Introduction
Breast cancer is prevalent among women worldwide
(1) and the incidence is
increasing rapidly with a large number of new cases being diagnosed
each year, especially in areas where it was previously not
prevalent, including a number of developing countries, such as
China, Brazil and Mexico (2–4). The
development of breast cancer is a complex process involving
multiple factors and a variety of cytokines, such as IL-6 and TGF-β
(5). Tumor migration and invasion
contribute to cancer development, recurrence and mortality in
patients with breast cancer, and 90% of patients with cancer suffer
mortality due to metastasis (6).
At present, except for chemotherapy, no effective drug treatment
has been identified that can directly inhibit tumor migration and
invasion in patients.
Metastasis is a programmed process that involves the
following steps: Malignant tumor cells separate from the original
location; invade the adjacent tissue via the surrounding
extracellular matrix and stromal cell layers; survive in the
circulatory system; translocate to distant tissues; re-initiate
proliferative capabilities; and subsequently develop into a new
tumor (7). Tumor metastasis
displays a number of unique characteristics, including migration
and invasion; however, the mechanisms of tumor migration and
invasion have not been fully determined. Recently, it was
identified that epithelial-mesenchymal transition (EMT), which is
the transformation of epithelial cells into mesenchymal cells, can
significantly enhance the motility of cancer cells (8).
EMT serves a vital role in tissue repair, embryonic
developmental processes, including gastrulation, somite
dissociation and neural crest development, and a number of
diseases, for example, breast cancer (9). EMT is considered to occur prior to
tumor migration and invasion (10). During EMT, epithelial cells lose
cell adhesions and undergo cytoskeletal alterations (11). In breast cancer, EMT is associated
with local invasion and migration, and is marked by the loss of
epithelial properties and the acquisition of mesenchymal properties
(12). Previous studies
demonstrated that a variety of factors can induce EMT, including
transforming growth factor β (TGF-β), Wnt, Notch and fibroblast
growth factor (13–16). Among these inducers, the
Wnt/β-catenin signaling pathway, which can be divided into
classical and non-classical signaling pathways, has attracted
increasing attention (17). Wnt
has been identified as the promoter of the pathway (18) and, as an upstream factor, β-catenin
affects the expression of downstream factors of the signaling
pathway, including c-Myc and Cyclin D1 (19). Previous studies have suggested that
the Wnt/β-catenin signaling pathway can influence developmental
embryonic processes, determine cell polarity and regulate cell
proliferation (20–23). Therefore, as the vital signaling
pathway during EMT, dysfunctions in the Wnt/β-catenin signaling
pathway are associated with a number of malignant tumors, including
colorectal and ovarian cancer (24). Khramtsov et al (25) demonstrated that the Wnt signaling
pathway enhanced migration and invasion during breast cancer. Lin
et al (26) observed that
suppressing the activity of the Wnt/β-catenin signaling pathway
reduced the invasiveness of osteosarcoma cells by reversing EMT.
Therefore, drugs that can interfere with the Wnt signaling pathway
during EMT may have potential therapeutic effects on breast
cancer.
Astragalus polysaccharide (APS) is one of the
main bioactive components extracted from the root of Astragalus
membranaceus, a traditional Chinese medicine that has been used
for thousands of years (27).
Previous studies demonstrated that APS displays immunoregulatory,
antitumor, antioxidant and anti-inflammatory effects (28,29).
Li et al (30) observed
that APS stimulated the immune response and inhibited breast cancer
cell growth. Lai et al (31) also demonstrated that APS displayed
anticancer activities in hepatoma cells by inhibiting cell growth,
and increasing the spleen and thymus indices. Additionally, Li
et al (32) demonstrated
that APS increased the sensitivity of ovarian cancer cells to
cisplatin, potentially by activating the JNK signaling pathway.
Zhou et al (33) also
demonstrated that the combination of APS and a polysaccharopeptide
had potent inhibitory effects on lung cancer. However, few studies
have identified the anti-migratory and anti-invasion effects of APS
on breast cancer. The aim of the present study was to investigate
the effect of APS on the migration and invasion of breast cancer
cells, and to identify the underlying molecular mechanisms.
Materials and methods
Cell lines and reagents
MCF-7 and MDA-MB-231 cells (Shanghai Cellular
Research Institute) were maintained in DMEM (HyClone; GE Healthcare
Life Sciences) supplemented with 10% FBS (HyClone; GE Healthcare
Life Sciences) and 1% penicillin/streptomycin. Cells were incubated
at 37°C with 5% CO2. Both cell lines were used in all
subsequent experiments. MCF-7 and MDA-MB-231 cells were cultured
with APS (800 µg/ml) and LiCl (10 mM, Sigma-Aldrich; Merck KGaA)
for 24 h for further investigation of the inhibitory effect of
APS.
Cell proliferation assay
Cells were plated (5×103 cells/well) into
96-well plates and cultured overnight. According to previous
literature (32,34,35),
cells were treated with 0, 25, 50, 100, 200, 400, 800 or 1,600
µg/ml APS (purity, >98%; Tianjin Cinorch Pharmaceutical Co.,
Ltd.) at 37°C for 24 h. Subsequently, 10 µl MTT (Sigma-Aldrich;
Merck KGaA) was added to each well for 4 h. Following the MTT
incubation, the medium was removed and the formazan crystals were
dissolved in 150 µl DMSO. Proliferation was analyzed at a
wavelength of 568 nm using a plate reader.
Wound healing assay
Cells (5×105 cells/well) were plated
in6-well plates and incubated in DMEM containing 10% FBS at 37°C.
When the cells reached 100% confluency, a 10-µl pipette tip was
used to make a single scratch through the center of the well and
PBS was used to remove the dislodged cells. Following treatment
with APS (0, 200, 400 or 800 µg/ml), cells were incubated in
serum-free DMEM for 24 h. The wounds were observed at 0 and 24 h
post-APS treatment using an inverted light microscope
(magnification, ×100). The relative area compared with control
group was measured.
Transwell Matrigel invasion assay
Transwell plates (24-well inserts; diameter, 6 mm;
pore size, 8 µm; Corning Life Sciences) were used to assess the
effect of APS on the invasion of breast cancer cells. Cells were
resuspended and diluted to a concentration of 2×105
cells/ml in serum-free DMEM. Transwell membranes were precoated
with Matrigel for 4 h at 37°C. Cells and different concentrations
of APS (0, 200, 400 or 800 µg/ml) were added to the upper chambers
of the Transwell plates. Complete medium (DMEM supplemented with
10% FPS and 1% penicillin/streptomycin) was plated in the lower
chambers of the Transwell plates. Following incubation for 24 h,
the Transwell membranes were removed and carefully cleaned using
PBS. Subsequently, the invading cells were stained with crystal
violet at room temperature for 20 min. Stained cells were counted
using an inverted light microscope (magnification, ×200).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Cells were treated with different concentrations of
APS (0, 200, 400 or 800 µg/ml) for 24 h. Total RNA was extracted
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and the concentration of RNA was measured at
wavelength of 260/280 nm using a spectrophotometer. RT-qPCR was
performed using the reverse transcription kit and the SYBR Green
Master mix (Vazyme, China) according to the manufacturer's protocol
and the ABI 7500 real-time PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The thermocycling conditions were:
Initial denaturation at 95°C for 10 min, then 40 cycles of
denaturation at 95°C for 30 sec and annealing and elongation at
60°C for 30 sec. The 2−ΔΔCq method was used for data
analysis (36). The following
primer pairs were used for qPCR: GAPDH forward,
5′-TCAAGAAGGTGGTGAAGCAGG-3′ and reverse,
5′-TCAAAGGTGGAGGAGTGGGT-3′; Wnt1 forward, 5′-CGCCTGTAACAGCTCGTCG-3′
and reverse, 5′-CGTGGCAGCACCAGTGGAAG-3′; β-catenin forward,
5′-CCAAGTGGGTGGTATAGAGG-3′ and reverse, 5′-AGTCCATAGTGAAGGCGAAC-3′;
E-cadherin forward, 5′-CGTAGCAGTGACGAATGTGG-3′ and reverse,
5′-CTGGGCAGTGTAGGATGTGA-3′; Snail forward,
5′-ATGCACATCCGAAGCCACA-3′ and reverse, 5′-TGACATCTGAGTGGGTCTGG-3′;
vimentin forward, 5′-TGAGTACCGGAGACAGGTGCAG-3′ and reverse,
5′-TAGCAGCTTCAACGGCAAAGTTC-3′; c-Myc forward,
5′-AACACACAACGTCTTGGAGC-3′ and reverse, 5′-GCACAAGAGTTCCGTAGCTG-3′;
and Cyclin D1 forward, 5′-CGGACTACAGGGGAGTTTTG-3′ and reverse,
5′-AGGAGGTTGGCATCGGGGT-3′. mRNA levels were normalized to the
internal reference gene GAPDH.
Western blot analysis
Cells were treated with 0, 200, 400 or 800 µg/ml APS
for 24 h. The nuclear and cytoplasmic proteins were extracted from
the cells using the Nuclear and Cytoplasmic Protein Extraction kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. The BCA method was used to protein
determination. Proteins (40 µg per lane) were separated by SDS-PAGE
on 10% gels and transferred to PVDF membranes. Subsequently, the
membranes were blocked with skimmed milk at room temperature for 2
h. Then the membranes were incubated at 4°C overnight with primary
antibodies targeted against: Vimentin (Cell Signaling Technology,
Inc., cat. no. 5741, dilution 1:1,000), Snail (Cell Signaling
Technology, Inc., cat. no. 3879, dilution 1:1,000), E-cadherin (BD
Biosciences, cat. no. 610812, dilution 1:1,000), β-catenin (Cell
Signaling Technology, Inc., cat. no. 8480, dilution 1:1,000), Wnt1
(Santa Cruz Biotechnology, Inc., cat. no. SC-514531, dilution
1:1,000), c-Myc (Santa Cruz Biotechnology, Inc., cat. no. SC-40,
dilution 1:1,000), Cyclin D1 (Santa Cruz Biotechnology, Inc., cat.
no. SC-8396, dilution 1:1,000), Histone H3 (Bioworld Technology,
Inc., cat. no. BS1405, dilution 1:1,000) and GAPDH (Santa Cruz
Biotechnology, Inc., cat. no. SC-47724, dilution 1:1,000).
