Introduction
Lung cancer has become the most common malignant
tumor worldwide, contributing to the highest mortality and
morbidity rate compared with all cancer types (1). Non-small-cell lung cancer (NSCLC)
accounts for >80% of all lung cancer cases (1). Patients with NSCLC typically have a
poor prognosis, which is primarily due to distant metastasis and
local recurrence (2). Several
patients with NSCLC are diagnosed at the late stages with local
infiltration and distant metastases; at which point, surgery is not
an effective treatment option. Therefore, identifying novel targets
to aid with the early diagnosis of NSCLC is required for improving
the efficacy of clinical treatment.
The Warburg effect is a phenomenon that cancer cells
rely on to sustain their proliferation by increasing the uptake of
glucose and the production of lactate. This phenomenon is a unique
energy metabolism present in cancer cells (3). Increasing evidence suggests that
enzymes involved in this process might be potential targets for
cancer therapy (4). Pyruvate
kinase (PK) is a key rate-limiting enzyme involved in tumor
glycolysis, which catalyzes the conversion of phosphoenolpyruvic
acid and ADP into pyruvate acid and ATP (5). In humans, four isoforms of PK have
been identified, including red blood cell PK, liver-type PK, and PK
muscle isozyme M1 and M2 (PKM1 and PKM2, respectively) (6). PKM2 has been reported to be
upregulated in tumor cells and cancer stem cells, playing a role in
glycolysis and tumor malignancy (7). Furthermore, a number of previous
studies have indicated that glycolysis is closely related to cell
proliferation and metastasis in NSCLC (8–11).
Long non-coding RNA (lncRNA) brain cytoplasmic RNA 1
(BCYRN1) is associated with the progress of numerous diseases
(12). In patients with
Alzheimer's disease, neurons display high levels of BCYRN1
expression, which regulates cell viability and apoptosis by
targeting β-site amyloid precursor protein-cleaving enzyme 1
(13). BCYRN1 also acts as either
an oncogene or a tumor suppressor in numerous types of human
cancer, including cervical, colorectal, breast and ovarian cancer,
as well as esophageal squamous cell carcinoma (14). During NSCLC, BCYRN1 enhances cell
migration and invasion by upregulating matrix metalloproteinases
(12). BCYRN1 has also been
identified as an oncogene in NSCLC (12); however, the mechanisms of action of
BCYRN1 are not completely understood.
The aim of the present study was to investigate the
role of BCYRN1 in NSCLC glycolysis. BCYRN1 was highly expressed in
NSCLC tissues and cells compared with the corresponding controls.
The function of BCYRN1 on the Warburg effect in NSCLC cell lines
was examined. Mechanistically, the results suggested that BCYRN1
induced cell glycolysis and tumor progression via the microRNA
(miRNA/miR)-149/PKM2 signaling pathway.
Materials and methods
Clinical samples
Written informed consent was obtained from patients
with NSCLC (male patients, 12 males; female patients, 8; mean age,
37.6 years; age range, 25–48 years) admitted to The Fourth
Affiliated Hospital of Harbin Medical University (Harbin, China)
between March 2017 and December 2018. Subsequently, 20 primary
NSCLC and paired normal tissues were collected, snap-frozen and
stored at −80°C. The matched ‘normal tissue’ was obtained from a 5
cm distance from the tumor margin, which were confirmed by a
pathologist to not contain tumor cells. The inclusion criteria were
as follows: i) Patients diagnosed with lung adenocarcinoma or lung
squamous carcinoma by histopathological examination; ii) tumors
identified as TNM stage I or IIIa; and iii) patients were subjected
to radical operation, and had not received systemic chemotherapy or
radiotherapy prior to surgery (15). Patients with autoimmune disease or
incomplete case data were excluded. The present study was approved
by the Clinical Research Ethics Committee of The Fourth Affiliated
Hospital of Harbin Medical University.
Cell lines and transfection
Human NSCLC cell lines (A549, H460 and H1299) and
the normal human bronchial epithelial cell line 16HBE were obtained
from the American Type Culture Collection and were cultured in DMEM
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2.
miR-149 mimic, pre-negative control (NC), miR-149
inhibitor, NC, small interfering (si)RNA targeting BCYRN1
(si-BCYRN1) or targeting PKM2 (si-PKM2), and si-NC were synthesized
by Guangzhou RiboBio Co., Ltd. The corresponding sequences were as
follows: miR-149 mimic, 5′-UCUGGCUCCGUGUCUUCACUCCC-3′; pre-NC,
5′-UUCUCCGAACGUGUCACGU-3′; miR-149 inhibitor,
5′-GGGAGUGAAGACACGGAGCCAGA-3′; NC, 5′-CAGUACUUUUGUGUAGUACAA-3′;
si-BCYRN1, 5′-CUCCAGAAAAAGGAAAAAAAAAA-3′; si-PKM2,
5′-GGAAAGAACAUCAAGAUUAUC-3′; si-NC, 5′-UUCUCCGAACGUGUCACGU-3′.
Plasmid pcDNA-BCYRN1 or pcDNA-PKM2 were constructed by Shanghai
GenePharma Co., Ltd. for the overexpression of BCYRN1 or PKM2 in
cells. A459 or H1299 cells (2×105) were transfected with
50 nM plasmid or 50 nM RNAs using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. After a 48-h incubation at 37°C, cells
were harvested for subsequent experiments. An empty plasmid was
used as the negative control.
