Introduction
Chronic liver disease, the leading cause of
mortality and morbidity globally, is characterized as fibrogenesis,
which increases the risk of liver cirrhosis and liver failure
(1). Persistent liver injury leads
to unresolved inflammation and the activation of myofibroblasts,
which express extracellular matrix (ECM) components to trigger the
generation of liver fibrosis (2).
However, observations in clinical studies and experimental animal
models suggest that the process of liver fibrosis is dynamic and
reversible, indicating that it contains at least two phases, the
developmental and recovery phases (3). Therefore, the guidance of clinical
medication may need to be upgraded according to the different
phases of this disease, which might have an influence on the
side-effects of treatments.
Leflunomide (LEF) is an anti-rheumatic drug with
immunosuppressive and anti-inflammatory functions (4). Teriflunomide, the metabolite of LEF,
induces cell cycle arrest in vigorously dividing lymphocytes, and
not in non-lymphatic cells (5).
Methotrexate (MTX) is a chemotherapy agent used for autoimmune
diseases due to its inhibitory effect on lymphocyte activation
(6). Of note, following failure of
monotherapy, LEF is used in combination with MTX, which has been
demonstrated to be effective and safe (6). However, clinical studies suggest that
this combination is associated with a significantly increased risk
of leucopenia and silent liver fibrosis in patients with rheumatoid
arthritis (RA) (7,8). Chronic liver diseases frequently
overlap with patients suffering from rheumatoid disease, which
requires the disease-modifying anti-rheumatic drugs to withstand an
excessive autoimmune response (9).
Thus, studying the multiple effects of the anti-rheumatic drugs in
the process of liver fibrosis is of importance. Additionally,
previous studies highlighted that the resolution of liver fibrosis
is mediated by myeloid cell-induced sinusoidal remodeling and the
involvement of macrophages (10,11).
Therefore, the potential side effects of the LEF+MTX combination
during the spontaneous recovery phase of liver fibrosis in mice was
investigated.
In the present study, it was demonstrated that the
LEF+MTX treatment impeded the step of hepatic fibrosis recovery.
The underlying mechanism was associated with hindering ECM
degradation and sinusoidal remodeling induced by the reduced
adhesion of myeloid cells. The present study elucidated the side
effects of the LEF+MTX combination in hepatic fibrosis recovery and
provided evidence for its use in clinical therapy.
Materials and methods
Animal experiment
A total of 50, 6-week old C57BL/6 mice weighing ~20
g were obtained from the Shanghai Model Organisms Center, Inc.
(Shanghai, China). Mice were divided into 4 groups, 6 mice per
group, for 2 individual experiments. All animal experiments were
approved by the Committee on Laboratory Animal Care of Fujian
Medical University (Fuzhou, China) and performed according to the
‘ARRIV’ guidelines (12). Female
mice aged 8 weeks were intraperitoneally injected with 200 µl 25%
carbon tetrachloride (CCl4)/olive oil (2 ml/kg) three
times per week for 12 weeks to induce hepatic fibrosis. A group of
mice served as the control, termed the NC group. The degree of
liver fibrosis in the mice was evaluated by histology.
Subsequently, the mice were randomly divided into two groups: A
group received LEF (15 mg/kg, oral administration) and MTX
(intraperitoneal administration of 0.15 mg/Kg,) every day for four
weeks and a group that received the relevant vehicle of LEF and MTX
via the same routes. Leflunomide (solvent, 0.5% sodium
carboxymethylcellulose) and methotrexate (solvent, PBS) were
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Hematoxylin-eosin staining
Whole liver tissues were fixed in 4%
paraformaldehyde at room temperature overnight and embedded in
paraffin. Sections (5 µm) of paraffin-embedded tissues were
subjected to hematoxylin staining for 8 min at room temperature.
Following washing in water for 1 h and dehydration in ethyl alcohol
for 10 min, the sections were subjected to eosin staining for 3 min
at room temperature. Following dehydration in ethyl alcohol, the
sections were mounted to detect pathological alterations. The
images (magnification, ×200) were obtained by an Axio Vert.A1
inverted microscope (Zeiss AG, Oberkochen, Germany).
Masson staining
Masson staining was performed with a Masson-Goldner
staining kit (Merck KGaA), following the manufacturer's protocol,
to detect collagen deposition in liver tissues. Briefly, rehydrated
sections (5 µm) were staining with Weigert's iron hematoxylin
staining solution for 5 min, followed by staining with Azophloxine
solution for 10 min, Tungstophosphoric acid orange G solution for 1
min and Light green SF solution for 2 min. Washes with 1% acetic
acid were performed between the staining steps. Following
dehydration in ethyl alcohol, the sections were mounted. All the
procedures were performed on room temperature. The images
(magnification, ×200) were obtained by an Axio Vert.A1 inverted
microscope (Zeiss AG, Oberkochen, Germany).
