Introduction
Ovarian cancer demonstrates the highest mortality
rate among gynecological cancers and it is difficult to diagnose at
an early stage (1). Thus, due to
the majority of diagnoses occurring at advanced stages, ovarian
cancer is difficult to treat and is often characterized by a low
5-year survival rate. In addition to surgical therapy, treatment of
ovarian cancer often consists of chemotherapy and immunotherapy
(2,3). Chemotherapy is the main adjuvant
therapy used for ovarian cancer, which not only affects the
condition of ovarian cancer but also exerts cytotoxic activity to
thoroughly eliminate cancer cells and prolong the survival time of
patients. Nonetheless, resistance of cancer cells to
chemotherapeutic drugs, including cisplatin (DDP), paclitaxel,
vincristine and bleomycin, is a growing concern. For example,
ovarian cancer cells are often resistant to paclitaxel and DDP
(4,5); DDP resistance is associated with an
increase in DNA repair and drug inactivation, as well as changes in
cellular accumulation, which prevent the efficacy of cancer
treatments (6). It has been
reported that STAT3 regulates microRNA (miRNA/miR)-216a to
facilitate ovarian cancer proliferation and DDP resistance
(7). Ai et al (8) reported that hypoxia-inducible
factor-1 α (HIF-1α) inhibited the response of DDP-resistant ovarian
cancer cells to DDP by redirecting aerobic glycolysis towards
mitochondrial oxidative phosphorylation, which promoted cellular
survival through the underproduction of reactive oxygen species.
Overall, this finding suggested that the HIF-1α-regulated cancer
metabolism pathway may be a novel target for overcoming DDP
resistance in ovarian cancer. Therefore, it is of importance to
investigate the specific molecular mechanisms underlying DDP
resistance in ovarian cancer to identify novel drug targets and
improve the survival rate of patients with ovarian cancer.
Propofol, a central nervous system anesthetic, is
often used in surgical operations in combination with inhalational
anesthetics and analgesics. In addition to its function as an
anesthetic, the antitumor effects of propofol have been
demonstrated in gastric, lung, cervical and breast cancer (9–12). A
previous study reported that propofol inhibited the invasion of
ovarian cancer cells and enhanced the apoptotic effect of
paclitaxel on ovarian cancer cells (13); however, the molecular mechanisms
underlying these specific roles of propofol in ovarian cancer are
largely unknown.
miRNAs are RNA molecules 21–23 nucleotides in
length, which do not encode proteins but regulate gene expression
through binding to specific miRNA-binding sites on target mRNAs
(14,15). miRNAs act as biological regulators
in a range of cellular processes, such as cell proliferation,
invasion and apoptosis or programmed cell death (16,17).
Notably, miR-374a negatively regulates its downstream genes to
control the proliferation and invasion of cancer cells (18). Emerging evidence indicated that the
transcription factor forkhead box O1 (FOXO1) serves important roles
in controlling drug resistance in cancer cells; for example, it has
been reported that FOXO1 contributes to paclitaxel-induced drug
resistance in ovarian cancer (19). In addition, Wang et al
(20) reported that the role of
FOXO1 in paclitaxel resistance was positively regulated by
thioredoxin-1 (Trx1), and this effect may depend on Trx1 nuclear
translocation, which was mediated by paclitaxel-induced reactive
oxygen species in ovarian cancer cells. Moreover, miR-374a
modulated DDP resistance in human ovarian cancer cells (21). These results suggest that the
expression of miR-374a is associated with drug resistance in
ovarian cancer cells; however, whether there is an interaction
between propofol and miR-374a remains to be identified.
In the present study, the effect of propofol on
miR-374a-induced proliferation and DDP resistance of ovarian cancer
cells was investigated. It was demonstrated that propofol inhibited
the growth and DDP resistance of ovarian cancer cells by decreasing
miR-374a expression and consequently regulating FOXO1 expression.
These findings provided a novel insight into the use of anesthetics
for the treatment of ovarian cancer.
Materials and methods
Cell culture
Human ovarian cancer cell lines A2780 and
DDP-resistant A2780/DDP were obtained from the American Type
Culture Collection and Shanghai Enzyme Research Biotechnology Co.,
Ltd., respectively. Cells were cultured in DMEM supplemented with
10% FBS (both purchased from Gibco; Thermo Fisher Scientific, Inc.)
and maintained in a humidified atmosphere of 5% CO2 at
37°C. Upon A2780 or A2780/DDP cells reaching 80–90% confluence, the
cells were subcultured or inoculated for further experiments.
Drug treatment
DDP and propofol were purchased from Sigma-Aldrich;
Merck KGaA. Different concentrations of propofol and DDP were used
to treat ovarian cancer cell lines in a cell viability assay using
different concentrations. For single agent treatment groups,
ovarian cancer cells were treated with 0, 1, 5, 10 or 20 µg/ml
propofol for 48 h at 37°C, or 0, 5, 10, 20 and 50 µM DDP for 48 h
at 37°C. For the combined treatment group, A2780 or A2780/DDP cells
were treated with 10 µg/ml propofol and 10 µM DDP for 48 h at 37°C.
Cells were transfected with miR-374a or NC mimics before treatment
with propofol and/or DDP.
Cell transfection
miR-374a mimics, miR-374a inhibitor and their
respective negative controls (NC) with random sequences (denoted as
mimics NC and inhibitor NC) were all synthesized by Shanghai
GenePharma Co., Ltd. The sequences were as follows: Mimics NC
forward, 5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′; miR-374a mimics forward,
5′-UUAUAAUACAACCUGAUAAGUG-3′ and reverse,
5′-CUUAUCAGGUUGUAUUAUAAUU-3′; inhibitors NC
5′-CAGUACUUUUGUGUAGUACAAA-3′; miR-374a inhibitor
5′-CACUUAUCAGGUUGUAUUAUAA-3′. Full-length FOXO1 sequences were
amplified and ligated into pcDNA3.1 plasmid (OriGene Technologies,
Inc.) and the recombined plasmid was referred to as FOXO1. The
cells were transfected with miR-374a mimics, mimics NC, miR-374a
inhibitor and inhibitor NC (50 nm) using Lipofectamine®
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Subsequently, the cells
were treated with 10 µg/ml propofol and 10 µM DDP treatment for 48
h. RNA extraction and western blotting were conducted 48 h
post-transfection.
Cell viability assay
Cell viability was measured using a Cell Counting
kit-8 (CCK-8) assay (Beyotime Institute of Biotechnology) according
to manufacturer's protocol. Briefly, 1×104 cells were
seeded into 96-well plates and following 10 µg/ml propofol
treatment or transfection, 10 µl CCK-8 solution was added into each
well. Following incubation for 4 h at 37°C, the absorbance was
measured at 490 nm using a spectrophotometer. Cell viability was
calculated according to the following formula: Cell viability (% of
control)=Experimental group (OD value)-Blank group (OD)/control
group (OD)-Blank group (OD).
Flow cytometric detection of apoptosis
and cell cycle progression
A total of 3×105 cells treated with
propofol were cultured in 6-well plates for 24 h. Cells were
harvested using trypsin, washed twice with PBS, and subsequently
subjected to an apoptosis assay with an Annexin V-FITC/PI Apoptosis
Detection kit (BD Pharmingen; BD Biosciences), according to the
manufacturer's protocol. Apoptotic cells were analyzed using a flow
cytometry system (BD FACSCalibur; BD Biosciences) using FL1
(excitation, 488 nm) and FL2 (excitation, 633 nm) channels. Data
were analyzed using BD CellQuest™ Pro Analysis software (BD
Biosciences).