Following primary incubation, the membranes were incubated with
horseradish peroxidase-conjugated secondary antibodies (OriGene
Technologies, Inc., cat. nos. TA130024 and TA130005, dilution
1:5,000) for 2 h at room temperature. Immunoreactive bands were
visualized using enhanced chemiluminescence (ECL) kit (Applygen
Technologies Inc.). Histone H3 and GAPDH were used as the loading
controls. The photographic density was quantitated and analyzed
using Glyko BandScan 5.0 software (Glyko).
Immunofluorescence staining
Cells at 60–80% confluence were seeded into
4-chamber dishes and incubated with different concentrations of APS
(0, 200, 400 or 800 µg/ml) for 24 h. Subsequently, cells were fixed
with 4% paraformaldehyde at room temperature for 15 min. Then cells
were blocked with 5% normal goat serum (Boster Biological
Technology.) at room temperature for 30 min. Subsequently, cells
were incubated withanti-Ki67 (Abcam, ab15580, dilution 1:200) and
anti-E-cadherin (BD Biosciences, cat. no. 610812, dilution 1:1,000)
primary antibodies at 4°C overnight. Following primary incubation,
cells were incubated with a fluorescence-labeled secondary antibody
(Boster Biological Technology., cat. no. BA1032, dilution 1:100) at
37°C for 30 min. Then cells were counterstained with DAPI at 37°C
for 5 min. Immunofluorescence staining was observed using a
biological fluorescent microscope (magnification, ×400). The images
were analyzed by Image-Pro Plus 6.0 (Media Cybernetics).
Statistical analysis
Data are presented as the mean ± standard deviation
of ≥3 repeated experiments. Statistical analyses were performed
using SPSS software (version 22.0; IBM Corp.). One-way ANOVA
followed by Dunnett's or Tukey's post hoc test was used to analyze
the data. P<0.05 was considered to indicate a statistically
significant difference.
Results
APS inhibits breast cancer cell
proliferation
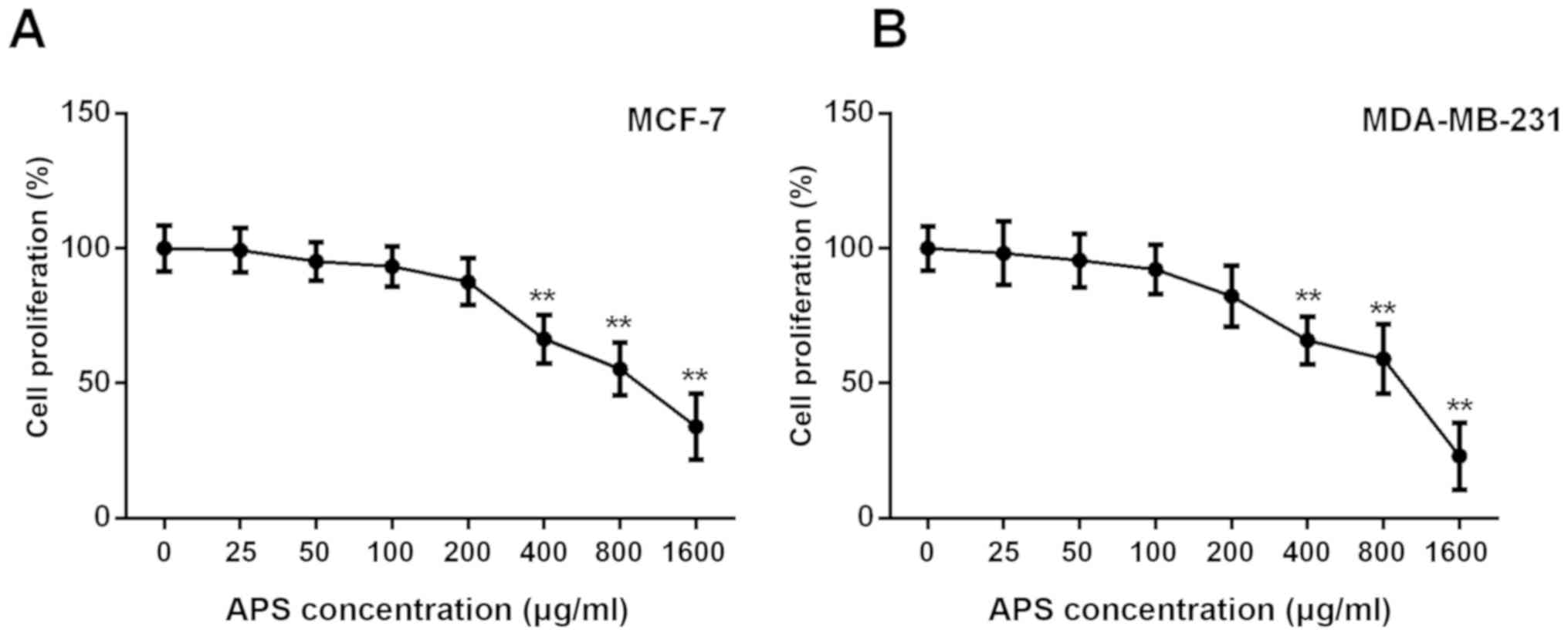
The MTT assay demonstrated that the proliferation of
MCF-7 and MDA-MB-231 cells was not significantly different between
cells treated with 0, 25, 50 and 100 µg/ml APS; however, the
proliferation of MCF-7 and MDA-MB-231 cells was significantly
inhibited by APS concentrations ≥400 µg/ml compared with the
untreated group (Fig. 1). The
proliferation rate of MCF-7 cells treated with 400, 800 and 1,600
µg/ml APS was significantly decreased by 33.6, 44.7 and 66.0%,
respectively, compared with the untreated group (Fig. 1A). Similarly, the proliferation
rate of MDA-MB-231 cells treated with 400, 800 and 1,600 µg/ml APS
was significantly decreased by 34.0, 40.9 and 76.9%, respectively,
compared with the untreated group (Fig. 1B). The IC50 value of APS
at 24 h was 945 µg/ml for MCF-7 cells and 817 µg/ml for MDA-MB-231
cells.
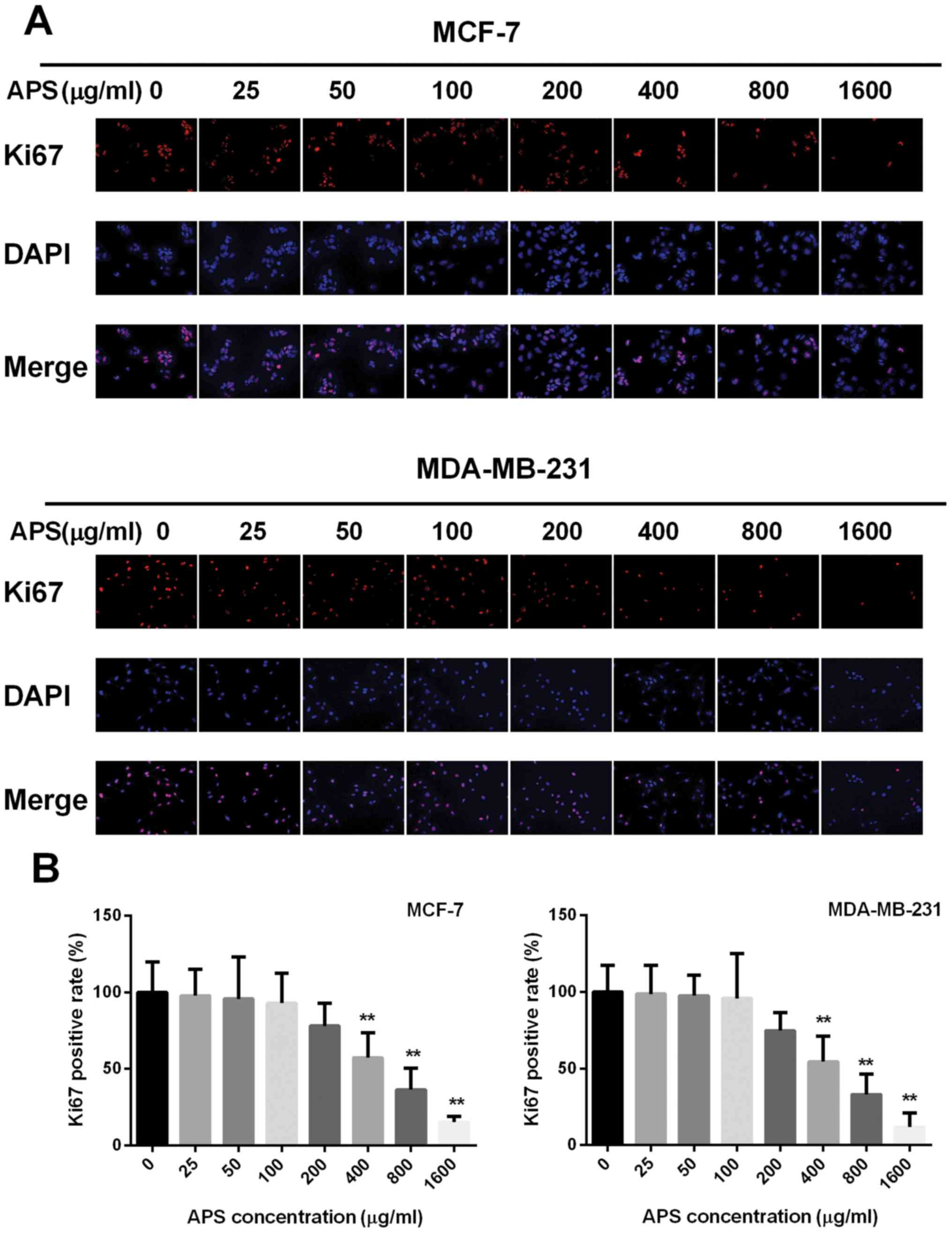
The effect of APS on cell proliferation was also
assessed using a Ki67 immunofluorescence assay. The percentage of
Ki67 positive cells decreased with increased APS concentration
(Fig. 2). The highest APS
concentration (1,600 µg/ml) demonstrated the most significant
anti-proliferative effects on MCF-7 and MDA-MB-231 cells compared
with the untreated group. Therefore, the lower APS concentrations
(0, 200, 400 and 800 µg/ml) that decreased the proliferation of
MCF-7 and MDA-MB-231 cells, compared with the untreated groups,
were selected for subsequent experiments (Fig. 2).