Immunofluorescence
Liver tissues were fixed in 4% paraformaldehyde and
cut using a cryostat. Sections (10 µm) were permeabilized with PBS
containing 0.5% Triton X-100. Following blocking with PBS/5% bovine
serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) for
2 h at room temperature, the sections were incubated with
Alexa-Fluor488-conjugated rat anti-mouse cluster of differentiation
(CD)11b antibody (1:200; cat. no. 101219; clone M1/70; BioLegend,
Inc., San Diego, CA, USA) at 37°C for 1 h in the dark. The sections
were counterstained with DAPI (BioLegend, Inc.) for 15 min at room
temperature. The images (magnification, ×200) were visualized by a
Zeiss LSM700 confocal microscope (Zeiss AG).
ELISA
Serum (~5 µl) contents were detected by procollagen
type III (cat. no. CSB-E17125m), collagen type IV (cat. no.
CSB-E08884m), hyaluronic acid (cat. no. CSB-E08121m) and laminin
(cat. no. CSB-EL012725MO) ELISA kits (all Cusabio, Wuhan, China),
according to the manufacturer's protocols. To detect the expression
levels of liver vascular endothelial growth factor (VEGF) protein,
liver tissues were homogenized in radioimmunoprecipitation (RIPA;
Cusabio Technology LLC, Wuhan, China) buffer on ice, and the
supernatants of the tissue homogenate were measured using a VEGF
ELISA kit (cat. no. CSB-E04756m; Cusabio, Wuhan, China).
Flow cytometry
Whole blood samples (100 µl) were incubated with red
blood cell lysis buffer (BD Pharmingen; BD Biosciences, San Jose,
CA, USA). Following centrifugation at 500 × g for 15 min at 4°C,
the cells were washed, stained with purified anti-CD16/32 (1/200;
clone 2.4G2; cat. no. 553141; BD Pharmingen; BD Biosciences), and
subsequently stained with fluorescein isothiocyanate-conjugated
CD11b (1:400; clone M1/70; cat. no. 557396; BD Pharmingen; BD
Biosciences) and allophycocyanin-conjugated Ly6C (1:20; clone
HK1.4; cat. no. 560595; BD Pharmingen; BD Biosciences) antibodies
for 30 min at 4°C. The cells were washed, resuspended in 200 µl
FACS buffer (BD Pharmingen; BD Biosciences) and analyzed with a
FACSAria (BD Biosciences). FlowJo software (version 10; FlowJo,
LLC, Ashland, OR, USA) was used for analysis. The number (N) cells
per ml in blood was calculated by the formula: N=collected cells ×
200/loading volume × 10.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) in liver tissues
Sections of liver tissues (4 mm) were rapidly frozen
in liquid nitrogen and homogenized in 1 ml TRIzol®
reagent (Thermo Fisher Scientific, Inc.). The RNA was exacted
following the manufacturer's protocol, and cDNA was synthesized
using SuperScriptIIreverse transcriptase (Invitrogen; Thermo Fisher
Scientific, Inc.) and oligodT primers (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
primers were as follows: VEGF-A forward, 5′-ATCCGCATGATCTGCATGG-3′
and reverse, 5′-ATCCGCATGATCTGCATGG-3′; beta-actin forward,
5′-CCACCATGTACCCAGGCATT-3′ and reverse, 5′-CGGACTCATCGTACTCCTGC-3′.
RT-qPCR was performed with cDNA in 15 µl reactions using SYBR-Green
Master Mix (Promega Corporation, Madison, WI, USA). The
thermocycling conditions were: 94 °C for 10 min, 94°C for 30 sec,
60°C for 30 sec and 72°C for 30 sec, repeated for 35 cycles. The
mRNA levels were first calculated using the 2−ΔΔCq
method and then normalized to β-actin level (13).
Statistical analysis
All graphs were manufactured using GraphPad Prism
version 7.0 software (GraphPad Software, Inc., La Jolla, CA, USA).
Statistical analysis was analyzed by one-way analysis of variance
followed by Tukey's multiple comparisons test. Data are expressed
as the mean ± standard deviation and collected from at least 6
mice. P<0.05 was considered to indicate a statistically
significant difference.
