Introduction
Acute aortic dissection (AAD) is a common and
critical clinical disease with a high rate of mortality; however,
its pathogenetic mechanism requires further investigation. Recent
studies suggested that AAD arises from problems with vascular
function. It is well established that vascular smooth muscle cells
(VSMCs) are essential regulators of vascular function (1). VSMCs are located in the middle
vascular layer of healthy arteries (2–4),
where they secrete vasoconstrictor proteins that help regulate
blood vessel tension and blood flow (5). VSMCs are the most important component
of the vascular middle layer and may contribute to AAD. VSMCs are
divided into two cell phenotypes: Contractile and synthetic
(6,7). Fusiform contractile VSMCs demonstrate
difficulty in secreting extracellular matrix proteins, and exhibit
poor proliferation and migration (8). The proliferative and migration
abilities of synthetic VSMCs are enhanced compared with the
systolic VSMCs (9–11). Synthetic VSMCs secrete a variety of
extracellular matrix components, including collagen, elastin and
proteoglycans (12,13). Upon atherosclerosis and arterial
restenosis, VSMCs undergo a transformation from the contractile to
the synthetic phenotype (14).
This transformation promotes the migration of SMCs into the intima,
enhances proliferation and promotes the secretion of extracellular
proteins (15). These phenotypic
transformations may underlie the basis of regulating the
composition and stability of blood vessels, which may eventually
lead to the formation of vascular lesions (16). Recent studies revealed that
extracellular factors and downstream signaling pathways participate
in transformation of VSMCs (17,18),
such as autophagy.
It was demonstrated that Rab7 participates in the
regulation of VSMC proliferation and migration (2). Autophagy, induced by platelet-derived
growth factor, serves an important role in the process of
transforming the phenotype of VSMCs from a contractile to a
synthetic form by preventing oxidative stress-induced cell death
(19). Rab proteins are
Ras-related small GTPases, which regulate exo- and endocytic
membrane trafficking by vesicle docking and fusion (20). As an important member of the Rab
GTPase superfamily, Rab7 promotes lysosomal biosynthesis and
maintains lysosomal function (21,22).
Furthermore, Rab7 serves a pivotal role in the fusion of vesicles
and lysosomes, and exhibit an important effect on autophagosome
maturation (23). Abnormal
expression or alterations in the activity of Rab7 may be associated
with cardiovascular diseases, lipid storage disorders and
neurodegenerative diseases (24–27).
Therefore, the present study hypothesized that phenotypic
transformation regulated by Rab7-mediated autophagy may be
associated with VSMC proliferation and invasion.
In the present study, AD VSMCs were treated with
small interfering (si)RNA or Rab7 overexpression plasmid to assess
phenotypic transformations and cellular behavior, including
proliferation, invasion, cell cycle and autophagy. The present
study aimed to identify the effects of Rab7 on autophagy in VSMCs
and determine whether -Rab7-regulated autophagy results in the
alteration of VSMC phenotype and cell behavior.
Materials and methods
Tissue sampling
A total of 51 AAD tissues were obtained from
patients with type A AD, who underwent aortic replacement
operations at The First Affiliated Hospital of Nanjing Medical
University (Nanjing, China) between October 2015 and October 2017
(Ethics no. 2016-SR-144). The main clinicopathological
characteristics of the patients are shown in Table I. All experimental protocols were
approved by the Ethics Committee of The First Affiliated Hospital
of Nanjing Medical University and all patients provided written
informed consent. Patients with hereditary connective tissue
defects, including Marfan syndrome or traumatic aneurysms, were
excluded. The control group comprised 14 non-aortic dissection
(NAD) aortic specimens collected from organ donors, containing
residual tissues from organ pruning, such as the aortic tissue
trimmed during heart transplantations. No significant differences
between the groups were observed with respect to the clinical
characteristics, including patient age, sex, smoking status,
hypertension or diabetes.
 | Table I.Association between Rab7 protein
expression and clinicopathological characteristics in patients with
AD. |
Table I.
Association between Rab7 protein
expression and clinicopathological characteristics in patients with
AD.
| Group | Sex
(female/male) | Age (years) | Blood glucose
(mmol/l) | Systolic pressure
(mmHg) | Diastolic pressure
(mmHg) |
|---|
| AD | 7/44 | 47.82±6.77 | 4.91±0.48 | 155.12±16.20 | 84.67±11.57 |
| NAD | 2/12 | 45.50±5.56 | 4.75±0.56 |
125.71±7.53b |
76.29±5.64c |
| Relative Rab7
expression | /a | / | / | 0.256 | 0.231 |
Immunohistochemistry
The fresh tissues were fixed in 4% paraformaldehyde
at room temperature for 48 h dehydrated in a graded ethanol series
and embedded in paraffin. Then, 5 µm thick sections were dewaxed
twice in xylene for 10 min and rehydrated in ethanol (75% ethanol 2
h, 95% ethanol 1.5 h, 95% ethanol 1.0 h, absolute ethyl alcohol 1.5
h and absolute ethyl alcohol 1.0 h) at room temperature. The
sections were washed with PBS (pH 7.4) 3 times for 3 min. Then, the
slides were boiled in 10 mM/l citrate buffer (pH 6.0) in an
autoclave for 3 min for antigen retrieval. Following slow cooling,
the sections were blocked by immersion in 3% methanolic peroxide
for 15 min at room temperature and incubated with the following
primary antibodies: Antibodies against Rab7 (ab50533; 1:500; Abcam,
Cambridge, UK) overnight at 4°C, followed by horseradish peroxidase
(HRP)-conjugated secondary antibodies for 2 h at room temperature
(31430; 1:5,000; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
After washing with water, the slides were counterstained with
hematoxylin for 2 min at room temperature, dehydrated and mounted
in resin mountant.
Cell culture
Primary human aortic VSMCs (HASMCs) were isolated
from the collected tissues using the tissue adhesion method
(28); subsequent experiments were
performed with 2nd-4th generation cells. Cells were cultured in
smooth muscle cell medium (Sciencell Research Laboratories, Inc.,
San Diego, CA, USA) containing 10% fetal bovine serum (Sciencell
Research Laboratories, Inc.). Cells were grown in cell culture
flasks (Corning, Inc., Corning, NY, USA) in a humidified
environment in 37°C with 5% CO2.
Cell transfection
siRab7, and Rab7 overexpression and control vectors
were obtained from Shanghai GeneChem Co., Inc. (Shanghai, China).
Sequences were: siRab7#1: 5′GUCUAGUUCCCUUCUGUGU(dTdT)3′; siRab7#2:
5′ACACAGAAGGGAACUAGAC(dTdT)3′. A total of 20 nM of siRNAs were
transfected into the cells using RNAiMAX (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. All
siRNAs were purchased from Shanghai GeneChem Co., Inc. (Shanghai,
China). Transfection efficiency was determined using western blot
analysis.
5′Ethynyl-2′-deoxyuridine (EdU)
assay
Briefly, the cells were plated into 12-well plates
at a density of 8×103 cells/well with 500 µl culture
medium. Cells were fixed at room temperature for 20 min using 4%
paraformaldehyde (PFA), then permeabilized with 0.2% Triton X-100
at room temperature for 20 min and blocked with 10% goat serum
(Abcam) in TBS containing 0.05% Tween-20 (TBST) for 1 h at room
temperature. The EdU assay was performed using the Click-iT Plus
EdU Alexa Fluor® 488 Imaging kit (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocols
to determine cell proliferation. Nuclei were counterstained with
DAPI at room temperature for 10 min. Under a confocal microscope
(Zeiss AG, Oberkochen, Germany), 15 fields/sample (magnification,
×200) were observed and the number of EdU-positive nuclei was
determined.