Cell Counting kit-8 (CCK-8) assay
Transfected A549 cells (1×104 cells/well)
were seeded in 96-well plates. After a 48-h culture, CCK-8 solution
(Dojindo Molecular Technologies, Inc.) (10 µl) was added into each
well and incubated for 4 h at 37°C. The absorbance was measured at
a wavelength of 450 nm by a microplate reader to analyze cell
proliferation. The cell survival rate=[(Aexperiment
group-Ablank)/(Acontrol group-Ablank)] ×100%
Matrigel invasion assay
Transfected A549 cells (2.5×103
cells/well) in serum-free DMEM were seeded into the upper chamber
of Transwell inserts pre-coated with Matrigel. DMEM supplemented
with 10% FBS was plated in the lower chamber. After incubation for
24 h at 37°C, the invasive cells were stained with 0.3% crystal
violet at room temperature for 20 min. Subsequently, the number of
invading cells in the lower chamber were counted using an inverted
fluorescence microscope IX71 (magnification, ×100).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the cells or tissues
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. After
quantification using a spectrophotometer, total RNA was reverse
transcribed into cDNA using the PrimeScript RT Reagent kit (Takara
Bio, Inc.), according to the manufacturer's protocol. The
conditions for RT were as follows: 70°C for 3 min, 42°C for 60 min
and 70°C for 15 min. Subsequently, qPCR was performed using the
SYBR Green kit (Takara Bio, Inc.) on an ABI 7900 system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The primer pairs that
were used for qPCR are presented in Table I. The following thermocycling
conditions were used for qPCR: Initial denaturation at 95°C for 30
sec, followed by 40 cycles at 95°C for 5 sec and at 60°C for 45
sec. Relative expression levels were quantified using the
2−ΔΔCq method (16) and
GAPDH or U6 was used as the internal reference gene for mRNA and
miRNA, respectively.
 | Table I.Primer sequences used for reverse
transcription--quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription--quantitative PCR.
| Gene | Primer sequences
(5′-3′) |
|---|
| BCYRN1 | F:
GCCTGTAATCCCAGCTCTCA |
|
| R:
GGGTTGTTGCTTTGAGGGAA |
| microRNA-149 | F:
AGCGCGUCUGGCUCCGUGUCUUC |
|
| R:
ATCCAGTGCAGGGTCCGAGG |
| PKM2 | F:
GCACACCGTATTCAGCTCTG |
|
| R:
TCCAGGAATGTGTCAGCCAT |
| GAPDH | F:
ACCACAGTCCATGCCATCAC |
|
| R:
TCCACCACCCTGTTGCTGTA |
| U6 | F:
GCTTCGGCAGCACATATACTAAAAT |
|
| R:
TACTGTGCGTTTAAGCACTTCGC |
Western blotting
Total protein was extracted from A549 and H1299
cells using the IP cell lysis buffer (Pierce; Thermo Fisher
Scientific, Inc.). Bicinchoninic acid protein assay kit (Takara
Biotechnology Co., Ltd.) was used to quantify protein
concentration. Proteins (30 µg/lane) were separated by SDS-PAGE on
10% gels. Subsequently, the separated proteins were transferred to
PVDF membranes (EMD Millipore) and blocked in 5% milk at 37°C for 2
h. The membranes were incubated overnight at 4°C with the primary
antibodies targeted against: PKM2 (1:1,000; cat. no. ab137852;
Abcam) and β-actin (1:2,000; cat. no. ab8227; Abcam). Following the
primary antibody incubation, the membranes were incubated with a
horseradish peroxidase conjugated secondary antibody (1:2,000; cat.
no. ab7090; Abcam) for 2 h at room temperature. Protein bands were
visualized using the enhanced chemiluminescence reagent (EMD
Millipore).
Glucose consumption and lactate
production assay
Following transfection, the culture media and the
cells were collected separately. A Glucose assay kit
(Sigma-Aldrich; cat. no. GAHK20; Merck KGaA) was used to measure
the glucose levels in the culture media according to the
manufacturer's protocol. Lactate levels in the culture media were
determined using a Lactic Acid assay kit (Biovision, Inc.; cat. no.
K606-100), according to the manufacturer's protocol.
Dual-luciferase reporter assay
The potential miRNAs that bind to BCYRN1 were
predicted using DIANA (diana.imis.athena-innovation.gr/DianaTools/index.php)
and the potential binding site between miR-149 and PKM2 was
predicted using Microrna (http://www.microrna.org/microrna/home.do). To
investigate the interaction between BCYRN1 and miR-149, and between
miR-149 and PKM2, the wild-type (WT) or mutant (MUT) BCYRN1
sequences and 3′-untranslated regions (3′-UTRs) of wild-type (WT)
or mutant (MUT) PKM2, were synthesized by Guangzhou RiboBio Co.,
Ltd. H1299 cells (2×105) were co-transfected with the
firefly luciferase reporter plasmid pmirGLO (Promega Corporation)
and 50 nM miR-149 mimic or pre-NC using Lipofectamine®
3000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. After a 48-h incubation at room
temperature, the luciferase activity was detected using a
Dual-Luciferase Reporter assay system (Promega Corporation),
according to the manufacturer's protocol. Firefly luciferase
activity was normalized to Renilla luciferase activity.
Statistical analysis
Statistical analyses were performed using SPSS
software (version 23; IBM Corp.). Data are presented as the mean ±
SD from three replicates. A paired Student's t-test was used to
assess statistical differences for the BCYRN1 express level between
tumor and adjacent healthy tissues. An unpaired Student's t-test or
one-way ANOVA followed by Tukey's post hoc test was used to assess
statistical differences between two groups or multiple groups,
respectively. P<0.05 was considered to indicate a statistically
significant difference.