Cell cycle analysis was performed using a kit from
Nanjing KeyGen Biotech Co., Ltd., according to the manufacturer's
protocol. Briefly, the cells were collected and fixed with 70%
ethanol at 4°C overnight. The cells were subsequently resuspended
in PBS, and were incubated in buffer containing 50 µg/ml PI and 10
µg/ml RNase A (Beyotime Institute of Biotechnology) for 30 min in
the dark at 37°C. Cell cycle progression was determined using a BD
LSR II flow cytometry system (BD Biosciences), and FlowJo software
v10.0.7 (Tree Star, Inc.) was used for further data analysis.
Detection of cell sensitivity to DDP
using an MTT assay
To evaluate cellular sensitivity to DDP, the
viability of cells treated with 5–50 µM DDP (Sigma-Aldrich; Merck
KGaA) for 48 h was analyzed using the MTT assay (Beyotime Institute
of Biotechnology). Briefly, a total of 5,000 cells were plated in
96-well plates and cultured overnight. Then, 10 µl MTT solution and
100 µl DMEM were added to each well for 4 h at 37°C. The solution
was subsequently discarded and 150 µl DMSO/well was added to
dissolve the formazan crystals. The plates were slightly agitated
for 10 min and the absorbance was determined at 490 nm using a
spectrophotometer. The half-maximal inhibitory concentration (IC50)
values were calculated by nonlinear regression analysis using
GraphPad Prism version 5.0 (GraphPad Software, Inc.) software.
Detection of caspase-3 and caspase-9
activity
Caspase-3 and caspase-9 activities were assessed by
the colorimetric method using Caspase-3 and Caspase-9 activity
assay kits (Beyotime Institute of Biotechnology), respectively,
according to the manufacturer's protocol. The cells were counted
and washed in PBS. The samples were collected by centrifugation
(600 × g; 5 min; 4°C), suspended in 100 µl lysis buffer (Beyotime
Institute of Biotechnology) and kept on ice for 20 min. Following
centrifugation (20,000 × g; 10 min; 4°C), the supernatant was
collected for analysis. A total of 2 mM Ac-DEVD-pNA or Ac-LEHD-pNA
was used to measure caspase-3 activity or caspase-9 activity,
respectively. The absorbance was measured at 405 nm using a
microplate reader, and caspase-3 and caspase-9 activity was
calculated based on the relative value of p-nitroanilide
content.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using RNAiso Plus
(Takara Biotechnology, Co., Ltd.). Following quantification, total
RNA was reverse transcribed into cDNA using a PrimeScript RT master
mix kit (Takara Biotechnology, Co., Ltd.), the reverse
transcription reaction conditions were set to 37°C for 30 min, and
reverse transcriptase inactivation was conducted at 85°C for 5 min.
qPCR was subsequently performed using the SYBR® Green
system (Takara Biotechnology, Co., Ltd.). The primer pairs used for
the qPCR are presented in Table I.
The following thermocycling conditions were used for qPCR:
Pre-denaturation at 95°C for 10 min, followed by a total of 40
cycles of denaturation at 95°C for 10 sec, annealing at 60°C for 50
sec and extension at 72°C for 32 sec. The relative expression
levels of miR-374a or FOXO1 were calculated using the
2−ΔΔCq method (22).
For mRNA expression levels, β-actin was used as the internal
control; for miRNA expression, U6 was used as the internal
control.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| MicroRNA-374a | F:
GCCGGCTTATAATACAACCTGA |
|
| R:
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCACTTA |
| FOXO1 | F:
TGCATTTCGCTACCCGAGTT |
|
| R:
GTGGCTGACAAGACTTAACTCAA |
| GAPDH | F:
CCAGGTGGTCTCCTCTGA |
|
| R:
GCTGTAGCCAAATCGTTGT |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
Western blotting
Cells were cultured in 6-well plates, collected and
total protein was extracted using RIPA lysis buffer (Beyotime
Institute of Biotechnology). Total protein was quantified using a
bicinchoninic acid assay kit (Bio-Rad Laboratories, Inc.) and 50 µg
protein samples were separated via SDS-PAGE gel on a 10% gel. The
separated proteins were subsequently transferred onto
nitrocellulose membranes (Merck KGaA) and blocked at room
temperature for 1 h with 5% skimmed milk. The membranes were
incubated with the following primary antibodies at 4°C overnight:
Anti-FOXO1 (1:1,000; ab39670; Abcam); anti-Bim (1:1,000; ab32158;
Abcam); anti-p27 (1:1,000; ab32034; Abcam); and anti-GAPDH
(1:2,000; ab8245; Abcam). The membranes were washed three times
with TBS containing 0.1% Tween 20. Following the primary antibody
incubation, membranes were incubated with goat anti-mouse/rabbit
IgG horseradish peroxidase-conjugated secondary antibodies
(1:5,000; A6715/R9130; Sigma-Aldrich; Merck KGaA) for 2 h at room
temperature. Protein bands were visualized using an ECL detection
kit (Pierce; Thermo Fisher Scientific, Inc.) and protein expression
was semi-quantified by ImageJ 1.52a (National Institutes of
Health).
Bioinformatics analysis
Bioinformatics tools, including RAID version 2.0
software (www.rna-society.org/raid/) and miRBase (www.mirbase.org) were performed to predict the targets
of miR-374a. The gene FOXO1 was identified as a potential target
gene of miR-374a.
Dual-luciferase reporter assay
The dual-luciferase reporter assay was conducted as
previously described (23), with a
slight modification. The mutation type 3′-untranslated region (UTR)
of FOXO1 gene was created by the QuikChange Site-Directed
Mutagenesis kit (Stratagene; Agilent Technologies, Inc.). The
wild-type or mutant (MUT) FOXO1 3′-UTR were cloned into psiCHECK™-2
vector (Promega Corporation). A total of 2×104 293T
cells (ATCC) were plated into 24-well plates and were subsequently
co-transfected with wild-type or MUT plasmids (200 ng) FOXO1 3′-UTR
vectors and miR-374a mimics or mimics NC using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 48 h. The firefly luciferase activity
of 293T cells was detected using the Dual-Luciferase Reporter assay
system (Promega Corporation), according to the manufacturer's
protocol. The firefly luciferase activity of 293T was compared with
Renilla luciferase activity.