Transwell invasion assay
Transwell invasion assays were performed using
Transwell chambers (pores, 8 mm; Corning, Inc.). Serum-free medium
containing 1×104 cells was added to the upper chamber
pre-coated with Matrigel (BD Biosciences, San Jose, CA, USA).
Following 24 h incubation in 37°C with 5% CO2 cells that
migrated to the lower surface of the membrane were fixed with
absolute ethanol and stained with 0.1% crystal violet at room
temperature for 10 min. Cells in the upper chamber were wiped from
the Matrigel with a cotton swab. Then, 3 visual fields were
randomly selected to screen the cells located on the lower surface
of the membrane under an inverted microscope. After calculating the
number of cells in every field, the average was counted and
analyzed.
Cell cycle analysis
Flow cytometry was performed to detect the cell
cycle. Cells were cultured to 80–90% confluence in 6-well plates.
Cells were transfected for 48 h, digested with 0.25% trypsin at
room temperature for 10 min, collected with pre-chilled 70% ethanol
at 4°C and maintained at 4°C for overnight fixation. Cells were
collected by centrifugation at 150 × g for 10 min at 4°C; the upper
layer was discarded and ethanol was washed away with PBS. The cells
were then stained with PI DNA staining solution (Hangzhou Lianke
Biology Technology, Co., Ltd., Hangzhou, China) at room temperature
for 30 min. A Beckman Gallios (AU39633; Beckman Coulter, Inc.,
Brea, CA, USA) and Wincycle version 3.0 (Beckman Coulter, Inc.)
were used for cell cycle analysis.
Detection of autophagy
Immunofluorescence and western blot analysis were
used to determine autophagy. For immunofluorescence assays, HASMCs
were cultured to 80–90% confluence in 6-well plates.
The cells were fixed in 4% PFA at room temperature
for 20 min, followed by 0.2% Triton X-100 for 20 min and
permeabilization with 10% goat serum in TBST for 1 h at room
temperature. Cells were incubated with rabbit polyclonal
anti-microtubule-associated protein light chain 3 (LC3) antibody
(ab48394; 1:100; Abcam, Cambridge, UK) at 4°C overnight. Cells were
then incubated with Alexa Fluor 488-labeled secondary antibody (5
µg/ml; A-11034; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature and imaged using an inverted fluorescence microscope.
The expression of Beclin-1 (ab62557; 1:1,000; Abcam), LC3 (ab48394;
1:1,000; Abcam) and P62 (ab56416; 1:1,000; Abcam) were detected by
western blotting (described below). Following the administration of
3-methyladenine (3-MA; 5 mM), a pharmacological inhibitor of
autophagy, to Rab7-overexpressing cells at 37°C with 5%
CO2 for 48 h, α-SMA (ab5694; 1:1,000; Abcam) and
Osteopintin (ab8448; 1:1,000; Abcam) were detected by western