Results
Combination LEF+MTX impedes the
recovery of liver pathology in the liver fibrosis in mice
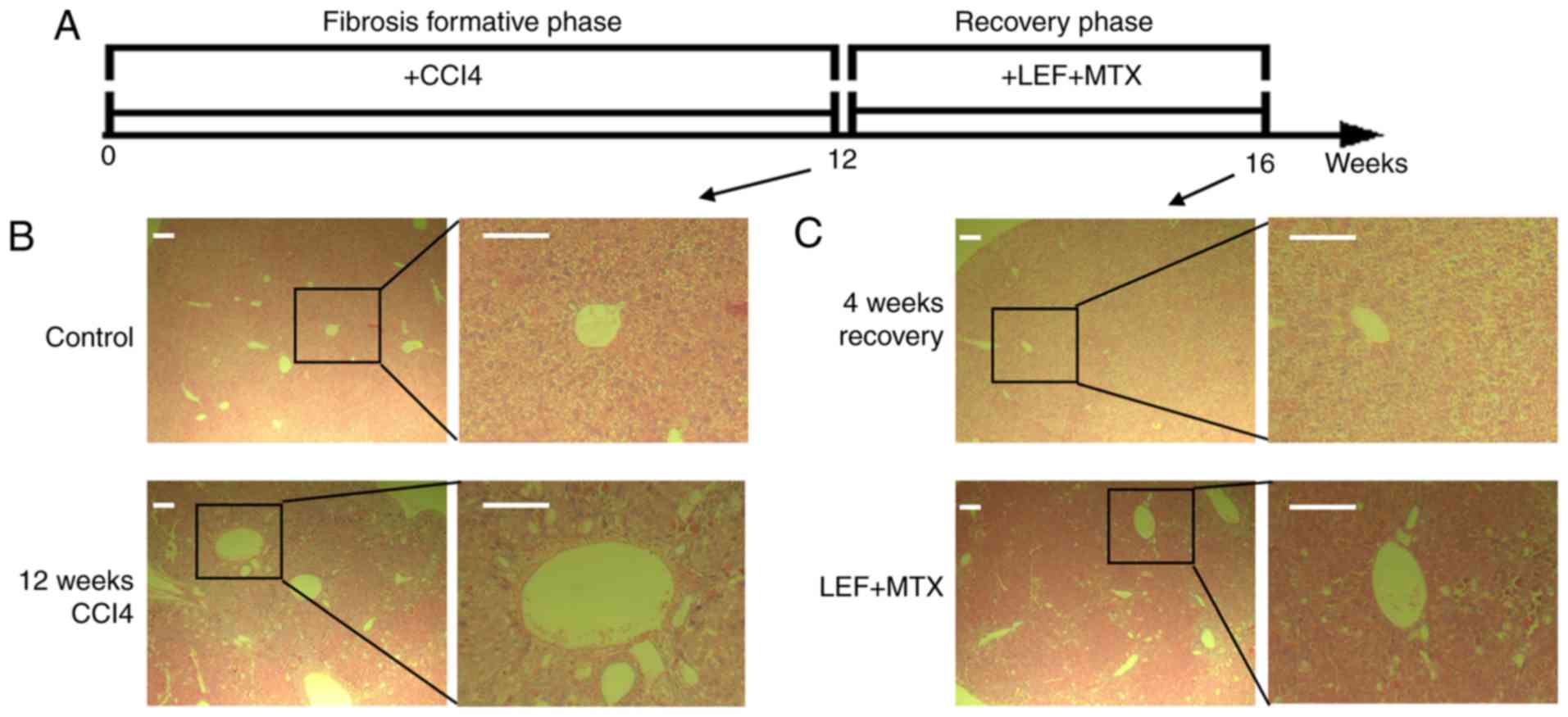
The animal experiments were performed as illustrated
in Fig. 1A. After 12 weeks of
CCl4 administration, five mice were randomly selected
for the determination of liver pathology, and illustrating a marked
alteration in the liver histology (Fig. 1B). Subsequently, the mice with
liver fibrosis were divided into two groups with or without
treatment with the LEF+MTX combination, and underwent recovery for
4 weeks. A completely spontaneous recovery was found in the mice
without further intervention (Fig.
1C, upper panel). By contrast, the mice that received the
LEF+MTX combination exhibited severe liver pathology compared with
the control group (Fig. 1C, lower
panel). These results demonstrated that the LEF+MTX combination had
an inhibitory effect on the recovery of liver fibrosis.