APS suppresses breast cancer cell
migration
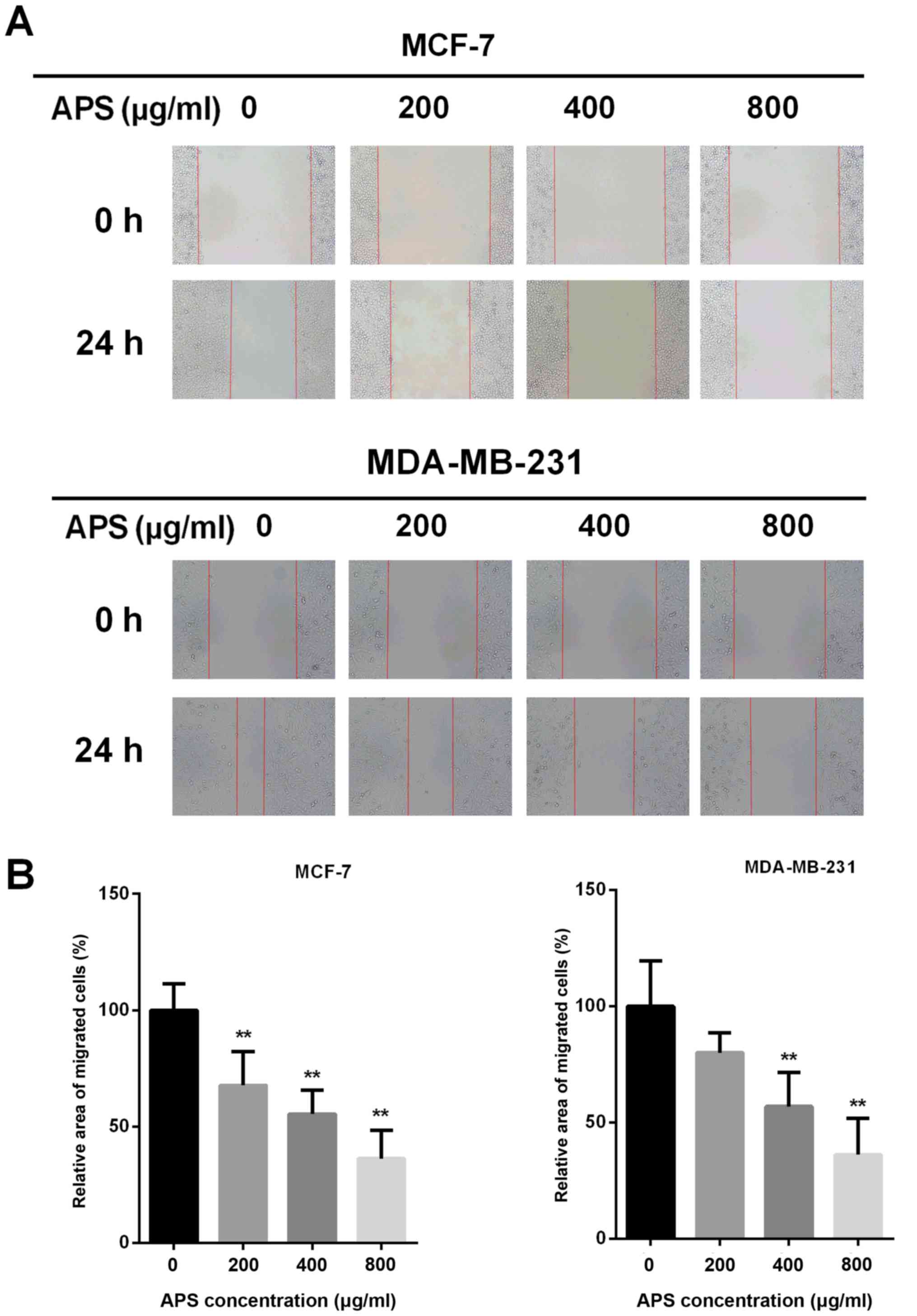
Due to EMT being closely associated with cancer cell
migration, a wound healing assay was performed to assess the effect
of APS on cell migration (Fig.
3A). The migratory potential of MCF-7 cells was decreased by
32.2, 44.6 and 63.7% after treatment with 200, 400 and 800 µg/ml
APS for 24 h, respectively, compared with the control group
(Fig. 3B). In MDA-MB-231 cells,
the migratory potential was decreased by 19.9, 43.1 and 63.8% after
treatment with 200, 400 and 800 µg/ml APS for 24 h, respectively,
compared with the control group (Fig.
3B).
APS inhibits breast cancer cell
invasion
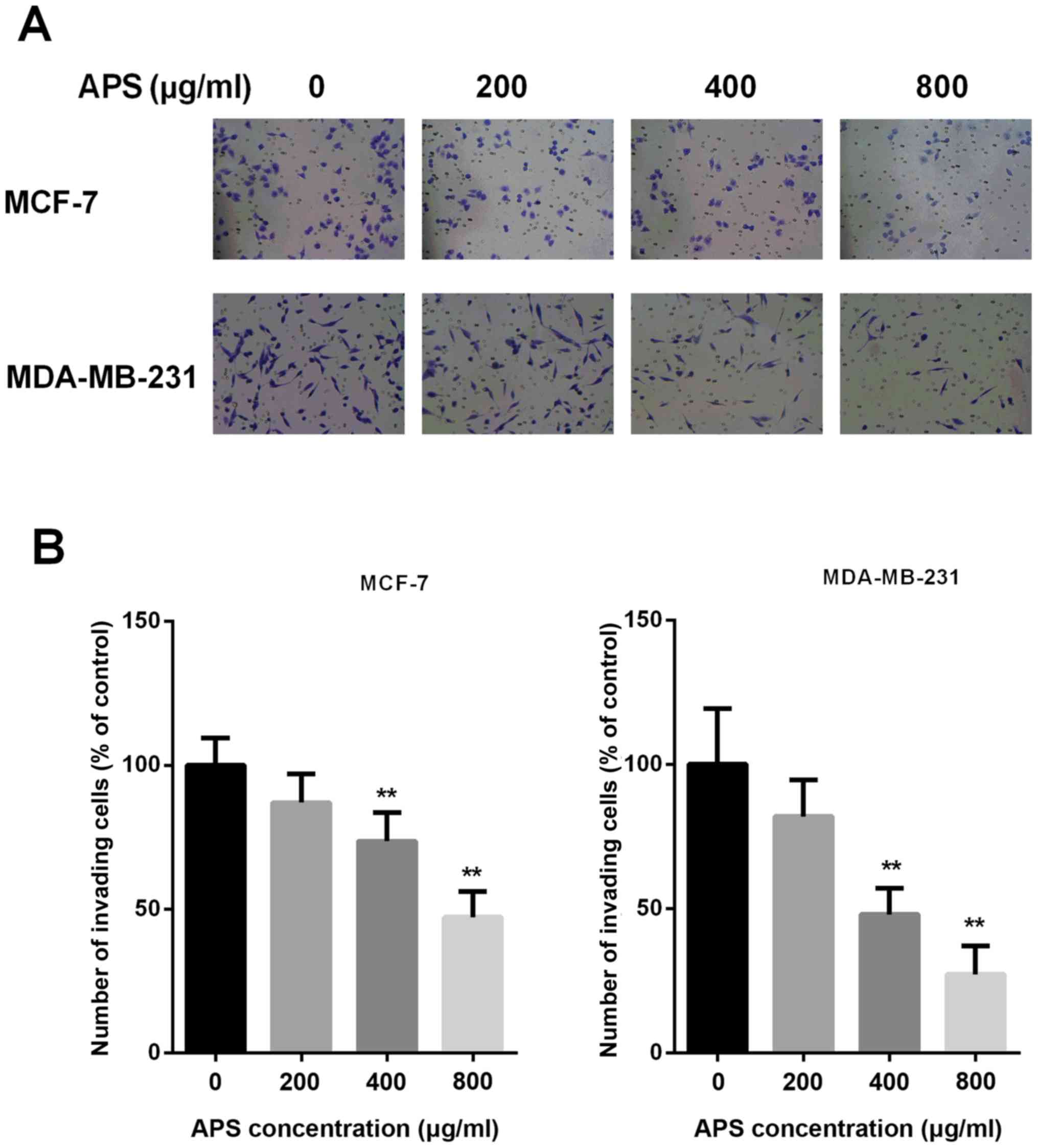
Invasion is an important characteristic that is
closely associated with EMT; therefore, Transwell Matrigel assays
were performed to assess whether APS inhibited the invasion of
breast cancer cells (Fig. 4A). The
results demonstrated that treatment with 200, 400 and 800 µg/ml APS
decreased the number of invasive MCF-7 cells by 13.1, 26.4 and
52.9%, respectively, compared with the control group (Fig. 4B). The number of invasive
MDA-MB-231 cells was also decreased by 18.0, 52.0 and 72.7%
following treatment with 200, 400 and 800 µg/ml APS, respectively,
compared with the control group (Fig.
4B). The results suggested that APS inhibited the invasion of
cells in a dose-dependent manner.