blotting (as described below).
Protein extraction and western
blotting
Following washes with PBS for 3 times, proteins were
extracted from the cells using radioimmunoprecipitation assay lysis
buffer containing phenylmethylsulfonyl fluoride and protease
inhibitors (R0010; Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China). A Bicinchoninic Acid Protein Assay kit
(Beyotime Institute of Biotechnology, Shanghai, China) was used to
quantify the total protein. SDS-PAGE (12%) was used to separate the
proteins (30 µg/lane) prior to transfer to polyvinylidene
difluoride membranes (0.45 µm; EMD Millipore, Billerica, MA, USA).
Subsequently, membranes were blocked with TBST and 5% fat-free dry
milk for 2 h at room temperature. Membranes were incubated with
primary antibodies (Beclin-1; ab62557; 1:1,000, LC3; ab48394;
1:1,000 and P62; 1:1,000; ab56416; all from Abcam) in TBST
containing 1% bovine serum albumin (ScienCell Research
Laboratories, Inc., San Diego, CA, USA) overnight at 4°C, followed
by incubation with horseradish peroxidase-conjugated secondary
antibody at an appropriate dilution with TBST (1:3,000; ab97080 and
ab97040; Abcam, UK) at room temperature for 2 h. Membranes were
washed three times for 10 min washes with TBST. A ChemiDoc XRS
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) and Supersignal
West Femto Trial kit (Thermo Fisher Scientific, Inc.) were used to
visualize the bands. Relative protein levels were determined using
Image J version 1.45s software (National Institutes of Health,
Bethesda, MD, USA).
Statistical analysis
All data are presented as the mean ± standard
deviation. SPSS version 17.0 software (SPSS, Inc., Chicago, IL,
USA) was used to perform statistical analysis. A Student's t-test
was applied to analyze the expression of Rab7 in AD tissues
compared with paired NAD tissues. The association between the
clinicopathological characteristics of patients and Rab7 expression
was determined using Fisher's exact test, analysis of variance
(ANOVA) and a Student's t-test. Data were analyzed via one-way
ANOVA for 3-group comparisons and two tail student's t-tests for
2-group comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results
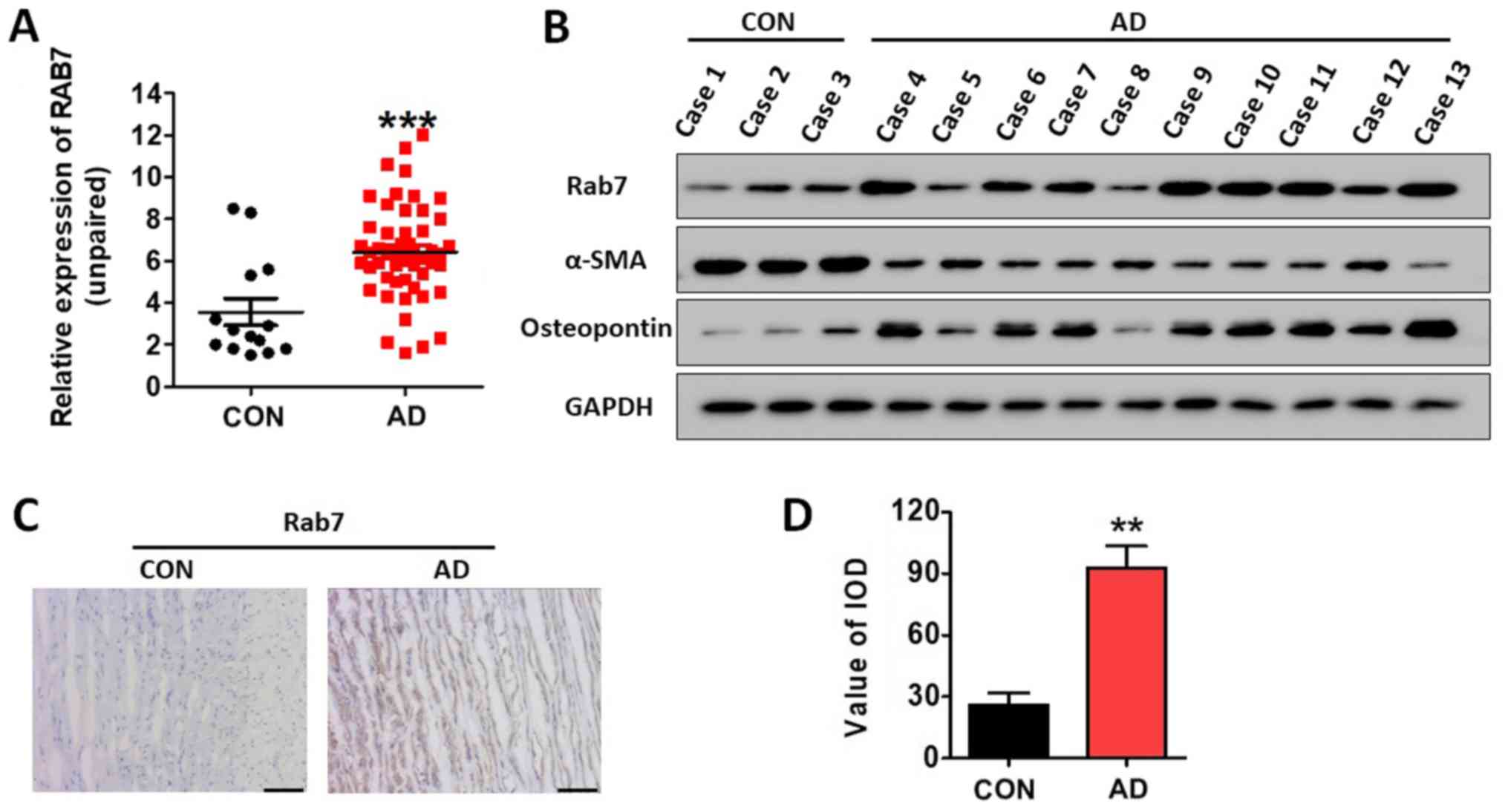
Rab7 is overexpressed in AD tissue and
the abundance of synthetic HASMCs is increased in AD
Rab7 protein expression levels in 14 NAD and 51 AD
specimens were determined by western blotting, and were normalized
to GAPDH. The results revealed that the expression levels of Rab7
were significantly increased in AD specimens compared with the NAD
samples (P<0.001; Fig. 1A).
Table I demonstrated the
