Hackman RM,
Mackenzie GG and Wang Y: Suppressing glucose metabolism with
epigallocatechin-3-gallate (EGCG) reduces breast cancer cell growth
in preclinical models. Food Funct. 9:5682–5696. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Baker KM and Bauer AC: Green tea catechin,
EGCG, suppresses PCB 102-induced proliferation in
estrogen-sensitive breast cancer cells. Int J Breast Cancer 2015.
1635912015.
|
|
32
|
Wang YQ, Lu JL, Liang YR and Li QS:
Suppressive effects of EGCG on cervical cancer. Molecules.
23:E23342018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Luo KW, Wei Chen, Lung WY, Wei XY, Cheng
BH, Cai ZM and Huang WR: EGCG inhibited bladder cancer SW780 cell
proliferation and migration both in vitro and in vivo via
down-regulation of NF-κB and MMP-9. J Nutr Biochem. 41:56–64. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weng LX, Wang GH, Yao H, Yu MF and Lin J:
Epigallocatechin gallate inhibits the growth of salivary adenoid
cystic carcinoma cells via the EGFR/Erk signal transduction pathway
and the mitochondria apoptosis pathway. Neoplasma. 64:563–570.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jen J, Tang YA, Lu YH, Lin CC, Lai WW and
Wang YC: Oct4 transcriptionally regulates the expression of long
non-coding RNAs NEAT1 and MALAT1 to promote lung cancer
progression. Mol Cancer. 16:1042017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang Y, Chen WJ, Gan TQ, Zhang XL, Xie
ZC, Ye ZH, Deng Y, Wang ZF, Cai KT, Li SK, et al: Clinical
significance and effect of lncRNA HOXA11-AS in NSCLC: A study based
on bioinformatics, in vitro and in vivo verification. Sci Rep.
7:55672017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang R, Li P, Zhang G, Lu C, Wang H and
Zhao G: Long non-coding RNA XLOC_008466 functions as an oncogene in
human non-small cell lung cancer by targeting miR-874. Cell Physiol
Biochem. 42:126–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ma YC, Li C, Gao F, Xu Y, Jiang ZB, Liu JX
and Jin LY: Epigallocatechin gallate inhibits the growth of human
lung cancer by directly targeting the EGFR signaling pathway. Oncol
Rep. 31:1343–1349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Florespérez A, Marchat LA, Sánchez LL,
Romero-Zamora D, Arechaga-Ocampo E, Ramírez-Torres N, Chávez JD,
Carlos-Reyes Á, Astudillo-de la Vega H, Ruiz-García E, et al:
Differential proteomic analysis reveals that EGCG inhibits HDGF and
activates apoptosis to increase the sensitivity of non-small cells
lung cancer to chemotherapy. Proteomics Clin Appl. 10:172–182.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sadava D, Whitlock E and Kane SE: The
green tea polyphenol, epigallocatechin-3-gallate inhibits
telomerase and induces apoptosis in drug-resistant lung cancer
cells. Biochem Biophys Res Commun. 360:233–237. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shervington A, Pawar V, Menon S, Thakkar D
and Patel R: The sensitization of glioma cells to cisplatin and
tamoxifen by the use of catechin. Mol Biol Rep. 36:1181–1186. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li Y, Shen X, Wang X, Li A, Wang P, Jiang
P, Zhou J and Feng Q: EGCG regulates the cross-talk between JWA and
topoisomerase IIα in non-small-cell lung cancer (NSCLC) cells. Sci
Rep. 5:110092015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lu LY, Ou N and Lu QB: Antioxidant induces
DNA damage, cell death and mutagenicity in human lung and skin
normal cells. Sci Rep. 3:31692013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hou Y, Zhu Q, Li Z, Peng Y, Yu X, Yuan B,
Liu Y, Liu Y, Yin L, Peng Y, et al: The FOXM1-ABCC5 axis
contributes to paclitaxel resistance in nasopharyngeal carcinoma
cells. Cell Death Dis. 8:e26592017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shanmugam T, Selvaraj M and Poomalai S:
Epigallocatechin gallate potentially abrogates fluoride induced
lung oxidative stress, inflammation via Nrf2/Keap1 signaling
pathway in rats: An in-vivo and in-silico study. Int
Immunopharmacol. 39:128–139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yun M, Lee D, Park MN, Kim EO, Sohn EJ,
Kwon BM and Kim SH: Cinnamaldehyde derivative (CB-PIC) sensitizes