Results
BCYRN1 induces glycolysis and
increases the expression levels of PKM2 in NSCLC cells
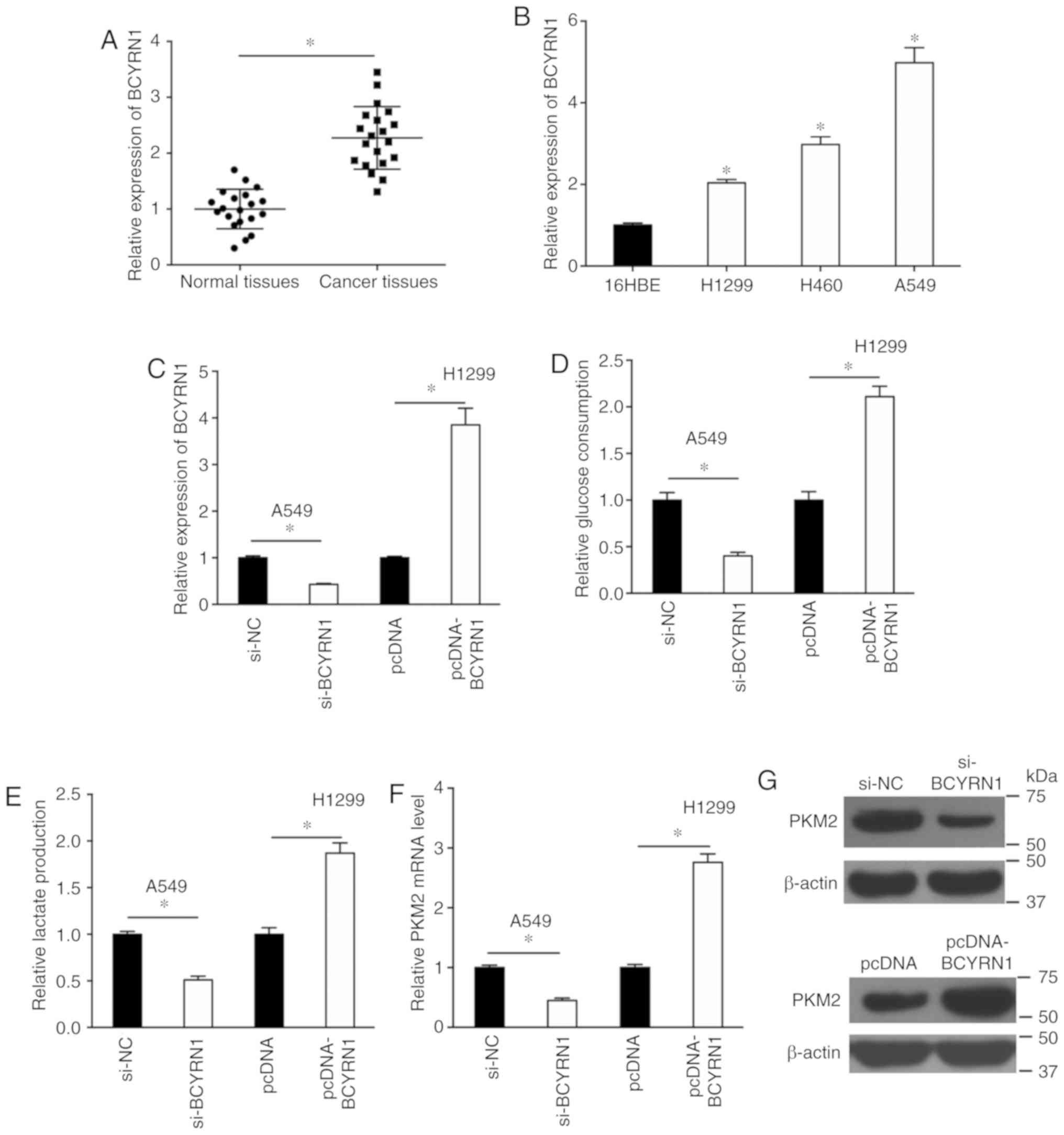
The expression of BCYRN1 in NSCLC tissues was
detected by RT-qPCR and the results suggested that BCYRN1
expression was significantly higher in tumor tissues compared with
adjacent normal tissues (Fig. 1A).
Additionally, BCYRN1 was significantly upregulated in a number of
NSCLC cell lines, including A549, H460 and H1299 cells, compared
with 16HBE cells (Fig. 1B). The
expression of BCYRN1 was significantly reduced by 57% in A549 cells
and increased by 3.85-fold in H1299 cells following transfection
with si-BCYRN1 or pcDNA-BCYRN1, respectively, compared with the
corresponding control (Fig. 1C).
Glucose consumption and lactate production assays were performed to
examine the effects of BCYRN1 on glycolysis. BCYRN1 knockdown
significantly decreased glucose consumption by 60% and lactate
production by 49% in A549 cells, and BCYRN1 overexpression
significantly increased glucose consumption and lactate production
in H1299 cells, compared with the corresponding controls (Fig. 1D and E). The mRNA and protein
expression levels of PKM2 were downregulated in A549 cells
following BCYRN1 knockdown, and conversely, BCYRN1 overexpression
upregulated the expression levels of PKM2 in H1299 cells, compared
with the corresponding controls (Fig.
1F and G).