association between Rab7 protein expression and the
clinicopathological characteristics in patients with AD. In
addition, the expression of Rab7, α-SMA and osteopontin was
determined by western blotting. It was observed that Rab7 and
osteopontin expression was notably increased in the AD specimens,
while α-SMA expression was decreased compared with the NAD
specimens (Fig. 1B). Using
immunohistochemistry, it was observed that the AD specimens had
significantly higher Rab7 expression compared with in NAD specimens
(Fig. 1C and D). These results
indicated that the abundance of contractile HASMCs was decreased
while synthetic HASMCs were increased in AD (29,30).
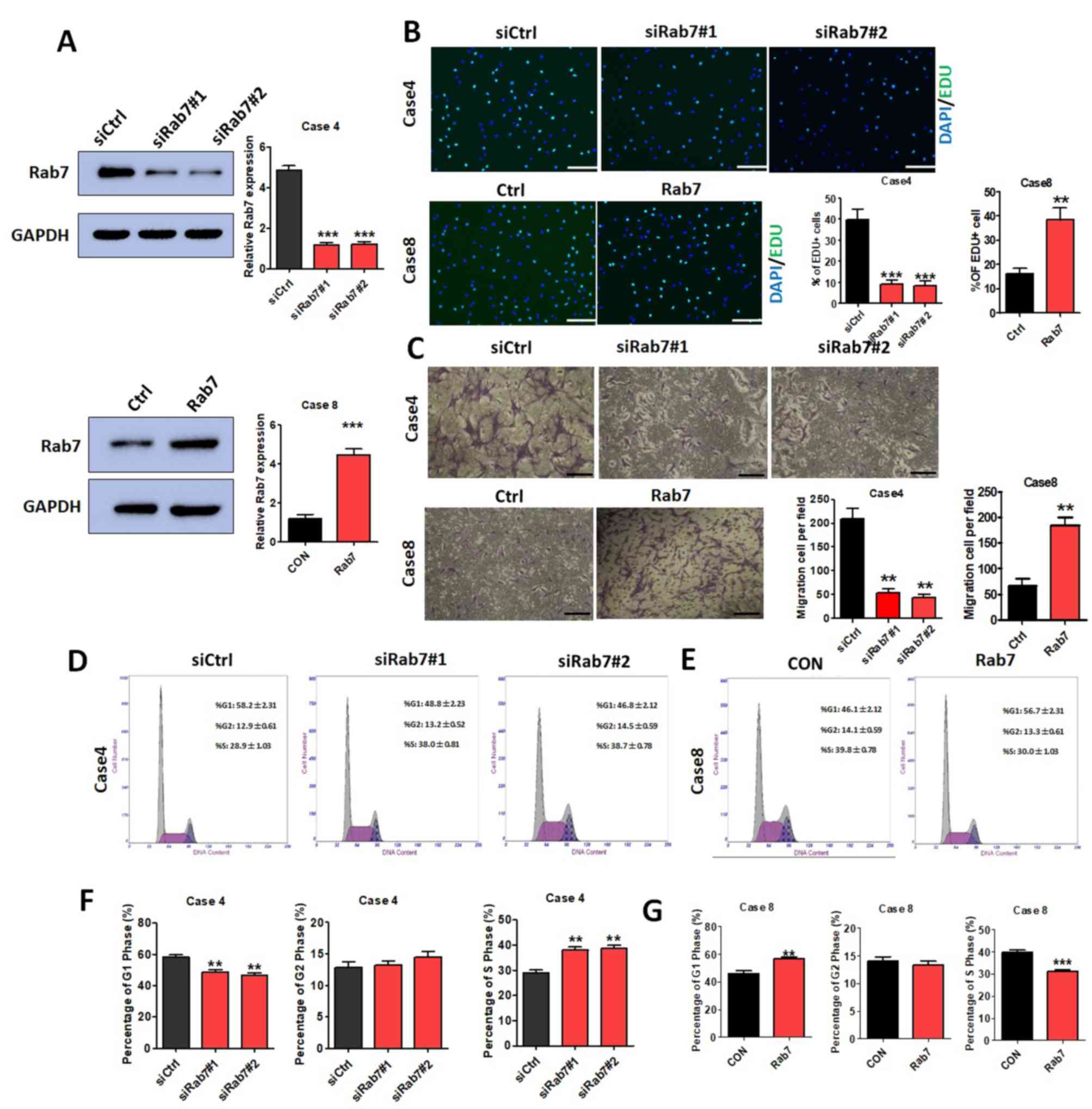
Rab7 regulates HASMC proliferation,
invasion and cell cycle
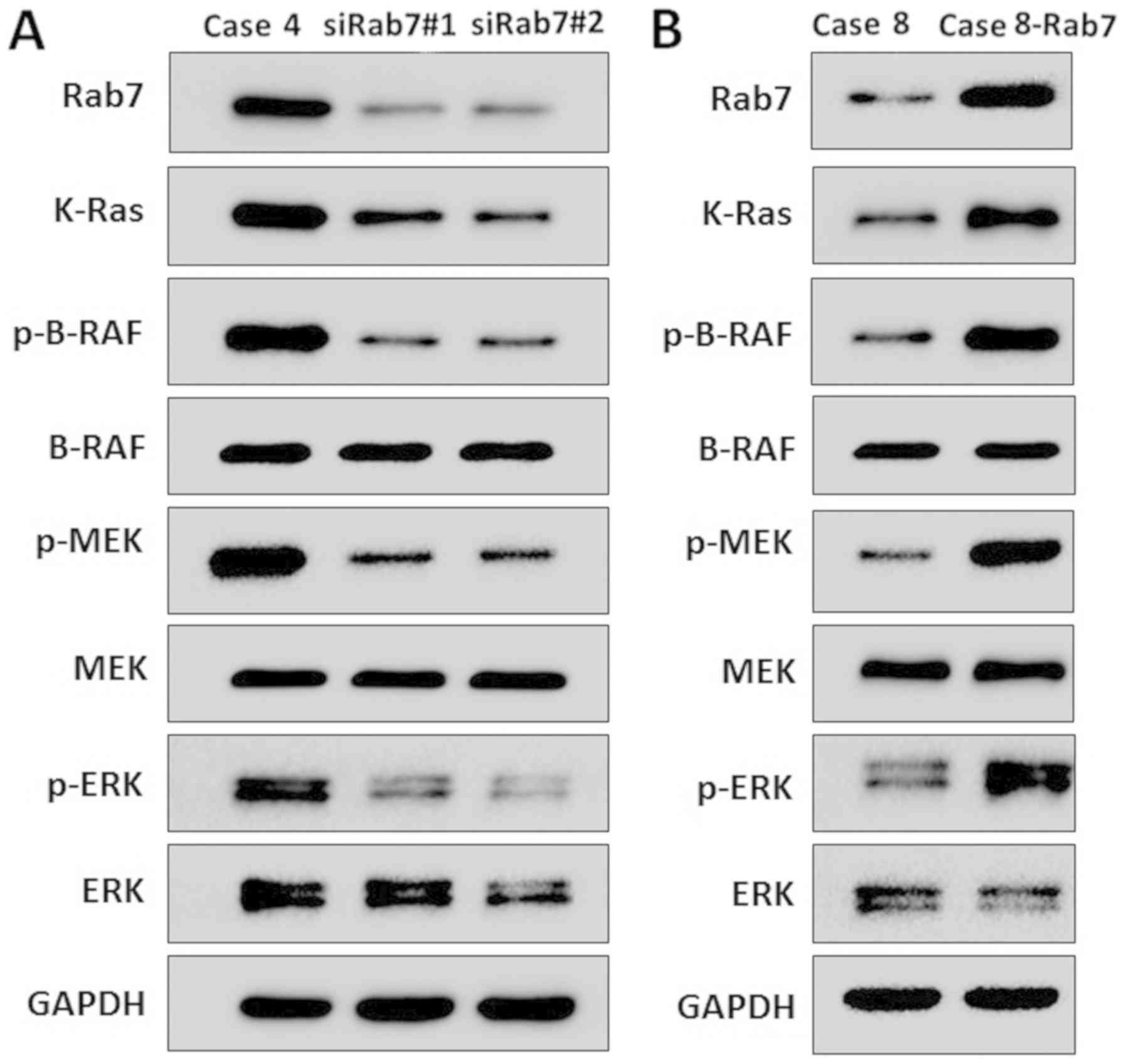
As presented in Fig.
1B, Rab7 expression in Case 4 of the AD group was markedly
increased compared with the NAD group and Rab7 expression in Case 8
was markedly decreased compared with other AD cases. Thus, Rab7
knockdown was performed in Case 4 HASMCs and Rab7 overexpression
was conducted in Case 8 HASMCs to investigate the effects of Rab7
on proliferation, invasion and the cell cycle. The transfection
efficiency was demonstrated for the Case 4 knockdown and Case 8
overexpression using western blot analysis (Fig. 2A).
EdU assays were performed to evaluate the effects of
Rab7 on proliferation. HASMC proliferation was significantly
decreased in Rab7 knockdown compared with the transfected siCtrl
Case 4 control (Fig. 2B) and Rab7
overexpression significantly promoted HASMC proliferation compared
with the transfected siCtrl Case 8 control (Fig. 2B). Rab7 knockdown in HASMCs using
siRab7#1 and siRab7#2 was observed to significantly decrease the
number of invaded cells compared with the Case 4 control; this was
significantly reversed in the Case 8 overexpression group compared
with the control (Fig. 2C). The
results indicated that Rab7 significantly affected the
proliferation and invasion of HASMCs.
Analysis of the cell cycle distribution revealed
that Rab7 was able to regulate the HASMC cycle. Compared with the
Case 4 control, significantly fewer cells were observed in the G1
phase, and a significantly higher proportion of cells in the G2 and
S phase were detected following knockdown of Rab7 with siRab7#1 or
siRab7#2 (Fig. 2D and F). Cell
cycle analysis of HASMCs overexpressing Rab7 exhibited opposing
results, with a significant increase in G1 cell number, and notable
decreases in the G2 and S phase cell number compared with the
control group (Fig. 2E and G). The
results suggested that Rab7 upregulation may induce cell cycle
arrest in the G1 phase.