Statistical analysis
Statistical analysis of all results was performed
using GraphPad Prism version 5.0 software (GraphPad Software,
Inc.). Each experiment was independently conducted≥3 times and all
data are presented as the mean ± SD. Significant differences
between groups were determined using Student's t-test or one-way
ANOVA followed by Tukey's post hoc test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Propofol decreases cell viability and
induces apoptosis of ovarian cancer cells
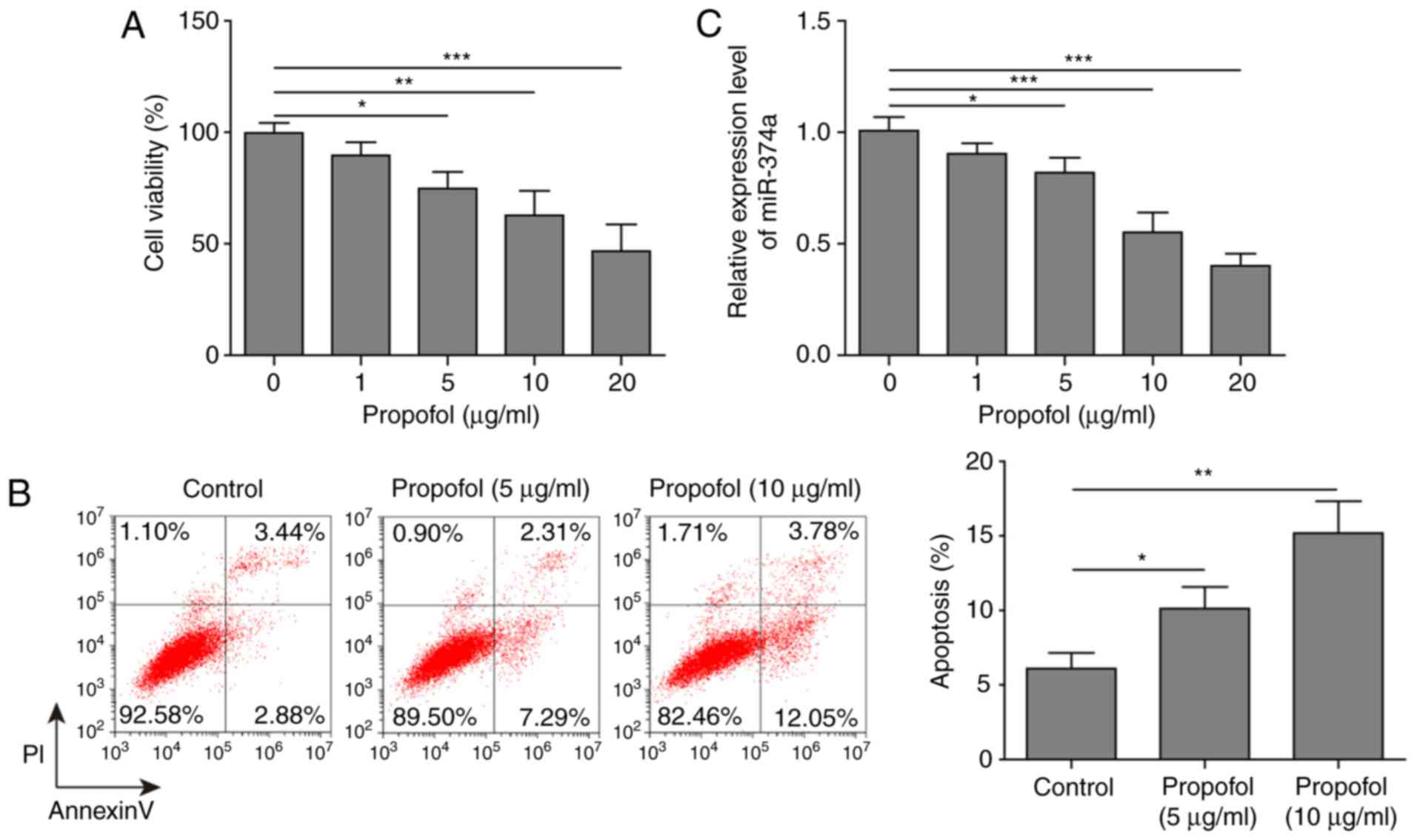
To explore the effect of propofol on A2780 cells,
the cell viability and apoptotic rates of A2780 cells treated with
0, 1, 5, 10 or 20 µg/ml propofol were determined. The results
demonstrated that propofol exerted a significant dose-dependent
inhibitory effect on cell viability and a significant
dose-dependent promoting effect on the apoptotic rate of A2780
cells compared with untreated cells (Fig. 1A and B). To determine the
association between the dose of propofol and miR-374a expression,
RT-qPCR analysis was performed. Propofol significantly decreased
the expression levels of miR-374a in A2780 cells in a
dose-dependent manner compared with untreated cells (Fig. 1C). Overall, it was demonstrated
that 10 µg/ml propofol inhibited ovarian cancer cell viability,
while still maintaining moderate cell viability; however, 20 µg/ml
led to significant cell damage compared with untreated cells
(Fig. 1A). Thus, 10 µg/ml propofol
was selected as the optimal concentration and used in subsequent
experiments.
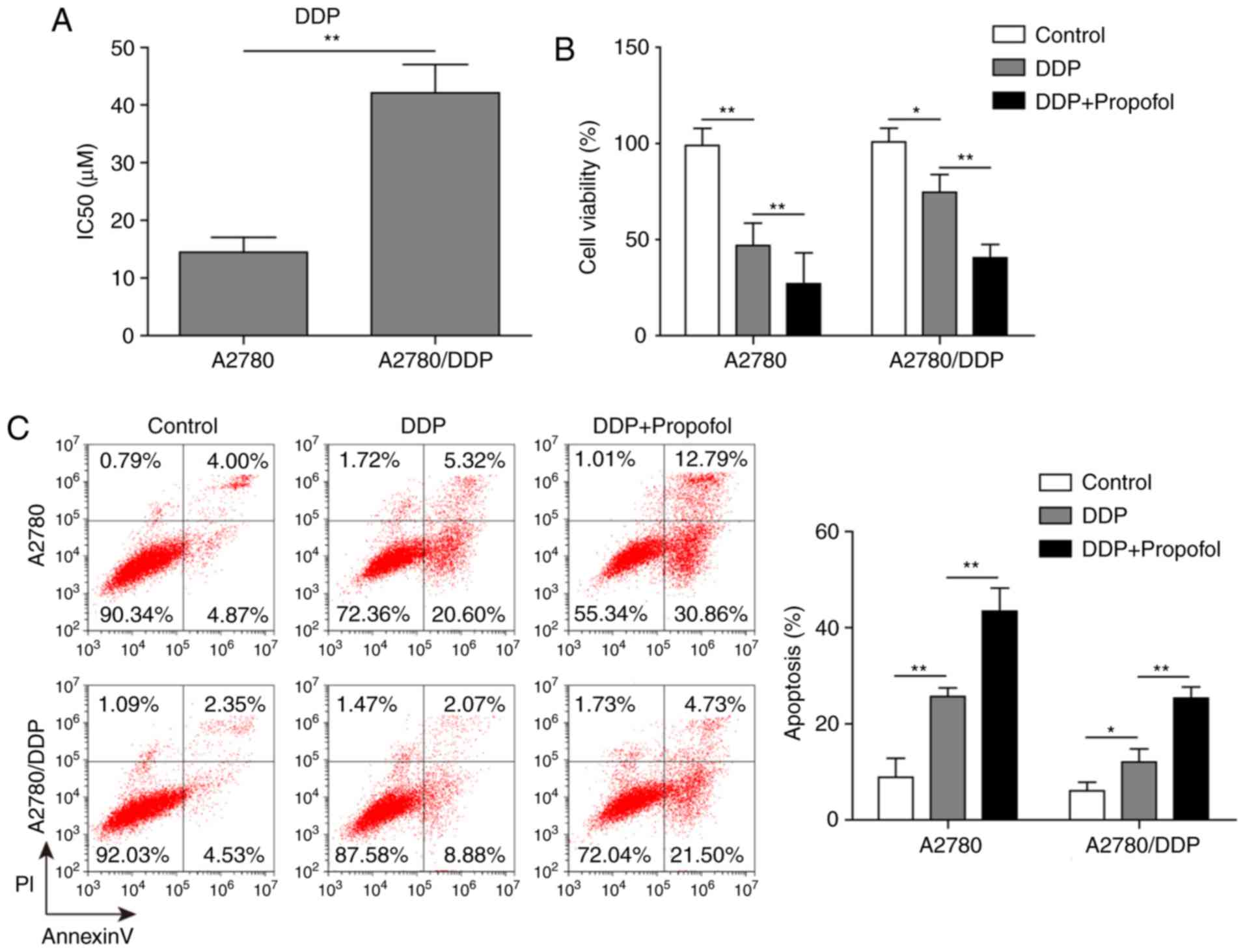
Propofol enhances DDP-induced
cytotoxicity in sensitive and resistant A2780 cell lines
The IC50 of DDP in resistant A2780/DDP
cells was significantly higher compared with A2780 cells (40.2 vs.
14.7 µM; Fig. 2A). This result
suggested that A2780/DDP cells were highly resistant to DDP. By
combining the findings of previous literature and the results of
the present study, a concentration near IC50 (10 µM DDP) was chosen
for subsequent experiments. To investigate the effect of propofol
on DDP resistance in ovarian cancer cells, an MTT assay and flow
cytometric analysis were used to assess the viability and apoptosis
of A2780/DDP and A2780 cells treated with DDP alone or combined
with propofol. Compared with the control group, the viability of
A2780 cells in the DDP-treated group was evidently reduced; but the
cell viability of A2780/DDP only decreased slightly; while the
viability of A2780/DDP cells in the propofol and DDP treatment
group was significantly reduced compared with the control group or
the DDP-only treatment group (Fig.