BCYRN1 regulates miR-149 expression
levels during glycolysis
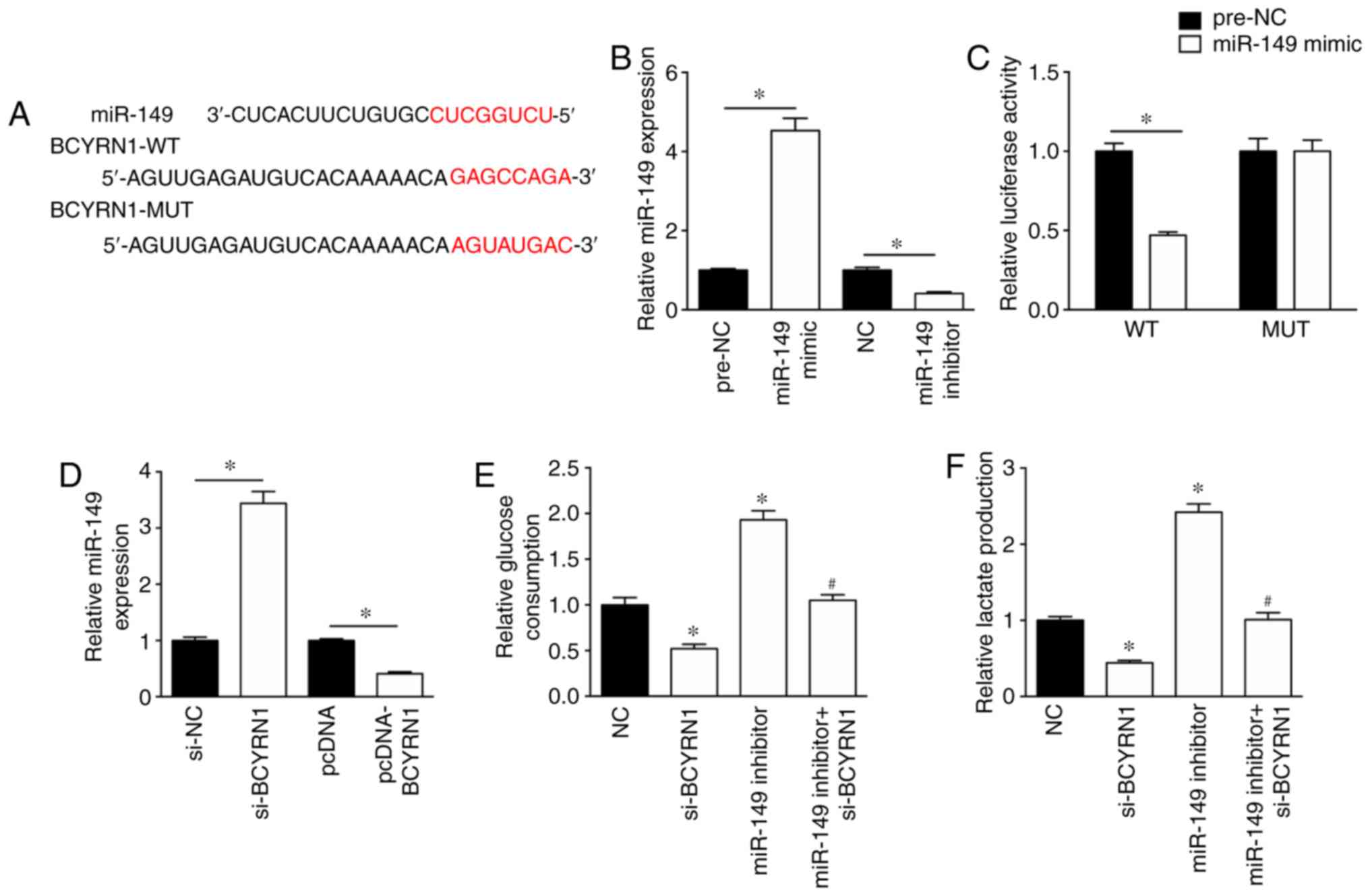
To investigate the mechanism underlying the role of
BCYRN1 during glycolysis, the potential miRNAs that bind to BCYRN1
were predicted using DIANA (diana.imis.athena-innovation.gr/DianaTools/index.php).
In silico results suggested that the sequence of BCYRN1
contained a possible miR-149 binding site (Fig. 2A). The miR-149 mimic significantly
enhanced the expression levels of miR-149 in A549 cells by
4.53-fold, whereas the expression levels of miR-149 were
significantly reduced by 41% in A549 cells transfected with miR-149
inhibitor, compared with the corresponding controls (Fig. 2B). In addition, the miR-149 mimic
inhibited the luciferase activity of the BCYRN1-WT luciferase
plasmid, but had no effect on the luciferase activity of the
BCYRN1-MUT luciferase plasmid in A549 cells (Fig. 2C). RT-qPCR was performed to assess
the effect of BCYRN1 on miR-149 expression. The results suggested
that miR-149 expression levels were significantly increased by
3.44-fold in A549 cells transfected with si-BCYRN1 compared with
cells transfected with si-NC (Fig.
2D). Additionally, miR-149 expression levels were significantly
downregulated in A549 cells following BCYRN1 overexpression
compared with cells transfected with the empty pcDNA vector
(Fig. 2D). The results of the
glucose consumption and lactate production assays suggested that
the miR-149 inhibitor rescued si-BCYRN1-induced inhibition of
glucose consumption and lactate production, compared with the
si-BCYRN1 (Fig. 2E and F).