APS decreases the expression of
EMT-associated molecules
EMT is closely associated with tumor metastasis. To
further investigate whether EMT was involved in the inhibitory
effect of APS on breast cancer cell migration and invasion, the
mRNA and protein expression levels of Snail, vimentin and
E-cadherin were measured using RT-qPCR and western blot analysis,
respectively (Fig. 5). The mRNA
expression levels of Snail were significantly decreased by APS (400
and 800 µg/ml) in MCF-7 and MDA-MB-231 cells compared with the
control group (Fig. 5A).
Furthermore, MCF-7 cells treated with 200, 400 and 800 µg/ml APS
demonstrated significantly decreased Snail protein expression
compared with the control group (Fig.
5D). Similarly, Snail protein expression was decreased by 19.5,
36.6 and 54.2% in MDA-MB-231 cells treated with 200, 400 and 800
µg/ml APS, respectively, compared with the control group (Fig. 5D). The mRNA expression of vimentin
was also significantly decreased by APS (400 and 800 µg/ml) in
MCF-7 and MDA-MB-231 cells compared with the control group
(Fig. 5A). In MCF-7 cells treated
with 200, 400 and 800 µg/ml APS, vimentin protein expression was
significantly decreased by 15.0, 36.0 and 51.8%, respectively,
compared with the control group (Fig.
5D). MDA-MB-231 cells also demonstrated significantly decreased
vimentin protein expression levels following treatment with 400 and
800 µg/ml APS compared with the control group (Fig. 5D).
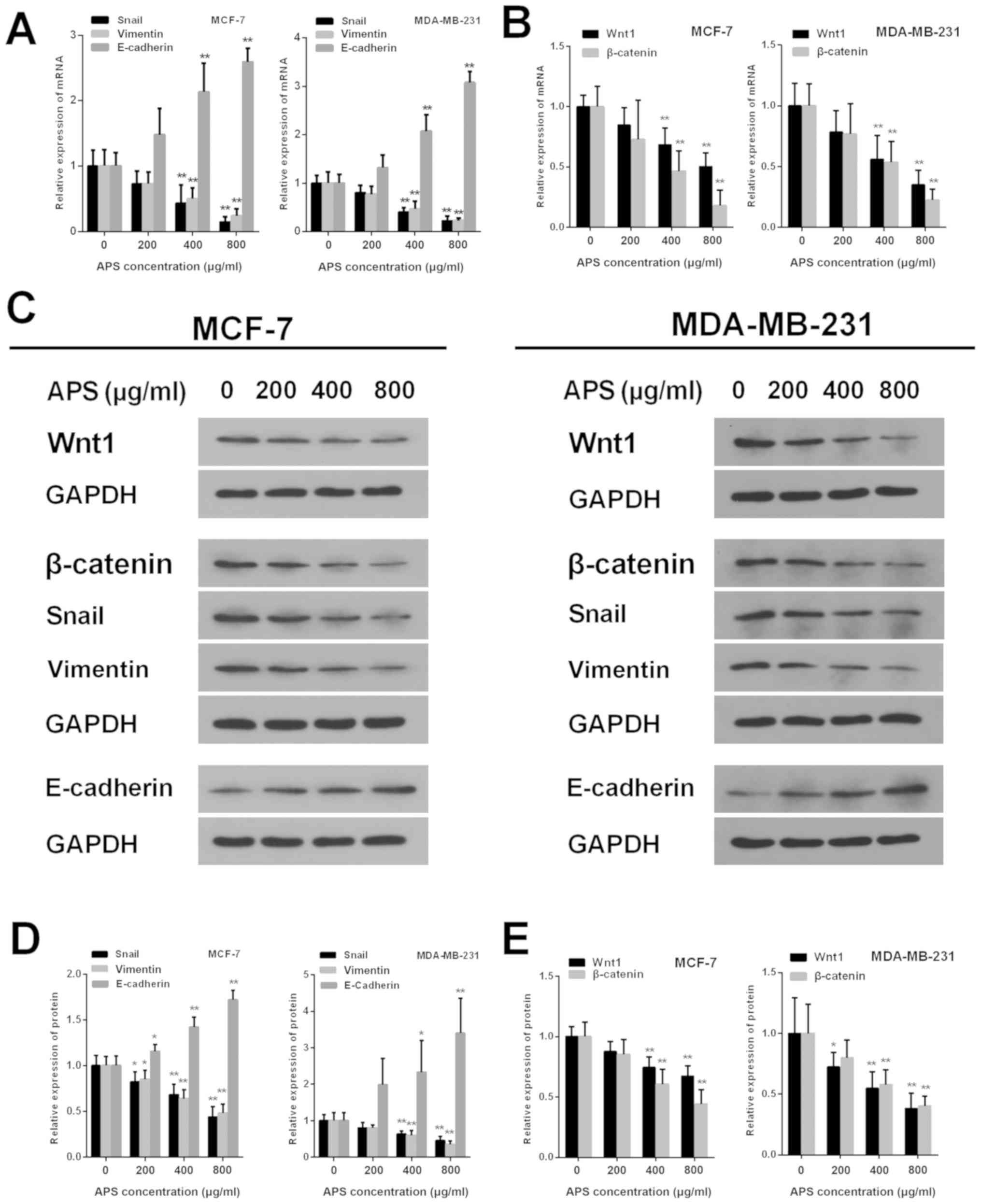 | Figure 5.APS alters the expression of Wnt1,
β-catenin and EMT-related molecules. mRNA expression levels of (A)
Snail, vimentin, E-cadherin, (B) Wnt1 and β-catenin were assessed
using reverse transcription-quantitative PCR. (C) Western blotting
was used to determine the protein expression levels of Wnt,
β-catenin and EMT-related molecules. The protein expression levels
of (D) Snail, vimentin, E-cadherin, (E) Wnt1 and β-catenin were
quantified. *P<0.05 and **P<0.01 vs. the respective control
group. APS, Astragalus polysaccharide; EMT,
epithelial-mesenchymal transition. |
The protein level of E-cadherin increased in MCF-7
and MDA-MB-231 cells as APS concentration increased. The mRNA
expression of E-cadherin was significantly increased in MCF-7 and
MDA-MB-231 cells treated with 400 and 800 µg/ml APS compared with
the control group (Fig. 5A).
Compared with the control group, the protein level of E-cadherin
inMCF-7 cells treated with 200, 400 and 800 µg/ml APS was
significantly increased (Fig. 5D).
Similarly, the protein level of E-cadherin was significantly
increased by 2.33 and 3.40-fold in MDA-MB-231 cells treated with
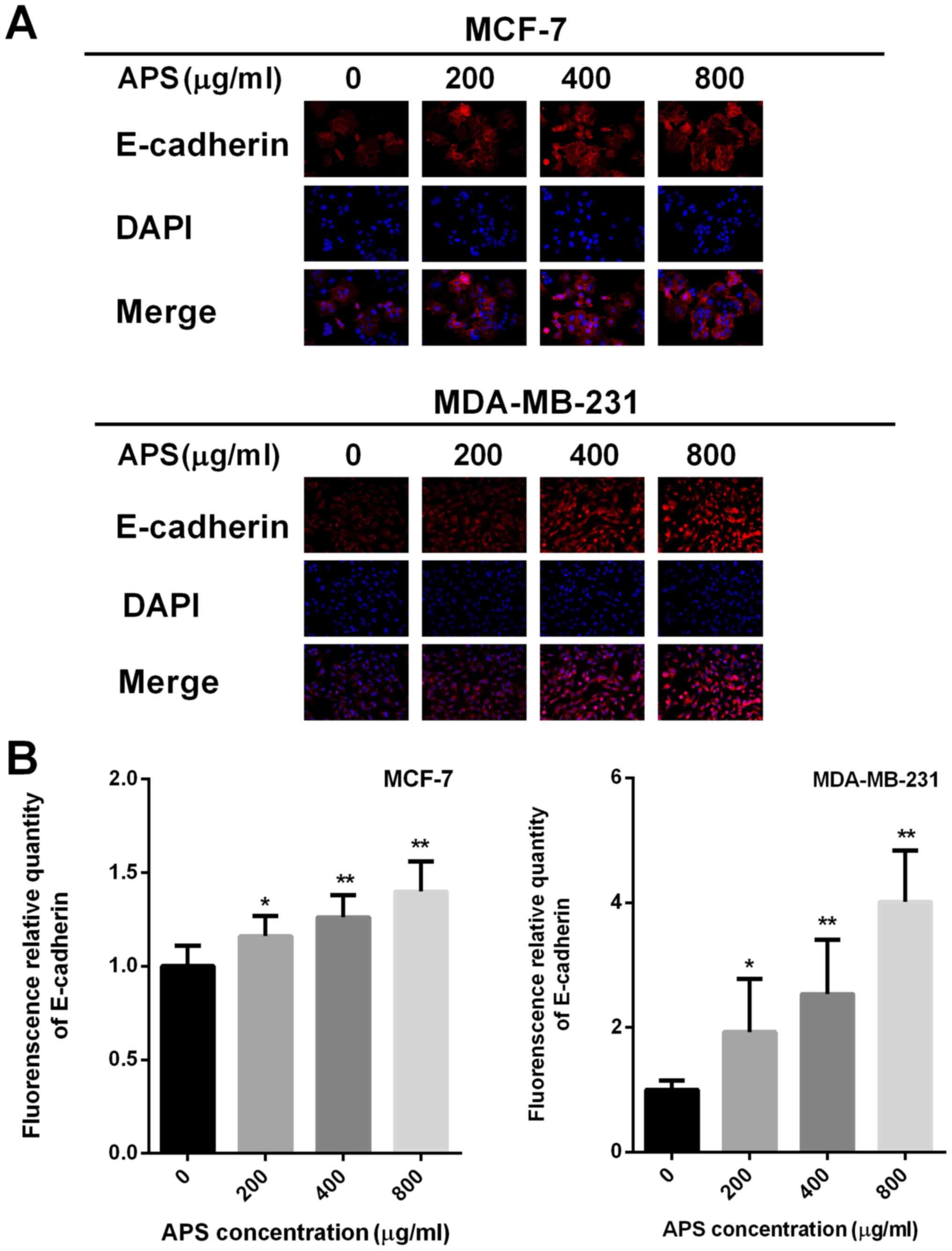
400 and 800 µg/ml APS, respectively (Fig. 5D). Immunofluorescence staining was
also used to assess E-cadherin expression (Fig. 6A). The relative fluorescence of
E-cadherin staining was significantly increased in MCF-7 and
MDA-MB-231 cells treated with 400 and 800 µg/ml APS compared with
the control group (Fig. 6B).