2B). In addition, treatment with DDP alone markedly increased
the apoptosis of A2780 cells, but the apoptosis of A2780/DDP cells
only increased slightly; whereas the proportion of apoptotic cells
was significantly increased in A2780/DDP cells treated with
propofol and DDP compared with control cells or cells treated with
DDP only (Fig. 2C). Taken
together, these results suggested that propofol pretreatment
inhibited the activity of ovarian cancer cells, induced apoptosis
and increased cell sensitivity to DDP.
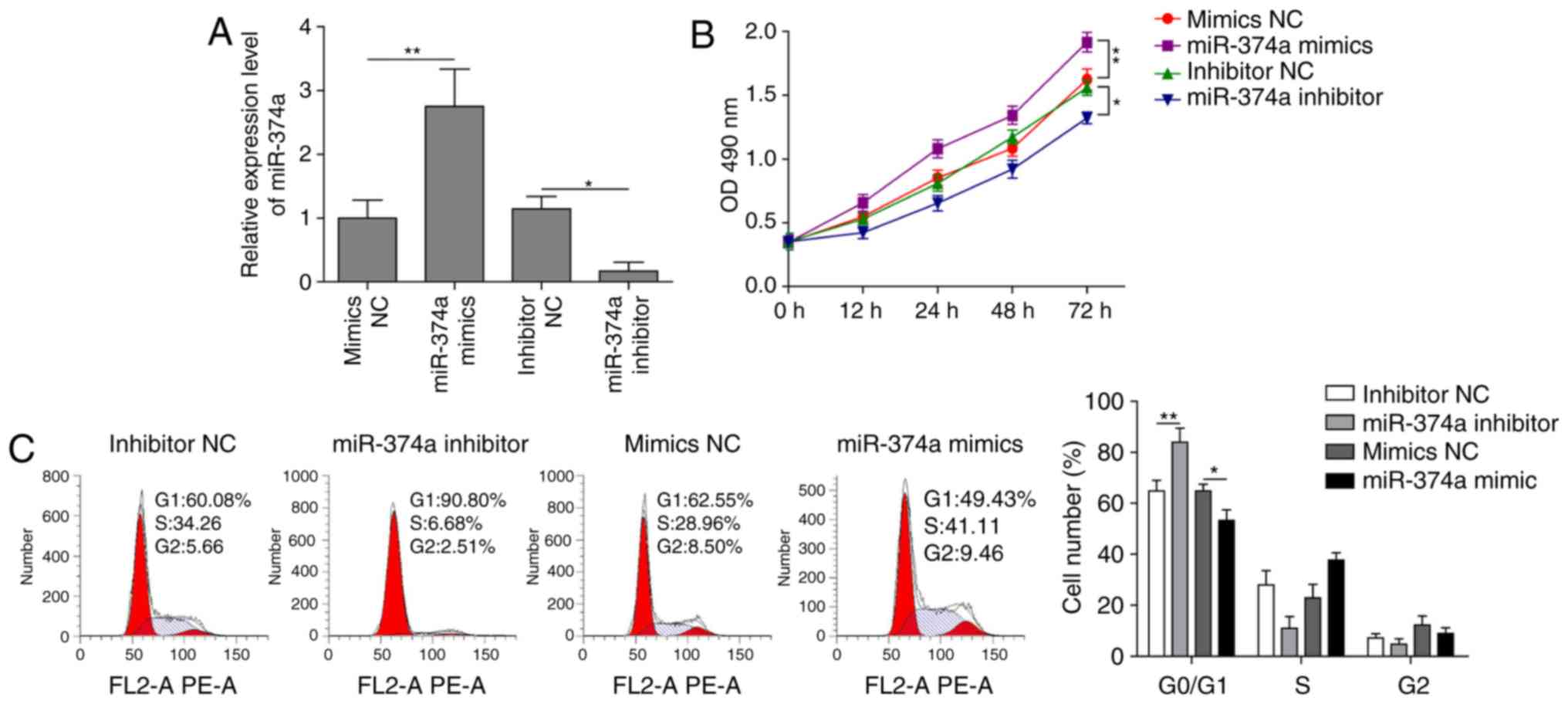
Downregulation of miR-374a decreases
the proliferation of ovarian cancer cells
RT-qPCR data demonstrated that propofol could
downregulate the expression of miR-374a. In order to further
determine the effect of miR-374a on the proliferation of A2780
cells, miR-374a was knocked down using a miR-374a inhibitor, or
overexpressed using miR-374a mimics, and the proliferative ability
of the cells was subsequently analyzed using the CCK-8 assay and
flow cytometry. miR-374a knockdown significantly decreased miR-374a
expression levels, whereas its overexpression significantly
increased miR-374a expression levels compared with their respective
controls (Fig. 3A), thus
demonstrating successful transfection efficiency. The miR-374a
mimics-transfected cells demonstrated significantly enhanced levels
of cell proliferation at 72 h compared with the mimics NC group,
whereas knockdown of miR-374a using the inhibitor significantly
inhibited the proliferation of A2780 cells compared with the NC
group (Fig. 3B). In addition, a
significant increase or decrease in the proportion of cells in the
G0/G1 phase was observed in cells transfected with the inhibitor or
mimics, respectively, compared with their respective NCs (Fig. 3C). Overall, these data revealed
that downregulation of miR-374a using an inhibitor blocked the cell
cycle at the G0/G1 phase and inhibited the proliferation of ovarian
cancer cells.
miR-374a negatively regulates FOXO1 to
promote proliferation and inhibit apoptosis of ovarian cancer
cells
In order to further determine the regulatory
mechanism of miR-374a, the present study explored the downstream
target genes of miR-374a involved in the proliferation of A2780
cells and validated the miRNA-target gene relationships. Target
prediction searches were performed using RAID version 2.0, which
indicated that miR-374a may directly target the 3′UTR of FOXO1
(Fig. 4A). A dual-luciferase
reporter gene assay revealed that the luciferase activity of cells
co-transfected with the miR-374a mimics and wild-type FOXO1 vector
was significantly decreased compared with cells co-transfected with
the mimics NC and wild-type FOXO1 vector (Fig. 4B). This suggested that miR-374a may
bind to the 3′UTR of FOXO1. The overexpression of miR-374a in A2780
cells induced by miR-374a mimics significantly decreased FOXO1 mRNA
and protein expression levels compared with the mimics NC group
(Fig. 4C and D), whereas knockdown
of miR-374a using the miR-374a inhibitor significantly increased
the expression levels of FOXO1 at both the mRNA and protein level
in A2780 cells compared with the inhibitor NC-transfected cells
(Fig. 4C and D). The mRNA and
protein expression levels of FOXO1 were significantly increased in
cells transfected with FOXO1 compared with pcDNA3.1-transfected
cells, indicating that FOXO1 was successfully overexpressed in
cells following the transfection with FOXO1 (Fig. 4E). Moreover, cells with miR-374a
overexpression were observed to have significantly enhanced
proliferation rates and decreased apoptotic rates compared with the
mimics NC group (Fig. 4F and G).
miR-374a overexpression alongside FOXO1 overexpression
significantly inhibited cell proliferation and increased apoptosis
compared with the miR-374a mimics alone (Fig. 4F and G). Therefore, these results
indicated that miR-374a may bind to and negatively regulate FOXO1,
facilitating the proliferation and inhibiting the apoptosis of
A2780 cells.