miR-149 regulates the expression of
PKM2
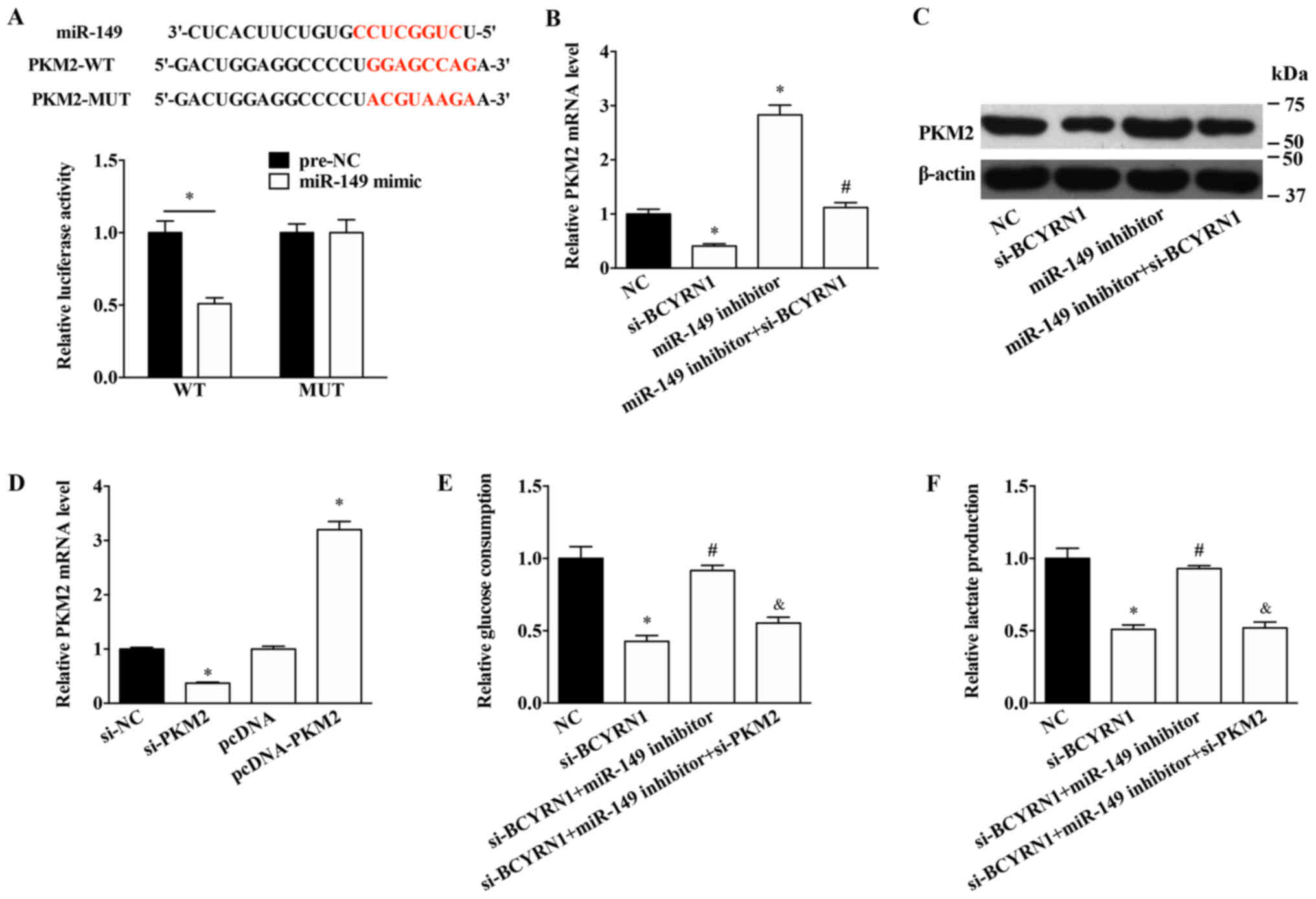
The potential mechanism underlying the effects of
BCYRN1 and miR-149 in A549 cells was further investigated. miR-149
was predicted to target the 3′-UTR of PKM2 (Fig. 3A). Notably, miR-149 overexpression
significantly inhibited the luciferase activity of the PKM2-WT
plasmid, but had no effect on the luciferase activity of the
PKM2-MUT plasmid (Fig. 3A).
miR-149 knockdown significantly upregulated the expression levels
of PKM2 and rescued the decreased expression levels of PKM2 induced
by si-BCYRN1 (Fig. 3B and C). To
further explore the role of PKM2 in BCYRN1/miR-149 function,
si-PKM2 was used to inhibit the expression of PKM2 and pcDNA-PKM2
was used to overexpress PKM2 in A549 cells (Fig. 3D). PKM2 knockdown significantly
reversed the effects of the miR-149 inhibitor on glucose
consumption and lactate production (Fig. 3E and F).
PKM2 is associated with the function
of BCYRN1 in A549 cells
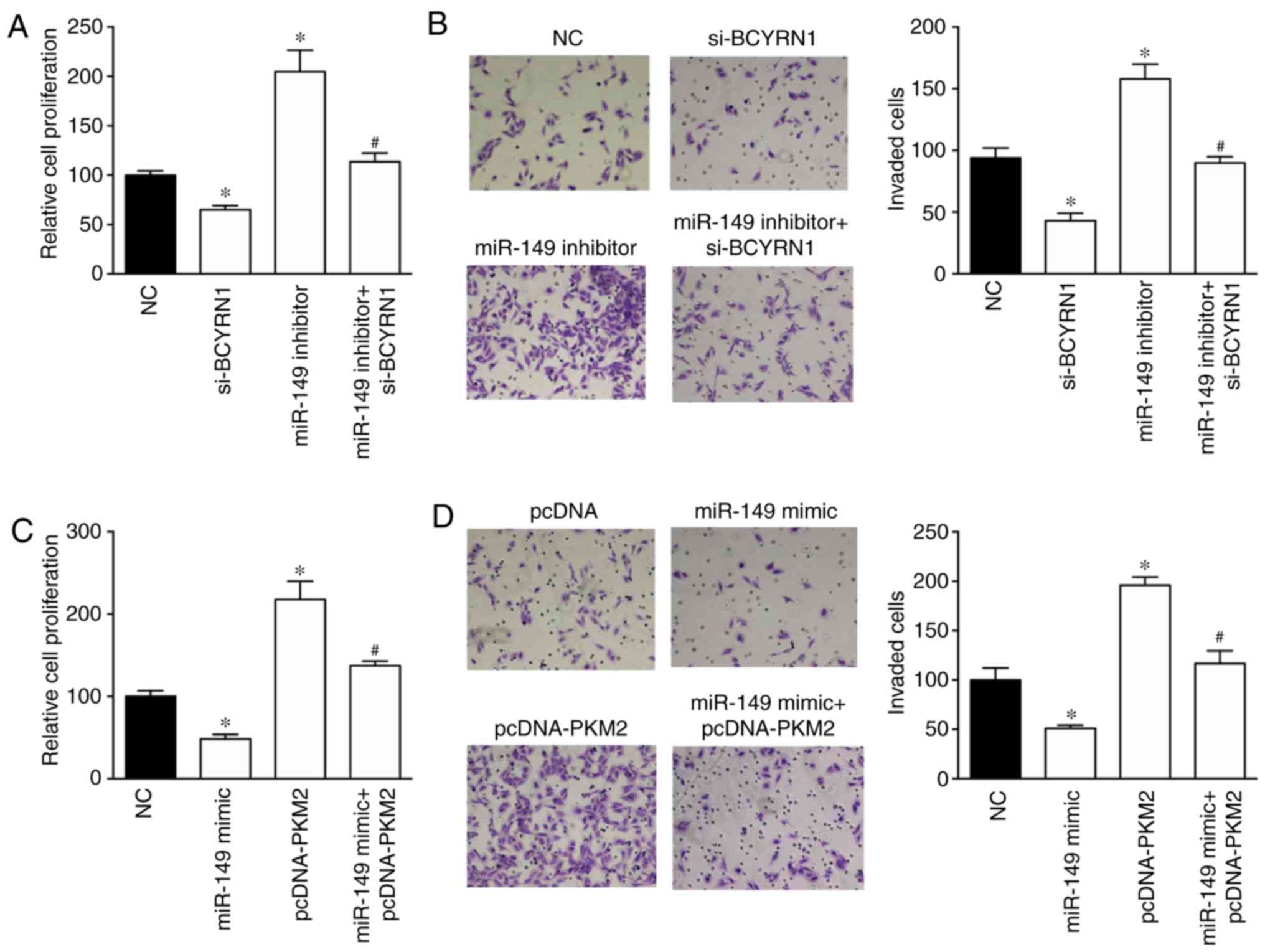
The effects of the BCYRN1/miR-149/PKM2 regulatory
pathway on NSCLC cell proliferation and invasion were investigated
using the CCK-8 and invasion assays. Cell proliferation and
invasion were inhibited by BCYRN1 knockdown and enhanced by the
miR-149 inhibitor in A549 cells (Fig.
4A and B). Notably, the miR-149 inhibitor rescued the
si-BCYRN1-induced inhibition of cell proliferation and invasion.
Furthermore, PKM2 overexpression induced a 2.17- and 1.96-fold
increase in proliferation and invasion in A549 cells compared with
NC group, respectively. In addition, PKM2 overexpression reversed
the miR-149 mimic-induced inhibition of cell proliferation and
invasion (Fig. 4C and D).
Discussion
Aerobic glycolysis, as one of the ‘hallmarks of
cancer’, converts glucose into pyruvate, which is then further
oxidized into lactate (17). The
resulting lactate contributes to destruction of the extracellular
matrix and provides a metastatic advantage for cancer cells,
leading to rapid proliferation and resistance to apoptosis
(18). Aerobic glycolysis during
cancer has attracted increasing attention in recent years, as it
has been suggested that targeting glycolysis might be a promising
treatment for cancer (19–21). Emerging data suggests that
glycolysis could be regulated by certain lncRNAs in cancer. For
example, Li et al (22)