chemo-resistant cancer cells to drug-induced apoptosis via
suppression of MDR1 and its upstream STAT3 and AKT signalling. Cell
Physiol Biochem. 35:1821–1830. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tang G, Zhang Z, Qian H, Chen J, Wang Y,
Chen X, Chen B and Chen Y: (−)-Epigallocatechin-3-gallate inhibits
osteosarcoma cell invasiveness by inhibiting the MEK/ERK signaling
pathway in human osteosarcoma cells. J Environ Pathol Toxicol
Oncol. 34:85–93. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ciccarese C, Massari F, Blanca A, Tortora
G, Montironi R, Cheng L, Scarpelli M, Raspollini MR, Vau N, Fonseca
J and Lopez-Beltran A: Tp53 and its potential therapeutic role as a
target in bladder cancer. Expert Opin Ther Targets. 21:401–414.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nuñez NN, Manlove AH and David SS: DNMT1
and cancer: An electrifying link. Chem Biol. 22:810–811. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu L, Liu J, Dai S, Wang X, Wu S, Wang J,
Huang L, Xiao X and He D: Reduced transthyretin expression in sera
of lung cancer. Cancer Sci. 98:1617–1624. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Park JY, Kim D, Yang M, Park HY, Lee SH,
Rincon M, Kreahling J, Plass C, Smiraglia DJ, Tockman MS and Kim
SJ: Gene silencing of SLC5A8 identified by genome-wide methylation
profiling in lung cancer. Lung Cancer. 79:198–204. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tsai CH, Cheng HC, Wang YS, Lin P, Jen J,
Kuo IY, Chang YH, Liao PC, Chen RH, Yuan WC, et al: Small GTPase
Rab37 targets tissue inhibitor of metalloproteinase 1 for
exocytosis and thus suppresses tumour metastasis. Nat Commun.
5:48042014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wu CY, Tseng RC, Hsu HS, Wang YC and Hsu
MT: Frequent down-regulation of hRAB37 in metastatic tumor by
genetic and epigenetic mechanisms in lung cancer. Lung Cancer.
63:360–367. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Dötsch MM, Kloten V, Schlensog M, Heide T,
Braunschweig T, Veeck J, Petersen I, Knüchel R and Dahl E: Low
expression of ITIH5 in adenocarcinoma of the lung is associated
with unfavorable patients' outcome. Epigenetics. 10:903–912. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen Y, Cui T, Knösel T, Yang L, Zöller K
and Petersen I: IGFBP7 is a p53 target gene inactivated in human
lung cancer by DNA hypermethylation. Lung Cancer. 73:38–44. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang L, Xiao H, Zhou H, Santiago S, Lee
JM, Garon EB, Yang J, Brinkmann O, Yan X, Akin D, et al:
Development of transcriptomic biomarker signature in human saliva
to detect lung cancer. Cell Mol Life Sci. 69:3341–3350. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Mclemore TL, Adelberg S, Liu MC, McMahon
NA, Yu SJ, Hubbard WC, Czerwinski M, Wood TG, Storeng R, Lubet RA,
et al: Expression of CYP1A1 gene in patients with lung cancer:
Evidence for cigarette smoke-induced gene expression in normal lung
tissue and for altered gene regulation in primary pulmonary
carcinomas. J Natl Cancer Inst. 82:1333–1339. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Taioli E, Ford J, Trachman J, Li Y,
Demopoulos R and Garte S: Lung cancer risk and CYP1A1 genotype in
African Americans. Carcinogenesis. 19:813–817. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Choi SH, Yang H, Lee SH, Ki JH, Nam DH and
Yoo HY: TopBP1 and Claspin contribute to the radioresistance of
lung cancer brain metastases. Mol Cancer. 13:2112014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Taniguchi H, Takeuchi S, Fukuda K,
Nakagawa T, Arai S, Nanjo S, Yamada T, Yamaguchi H, Mukae H and
Yano S: AREG triggered EGFR activation confers in vivo
crizotinib-resistance of EML4-ALK lung cancer and circumvention by
EGFR inhibitors. Cancer Sci. 108:53–60. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Taron M, Rosell R, Felip E, Mendez P,
Souglakos J, Ronco MS, Queralt C, Majo J, Sanchez JM, Sanchez JJ
and Maestre J: BRCA1 mRNA expression levels as an indicator of
chemoresistance in lung cancer. Hum Mol Genet. 13:2443–2449. 2004.
View Article : Google Scholar : PubMed/NCBI
|