APS inhibits EMT by suppressing the
activity of the Wnt/β-catenin signaling pathway
The Wnt signaling pathway is an important classical
pathway in EMT (17). The present
study investigated whether APS exerted its antitumor effect by
suppressing the activity of the Wnt/β-catenin signaling pathway.
Wnt1 is one of the activators of the signaling pathway; therefore,
the mRNA and protein expression levels of Wnt1 were assessed by
RT-qPCR and western blotting, respectively. The mRNA expression
levels of Wnt1 were significantly decreased in MCF-7 and MDA-MB-231
cells treated with 400 and 800 µg/ml APS compared with the control
group (Fig. 5B). Wnt1 protein
expression was also significantly decreased in MCF-7 and MDA-MB-231
cells treated with 400 and 800 µg/ml APS compared with the control
group (Fig. 5E). β-catenin is a
key factor that activates upstream and downstream factors of the
Wnt/β-catenin signaling pathway (19). The mRNA and protein expression
levels of β-catenin were also significantly decreased in MCF-7 and
MDA-MB-231 cells treated with 400 and 800 µg/ml APS compared with
the control group (Fig. 5B and
E).
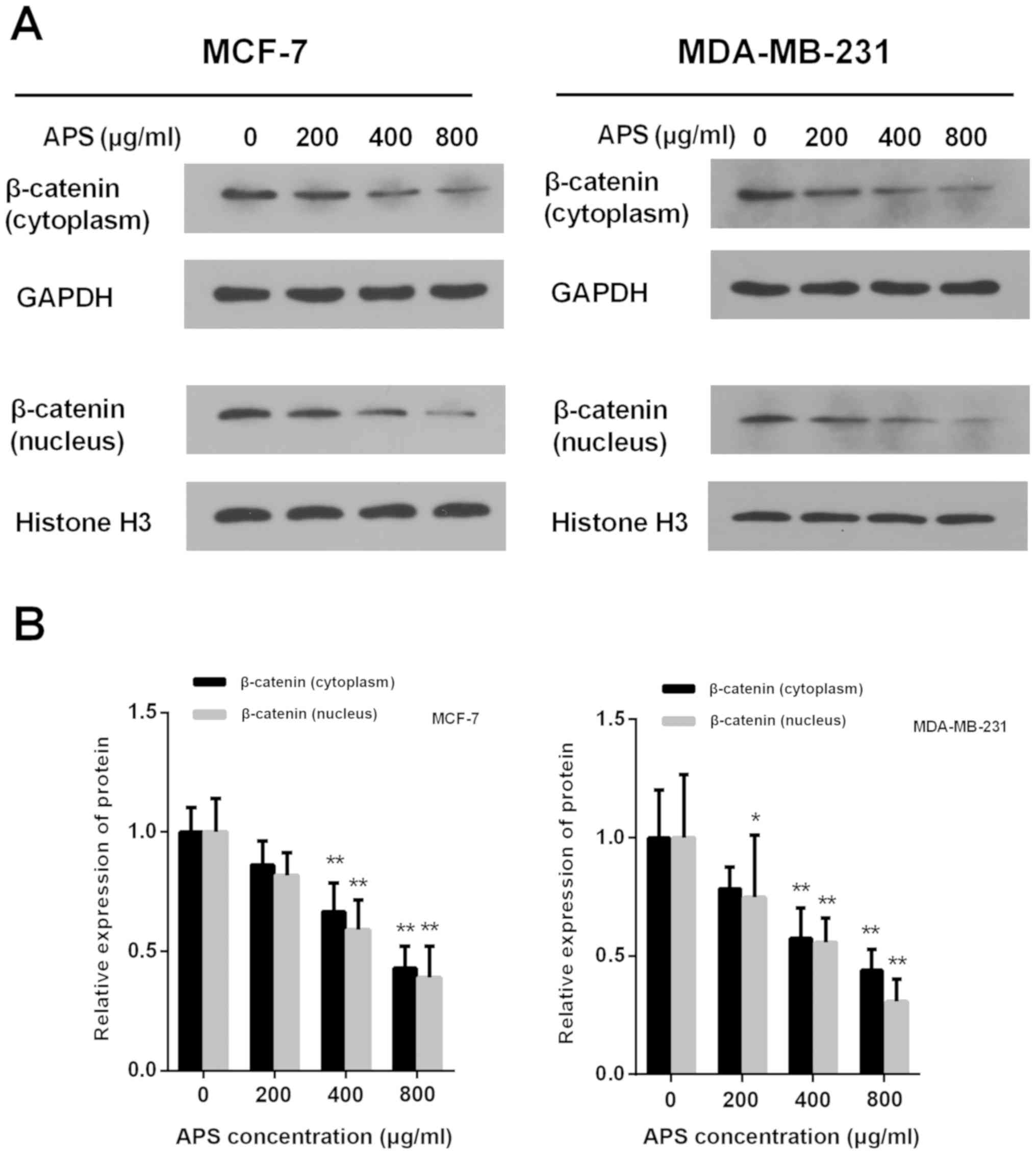
The accumulation of β-catenin in the cytoplasm and
its translocation into the nucleus are critical for the activation
of β-catenin target gene transcription (18). Treatment with APS (400 and 800
µg/ml) significantly decreased the protein expression level of
cytoplasmic and nuclear β-catenin compared with the control group
(Fig. 7). In MCF-7 cells treated
with 200, 400 and 800 µg/ml APS, the expression of cytoplasmic
β-catenin was decreased by 13.8, 33.4 and 57.1%, respectively,
compared with the control group (Fig.
7B). In MDA-MB-231 cells treated with 200, 400 and 800 µg/ml
APS, the expression of cytoplasmic β-catenin was decreased by 21.6,
42.5 and 56.1%, respectively, compared with the control group
(Fig. 7B). In MCF-7 and MDA-MD-231
cells treated with 400 and 800 µg/ml APS, the expression of nuclear
β-catenin was significantly decreased compared with the control
group (Fig. 7B).
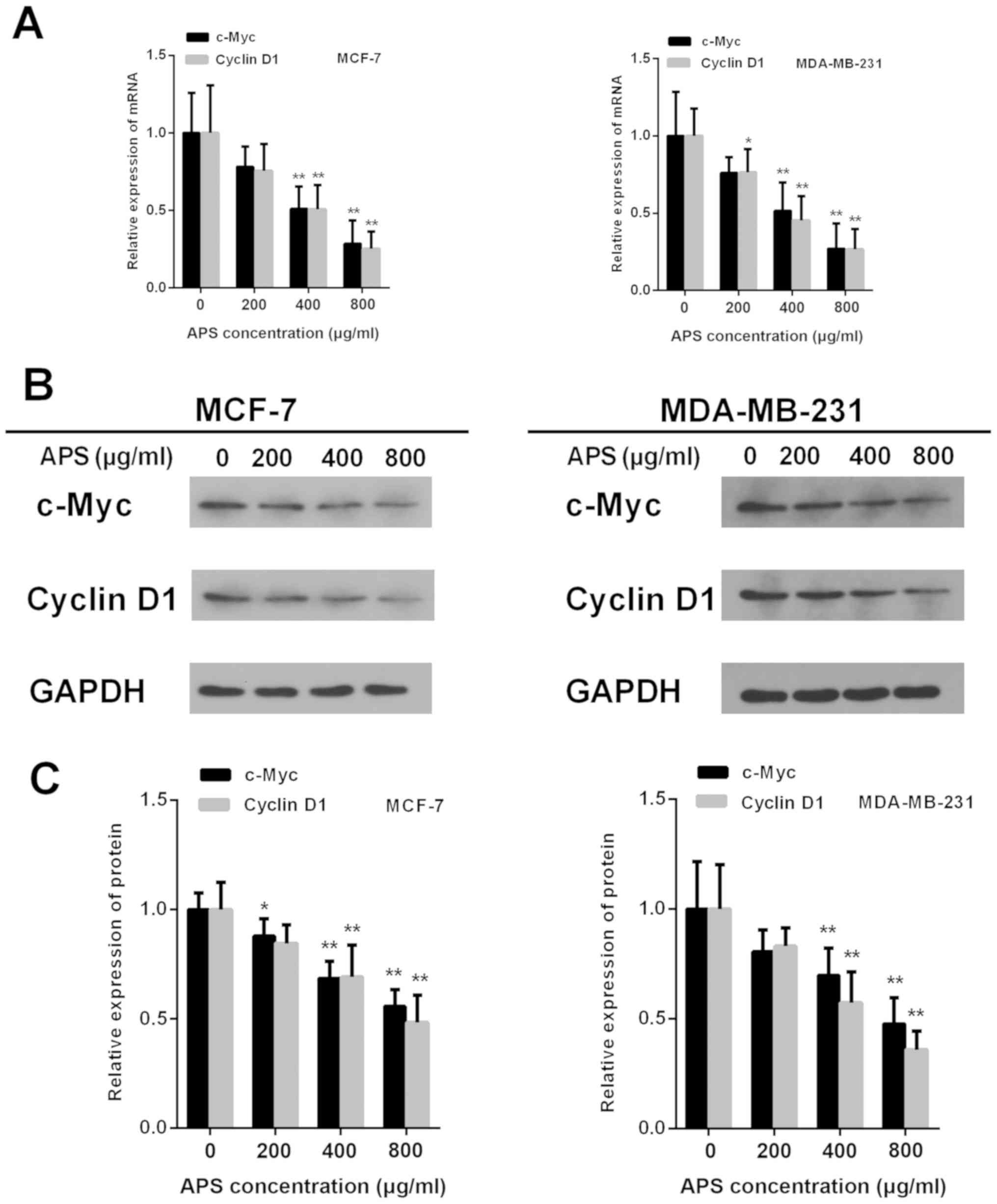
Cyclin D1and c-Myc are downstream factors of the
Wnt/β-catenin signaling pathway (18). The mRNA expression of c-Myc was
significantly decreased in MCF-7 and MDA-MB-231 cells treated with
400 and 800 µg/ml APS cells compared with the control group
(Fig. 8A). The results also
demonstrated that the protein expression of c-Myc was significantly
decreased by 12.1, 31.4 and 44.3% in MCF-7 cells treated with 200,
400 and 800 µg/ml APS, respectively, compared with the control
group (Fig. 8B and C). In
MDA-MB-231 cells, the protein expression level of c-Myc was
significantly decreased by 30.2 and 52.3% following treatment with
400 and 800 µg/ml APS, respectively, compared with the control
group (Fig. 8B and C).