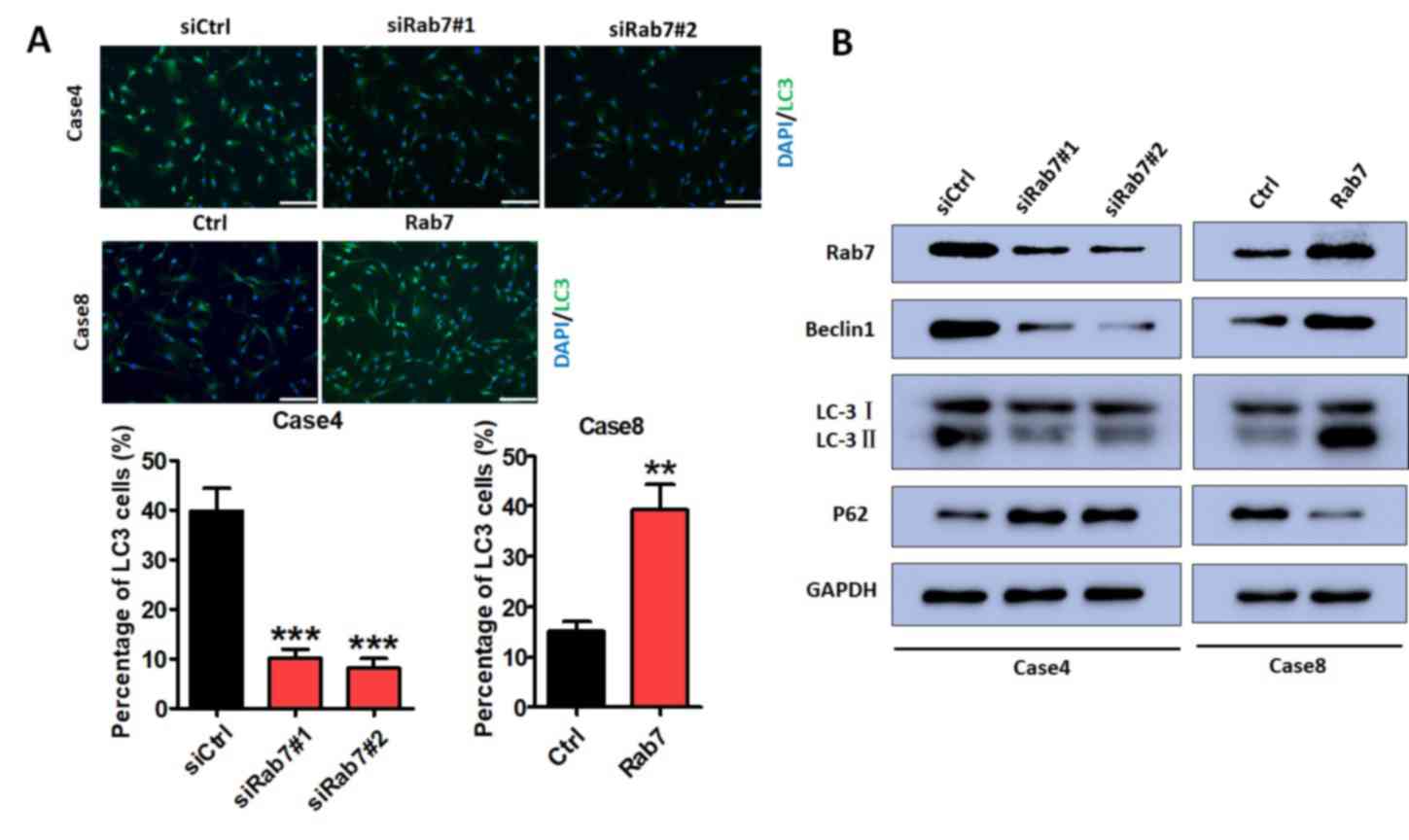
Rab7 regulates autophagy
Immunofluorescence assays were performed to
determine the effects of Rab7 on autophagy. Based on the
literature, Beclin1, LC3-II and P62 were selected as markers in the
evaluation of autophagy. Immunofluorescence assays demonstrated
that HASMC autophagy was significantly decreased following Rab7
knockdown in Case 4 compared with the control cells (Fig. 3A); overexpression promoted HASMC
autophagy compared with Case 8 control cells (Fig. 3A). Rab7 knockdown in Case 4 notably
decreased Beclin1 and LC3-II, and increased P62 expression compared
with the control. These effects were reversed in Rab7
overexpressing cells compared with the Case 8 control (Fig. 3B). This suggested that Rab7
inducted HASMC autophagy.
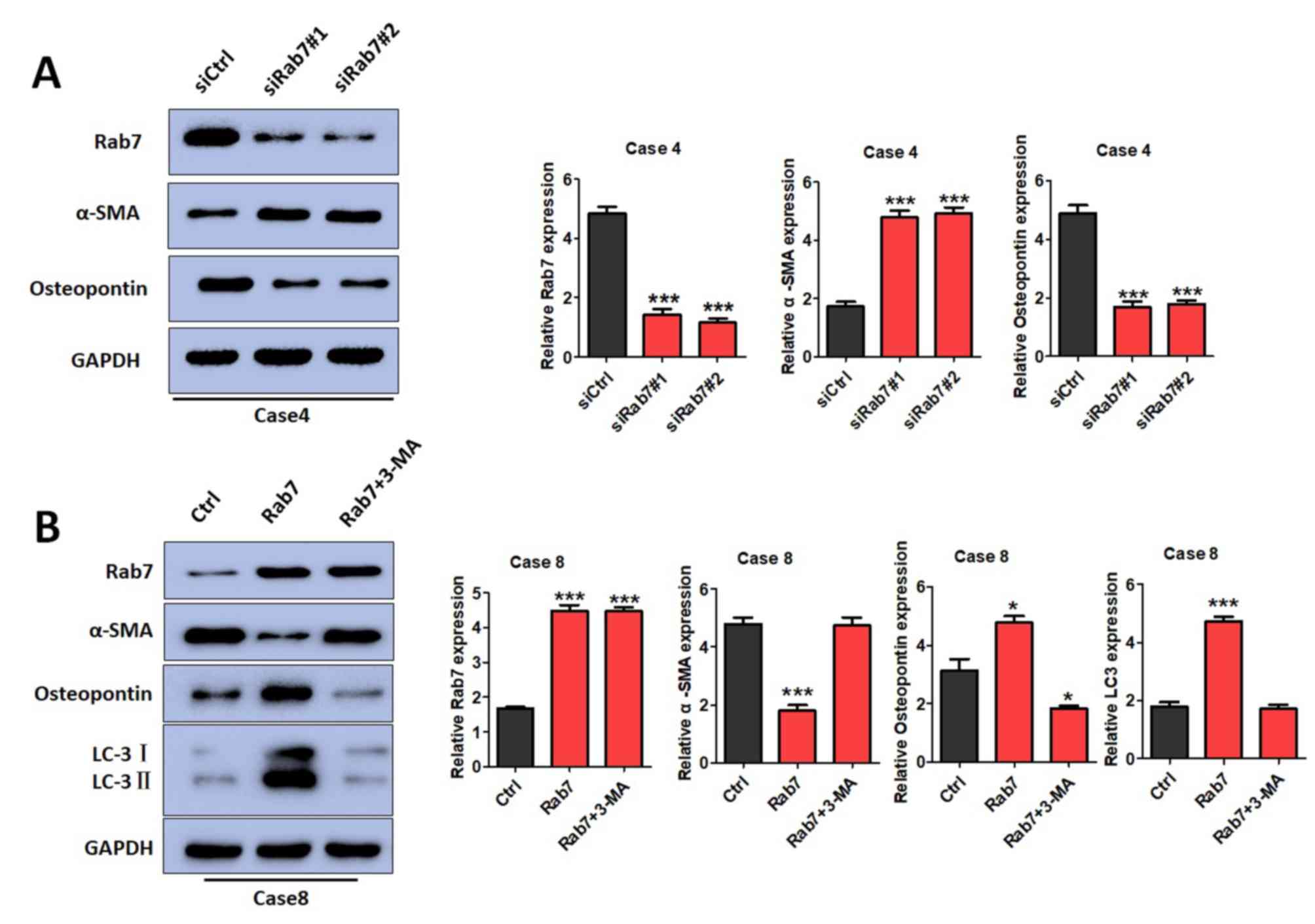
Rab7 regulates the phenotypic
transformations of HASMCs by inducing autophagy
The effects of Rab7 on phenotypic transformations
were determined by detecting α-SMA and osteopontin expression.
α-SMA is mainly expressed in contractile HASMCs, while osteopontin
is mainly expressed in synthetic HASMCs (31,32).
It was demonstrated that α-SMA expression was significantly
increased in Rab7 knockdown cells of Case 4 compared with the
control cells (Fig. 4A) and Rab7
overexpression in Case 8 cells inhibited α-SMA expression compared
with the control cells (Fig. 4B).
Furthermore, osteopontin expression was decreased in Rab7 knockdown
cells and increased in Rab7 overexpression cells compared with the
Case 4 and Case 8 controls, respectively (Fig. 4A and B). The results indicated that
Rab7 significantly promoted HASMC phenotypic transformations from a
contractile to a synthetic phenotype.