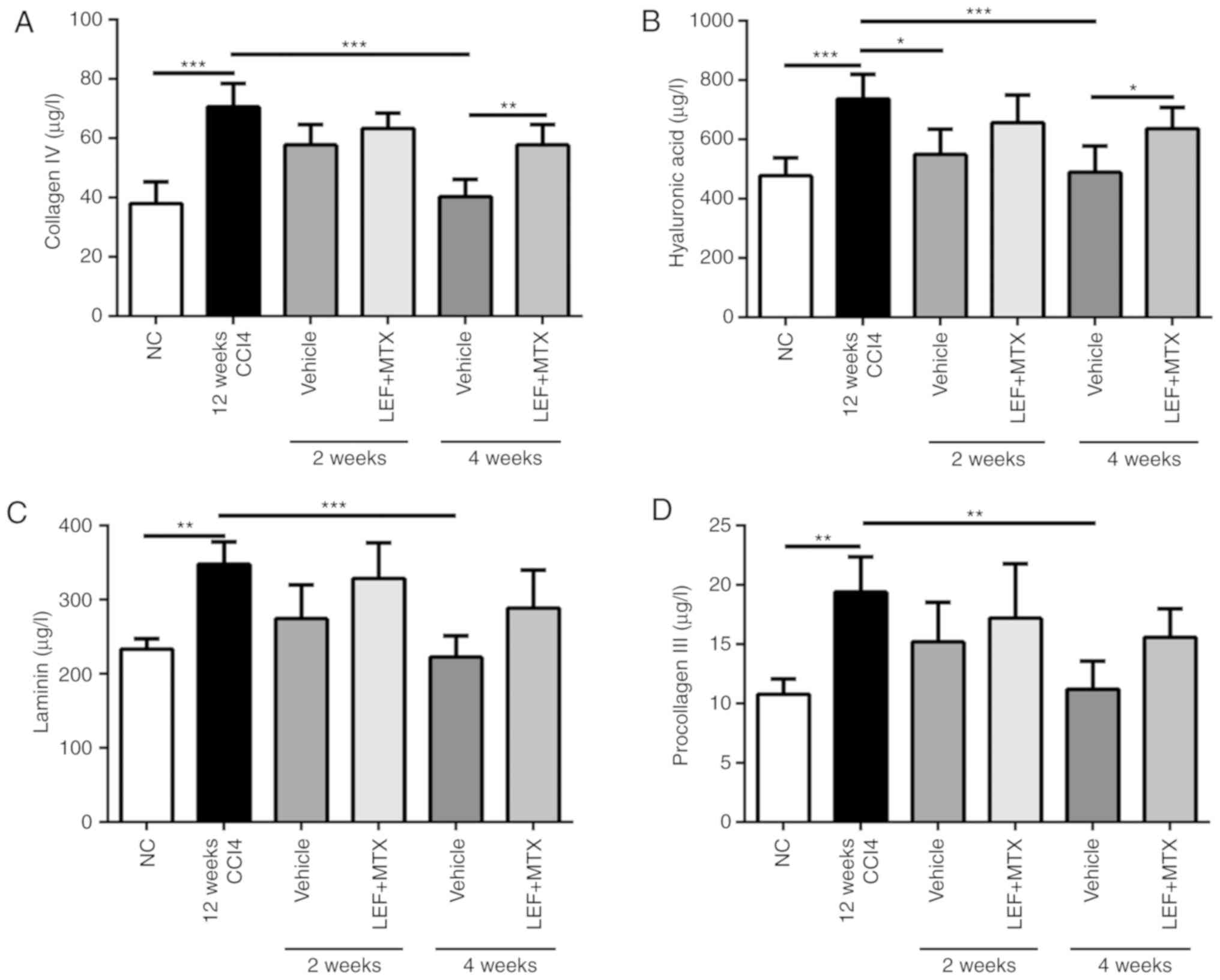
A high level of collagen degradation
is observed in mice receiving LEF+MTX treatment
Liver fibrosis levels are accompanied by increasing
levels of serum indicators, including hyaluronic acid, laminin,
procollagen type III and collagen IV (14). The mice which underwent 12 weeks of
CCl4 administration demonstrated a marked increase in
serum parameters. Following grouping, the mice without LEF+MTX
treatment exhibited a continuous decline of these indicators, which
almost returned to their normal levels after 4 weeks (Fig. 2). By contrast, no significant
decrease of these indicators was observed in the mice that received
the LEF+MTX combination. In addition, compared with mice without
LEF+MTX treatment, mice that were administrated with LEF+MTX
combination showed the higher levels of collagen IV and hyaluronic
acid.