Furthermore, the mRNA and protein expression levels of Cyclin D1
were significantly decreased in MCF-7 and MDA-MB-231 cells treated
with 400 and 800 µg/ml APS compared with the control group
(Fig. 8).
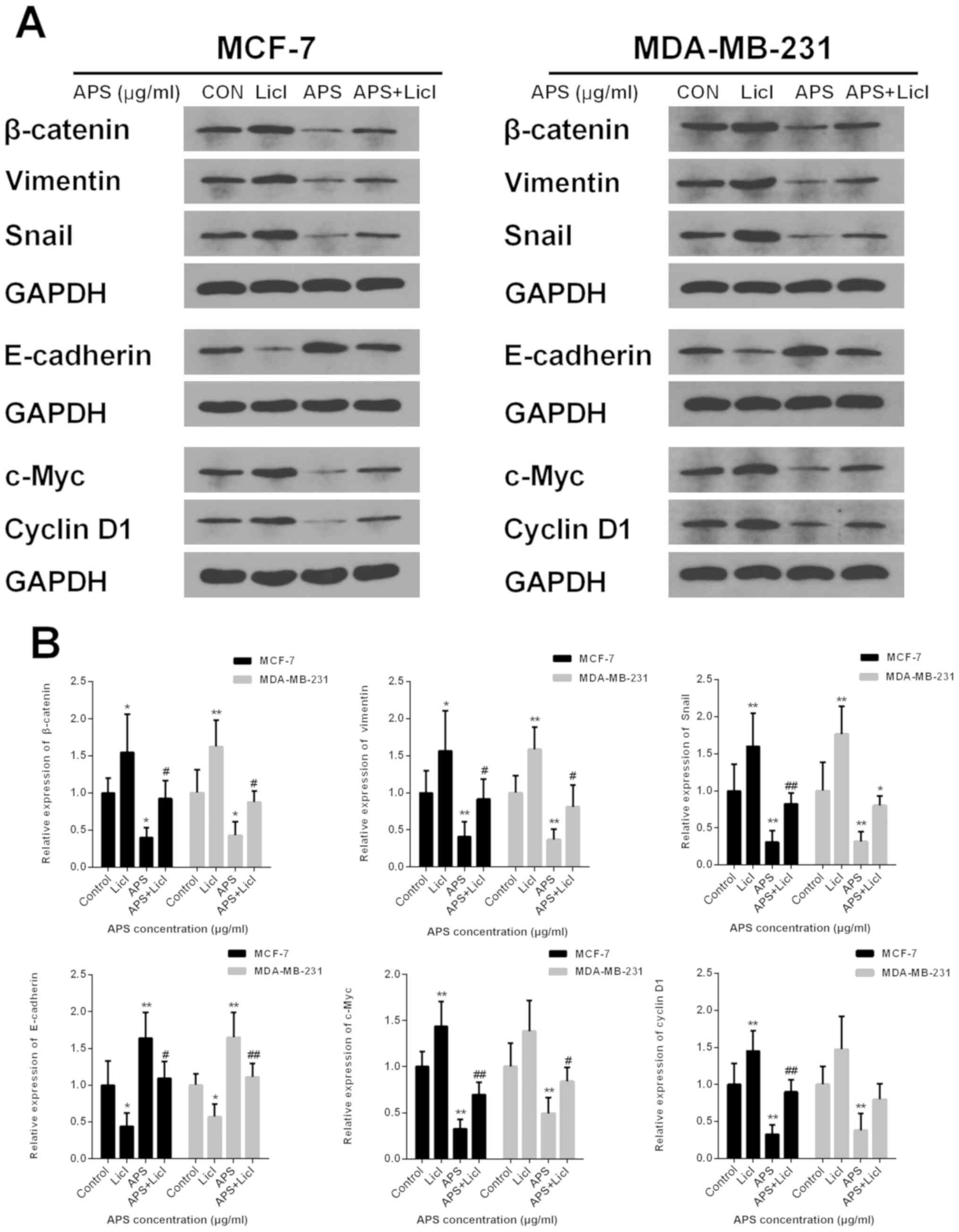
Lithium chloride (LiCl) reverses the
inhibitory effect of APS on the Wnt/β-catenin signaling
pathway
LiCl, an agonist of the Wnt/β-catenin signaling
pathway, was used in the present study to further investigate the
inhibitory effect of APS. MCF-7 and MDA-MB-231 cells were cultured
with APS (800 µg/ml) and LiCl (10 mM) for 24 h. The protein
expression levels of β-catenin, Snail, vimentin, c-Myc and Cyclin
D1 were significantly decreased and the protein level of E-cadherin
was significantly increased in the APS-treated group compared with
the control group (Fig. 9).
However, LiCl treatment reversed the APS-induced effects on protein
expression (Fig. 9). The results
further suggested that the mechanism underlying the inhibitory
effects of APS involved the Wnt/β-catenin signaling pathway.
Discussion
Tumor migration and invasion are important causes of
increased mortality in patients with breast cancer (6). EMT can significantly enhance the
migratory ability of cancer cells, including breast cancer cells,
which can lead to tumor migration and invasion (9); therefore, drugs targeting the EMT
pathway may reduce mortality in patients with breast cancer
(37). In recent years, a number
of studies have reported that traditional Chinese medicine displays
significant anticancer effects (38–40).
AM, which is an important component of the qi-supplementing
formula, is widely used in China (41) and the main extract of AM is APS.
APS is primarily composed of α-1,4-(1,6)-glucan, rhamnus-galacturonic acid
polysaccharide I, arabic-galactopolysaccharide and
arabic-galactoprotein polysaccharide (42). APS has been demonstrated to display
therapeutic effects in multiple diseases, including a variety of
tumor diseases, such as hepatocellular carcinoma and lung carcinoma
(28–33,43).
However, there are only a few studies that have reported the effect
of APS on breast cancer, and these studies primarily focused on the
effects of APS on the proliferation and immune regulation of breast
cancer cells (30,44). The antimigratory and anti-invasion
effects, as well as the specific mechanisms of APS on breast cancer
cells, are still unclear. The results of the present study
suggested that APS significantly decreased the migration and
invasion of breast cancer cells in vitro. Furthermore, the
results suggested that the mechanism underlying the effects of APS
was closely associated with EMT by downregulating the activity of
the Wnt/β-catenin signaling pathway. Therefore, the present study
may provide rationale for further investigation into the effects of
the bioactive components of APS on breast cancer.
MCF-7 and MDA-MB-231 cells differ greatly in cell
morphology, estrogen receptor expression, and migration and
invasion potential (45).
MDA-MB-231 cells proliferate rapidly and are prone to tumorigenesis
(45), whereas MCF-7 cells are an
estrogen receptor-positive cell line. The migration and invasion of
MCF-7 cells is weaker compared with MDA-MB-231 cells; however,
previous studies have demonstrated that the migratory and invasive
abilities of MCF-7 cells are significantly enhanced under the
effect of multiple molecules, including estrogen and Wnt/β-catenin
signaling pathway-associated cytokines (46,47).
A number of studies have used the two aforementioned cell lines to
study the migration and invasion of breast cancer cells; therefore,
the two cell lines were selected for the present study (48–50).
Yang et al (51)
demonstrated that APS inhibited the proliferation of hepatocellular
carcinoma cells partly via immunomodulation. In the present study,
an MTT assay and Ki67 immunostaining assay demonstrated that APS
decreased the proliferation of breast cancer cells in a
dose-dependent manner. Tumor migration and invasion involves the
following steps: Tumor cells undergo a decrease in homogenous
adhesion; detach from the original site of the tumor; adhere to the
extracellular matrix (ECM); degrade the ECM; penetrate the ECM
surrounding blood vessels; and enter circulation (52). In the present study, the number of
migratory cells was significantly decreased in the APS-treated
groups compared with the untreated group, as measured by a wound
healing assay. The present results suggested that APS inhibited
breast cancer cell migration. Additionally, the results of the
Transwell Matrigel assay further suggested that the number of
invading cells was decreased in the APS-treated groups compared
with the untreated group. The present results suggested that APS
may inhibit breast cancer cell migration and invasion.
EMT is a vital mechanism during cell growth, tissue
repair and organ fibrosis (9);
however, it is also closely associated with tumor invasion and
migration (53). Previous studies
have demonstrated that via the EMT process, cells gain invasive and
anti-apoptotic abilities (10–12,54).
The hallmark changes of EMT include a reduction in intercellular
adhesion and a loss of cell polarity (55). It is well established that the Wnt
signaling pathway can induce EMT, which subsequently leads to cell
invasion and migration (56,57),
and a number of previous studies demonstrated that the
Wnt/β-catenin signaling pathway promotes breast cancer progression
(58–60). The Wnt signaling pathway is
composed of two different intracellular signaling pathways, the
canonical and non-canonical pathway (61–63).