To further investigate whether the phenotypic
transformation of SMCs is regulated by Rab7-mediated autophagy, a
pharmacological inhibitor of autophagy, 3-MA, was administered to
Rab7-overexpressing cells to suppress autophagy. When autophagy was
inhibited by 3-MA, it was observed that α-SMA, osteopontin and LC3
expression was markedly altered compared with the control (Fig. 4B). In conclusion, Rab7 was observed
to induce autophagy, which may be associated with the phenotypic
transformation of HASMCs.
Rab7 activates the
Ras/Raf/mitogen-activated protein kinase (MAPK) kinase
(MEK)/extracellular signal-regulated kinase (ERK) signaling pathway
in HASMCs
The Ras/Raf/MEK/ERK signaling pathway serves an
important role in autophagy (33,34).
The expression levels of various molecules of the Ras/Raf/MEK/ERK
signaling pathway were analyzed by western blotting. Rab7 knockdown
resulted in decreased expression of K-Ras, phosphorylated (p)-BRAF,
p-MEK and p-ERK in Case 4 cells compared with the transformed
siCtrl control (Fig. 5A). In
Rab7-overexpressing cells, the expression of K-Ras, p-BRAF, p-MEK
and p-ERK was increased in Case 8 cells compared with the control
(Fig. 5B). As the Ras/Raf/MEK/ERK
signaling pathway is associated with autophagy (35), the results of the present study
suggested that Rab7 induced autophagy by activating the
Ras/Raf/MEK/ERK signaling pathway in HASMCs.
Discussion
Analyses into the pathogenesis of AD have become
more widespread and various factors that contribute to the
development of AD have been identified, such as hereditary
diseases, including Marfan syndrome, hypertension, atherosclerosis
and autoimmune diseases (36–38);
however, the molecular mechanism underlying the development and
progression of AD remains unknown. It is well established that
normal aortic media comprises numerous regularly arranged VSMCs and
extracellular matrix (39). As
SMCs are associated with the structure and function of the aorta,
there is the potential for serving important roles in AD
development (40–42). In healthy individuals, contractile
VSMCs serve a crucial role in maintaining vascular wall structure
and function (43). In patients
with AD, a significant increase in the number and ratio of
synthetic VSMCs was observed, which resulted in decreased aortic
elasticity and rupture of the vessel wall (44,45).
These studies suggested that the phenotypic conversion of VSMCs may
be the key factor underlying the occurrence and development of
AD.
Previous studies have reported that numerous
factors, including vasoconstrictor agonists and extracellular
matrices regulate the phenotypic conversion of VSMCs (46,47).
While contractile VSMCs express α-SMA extensively, synthetic VSMCs
exhibit high expression of osteopontin (31). The present study reported that Rab7
downregulation resulted in increased α-SMA and osteopontin
expression. These results suggested that Rab7 may promote the
phenotypic conversion of contractile to synthetic VSMCs; however,
the specific mechanism remains unclear. Rab7, a member of the Rab
GTPase superfamily, serves an unique role in the regulation of
autophagy (48). Several studies
have revealed that autophagy promotes the phenotypic conversion of
contractile to synthetic VSMCs (19,49,50).
Therefore, the present study proposed that autophagy may be
affected by the phenotypic transformations VSMCs mediated by Rab7.
To further verify this hypothesis, the autophagy inhibitor 3-MA was
applied to cells to investigate whether Rab7 promotes the
phenotypic transformations of VSMCs. It was observed that the
effects of Rab7 on phenotypic transformations were reversed
following autophagy inhibition. In conclusion, autophagy may be a
key component in Rab7-inducing phenotypic transformations. The
effects of Rab7 on phenotypic transformations were revealed via the
suppression of autophagy in Rab7 knockdown cells; the phenotypic
transformations were associated with autophagy in Rab7
overexpressing cells.
Furthermore, as autophagy is regulated by the
Ras/Raf/MEK/ERK signaling pathway (33,35),
it was proposed that Rab7 may induce autophagy-mediated phenotypic
transformations via the Ras/Raf/MEK/ERK signaling pathway. It was
observed that Rab7 knockdown or overexpression regulated the
expression of Ras/Raf/MEK/ERK signaling pathway-associated
proteins. Future experiments are required investigate the effects
of the Ras/Raf/MEK/ERK signaling pathway on Rab7-mediated VSMC
phenotypic transformations.
The present study revealed that Rab7 promoted VSMC
proliferation and invasion in AD. This phenomenon may be associated
with autophagy and phenotypic transformations. Autophagy has been
considered to be the basic catabolic mechanism that is essential
for cell survival, differentiation, development and the cellular
response to stress via the regulating the actions of numerous
lysosomes (51–53). In aortic endothelial cells,
autophagy promotes tube formation and endothelial cell migration by
inducing angiogenesis (54).
Additionally, in lung cancer cells, autophagy facilitates
proliferation, migration and invasion (55). VSMCs of the synthetic phenotype
have enhanced proliferative and migration abilities compared with
the contractile phenotype (56,57).
Compared with healthy controls, the present study positively
correlated VSMC proliferation and invasion with autophagy and
phenotypic transformations in AD. In the present study, autophagy
and phenotypic transformations were reported to be regulated by
Rab7, which in turn was supported by Rab7 promoting VSMC
proliferation and invasion in AD; however, the exact mechanism of
action remains unclear and requires further investigation.
There are certain limitations to the present study.
Synthetic VSMCs synthesize and secrete extracellular matrices
(58); however, the expression of
extracellular matrix proteins, including collagen and elastin were
not detected in the present study. Therefore, the role of Rab7 and
the Ras/Raf/MEK/ERK signaling pathway in extracellular
matrix-associated protein secretion remains unknown. Additionally,
as VSMC phenotypic conversion is associated with AD development and
various cardiovascular events including coronary heart disease and
vascular complications in diabetes (59), further investigation into
phenotypic transition is essential for the prevention and treatment
of particular cardiovascular diseases.
In summary, Rab7 promoted the proliferation and
invasion of VSMCs in AD, which may be mediated by the regulation of
autophagy and phenotypic transformation. Additionally, Rab7
expression was associated with the Ras/Raf/MEK/ERK signaling
pathway. These results indicate the important role of Rab7 in the
etiology of AD and suggested that Rab7 may be considered as a novel
therapeutic target for the treatment of AD; however, further
experiments are required to determine the underlying mechanism of
Rab7 in the regulation of autophagy and phenotypic
transformation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the ‘333’ Project
of Jiangsu Province (grant no. BRA2014336) and the Natural Science
Foundation of Jiangsu Province (grant no. BK20151590).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
KH and HS were in charge of completing most of the
experiments. JZ, RZ, JG and ML aided in the collection of
specimens. JZ and RZ performed western blotting, and JG and ML
conducted the cell experiments. YS provided project design and
guidance.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Nanjing Medical
University (Nanjing, China) and all patients provided written
informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hecht E, Freise C, Websky KV, Nasser H,
Kretzschmar N, Stawowy P, Hocher B and Querfeld U: The matrix
metalloproteinases 2 and 9 initiate uraemic vascular
calcifications. Nephrol Dial Transplant. 31:789–797. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu K, Fang C, Shen Y, Liu Z, Zhang M, Ma
B and Pang X: Hypoxia-inducible factor 1a induces phenotype switch
of human aortic vascular smooth muscle cell through PI3K/AKT/AEG-1
signaling. Oncotarget. 8:33343–33352. 2017.PubMed/NCBI
|
|
3
|
Zhong Y, Feng J, Li J and Fan Z: Curcumin
prevents lipopolysaccharide-induced matrix metalloproteinase-2
activity via the Ras/MEK1/2 signaling pathway in rat vascular
smooth muscle cells. Mol Med Rep. 16:4315–4319. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Freise C, Bobb V and Querfeld U: Collagen
XIV and a related recombinant fragment protect human vascular
smooth muscle cells from calcium-/phosphate-induced
osteochondrocytic transdifferentiation. Exp Cell Res. 358:242–252.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hogarth DK, Sandbo N, Taurin S, Kolenko V,
Miano JM and Dulin NO: Dual role of PKA in phenotypic modulation of
vascular smooth muscle cells by extracellular ATP. Am J Physiol
Cell Physiol. 287:C449–C456. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pei C, Qin S, Wang M and Zhang S:
Regulatory mechanism of human vascular smooth muscle cell
phenotypic transformation induced by NELIN. Mol Med Rep.