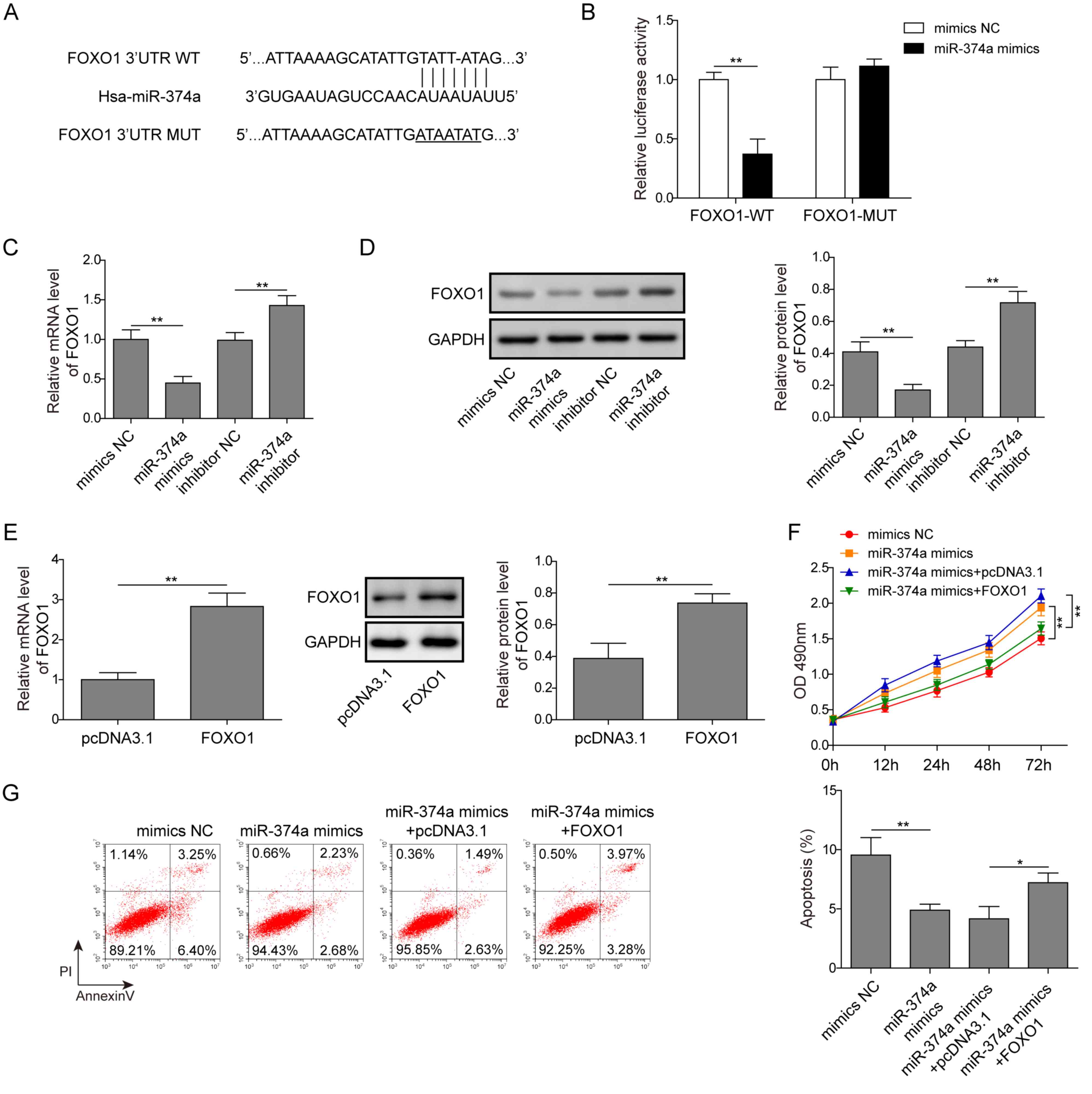 | Figure 4.miR-374a negatively regulates FOXO1
to promote the proliferation and induce the apoptosis of ovarian
cancer cells. (A) A potential binding site of miR-374a in FOXO1 was
predicted. (B) A2780 cells were co-transfected with miR-374a mimics
or mimics NC and luciferase reporter plasmids carrying WT or MUT
FOXO1 3′UTR, and luciferase activity was measured. (C and D) A2780
cells were transfected with mimics NC, miR-374a mimics, inhibitor
NC or miR-374a inhibitor and the relative (C) gene and (D) protein
expression levels of FOXO1 were analyzed using RT-qPCR and western
blotting, respectively. (E) A2780 cells were transfected with empty
pcDNA3.1 plasmid or FOXO1 plasmid, and the relative gene and
protein expression levels of FOXO1 were analyzed using RT-qPCR and
western blotting, respectively. (F and G) A2780 cells were
transfected with mimics NC, miR-374a mimics, miR-374a mimics and
pcDNA3.1, or miR-374a mimics and FOXO1. (F) Cell proliferation was
analyzed using a Cell Counting kit-8 assay. (G) Apoptosis was
detected using flow cytometry. Data are presented as the mean ± SD,
n=3. *P<0.05 and **P<0.01. FOXO1, forkhead box O1; miR,
microRNA; MUT, mutant; NC, negative control; RT-qPCR, reverse
transcription-quantitative PCR; WT, wild-type. |
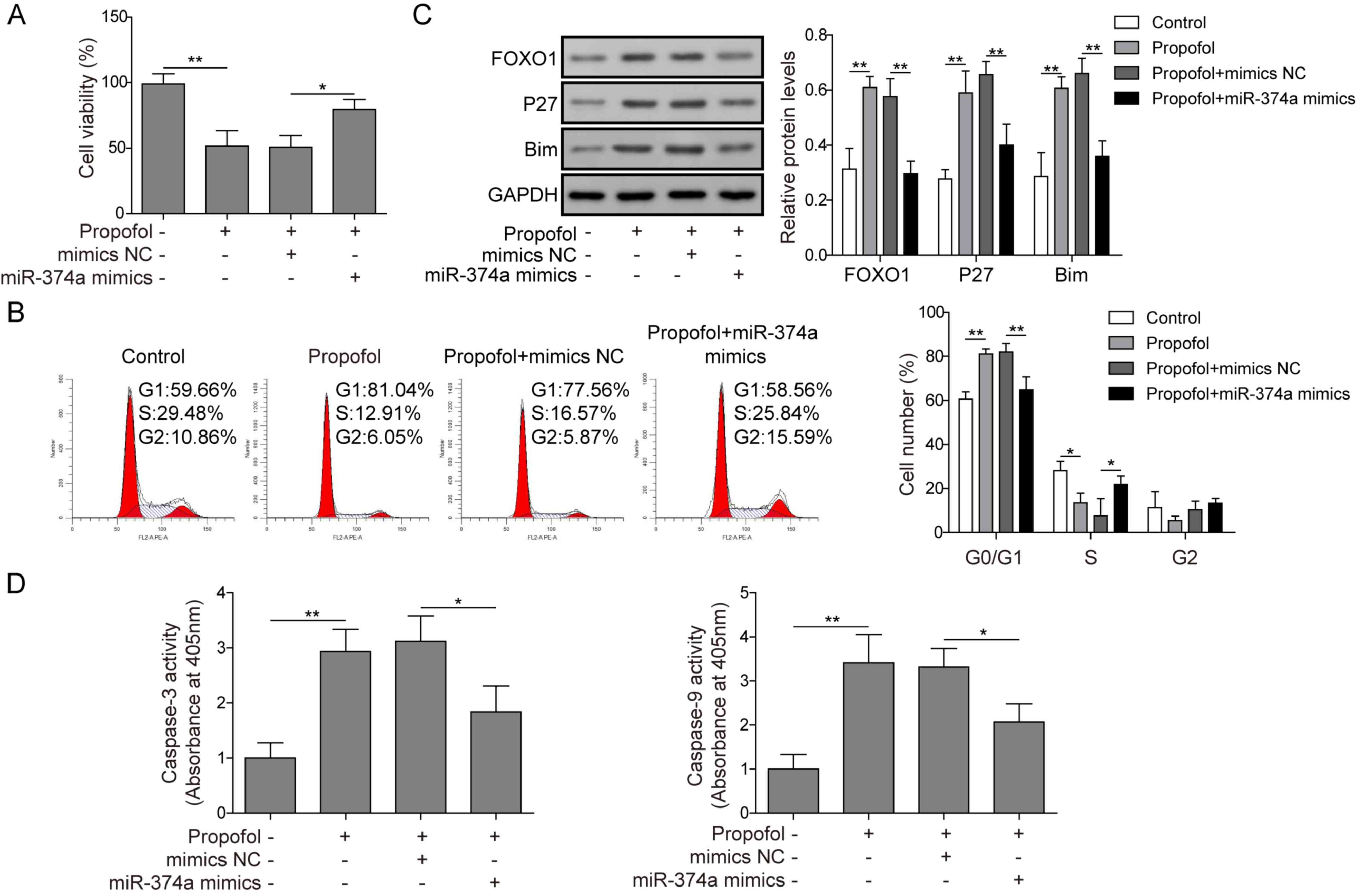
Propofol inhibits proliferation and
accelerates apoptosis of ovarian cancer cells through regulating
miR-374a/FOXO1
To further determine whether propofol inhibited the
proliferation of ovarian cancer cells by modulating the
miR-374a/FOXO1 signaling axis, the effects of propofol alone or
combined with miR-374a overexpression on the cell viability, cell
cycle progression and expression of proteins related to FOXO1
signaling were analyzed. Cell viability was significantly reduced
in the propofol treatment group compared with the control group,
whereas viability was significantly enhanced in the propofol
treatment and miR-374a overexpression group compared with the
propofol treatment and mimics NC group (Fig. 5A). Furthermore, propofol
significantly increased the proportion of cells in the G0/G1 phase
of the cell cycle compared with the control group and decreased the
proportion of cells in the S phase (Fig. 5B), whereas propofol combined with
miR-374a overexpression significantly decreased the proportion of