reported that urothelial cancer-associated 1 was involved in the
proliferation and glycolysis of bladder cancer cells via the
induction of hexokinase 2 (HK2) expression. c-Myc-activated lncRNA
macrophage migration inhibitory factor inhibited aerobic glycolysis
and tumorigenesis in cancer cells by downregulating c-Myc and
upregulating F-box and WD repeat domain containing 7 expression
(23). Furthermore, Pvt1 oncogene
has been reported to promote osteosarcoma cell glucose metabolism,
cell proliferation and motility via the miR-497/HK2 signaling
pathway (24).
In the present study, increased glucose consumption
and lactate production was observed in NSCLC cells overexpressing
BCYRN1. Additionally, higher expression levels of BCYRN1 were
observed in NSCLC tissues and cell lines compared with normal
control tissues and cell lines. Consistent with these results, Hu
and Lu (12) also reported that
BCYRN1 acted as an oncogene in NSCLC as evidenced by the high cell
proliferation and metastasis induced by BCYRN1. BCYRN1 has been
reported to function as an oncogene in colorectal and cervical
cancer, as well as in gastric carcinoma (25–27).
Mechanistically, BCYRN1 may regulate gene expression by targeting
miRNAs. For example, BCYRN1 targeted miR-138 or miR-150 to alter
the proliferation and metastasis of cervical cancer cells or airway
smooth muscle cells, respectively (25,28).
To further investigate the molecular mechanisms
underlying the effects of BCYRN1 on NSCLC, bioinformatics analysis
was performed to identify the potential targets of BCYRN1 and
suggested that miR-149 might be a target of BCYRN1. The results of
the luciferase activity assay further suggested that BCYRN1
interacted with miR-149 and regulated the expression of miR-149 in
NSCLC cells. In addition, the miR-149 inhibitor rescued the
si-BCYRN1-induced effects on glucose consumption, lactate
production and cell proliferation and invasion. miR-149 has been
identified as a tumor suppressor in several types of cancer,
including gastric cancer, and renal cell and colonic carcinoma
(29–31). Zhao et al (32) reported that miR-149 decreased human
NSCLC growth and metastasis by inhibiting the forkhead box
M1/cyclin D1/matrix metallopeptidase 2 axis. Furthermore, in
cervical cancer cells, miR-149 regulated proliferation and
apoptosis via regulating GIT ArfGAP1 (33).