The canonical pathway involves the activation of β-catenin as a
result of triggering by Wnt1, Wnt3a and Wnt8. However, in the
non-canonical pathway, Wnt proteins can trigger other effectors,
including JNK (64–66). Wnt1 is one of the activators of the
Wnt/β-catenin pathway (18).
Following activation by Wnt1, β-catenin translocates to the nucleus
to regulate the transcription of downstream genes, including Snail,
vimentin and E-cadherin (57).
Snail can induce EMT and increase cell motility to promote cell
differentiation, and it has been reported that Snail expression is
abnormal during tumor proliferation and metastasis (67). Vimentin is a downstream molecule of
the tumor-related signaling pathway that is important for EMT
during malignant transformation (68). Previous studies demonstrated that
vimentin can regulate cell-cell adhesion to promote tumor migration
and invasion (69,70). Furthermore, E-cadherin participates
in the regulation of epithelium formation, maintaining homeostasis
and forming adhesive connections. E-cadherin is also involved in
the formation of polarized sheets of epithelial cells, and
downregulation of E-cadherin can lead to decreased cell adhesion
and aggregation (71). It was
identified that E-cadherin displays a decreasing trend during EMT
due to increased promoter methylation, which occurs during breast
cancer (72,73). The present study assessed the
expression of EMT-associated molecules, including Wnt1, β-catenin,
Snail, vimentin and E-cadherin. Consistent with previous studies
(72,73), the present results suggested that
Wnt1, β-catenin, Snail and vimentin expression was increased, while
E-cadherin expression was decreased in MCF-7 and MDA-MB-231 cells.
However, APS treatment significantly downregulated Wnt1, vimentin,
Snail and β-catenin expression, and upregulated E-cadherin
expression in a dose-dependent manner. The intracellular
localization of β-catenin is important for the activation of
downstream target genes (57);
therefore, cytoplasmic and nuclear β-catenin expression levels
following APS treatment were determined. The present results
demonstrated that APS reduced the expression levels of cytoplasmic
and nuclear β-catenin in a dose-dependent manner. Additionally,
Cyclin D1 and c-Myc are produced following β-catenin activation
(19). c-Myc, a nuclear protein
transcription factor, is associated with the activation of genes
that are related to multiple cellular processes, such as
proliferation, differentiation, and apoptosis. c-Myc is also
important for cell cycle modulation and malignant transformation of
cells (74). Cyclin D1, an
oncogene, can regulate the function of the cell cycle function, and
its abnormal expression can disrupt the cell cycle and promote
tumorigenesis (75,76). The present results suggested that
APS decreased the transcription of c-Myc and Cyclin D1 in a
dose-dependent manner. However, further investigation into the
effect of APS when protein degradation is inhibited is
required.
The results also demonstrated that LiCl, an agonist
of the Wnt signaling pathway, partly reversed the inhibitory
effects of APS on breast cancer cells. Therefore, the results of
the present study suggested that APS may modulate EMT in breast
cancer via the Wnt/β-catenin signaling pathway. Collectively, the
present results suggested that the Wnt/β-catenin signaling pathway
may serve as an effective target for the inhibition of breast
cancer cell migration and invasion.
In conclusion, the present study suggested that APS
significantly decreased the migration and invasion of breast cancer
cells in vitro by inhibiting EMT and modulating the
expression of components of the Wnt/β-catenin signaling pathway.
The present study identified a novel mechanism of APS and
demonstrated that APS may serve as a potential therapeutic agent
for breast cancer. The effect of APS in other EMT-associated
pathways, including the TGF-β signaling pathway, and which
component of APS displays the antitumor effect requires further
investigation.
Acknowledgements
The authors would like to thank Mrs. Qiyan Wang
(School of Life Science, Beijng University of Chinese Medicine) for
editing the manuscript.
Funding
The present study was supported by The Qingdao
Post-Doctoral Research Project and The Natural Science Foundation
of Shandong Province (grant no. ZR2017BH067).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SY and HW conceived and design the study. SY drafted
the manuscript. SY and WX performed the experiments. SS, BY and GW
performed the statistical analyses. HW and BY revised the
manuscript. All authors read and approved the final manuscript to
be published. All authors agreed to be accountable for the work in
ensuring that questions related to the integrity of any part of the
work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tao Z, Shi A, Lu C, Song T, Zhang Z and
Zhao J: Breast cancer: Epidemiology and etiology. Cell Biochem
Biophys. 72:333–338. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sharma R: Breast cancer incidence,
mortality and mortality-to-incidence ratio (MIR) are associated
with human development, 1990–2016: Evidence from Global Burden of
Disease Study 2016. Breast Cancer. 26:428–445. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bhat V, Allan AL and Raouf A: Role of the
microenvironment in regulating normal and cancer stem cell
activity: Implications for breast cancer progression and therapy
response. Cancers (Basel). 11(pii): E12402019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Benson JR and Jatoi I: The global breast
cancer burden. Future Oncol. 8:697–702. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weidle UH, Birzele F and Nopora A:
MicroRNAs as potential targets for therapeutic intervention with
metastasis of non-small cell lung cancer. Cancer Genomics
Proteomics. 16:99–119. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yan L, Xu F and Dai CL: Relationship
between epithelial-to-mesenchymal transition and the inflammatory
microenvironment of hepatocellular carcinoma. J Exp Clin Cancer
Res. 37:2032018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Medici D, Shore EM, Lounev VY, Kaplan FS,
Kalluri R and Olsen BR: Conversion of vascular endothelial cells
into multipotent stem-like cells. Nat Med. 16:1400–1406. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bai J, Kwok WC and Thiery JP: Traditional
Chinese medicine and regulatory roles on epithelial-mesenchymal
transitions. Chin Med. 14:342019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shang BQ, Li ML, Quan HY, Hou PF, Li ZW,
Chu SF, Zheng JN and Bai J: Functional roles of circular RNAs
during epithelial-to-mesenchymal transition. Mol Cancer.
18:1382019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sjoberg E, Meyrath M, Milde L, Herrera M,
Lovrot J, Hagerstrand D, Frings O, Bartish M, Rolny C, Sonnhammer
E, et al: A novel ACKR2-dependent role of fibroblast-derived CXCL14
in Epithelial-to-mesenchymal transition and metastasis of breast
cancer. Clin Cancer Res. 25:3702–3717. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McCormack N and O'Dea S: Regulation of
epithelial to mesenchymal transition by bone morphogenetic
proteins. Cell Signal. 25:2856–2862. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Farahmand L, Darvishi B, Majidzadeh AK and
Madjid Ansari A: Naturally occurring compounds acting as potent
anti-metastatic agents and their suppressing effects on Hedgehog
and WNT/β-catenin singaling pathways. Cell Prolif. 502017.doi:
10.1111/cpr.12299.
|
|
15
|
Heldin CH, Vanlandewijck M and Moustakas
A: Regulation of EMT by TGFβ in cancer. FEBS Lett. 586:1959–1970.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Espinoza I and Miele L: Deadly crosstalk:
Notch signaling at the intersection of EMT and cancer stem cells.
Cancer Lett. 341:41–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang P, Gao XY, Yang SQ, Sun ZX, Dian LL,
Qasim M, Phyo AT, Liang ZS and Sun YF: Jatrorrhizine inhibits
colorectal carcinoma proliferation and metastasis through
Wnt/β-catenin signaling pathway and epithelial-mesenchymal
transition. Drug Des Devel Ther. 13:2235–2247. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ram Makena M, Gatla H, Verlekar D,
Sukhavasi S, K Pandey M and C Pramanik K: Wnt/β-catenin signaling:
The culprit in pancreatic carcinogenesis and therapeutic
resistance. Int J Mol Sci. 20(pii): E42422019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang W, Li Q, Yang T, Li D, Ding F, Sun H
and Bai G: Anti-cancer effect of Aquaporin 5 silencing in
colorectal cancer cells in association with inhibition of
Wnt/β-catenin pathway. Cytotechnology. 70:615–624. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu XL, Meng J, Zhang XT, Liang XH, Zhang
F, Zhao GR and Zhang T: ING5 inhibits lung cancer invasion and
epithelial-mesenchymal transition by inhibiting the WNT/β-catenin
pathway. Thorac Cancer. 10:848–855. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Okura T, Ohkawara B, Takegami Y, Ito M,
Masuda A, Seki T, Ishiguro N and Ohno K: Mianserin suppresses
R-spondin 2-induced activation of Wnt/β-catenin signaling in
chondrocytes and prevents cartilage degradation in a rat model of
osteoarthritis. Sci Rep. 9:28082019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brunt L and Scholpp S: The function of
endocytosis in Wnt signaling. Cell Mol Life Sci. 75:785–795. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Krishnamurthy N and Kurzrock R: Targeting
the Wnt/beta-catenin pathway in cancer: Update on effectors and
inhibitors. Cancer Treat Rev. 62:50–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Khramtsov AI, Khramtsova GF, Tretiakova M,
Huo D, Olopade OI and Goss KH: Wnt/beta-catenin pathway activation
is enriched in basal-like breast cancers and predicts poor outcome.