G0/G1 phase cells compared with the propofol and mimics NC-treated
cells and increased the proportion of S phase cells (Fig. 5B). p27 is a CDK inhibitor that
negatively regulates cell cycle progression (24) and Bim is a pro-apoptotic protein,
the dysregulation of which is closely related to tumorigenesis and
drug resistance (25). Western
blotting was used to analyze the protein expression levels of
FOXO1, p27 and Bim. Following propofol treatment, the expression
levels of FOXO1, p27 and Bim in cells were significantly increased
compared with the control (Fig.
5C). However, following treatment with propofol and the
overexpression of miR-374a, the expression levels of FOXO1, p27 and
Bim in cells were significantly decreased compared with cells
treated with propofol and mimics NC (Fig. 5C). Propofol significantly increased
caspase-3 and caspase-9 activities in cells compared with untreated
cells (Fig. 5D), whereas propofol
combined with miR-374a overexpression significantly decreased
caspase-3 and caspase-9 activities compared with cells treated with
propofol and mimics NC (Fig. 5D).
Taken together, these data further demonstrated that propofol
suppressed proliferation and accelerated apoptosis of ovarian
cancer cells via regulation of the miR-374a/FOXO1 signaling
axis.
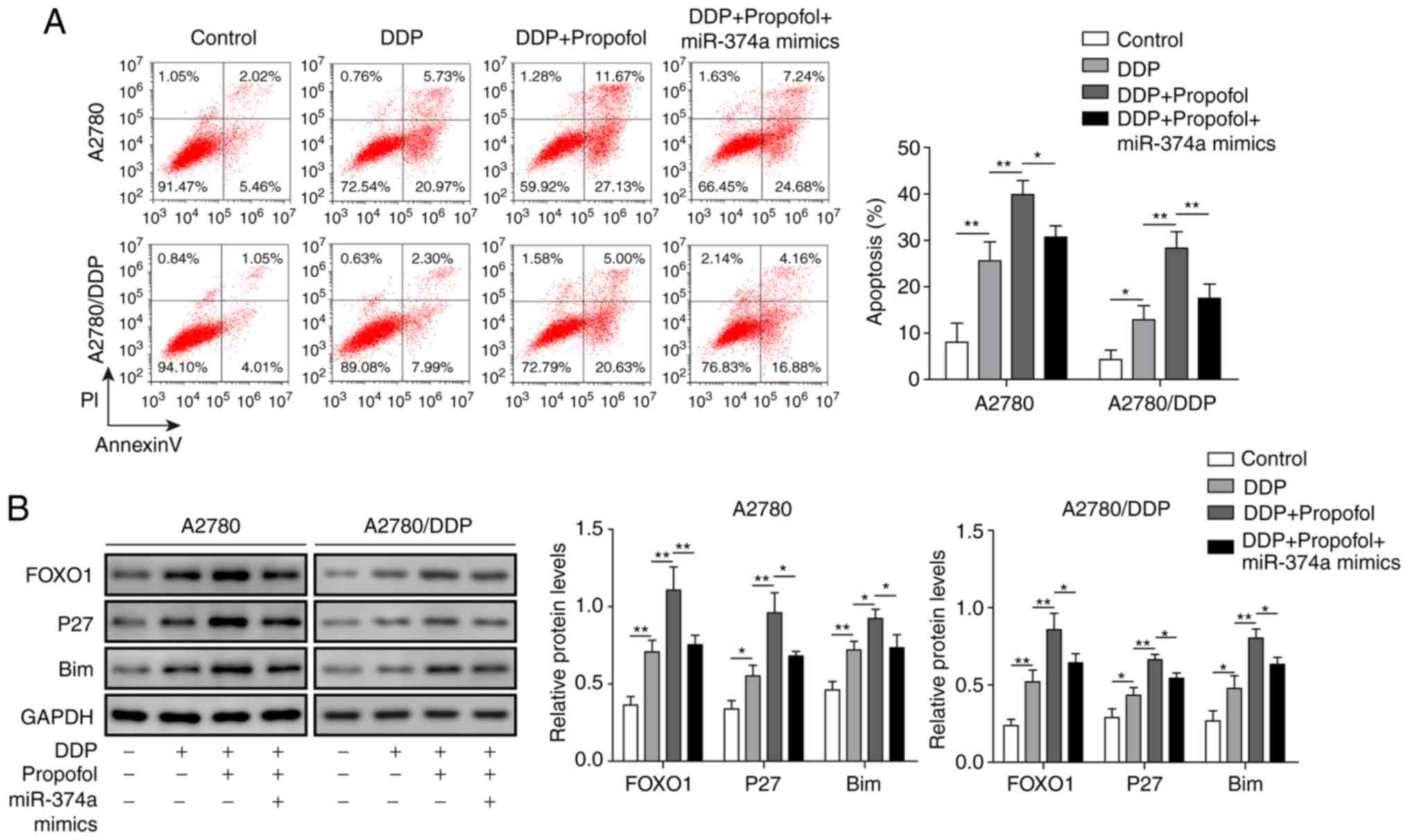
Propofol inhibits DDP resistance in
ovarian cancer cells by regulating the miR-374a/FOXO1 signaling
axis
The molecular signaling mechanisms involved in the
effect of propofol on the drug sensitivity of ovarian cancer cells
were subsequently investigated. Following treatment with propofol
and DDP, the apoptotic rate of A2780 and A2780/DDP cells was
significantly increased compared with DDP only-treated cells;
however, this was significantly reduced following treatment of
cells with propofol, DDP and the miR-374a mimics (Fig. 6A). The protein expression levels of
FOXO1, p27 and Bim in A2780 and A2780/DDP cells were significantly
increased following DDP and propofol pretreatment compared with
cells treated with DDP alone, but were significantly decreased
following simultaneous treatment with propofol, DDP and miR-374a
mimics (Fig. 6B). Overall, these
data revealed that propofol may exert a positive effect by
increasing the sensitivity of ovarian cancer cells to DDP by
inhibiting the expression of miR-374a and subsequently promoting
FOXO1 signaling.