Reduced monocytes in blood are
associated with the impaired resolution of fibrosis
It has been demonstrated that combination LEF+MTX
has a potent effect on the inhibition of leucopoiesis (7). The absolute numbers and percentages
of monocytes in blood from the mice that experienced recovery at 2
weeks were measured. The gating strategy used to identify the
inflammatory monocytes (iMo, CD11b+Ly6Chigh)
and the patrolling monocytes (pMo, CD11b+Ly6Cint) is
illustrated in Fig. 3A. Treating
mice with CCl4 for 12 weeks significantly increased the
number of monocytes in the blood. After 2 weeks of recovery, iMo
and pMo were decreased in the mice, whether or not they were
treated with LEF+MTX (Fig. 3B-E).
Compared with the vehicle group, either the absolute numbers or
cell percentages of iMo and pMo were decreased in the
LEF+MTX-treated mice. In summary, the LEF+MTX combination
significantly reduced the levels of two monocyte subsets in the
mice that underwent fibrosis recovery, and this may cause the
impaired recruitment of monocytes to the site of liver fibrosis,
although this requires further investigation.
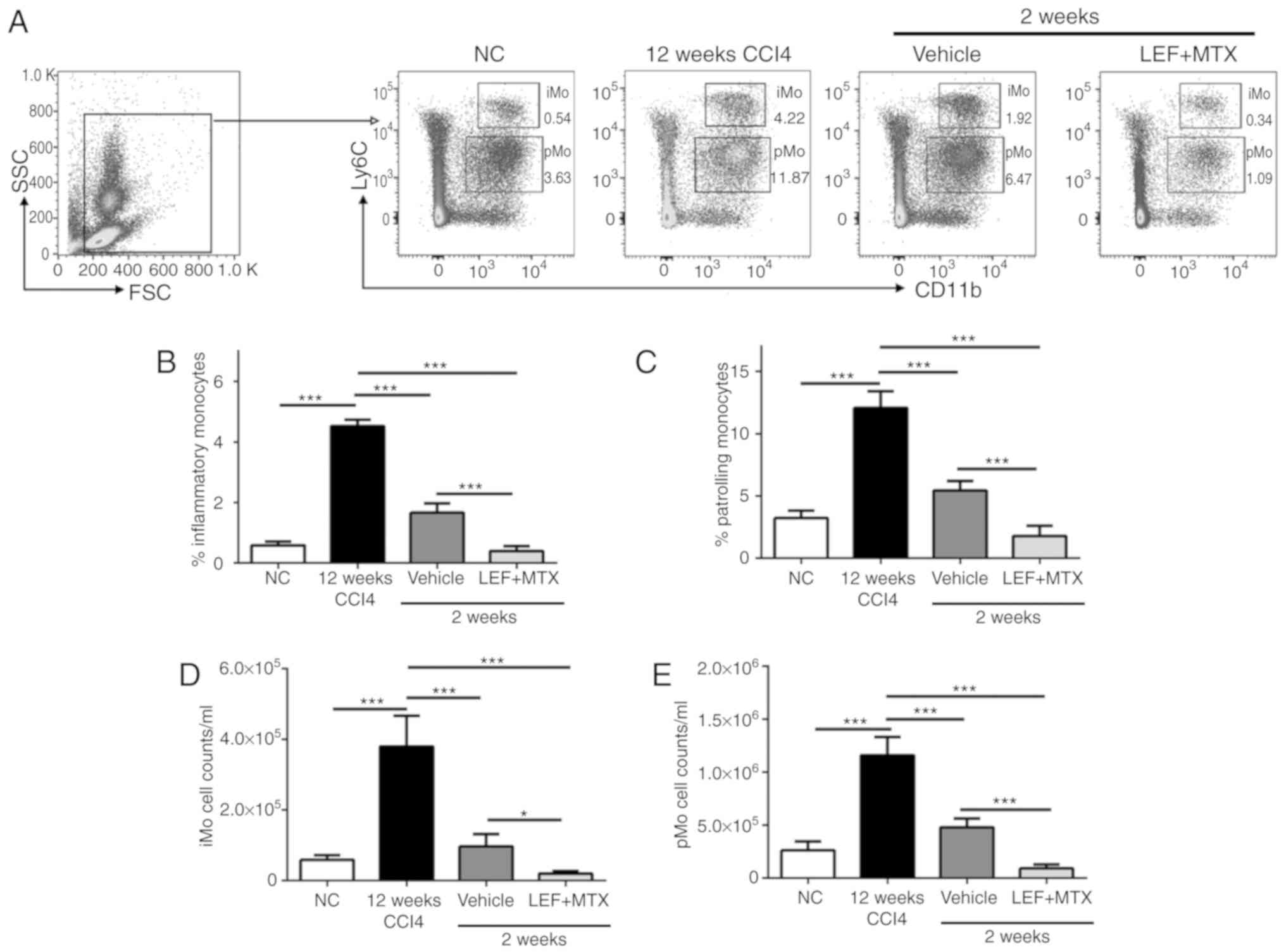 | Figure 3.Monocyte subsets are significantly
decreased following treatment with LEF+MTX. Whole blood (200 µl)
was collected, and the red blood cells were removed. (A) The gating
strategy and representative images of flow cytometry for
CD11b+Ly6C+ cells. The percentages of (B) iMo
(CD11b+Ly6Chigh) and of (C) pMo
(CD11b+Ly6Cint) were measured by flow cytometry. The
absolute numbers of (D) iMo and (E) pMo are also presented. Data
are presented as the mean ± standard deviation (n=6). *P<0.05
and ***P<0.001. CCl4, carbon tetrachloride; CD,
cluster of differentiation; iMo, inflammatory monocytes; LEF,
leflunomide; MTX, methotrexate; NC, normal control; pMo, patrolling
monocytes; FSC, forward scatter; SSC, side scatter. |
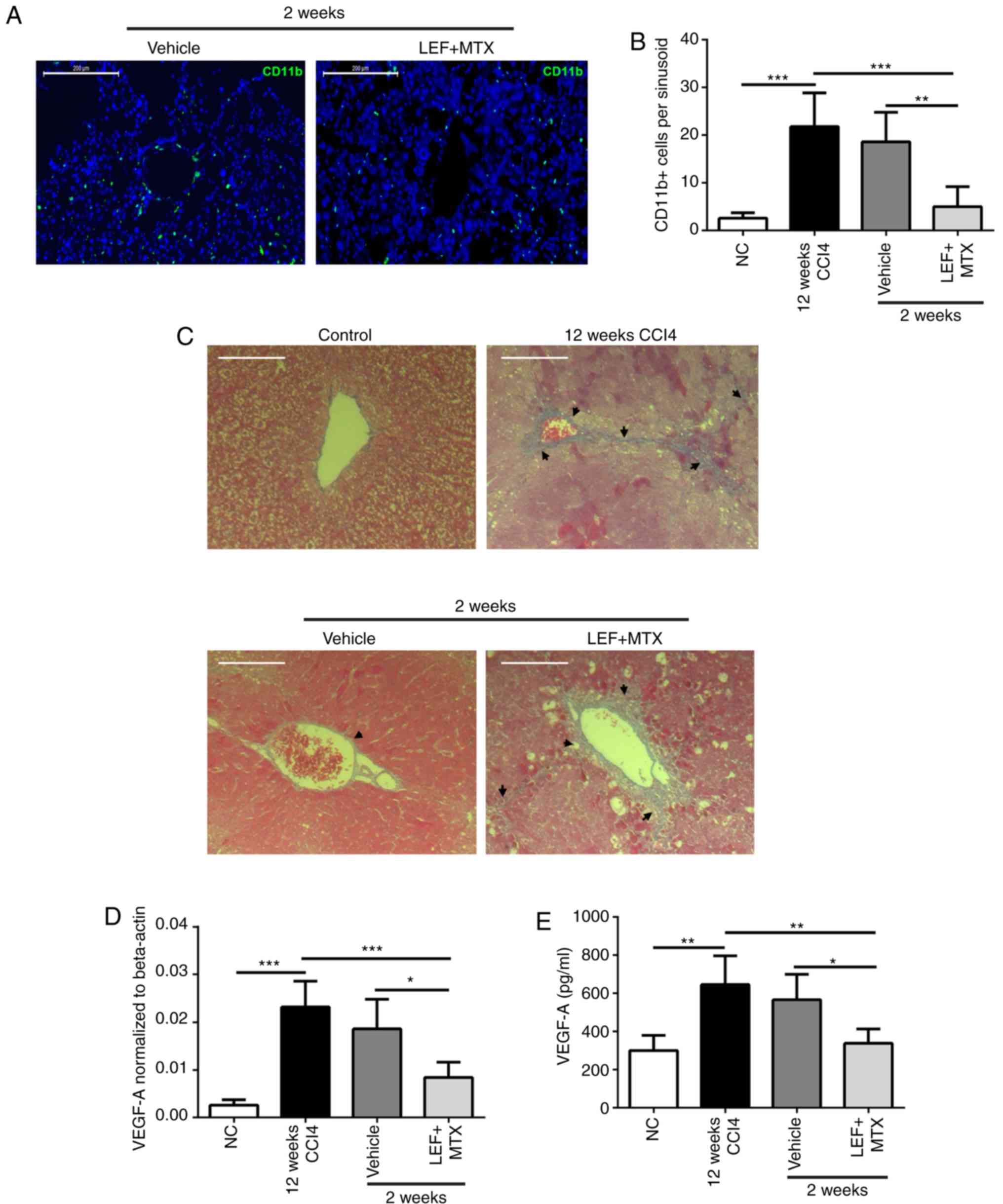
Combination LEF+MTX suppresses
fibrosis resolution mediated by myeloid cells during the recovery
phase
To reveal the underlying association between myeloid
cells in the blood and fibrosis resolution, CD11b
immunohistochemistry and Masson's trichrome staining were performed
on the liver tissues. The results demonstrated that during the
recovery phase, abundant CD11b+ cells were adhered to