The coding gene downstream of the BCYRN1/miR-149
axis was further investigated in NSCLC cells. Finally, PKM2 was
identified as a potential target of miR-149 and was upregulated by
the miR-149 inhibitor. PKM2, as the critical isoenzyme of PK,
serves a role in maintaining the metabolic program of cancer cells
(34). Inhibition of PKM2
expression was previously reported to decrease glucose consumption
and lactate production (35), and
elevated expression of PKM2 has been reported in colon and lung
cancer (36,37). Previous studies have suggested that
PKM2 contributes to cancer metabolism by promoting
hypoxia-inducible factor-1 expression or coordinating with c-Myc
(38,39). In the present study, PKM2 knockdown
significantly reversed the miR-149 inhibitor-induced effects on
glucose consumption and lactate production. PKM2 overexpression
also reversed the miR-149 mimic-induced reduction of cell
proliferation and invasion.
Collectively, the present study suggested that
BCYRN1 participated in glycolysis during NSCLC via regulating the
miR-149/PKM2 axis, and accelerating cell proliferation and
invasion. The results suggested that the BCYRN1/miR-149/PKM2
signaling pathway was related to regulation of the Warburg effect
and therefore might be a potential therapeutic target for
NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
NL, CW and JZ designed the study. NL performed the
experiments and wrote the paper. CW performed the statistical
analyses. FS, TW and HAC performed the cell experiments.
Ethics approval and consent to
participate
The present study was approved by the Clinical
Research Ethics Committee of the Fourth Affiliated Hospital of
Harbin Medical University. All patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jamal-Hanjani M, Wilson GA, McGranahan N,
Birkbak NJ, Watkins TBK, Veeriah S, Shafi S, Johnson DH, Mitter R,
Rosenthal R, et al: Tracking the evolution of non-small-cell lung
cancer. N Engl J Med. 376:2109–2121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Detterbeck FC, Boffa DJ, Kim AW and Tanoue
LT: The eighth edition lung cancer stage classification. Chest.
151:193–203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schwartz L, Supuran CT and Alfarouk KO:
The Warburg effect and the hallmarks of cancer. Anticancer Agents
Med Chem. 17:164–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ngo DC, Ververis K, Tortorella SM and
Karagiannis TC: Introduction to the molecular basis of cancer
metabolism and the Warburg effect. Mol Biol Rep. 42:819–823. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Allen AE and Locasale JW: Glucose
metabolism in cancer: The saga of pyruvate kinase continues. Cancer
Cell. 33:337–339. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Israelsen WJ and Vander Heiden MG:
Pyruvate kinase: Function, regulation and role in cancer. Semin
Cell Dev Biol. 43:43–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu J, Chen M, Gao S, Yuan J, Zhu Z and Zou
X: LY294002 inhibits the Warburg effect in gastric cancer cells by
downregulating pyruvate kinase M2. Oncol Lett. 15:4358–4364.
2018.PubMed/NCBI
|
|
8
|
Li J, Cheng D, Zhu M, Yu H, Pan Z, Liu L,
Geng Q, Pan H, Yan M and Yao M: OTUB2 stabilizes U2AF2 to promote
the Warburg effect and tumorigenesis via the AKT/mTOR signaling
pathway in non-small cell lung cancer. Theranostics. 9:179–195.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu QF, Liu Y, Fan Y, Hua SN, Qu HY, Dong
SW, Li RL, Zhao MY, Zhen Y, Yu XL, et al: Alpha-enolase promotes
cell glycolysis, growth, migration, and invasion in non-small cell
lung cancer through FAK-mediated PI3K/AKT pathway. J Hematol Oncol.
8:222015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao W, Li W, Dai W, Huang N and Qiu J:
LINK-A promotes cell proliferation through the regulation of
aerobic glycolysis in non-small-cell lung cancer. Onco Targets
Ther. 11:6071–6080. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen H, Zhang M, Zhang W, Li Y, Zhu J,
Zhang X, Zhao L, Zhu S and Chen B: Downregulation of BarH-like
homeobox 2 promotes cell proliferation, migration and aerobic
glycolysis through Wnt/β-catenin signaling, and predicts a poor
prognosis in non-small cell lung carcinoma. Thorac Cancer.
9:390–399. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu T and Lu YR: BCYRN1, a c-MYC-activated
long non-coding RNA, regulates cell metastasis of non-small-cell
lung cancer. Cancer Cell Int. 15:362015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Zheng L, Jiang A, Mo Y and Gong Q:
Identification of the biological affection of long noncoding RNA
BC200 in Alzheimer's disease. Neuroreport. 29:1061–1067. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Booy EP, McRae EK, Koul A, Lin F and
McKenna SA: The long non-coding RNA BC200 (BCYRN1) is critical for
cancer cell survival and proliferation. Mol Cancer. 16:1092017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: The 7th Edition of the AJCC Cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lim SO, Li CW, Xia W, Lee HH, Chang SS,
Shen J, Hsu JL, Raftery D, Djukovic D, Gu H, et al: EGFR signaling
enhances aerobic glycolysis in triple-negative breast cancer cells
to promote tumor growth and immune escape. Cancer Res.
76:1284–1296. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kong XZ, Hu SS, Sun Z, Zuo LH, Kang J, Zhu
ZF, Tian X and Zhang XJ: Regulation of aerobic glycolysis by long
non-coding RNAs in cancer. Biochem Biophys Res Commun. 479:28–32.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Goncalves MD and Cantley LC: A glycolysis
outsider steps into the cancer spotlight. Cell Metab. 28:3–4. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen R, Zhu S, Fan XG, Wang H, Lotze MT,
Zeh HJ III, Billiar TR, Kang R and Tang D: High mobility group
protein B1 controls liver cancer initiation through yes-associated
protein-dependent aerobic glycolysis. Hepatology. 67:1823–1841.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Lu JH, Wu QN, Jin Y, Wang DS, Chen
YX, Liu J, Luo XJ, Meng Q, Pu HY, et al: LncRNA LINRIS stabilizes
IGF2BP2 and promotes the aerobic glycolysis in colorectal cancer.