Discussion
Previous studies have demonstrated that ovarian
cancer cells can acquire DDP resistance (26,27);
drug resistance reduces the sensitivity of ovarian cancer cells to
chemotherapeutic drugs, promotes the proliferation and metastasis
of cancer cells, and shortens the survival time of patients
(28). Thus, a detailed study of
the mechanisms underlying drug resistance in ovarian cancer is
required to provide a deeper understanding of the inhibition of
ovarian cancer cells.
Propofol is an intravenous anesthetic that is mainly
used for the induction and maintenance of general anesthesia during
surgery (29,30). However, it has been reported that
propofol may inhibit the proliferation of cancer cells. Zhang et
al (31) demonstrated that
propofol promoted the apoptosis of cervical cancer cells by
regulating the mTOR signaling pathway. Liu et al (32) also concluded that propofol
suppressed the growth, migration and invasion of hepatocellular
carcinoma cells by downregulating miRNA expression. Propofol
enhanced the apoptotic effect of DDP on cervical cancer, which
subsequently increased the sensitivity of cancer cells to the
chemotherapeutic effect of DDP (33), whereas another study reported that
propofol altered the resistance of prostate cancer cells to
docetaxel by inducing hypoxia (34). In addition, Huang et al
(35) observed that propofol
inhibited the growth and invasive ability of ovarian cancer cells
by modulating the miR-9/NF-kB signaling pathway. The results of the
present study suggested that propofol alone inhibited the growth of
ovarian cancer cells and enhanced apoptosis; however, the
mechanisms through which propofol induced apoptosis and inhibited
the proliferation of cancer cells are not well understood. This
study demonstrated that the expression levels of miR-374a were
significantly decreased by propofol in a dose-dependent manner. It
was further observed that overexpression of miR-374a reversed
propofol-induced cell death and the apoptosis of ovarian cancer
cells, suggesting that miR-374a may serve a crucial role in
propofol-induced antitumor effects. DDP is a common
chemotherapeutic drug used in the clinic, which can induce
cytotoxicity or damage ovarian cancer cells; however, owing to the
increased usage of chemotherapeutic drugs, cancer cells mutate and
exhibit reduced sensitivity towards DDP (36). In the present study, it was
discovered that combining propofol with DDP reduced the viability
of DDP-resistant ovarian cancer cells and promoted apoptosis,
demonstrating an overall effect similar to that seen of each drug
alone. Therefore, this suggested that propofol and DDP may exhibit
an additive effect on the cytotoxicity over ovarian cancer
cells.
miR-374a serves an important physiological role in
drug resistance and proliferation of cancer cells; Li et al
(21) discovered that miR-374a is
a DDP-resistant regulator in human ovarian cancer cells and the
expression levels of miR-374a in DDP-resistant ovarian cancer cells
were higher than those in non-DDP-resistant ovarian cancer cells
(19). FOXO1, a vital
transcription factor that mainly participates in cell
proliferation, is closely associated with drug resistance of cancer
cells (37). In the present study,
knockdown of miR-374a using a miR-374a inhibitor inhibited the
growth of ovarian cancer cells and promoted cell cycle arrest at
the G0/G1 phase. In addition, miR-374a was observed to bind to
FOXO1 and negatively regulate FOXO1 expression. He et al
(38) demonstrated that in human
osteosarcoma, miR-374a enhanced cell growth by inhibiting
downstream FOXO1 signaling, which is consistent with the results of
the current study in human ovarian cancer cells. Furthermore, p27
is a CDK inhibitor that has a negative regulatory effect on
proliferation and is a downstream target of FOXO1; consequently,
FOXO1 inhibits cell proliferation by upregulating p27 (39–42).
The Bim protein belongs to the Bcl-2 protein family and exerts a
pro-apoptotic effect on cells (43,44);
previous studies have confirmed that FOXO1 regulates cell apoptosis
through its downstream targets, including Bim (45,46).
The present study confirmed that propofol may regulate
miR-374a/FOXO1 signaling to promote the expression of p27 and Bim,
and to inhibit the proliferation and promote the apoptosis of
ovarian cancer cells, thereby enhancing the sensitivity of ovarian
cancer cells to DDP.
Although the present study revealed a novel
downstream molecular mechanism underlying the apoptosis of ovarian
cancer cells caused by propofol treatment in vitro, further
investigations are required to deeply understand this complex
mechanism. However, in vivo systems are more complex and
often provide more uncertainty than in vitro models; thus,
whether this mechanism could be applied to an animal model is
unknown. Therefore, in the future, this novel molecular mechanism
will require further verification in more cell lines and in
vivo mice models to prove its potential as a therapeutic
target.
In conclusion, the present study demonstrated that
miR-374a negatively regulated FOXO1 expression to promote the
proliferation and inhibit the apoptosis of ovarian cancer cells.
Propofol decreased miR-374a expression to subsequently enhance
FOXO1 expression and inhibit the proliferation of ovarian cancer
cells, thus improving the sensitivity of ovarian cancer cells to
DDP. This provides a theoretical basis for the adjuvant treatment
of ovarian tumors with propofol.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the author on reasonable request.
Authors' contributions
YS and ZC conceived and designed the study, drafted
the manuscript and revised the manuscript. YS, YP, LY, LM and MZ
acquired analyzed and interpreted the data. All the authors
approved publication of the final version and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Grunewald T and Ledermann JA: Targeted
therapies for ovarian cancer. Best Pract Res Clin Obstet Gynaecol.
41:139–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jaaback K, Johnson N and Lawrie TA:
Intraperitoneal chemotherapy for the initial management of primary
epithelial ovarian cancer. Cochrane Database Syst Rev.
1:CD0053402016.
|
|
3
|
Zsiros E, Tanyi J, Balint K and Kandalaft
LE: Immunotherapy for ovarian cancer: Recent advances and
perspectives. Curr Opin Oncol. 26:492–500. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shi C and Wang M: LINC01118 modulates
paclitaxel resistance of epithelial ovarian cancer by regulating
miR-134/ABCC1. Med Sci Monit. 24:8831–8839. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Papaevangelou E, Almeida GS, Box C,
deSouza NM and Chung YL: The effect of FASN inhibition on the
growth and metabolism of a cisplatin-resistant ovarian carcinoma
model. Int J Cancer. 143:992–1002. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen SH and Chang JY: New insights into
mechanisms of Cisplatin resistance: From tumor cell to
microenvironment. Int J Mol Sci. 20:E41362019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jin P, Liu Y and Wang R: STAT3 regulated
miR-216a promotes ovarian cancer proliferation and cisplatin
resistance. Biosci Rep. 38:BSR201805472018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ai Z, Lu Y, Qiu S and Fan Z: Overcoming
cisplatin resistance of ovarian cancer cells by targeting
HIF-1-regulated cancer metabolism. Cancer Lett. 373:36–44. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang C, Gao J, Yan N, Wu B, Ren Y, Li H
and Liang J: Propofol inhibits the growth and survival of gastric
cancer cells in vitro through the upregulation of ING3.
Oncol Rep. 37:587–593. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang N, Liang Y, Yang P, Yang T and Jiang
L: Propofol inhibits lung cancer cell viability and induces cell
apoptosis by upregulating microRNA-486 expression. Braz J Med Biol
Res. 50:e57942017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen X, Li K and Zhao G: Propofol inhibits
HeLa cells by impairing autophagic flux via AMP-activated protein
kinase (AMPK) activation and endoplasmic reticulum stress regulated
by calcium. Med Sci Monit. 24:2339–2349. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu B, Gao W, Zhou H, Miao X, Chang Y, Wang
L, Xu M and Ni G: Propofol induces apoptosis of breast cancer cells
by downregulation of miR-24 signal pathway. Cancer Biomark.