the sinusoidal endothelium, while the LEF+MTX treatment
significantly reduced the number of CD11b+ cells per
sinusoid (Fig. 4A and B). Notably,
the results of the Masson's trichrome staining suggested a marked
degradation of the ECM in liver sinusoids from mice that
experienced the spontaneous recovery, although not in the mice
treated with the LEF+MTX combination (Fig. 4C). Subsequently, the expression
levels of VEGF-A, a cytokine involved in angiogenesis which is also
present in this phase, were investigated (10). The mRNA and protein expression
levels of VEGF-A demonstrated that compared with the mice that had
undergone recovery at 2 weeks, the VEGF-A expression levels in the
LEF+MTX-treated mice were decreased (Fig. 4D and E). These results indicated
that the LEF+MTX combination may suppress the presence of myeloid
cells in the liver sinusoidal endothelium, impeding the fibrosis
recovery.
Discussion
Hepatic fibrosis, a pathological consequence of
chronic liver diseases with differing etiologies, is characterized
by the deposition of ECM produced by activated hepatic stellate
cells in response to persistent inflammation and unremitting injury
(2). A previous clinical study
demonstrated that treatment with combined LEF and MTX in patients
with RA led to an increased risk of silent liver fibrosis (8). In the present study, part of the
underlying mechanism of this was assessed in experimental
animals.
Previous studies have proposed that LEF exerts an
inhibitory effect on the formation of hepatic fibrosis,
accelerating the fibrosis recovery that involves the enhanced
apoptosis of hepatic stellate cells (15,16).
Additionally, MTX is associated with the increased morbidity of
hepatic fibrosis and increased myelosuppression, which is
characterized by fewer white blood cells in the blood, particularly
myeloid cell subsets (17,18). In the recovery phase of liver
fibrosis, it was observed that the LEF+MTX combination
significantly impeded the resolution of the fibrotic scar, and the
underlying mechanism may be associated with impaired myeloid
cell-induced remodeling of the liver sinusoids. Inflammatory
monocytes selectively respond to infection and tissue damage, and
are also able to digest damaged tissue (19,20).
pMo, with attenuated inflammatory properties, exhibit a fundamental