Mol Cancer. 18:1742019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li T, Sun X and Jiang X: UCA1 involved in
the metformin-regulated bladder cancer cell proliferation and
glycolysis. Tumour Biol. 39:10104283177108232017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang P, Cao L, Fan P, Mei Y and Wu M:
LncRNA-MIF, a c-Myc-activated long non-coding RNA, suppresses
glycolysis by promoting Fbxw7-mediated c-Myc degradation. EMBO Rep.
17:1204–1220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song J, Wu X, Liu F, Li M, Sun Y, Wang Y,
Wang C, Zhu K, Jia X, Wang B and Ma X: Long non-coding RNA PVT1
promotes glycolysis and tumor progression by regulating miR-497/HK2
axis in osteosarcoma. Biochem Biophys Res Commun. 490:217–224.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peng J, Hou F, Feng J, Xu SX and Men XY:
Long non-coding RNA BCYRN1 promotes the proliferation and
metastasis of cervical cancer via targeting microRNA-138 in vitro
and in vivo. Oncol Lett. 15:5809–5818. 2018.PubMed/NCBI
|
|
26
|
Gu L, Lu L, Zhou D and Liu Z: Long
noncoding RNA BCYRN1 promotes the proliferation of colorectal
cancer cells via Up-regulating NPR3 expression. Cell Physiol
Biochem. 48:2337–2349. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ren H, Yang X, Yang Y, Zhang X, Zhao R,
Wei R, Zhang X and Zhang Y: Upregulation of LncRNA BCYRN1 promotes
tumor progression and enhances EpCAM expression in gastric
carcinoma. Oncotarget. 9:4851–4861. 2017.PubMed/NCBI
|
|
28
|
Zhang XY, Tang XY, Ma LJ, Guo YL, Li XS,
Zhao LM, Tian CJ, Cheng DJ, Chen ZC and Zhang LX: Schisandrin B
down-regulated lncRNA BCYRN1 expression of airway smooth muscle
cells by improving miR-150 expression to inhibit the proliferation
and migration of ASMC in asthmatic rats. Cell Prolif.
50:e123822017. View Article : Google Scholar
|
|
29
|
Jin L, Li Y, Liu J, Yang S, Gui Y, Mao X,
Nie G and Lai Y: Tumor suppressor miR-149-5p is associated with
cellular migration, proliferation and apoptosis in renal cell
carcinoma. Mol Med Rep. 13:5386–5392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Y, Zheng X, Zhang Z, Zhou J, Zhao G,
Yang J, Xia L, Wang R, Cai X, Hu H, et al: MicroRNA-149 inhibits
proliferation and cell cycle progression through the targeting of
ZBTB2 in human gastric cancer. PLoS One. 7:e416932012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang F, Ma YL, Zhang P, Shen TY, Shi CZ,
Yang YZ, Moyer MP, Zhang HZ, Chen HQ, Liang Y and Qin HL: SP1
mediates the link between methylation of the tumour suppressor
miR-149 and outcome in colorectal cancer. J Pathol. 229:12–24.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao L, Liu L, Dong Z and Xiong J: miR-149
suppresses human non-small cell lung cancer growth and metastasis
by inhibiting the FOXM1/cyclin D1/MMP2 axis. Oncol Rep.
38:3522–3530. 2017.PubMed/NCBI
|
|
33
|
Qian B, Zhao L, Wang X, Xu J, Teng F, Gao
L and Shen R: RETRACTED: miR-149 regulates the proliferation and
apoptosis of cervical cancer cells by targeting GIT1. Biomed
Pharmacother. 105:1106–1116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dayton TL, Jacks T and Vander Heiden MG:
PKM2, cancer metabolism, and the road ahead. EMBO Rep.
17:1721–1730. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dong G, Mao Q, Xia W, Xu Y, Wang J, Xu L
and Jiang F: PKM2 and cancer: The function of PKM2 beyond
glycolysis. Oncol Lett. 11:1980–1986. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Papadaki C, Sfakianaki M, Lagoudaki E,
Giagkas G, Ioannidis G, Trypaki M, Tsakalaki E, Voutsina A,
Koutsopoulos A, Mavroudis D, et al: PKM2 as a biomarker for
chemosensitivity to front-line platinum-based chemotherapy in
patients with metastatic non-small-cell lung cancer. Br J Cancer.
111:1757–1764. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He J, Xie G, Tong J, Peng Y, Huang H, Li
J, Wang N and Liang H: Overexpression of microRNA-122 re-sensitizes
5-FU-resistant colon cancer cells to 5-FU through the inhibition of
PKM2 in vitro and in vivo. Cell Biochem Biophys. 70:1343–1350.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang HJ, Hsieh YJ, Cheng WC, Lin CP, Lin
YS, Yang SF, Chen CC, Izumiya Y, Yu JS, Kung HJ and Wang WC: JMJD5
regulates PKM2 nuclear translocation and reprograms HIF-1α-mediated
glucose metabolism. Proc Natl Acad Sci USA. 111:279–284. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Luan W, Wang Y, Chen X, Shi Y, Wang J,
Zhang J, Qian J, Li R, Tao T, Wei W, et al: PKM2 promotes glucose
metabolism and cell growth in gliomas through a mechanism involving
a let-7a/c-Myc/hnRNPA1 feedback loop. Oncotarget. 6:13006–13018.
2015. View Article : Google Scholar : PubMed/NCBI
|