21:513–519. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang P, Chen J, Mu LH, Du QH, Niu XH and
Zhang MY: Propofol inhibits invasion and enhances
paclitaxel-induced apoptosis in ovarian cancer cells through the
suppression of the transcription factor slug. Eur Rev Med Pharmacol
Sci. 17:1722–1729. 2013.PubMed/NCBI
|
|
14
|
Kumar S, Boon RA, Maegdefessel L, Dimmeler
S and Jo H: Role of noncoding RNAs in the pathogenesis of abdominal
aortic aneurysm. Circ Res. 124:619–630. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu B, Li J and Cairns MJ: Identifying
miRNAs, targets and functions. Brief Bioinform. 15:1–19. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma JB, Hu SL, Zang RK, Su Y, Liang YC and
Wang Y: MicroRNA-487a promotes proliferation of esophageal cancer
cells by inhibiting p62 expression. Eur Rev Med Pharmacol Sci.
23:1502–1512. 2019.PubMed/NCBI
|
|
17
|
He H, Zhao X, Zhu Z, Du L, Chen E, Liu S,
Li Q, Dong J, Yang J and Lei L: MicroRNA-3191 promotes migration
and invasion by downregulating TGFBR2 in colorectal cancer. J
Biochem Mol Toxicol. e223082019.doi: 10.1002/jbt.22308 (Epub ahead
of print). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu H, Liu Y, Shu XO and Cai Q: MiR-374a
suppresses lung adenocarcinoma cell proliferation and invasion by
targeting TGFA gene expression. Carcinogenesis. 37:567–575. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Goto T, Takano M, Hirata J and Tsuda H:
The involvement of FOXO1 in cytotoxic stress and drug-resistance
induced by paclitaxel in ovarian cancers. Br J Cancer.
98:1068–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang J, Yang H, Li W, Xu H, Yang X and Gan
L: Thioredoxin 1 upregulates FOXO1 transcriptional activity in drug
resistance in ovarian cancer cells. Biochim Biophys Acta.
1852:395–405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li N, Yang L, Wang H, Yi T, Jia X, Chen C
and Xu P: MiR-130a and MiR-374a function as novel regulators of
cisplatin resistance in human ovarian cancer A2780 cells. PLoS One.
10:e01288862015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang L, Hu C, Chao H, Wang R, Lu H, Li H
and Chen H: miR-29c regulates resistance to paclitaxel in
nasopharyngeal cancer by targeting ITGB1. Exp Cell Res. 378:1–10.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yue ZX, Gao RQ, Gao C, Liu SG, Zhao XX,
Xing TY, Niu J, Li ZG, Zheng HY and Ding W: The prognostic
potential of coilin in association with p27 expression in pediatric
acute lymphoblastic leukemia for disease relapse. Cancer Cell Int.
18:1062018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ajabnoor GM, Crook T and Coley HM:
Paclitaxel resistance is associated with switch from apoptotic to
autophagic cell death in MCF-7 breast cancer cells. Cell Death Dis.
3:e2602012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gong S, Chen Y, Meng F, Zhang Y, Wu H, Li
C and Zhang G: RCC2, a regulator of the RalA signaling pathway, is
identified as a novel therapeutic target in cisplatin-resistant
ovarian cancer. FASEB J. 33:5350–5365. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li X, Zou Z, Tang J, Zheng Y, Liu Y, Luo
Y, Liu Q and Wang Y: NOS1 upregulates ABCG2 expression contributing
to DDP chemoresistance in ovarian cancer cells. Oncol Lett.
17:1595–1602. 2019.PubMed/NCBI
|
|
28
|
Norouzi-Barough L, Sarookhani MR, Sharifi
M, Moghbelinejad S, Jangjoo S and Salehi R: Molecular mechanisms of
drug resistance in ovarian cancer. J Cell Physiol. 233:4546–4562.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chidambaran V, Costandi A and D'Mello A:
Propofol: A review of its role in pediatric anesthesia and
sedation. CNS Drugs. 29:543–563. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fan W, Zhu X, Wu L, Wu Z, Li D, Huang F
and He H: Propofol: An anesthetic possessing neuroprotective
effects. Eur Rev Med Pharmacol Sci. 19:1520–1529. 2015.PubMed/NCBI
|
|
31
|
Zhang D, Zhou XH, Zhang J, Zhou YX, Ying
J, Wu GQ and Qian JH: Propofol promotes cell apoptosis via
inhibiting HOTAIR mediated mTOR pathway in cervical cancer. Biochem
Biophys Res Commun. 468:561–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu SQ, Zhang JL, Li ZW, Hu ZH, Liu Z and
Li Y: Propofol inhibits proliferation, migration, invasion and
promotes apoptosis through down-regulating miR-374a in
hepatocarcinoma cell lines. Cell Physiol Biochem. 49:2099–2110.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li H, Lu Y, Pang Y, Li M, Cheng X and Chen
J: Propofol enhances the cisplatin-induced apoptosis on cervical
cancer cells via EGFR/JAK2/STAT3 pathway. Biomed Pharmacother.
86:324–333. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qian J, Shen S, Chen W and Chen N:
Propofol reversed hypoxia-induced docetaxel resistance in prostate
cancer cells by preventing epithelial-mesenchymal transition by
inhibiting hypoxia-inducible factor 1α. Biomed Res Int.
2018:41742322018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang X, Teng Y, Yang H and Ma J: Propofol
inhibits invasion and growth of ovarian cancer cells via regulating
miR-9/NF-κB signal. Braz J Med Biol Res. 49:e57172016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao C, Lu E, Hu X, Cheng H, Zhang JA and
Zhu X: S100A9 regulates cisplatin chemosensitivity of squamous
cervical cancer cells and related mechanism. Cancer Manag Res.
10:3753–3764. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
He W, Feng L, Xia D and Han N: MiR-374a
promotes the proliferation of human osteosarcoma by downregulating
FOXO1 expression. Int J Clin Exp Med. 8:3482–3489. 2015.PubMed/NCBI
|
|
39
|
Xu Y, Bei Y, Shen S, Zhang J, Lu Y, Xiao J
and Li X: MicroRNA-222 promotes the proliferation of pulmonary
arterial smooth muscle cells by targeting P27 and TIMP3. Cell
Physiol Biochem. 43:282–292. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Roy A and Banerjee S: p27 and leukemia:
Cell cycle and beyond. J Cell Physiol. 230:504–509. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li Y, Deng S, Peng J, Wang X, Essandoh K,
Mu X, Peng T, Meng ZX and Fan GC: MicroRNA-223 is essential for
maintaining functional β-cell mass during diabetes through
inhibiting both FOXO1 and SOX6 pathways. J Biol Chem.
294:10438–10448. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Suzuki R, Amatya VJ, Kushitani K, Kai Y,
Kambara T and Takeshima Y: MiR-182 and miR-183 promote cell
proliferation and invasion by targeting FOXO1 in mesothelioma.
Front Oncol. 8:4462018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li Y, Deutzmann A and Felsher DW:
BIM-mediated apoptosis and oncogene addiction. Aging (Albany NY).
8:1834–1835. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li L, Wang C, Wen Y, Hu Y, Xie Y, Xu M,
Liang M, Liu W, Liu L and Wu Y: ERK1/2 and the Bcl-2 family
proteins Mcl-1, tBid, and Bim are involved in inhibition of
apoptosis during persistent chlamydia psittaci infection.
Inflammation. 41:1372–1383. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Huang C, Chen D, Zhu H, Lv S, Li Q and Li
G: LITAF enhances radiosensitivity of human glioma cells via the
FoxO1 pathway. Cell Mol Neurobiol. 39:871–882. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qin Y, Li L, Wang F, Zhou X, Liu Y, Yin Y
and Qi X: Knockdown of Mir-135b sensitizes colorectal cancer cells
to oxaliplatin-induced apoptosis through increase of FOXO1. Cell
Physiol Biochem. 48:1628–1637. 2018. View Article : Google Scholar : PubMed/NCBI
|