12:7310–7316. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yan Y, Wang C, Lu Y, Gong H, Wu Z, Ma X,
Li H, Wang B and Zhang X: Mineralocorticoid receptor antagonism
protects the aorta from vascular smooth muscle cell proliferation
and collagen deposition in a rat model of adrenal
aldosterone-producing adenoma. J Physiol Biochem. 74:17–24. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lv P, Zhang F, Yin YJ, Wang YC, Gao M, Xie
XL, Zhao LL, Dong LH, Lin YL, Shu YN, et al: SM22α inhibits
lamellipodium formation and migration via Ras-Arp2/3 signaling in
synthetic VSMCs. Am J Physiol Cell Physiol. 311:C758–C767. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Potier M, Gonzalez JC, Motiani RK,
Abdullaev IF, Bisaillon JM, Singer HA and Trebak M: Evidence for
STIM1- and Orai1-dependent store-operated calcium influx through
ICRAC in vascular smooth muscle cells: Role in proliferation and
migration. FASEB J. 23:2425–2437. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo RW, Yang LX, Li MQ, Pan XH, Liu B and
Deng YL: Stim1- and Orai1-mediated store-operated calcium entry is
critical for angiotensin II-induced vascular smooth muscle cell
proliferation. Cardiovasc Res. 93:360–370. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meng YH, Tian C, Liu L, Wang L and Chang
Q: Elevated expression of connective tissue growth factor,
osteopontin and increased collagen content in human ascending
thoracic aortic aneurysms. Vascular. 22:20–27. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wanjare M, Kusuma S and Gerecht S:
Defining differences among perivascular cells derived from human
pluripotent stem cells. Stem Cell Reports. 2:561–575. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pahk K, Joung C, Jung SM, Young Song H,
Yong Park J, Woo Byun J, Lee YS, Chul Paeng J, Kim C, Kim S and Kim
WK: Visualization of synthetic vascular smooth muscle cells in
atherosclerotic carotid rat arteries by F-18 FDG PET 7.
69892017.PubMed/NCBI
|
|
15
|
Mack CP: Signaling mechanisms that
regulate smooth muscle cell differentiation. Arterioscler Thromb
Vasc Biol. 31:1495–1505. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pyle AL and Young PP: Atheromas feel the
pressure: Biomechanical stress and atherosclerosis. Am J Pathol.
177:4–9. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma J, Feng Y, Li Z and Tang C: The effect
of adrenomedullin and proadrenomedullin N-terminal 20 peptide on
angiotensin II induced vascular smooth muscle cell proliferation.
Iran J Basic Med Sci. 19:49–54. 2016.PubMed/NCBI
|
|
18
|
Chuang TD, Pearce WJ and Khorram O:
miR-29c induction contributes to downregulation of vascular
extracellular matrix proteins by glucocorticoids. Am J Physiol Cell
Physiol. 309:C117–C125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Salabei JK, Cummins TD, Singh M, Jones SP,
Bhatnagar A and Hill BG: PDGF-mediated autophagy regulates vascular
smooth muscle cell phenotype and resistance to oxidative stress.
Biochem J. 451:375–388. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mima J: Reconstitution of membrane
tethering mediated by Rab-family small GTPases. Biophys Rev.
10:543–549. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Distefano MB, Kjos I, Bakke O and Progida
C: Rab7b at the intersection of intracellular trafficking and cell
migration. Commun Integr Biol. 8:e10234922015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jimenez-Orgaz A, Kvainickas A, Nägele H,
Denner J, Eimer S, Dengjel J and Steinberg F: Control of RAB7
activity and localization through the retromer-TBC1D5 complex
enables RAB7-dependent mitophagy. EMBO J. 37:235–254. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Noda T and Yoshimori T: Between canonical
and antibacterial autophagy: Rab7 is required for GAS-containing
autophagosome-like vacuole formation. Autophagy. 6:419–420. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cataldo AM, Mathews PM, Boiteau AB,
Hassinger LC, Peterhoff CM, Jiang Y, Mullaney K, Neve RL, Gruenberg
J and Nixon RA: Down syndrome fibroblast model of Alzheimer-related
endosome pathology: Accelerated endocytosis promotes late endocytic
defects. Am J Pathol. 173:370–384. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang M, Chen L, Wang S and Wang T: Rab7:
Roles in membrane trafficking and disease. Biosci Rep. 29:193–209.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ismail S, Gherardi MJ, Froese A, Zanoun M,
Gigoux V, Clerc P, Gaits-Iacovoni F, Steyaert J, Nikolaev VO and
Fourmy D: Internalized receptor for glucose-dependent
insulinotropic peptide stimulates adenylyl cyclase on early
endosomes. Biochem Pharmacol. 120:33–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han J, Pan XY, Xu Y, Xiao Y, An Y, Tie L,
Pan Y and Li XJ: Curcumin induces autophagy to protect vascular
endothelial cell survival from oxidative stress damage. Autophagy.
8:812–825. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Corbera-Bellalta M, Planas-Rigol E, Lozano
E, Terrades-García N, Alba MA, Prieto-González S, García-Martínez
A, Albero R, Enjuanes A, Espígol-Frigolé G, et al: Blocking
interferon γ reduces expression of chemokines CXCL9, CXCL10 and
CXCL11 and decreases macrophage infiltration in ex vivo cultured
arteries from patients with giant cell arteritis. Ann Rheum Dis.
75:1177–1186. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun HJ, Ren XS, Xiong XQ, Chen YZ, Zhao
MX, Wang JJ, Zhou YB, Han Y, Chen Q, Li YH, et al: NLRP3
inflammasome activation contributes to VSMC phenotypic
transformation and proliferation in hypertension. Cell Death Dis.
8:e30742017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang X, Zhao JF, Zhao F, Yan JF, Yang F,
Huang XJ, Chen G, Fu HY and Lv BD: The protective effect of
salidroside on hypoxia-induced corpus cavernosum smooth muscle cell
phenotypic transformation. Evid Based Complement Alternat Med.
2017:35302812017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cao C, Ji X, Luo X and Zhong L: Gingipains
from Porphyromonas gingivalis promote the transformation and
proliferation of vascular smooth muscle cell phenotypes. Int J Clin
Exp Med. 8:18327–18334. 2015.PubMed/NCBI
|
|
32
|
Tian L, Chen K, Cao J, Han Z, Gao L, Wang
Y, Fan Y and Wang C: Galectin-3-induced oxidized low-density
lipoprotein promotes the phenotypic transformation of vascular
smooth muscle cells. Mol Med Rep. 12:4995–5002. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Butler DE, Marlein C, Walker HF, Frame FM,
Mann VM, Simms MS, Davies BR, Collins AT and Maitland NJ:
Inhibition of the PI3K/AKT/mTOR pathway activates autophagy and
compensatory Ras/Raf/MEK/ERK signalling in prostate cancer.