characteristic of patrolling the endothelium and, notably, they are
the principal producers of VEGF-A and drive healing via the
accumulation of myofibroblasts, angiogenesis and the deposition of
ECM (20). These characteristics
of monocyte subsets support the observation that the LEF+MTX
combination inhibited the number of monocytes in circulation,
leading to fewer monocytes reaching the site of liver fibrosis
recovery. A previous study on the recovery of liver fibrosis
suggested that myeloid cell-driven VEGF is indispensable for the
degradation of the ECM in liver sinusoids, and that this process is
dependent on the regulation of sinusoidal cells (10). This notion is consistent with the
present data, in that decreased expression of liver VEGF-A was
associated with impaired ECM degradation. In addition, a previous
in vivo study suggested that LEF administration increases
hepatic exposure to MTX, which may lead to additional liver injury
(21). This observation may be
another reason why the LEF+MTX administration impeded fibrosis
recovery.
In the present study, it was demonstrated that
during the recovery phase of CCl4-induced liver
fibrosis, the LEF+MTX treatment impeded fibrosis resolution by
reducing the number of myeloid cells, which may promote the
revascularization of sinusoidal endothelial cells, leading to
reduced ECM degradation. The present study highlighted a potential
side effect of the LEF+MTX combination in the context of recovery
from hepatic fibrosis-associated diseases.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ML, RG and ZS performed the animal experiments and
analyzed data. ML, SK and DZ contributed to the conception and
design of this study. ML drafted this manuscript. All authors have
reviewed this manuscript and approved for publication.
Ethics approval and consent to
participate
All animal experiments were approved by the
Committee on Laboratory Animal Care of Fujian Medical University
(Fuzhou, China) and performed according to the ‘ARRIV’
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests
References
|
1
|
Patten DA and Shetty S: Chronic liver
disease: Scavenger hunt for novel therapies. Lancet. 391:104–105.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tsuchida T and Friedman SL: Mechanisms of
hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol.