Oncotarget. 8:56698–56713. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Nie H, Zhao X, Qin Y and Gong X:
Bicyclol induces cell cycle arrest and autophagy in HepG2 human
hepatocellular carcinoma cells through the PI3K/AKT and
Ras/Raf/MEK/ERK pathways. BMC Cancer. 16:7422016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xin L, Ma X, Xiao Z, Yao H and Liu Z:
Coxsackievirus B3 induces autophagy in HeLa cells via the
AMPK/MEK/ERK and Ras/Raf/MEK/ERK signaling pathways. Infect Genet
Evol. 36:46–54. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Agg B, Benke K, Szilveszter B, Pólos M,
Daroczi L, Odler B, Nagy ZB, Tarr F, Merkely B and Szabolcs Z:
Possible extracardiac predictors of aortic dissection in Marfan
syndrome. BMC Cardiovasc Disord. 14:472014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gaddum NR, Keehn L, Guilcher A, Gomez A,
Brett S, Beerbaum P, Schaeffter T and Chowienczyk P: Altered
dependence of aortic pulse wave velocity on transmural pressure in
hypertension revealing structural change in the aortic wall.
Hypertension. 65:362–369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nienaber CA, Clough RE, Sakalihasan N,
Suzuki T, Gibbs R, Mussa F, Jenkins MP, Thompson MM, Evangelista A,
Yeh JS, et al: Aortic dissection. Nat Rev Dis Primers. 2:160532016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lacolley P, Regnault V, Segers P and
Laurent S: Vascular smooth muscle cells and arterial stiffening:
Relevance in development, aging, and disease. Physiol Rev.
97:1555–1617. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Docherty CK, Carswell A, Friel E and
Mercer JR: Impaired mitochondrial respiration in human carotid
plaque atherosclerosis: A potential role for Pink1 in vascular
smooth muscle cell energetics. Atherosclerosis. 268:1–11. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Iida Y, Tanaka H, Sano H, Suzuki Y,
Shimizu H and Urano T: Ectopic expression of PCSK9 by smooth muscle
cells contributes to aortic dissection. Ann Vasc Surg. 48:195–203.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wei X, Sun Y, Wu Y, Zhu J, Gao B, Yan H,
Zhao Z, Zhou J and Jing Z: Downregulation of Talin-1 expression
associates with increased proliferation and migration of vascular
smooth muscle cells in aortic dissection. BMC Cardiovasc Disord.
17:1622017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang L, Gao L, Nickel T, Yang J, Zhou J,
Gilbertsen A, Geng Z, Johnson C, Young B, Henke C, et al: Lactate
promotes synthetic phenotype in vascular smooth muscle cells. Circ
Res. 121:1251–1262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Perrucci GL, Rurali E, Gowran A, Pini A,
Antona C, Chiesa R, Pompilio G and Nigro P: Vascular smooth muscle
cells in Marfan syndrome aneurysm: The broken bricks in the aortic
wall. Cell Mol Life Sci. 74:267–277. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cai YL and Wang ZW: The expression and
significance of IL-6, IFN-γ, SM22α, and MMP-2 in rat model of
aortic dissection. Eur Rev Med Pharmacol Sci. 21:560–568.
2017.PubMed/NCBI
|
|
46
|
Zhou W, Liu W, Liao H, Cao Z, Xie H, Zhang
S and Chen M: Testosterone suppresses oxidized low-density
lipoprotein-induced vascular smooth muscle cell phenotypic
transition and proliferation. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi.
31:775–778. 2015.(In Chinese). PubMed/NCBI
|
|
47
|
Wang L, Zheng J, Du Y, Huang Y, Li J, Liu
B, Liu CJ, Zhu Y, Gao Y, Xu Q, et al: Cartilage oligomeric matrix
protein maintains the contractile phenotype of vascular smooth
muscle cells by interacting with alpha(7)beta(1) integrin. Circ Re.
106:514–525. 2010. View Article : Google Scholar
|
|
48
|
Zhan L, Chen S, Li K, Liang D, Zhu X, Liu
L, Lu Z, Sun W and Xu E: Autophagosome maturation mediated by Rab7
contributes to neuroprotection of hypoxic preconditioning against
global cerebral ischemia in rats. Cell Death Dis. 8:e29492017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Song TF, Huang LW, Yuan Y, Wang HQ, He HP,
Ma WJ, Huo LH, Zhou H, Wang N and Zhang TC: LncRNA MALAT1 regulates
smooth muscle cell phenotype switch via activation of autophagy.
Oncotarget. 9:4411–4426. 2017.PubMed/NCBI
|
|
50
|
Zhao Y, Guo Y, Jiang Y, Zhu X, Liu Y and
Zhang X: Mitophagy regulates macrophage phenotype in diabetic
nephropathy rats. Biochem Biophys Res Commun. 494:42–50. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Levine B and Klionsky DJ: Development by
self-digestion: Molecular mechanisms and biological functions of
autophagy. Dev Cell. 6:463–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Matsuo T and Sadzuka Y: Extracellular
acidification by lactic acid suppresses glucose deprivation-induced
cell death and autophagy in B16 melanoma cells. Biochem Biophys Res
Commun. 496:1357–1361. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Grootaert MOJ, Moulis M, Roth L, Martinet
W, Vindis C, Bennett MR and De Meyer GRY: Vascular smooth muscle
cell death, autophagy and senescence in atherosclerosis. Cardiovasc
Res. 114:622–634. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Slevin M, Krupinski J, Rovira N, Turu M,
Luque A, Baldellou M, Sanfeliu C, de Vera N and Badimon L:
Identification of pro-angiogenic markers in blood vessels from
stroked-affected brain tissue using laser-capture microdissection.
BMC Genomics. 10:1132009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhan Z, Xie X, Cao H, Zhou X, Zhang XD,
Fan H and Liu Z: Autophagy facilitates TLR4- and TLR3-triggered
migration and invasion of lung cancer cells through the promotion
of TRAF6 ubiquitination. Autophagy. 10:257–268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Stone JD, Narine A, Shaver PR, Fox JC,
Vuncannon JR and Tulis DA: AMP-activated protein kinase inhibits
vascular smooth muscle cell proliferation and migration and
vascular remodeling following injury. Am J Physiol Heart Circ
Physiol. 304:H369–H381. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu F, Ahmed AS, Kang X, Hu G, Liu F, Zhang
W and Zhou J: MicroRNA-15b/16 attenuates vascular neointima
formation by promoting the contractile phenotype of vascular smooth
muscle through targeting YAP. Arterioscler Thromb Vasc Biol.
35:2145–2152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yan C: Cyclic nucleotide phosphodiesterase
1 and vascular aging. Clin Sci (Lond). 129:1077–1081. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
McCarty MF and DiNicolantonio JJ: The
molecular biology and pathophysiology of vascular calcification.
Postgrad Med. 126:54–64. 2014. View Article : Google Scholar : PubMed/NCBI
|