14:397–411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pellicoro A, Ramachandran P, Iredale JP
and Fallowfield JA: Liver fibrosis and repair: Immune regulation of
wound healing in a solid organ. Nat Rev Immunol. 14:181–194. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Janssen NM and Genta MS: The effects of
immunosuppressive and anti-inflammatory medications on fertility,
pregnancy, and lactation. Arch Intern Med. 160:610–619. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dougados M, Emery P, Lemmel EM, Zerbini
CA, Brin S and van Riel P: When a DMARD fails, should patients
switch to sulfasalazine or add sulfasalazine to continuing
leflunomide? Ann Rheum Dis. 64:44–51. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wessels JA, Huizinga TW and Guchelaar HJ:
Recent insights in the pharmacological actions of methotrexate in
the treatment of rheumatoid arthritis. Rheumatology (Oxford).
47:249–255. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rozman B: Clinical pharmacokinetics of
leflunomide. Clin Pharmacokinet. 41:421–430. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee SW, Park HJ, Kim BK, Han KH, Lee SK,
Kim SU and Park YB: Leflunomide increases the risk of silent liver
fibrosis in patients with rheumatoid arthritis receiving
methotrexate. Arthritis Res Ther. 14:R2322012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ferri C, Sebastiani M, Antonelli A, Colaci
M, Manfredi A and Giuggioli D: Current treatment of hepatitis
C-associated rheumatic diseases. Arthritis Res Ther. 14:2152012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kantari-Mimoun C, Castells M, Klose R,
Meinecke AK, Lemberger UJ, Rautou PE, Pinot-Roussel H, Badoual C,
Schrödter K, Österreicher CH, et al: Resolution of liver fibrosis
requires myeloid cell-driven sinusoidal angiogenesis. Hepatology.
61:2042–2055. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Duffield JS, Forbes SJ, Constandinou CM,
Clay S, Partolina M, Vuthoori S, Wu S, Lang R and Iredale JP:
Selective depletion of macrophages reveals distinct, opposing roles
during liver injury and repair. J Clin Invest. 115:56–65. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kilkenny C, Browne W, Cuthill IC, Emerson
M and Altman DG; National Centre for the Replacement, Refinement
and Reduction of Amimals in Research, : Animal research: Reporting
in vivo experiments-the ARRIVE guidelines. J Cereb Blood Flow
Metab. 31:991–993. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fontana RJ, Dienstag JL, Bonkovsky HL,
Sterling RK, Naishadham D, Goodman ZD, Lok AS, Wright EC and Su GL;
HALT-C Trial Group, : Serum fibrosis markers are associated with
liver disease progression in non-responder patients with chronic
hepatitis C. Gut. 59:1401–1409. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yao HW, Li J, Chen JQ and Xu SY:
Inhibitory effect of leflunomide on hepatic fibrosis induced by
CCl4 in rats. Acta Pharmacol Sin. 25:915–920. 2004.PubMed/NCBI
|
|
16
|
Tang X, Yang J and Li J: Accelerative
effect of leflunomide on recovery from hepatic fibrosis involves
TRAIL-mediated hepatic stellate cell apoptosis. Life Sci.
84:552–557. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bjorkman DJ, Boschert M, Tolman KG, Clegg
DO and Ward JR: The effect of long-term methotrexate therapy on
hepatic fibrosis in rheumatoid arthritis. Arthritis Rheum.
36:1697–1701. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bilasy SE, Essawy SS, Mandour MF, Ali EA
and Zaitone SA: Myelosuppressive and hepatotoxic potential of
leflunomide and methotrexate combination in a rat model of
rheumatoid arthritis. Pharmacol Rep. 67:102–114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Landsman L, Varol C and Jung S: Distinct
differentiation potential of blood monocyte subsets in the lung. J
Immunol. 178:2000–2007. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nahrendorf M, Swirski FK, Aikawa E,
Stangenberg L, Wurdinger T, Figueiredo JL, Libby P, Weissleder R
and Pittet MJ: The healing myocardium sequentially mobilizes two
monocyte subsets with divergent and complementary functions. J Exp
Med. 204:3037–3047. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang L, Ma L, Lin Y, Liu X, Xiao L, Zhang
Y, Xu Y, Zhou H and Pan G: Leflunomide increases hepatic exposure
to methotrexate and its metabolite by differentially regulating
multidrug resistance-associated protein Mrp2/3/4 transporters via
peroxisome proliferator-activated receptor α activation. Mol
Pharmacol. 93:563–574. 2018. View Article : Google Scholar : PubMed/NCBI
|