Am J Pathol. 176:2911–2920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin CH, Ji T, Chen CF and Hoang BH: Wnt
signaling in osteosarcoma. Adv Exp Med Biol. 804:33–45. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bamodu OA, Kuo KT, Wang CH, Huang WC, Wu
ATH, Tsai JT, Lee KY, Yeh CT and Wang LS: Astragalus
polysaccharides (PG2) Enhances the M1 polarization of
macrophages, functional maturation of dendritic cells, and T
cell-mediated anticancer immune responses in patients with lung
cancer. Nutrients. 11(pii): E22642019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jin M, Zhao K, Huang Q and Shang P:
Structural features and biological activities of the
polysaccharides from Astragalus membranaceus. Int J Biol
Macromol. 64:257–266. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xie JH, Jin ML, Morris GA, Zha XQ, Chen
HQ, Yi Y, Li JE, Wang ZJ, Gao J, Nie SP, et al: Advances on
bioactive polysaccharides from medicinal plants. Crit Rev Food Sci
Nutr. 56 (Suppl 1):S60–S84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li W, Hu X, Wang S, Wang H, Parungao R,
Wang Y, Liu T and Song K: Detection and evaluation of Anti-cancer
efficiency of astragalus polysaccharide via a tissue engineered
tumor model. Macromol Biosci. 18:e18002232018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lai X, Xia W, Wei J and Ding X:
Therapeutic effect of Astragalus polysaccharides on hepatocellular
carcinoma H22-bearing mice. Dose Response. 15:15593258166851822017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li C, Hong L, Liu C, Min J, Hu M and Guo
W: Astragalus polysaccharides increase the sensitivity of SKOV3
cells to cisplatin. Arch Gynecol Obstet. 297:381–386. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou X, Liu Z, Long T, Zhou L and Bao Y:
Immunomodulatory effects of herbal formula of astragalus
polysaccharide (APS) and polysaccharopeptide (PSP) in mice with
lung cancer. Int J Biol Macromol. 106:596–601. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li W, Song K, Wang S, Zhang C, Zhuang M,
Wang Y and Liu T: Anti-tumor potential of astragalus
polysaccharides on breast cancer cell line mediated by macrophage
activation. Mater Sci Eng C Mater Biol Appl. 98:685–695. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang JX, Han YP, Bai C and Li Q: Notch1/3
and p53/p21 are a potential therapeutic target for APS-induced
apoptosis in non-small cell lung carcinoma cell lines. Int J Clin
Exp Med. 8:12539–12547. 2015.PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang XG, Zhu LC, Wang YJ, Li YY and Wang
D: Current advance of therapeutic agents in clinical trials
potentially targeting tumor plasticity. Front Oncol. 9:8872019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu Y, Hua W, Li Y, Xian X, Zhao Z, Liu C,
Zou J, Li J, Fang X and Zhu Y: Berberine suppresses colon cancer
cell proliferation by inhibiting the SCAP/SREBP-1 signaling
pathway-mediated lipogenesis. Biochem Pharmacol. 174:1137762019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song M, Wang X, Luo Y, Liu Z, Tan W, Ye P,
Fu Z, Lu F, Xiang W, Tang L, et al: Cantharidin suppresses gastric
cancer cell migration/invasion by inhibiting the PI3K/Akt signaling
pathway via CCAT1. Chem Biol Interact. 317:1089392020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen T, Lin J, Tang D, Zhang M, Wen F, Xue
D and Zhang H: Paris saponin H suppresses human hepatocellular
carcinoma (HCC) by inactivation of Wnt/β-catenin pathway in vitro
and in vivo. Int J Clin Exp Pathol. 12:2875–2886. 2019.PubMed/NCBI
|
|
41
|
Shan H, Zheng X and Li M: The effects of
Astragalus membranaceus active extracts on autophagy-related
diseases. Int J Mol Sci. 20(pii): E19042019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sun S, Yang S, An N, Wang G, Xu Q, Liu J
and Mao Y: Astragalus polysaccharides inhibits cardiomyocyte
apoptosis during diabetic cardiomyopathy via the endoplasmic
reticulum stress pathway. J Ethnopharmacol. 238:1118572019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhao M, Zhang ZF, Ding Y, Wang JB and Li
Y: Astragalus polysaccharide improves palmitate-induced insulin
resistance by inhibiting PTP1B and NF-κB in C2C12 myotubes.
Molecules. 17:7083–7092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhou L, Liu Z, Wang Z, Yu S, Long T, Zhou
X and Bao Y: Astragalus polysaccharides exerts immunomodulatory
effects via TLR4-mediated MyD88-dependent signaling pathway in
vitro and in vivo. Sci Rep. 7:448222017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Karamanou K, Franchi M, Vynios D and
Brezillon S: Epithelial-to-mesenchymal transition and invadopodia
markers in breast cancer: Lumican a key regulator. Semin Cancer
Biol. Aug 8–2019.(Epub ahead of print). doi:
10.1016/j.semcancer.2019.08.003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Song C, Sun P, He Q, Liu LL, Cui J and Sun
LM: Long non-coding RNA LINC01287 promotes breast cancer cells
proliferation and metastasis by activating Wnt/β-catenin signaling.
Eur Rev Med Pharmacol Sci. 23:4234–4242. 2019.PubMed/NCBI
|
|
47
|
Zhang Y, Fang J, Zhao H, Yu Y, Cao X and
Zhang B: Downregulation of microRNA-1469 promotes the development
of breast cancer via targeting HOXA1 and activating PTEN/PI3K/AKT
and Wnt/β-catenin pathways. J Cell Biochem. 120:5097–5107. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ma Y, Yan F, Wei W, Deng J, Li L, Liu L
and Sun J: Litchi seed aqueous extracts play a role in suppression
of epithelial-mesenchymal transition, invasion and migration in
breast cancer cells. Cell Cycle. 1–9. 2020.doi:
10.1080/15384101.2019.1710912 (Epub ahead of print).
|
|
49
|
Duan X, Guo G, Pei X, Wang X, Li L, Xiong
Y and Qiu X: Baicalin inhibits cell viability, migration and
invasion in breast cancer by regulating miR-338-3p and MORC4. Onco
Targets Ther. 12:11183–11193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wei T, Xiaojun X and Peilong C:
Magnoflorine improves sensitivity to doxorubicin (DOX) of breast
cancer cells via inducing apoptosis and autophagy through AKT/mTOR
and p38 signaling pathways. Biomed Pharmacother. 121:1091392020.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang B, Xiao B and Sun T: Antitumor and
immunomodulatory activity of Astragalus membranaceus
polysaccharides in H22 tumor-bearing mice. Int J Biol Macromol.
62:287–290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Venning FA, Wullkopf L and Erler JT:
Targeting ECM disrupts cancer progression. Front Oncol. 5:2242015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tsubakihara Y and Moustakas A:
Epithelial-mesenchymal transition and metastasis under the control
of transforming growth factor β. Int J Mol Sci. 19(pii): E36722018.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Vijay GV, Zhao N, Den Hollander P, Toneff
MJ, Joseph R, Pietila M, Taube JH, Sarkar TR, Ramirez-Pena E,
Werden SJ, et al: GSK3β regulates epithelial-mesenchymal transition
and cancer stem cellproperties in triple-negative breast cancer.
Breast Cancer Res. 21:372019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang Y and Zhou BP: Epithelial-mesenchymal
Transition-A Hallmark of breast cancer metastasis. Cancer Hallm.
1:38–49. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fu Y, Zheng S, An N, Athanasopoulos T,
Popplewell L, Liang A, Li K, Hu C and Zhu Y: β-catenin as a
potential key target for tumor suppression. Int J Cancer.
129:1541–1551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rahmani F, Amerizadeh F, Hassanian SM,
Hashemzehi M, Nasiri SN, Fiuji H, Ferns GA, Khazaei M and Avan A:
PNU-74654 enhances the antiproliferative effects of 5-FU in breast
cancer and antagonizes thrombin-induced cell growth via the Wnt
pathway. J Cell Physiol. 234:14123–14132. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hseu YC, Lin YC, Rajendran P, Thigarajan
V, Mathew DC, Lin KY, Way TD, Liao JW and Yang HL: Antrodia
salmonea suppresses invasion and metastasis in triple-negative
breast cancer cells by reversing EMT through the NF-κB and
Wnt/β-catenin signaling pathway. Food Chem Toxicol. 124:219–230.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang K, Liu P, Tang H, Xie X, Kong Y,
Song C, Qiu X and Xiao X: AFAP1-AS1 promotes epithelial-mesenchymal
transition and tumorigenesis through Wnt/β-catenin signaling
pathway in Triple-negative breast cancer. Front Pharmacol.
9:12482018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Jamieson C, Sharma M and Henderson BR: Wnt
signaling from membrane to nucleus: β-catenin caught in a loop. Int
J Biochem Cell Biol. 44:847–850. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Rao TP and Kuhl M: An updated overview on
Wnt signaling pathways: A prelude for more. Circ Res.
106:1798–1806. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Carayol N and Wang CY: IKKalpha stabilizes
cytosolic beta-catenin by inhibiting both canonical and
non-canonical degradation pathways. Cell Signal. 18:1941–1946.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
De A: Wnt/Ca2+ signaling
pathway: A brief overview. Acta Biochim Biophys Sin (Shanghai).
43:745–756. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang Y: Wnt/Planar cell polarity
signaling: A new paradigm for cancer therapy. Mol Cancer Ther.
8:2103–2109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Veeman MT, Axelrod JD and Moon RT: A
second canon. Functions and mechanisms of beta-catenin-independent
Wnt signaling. Dev Cell. 5:367–377. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kang Y and Massague J:
Epithelial-mesenchymal transitions: Twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Danielsson F, Peterson MK, Caldeira Araujo
H, Lautenschläger F and Gad AKB: Vimentin diversity in health and
disease. Cells. 7(pii): E1472018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tsuruta D and Jones JC: The vimentin
cytoskeleton regulates focal contact size and adhesion of
endothelial cells subjected to shear stress. J Cell Sci.
116:4977–4984. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Maskarinec G, Ju D, Fong J, Horio D, Chan
O, Loo LWM and Hernandez BY: Mammographic density and breast tissue
expression of inflammatory markers, growth factors and vimentin.
BMC Cancer. 18:11912018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hirohashi S: Inactivation of the
E-cadherin-mediated cell adhesion system in human cancers. Am J
Pathol. 153:333–339. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Aristizabal-Pachon AF and Takahashi CS:
Effect of genetics, epigenetics and variations in the
transcriptional expression of cadherin-E in breast cancer
susceptibility. Biomedica. 36:593–602. 2016.(In Spanish).
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang Q, Gun M and Hong XY: Induced
Tamoxifen resistance is mediated by increased methylation of
E-cadherin in estrogen receptor-expressing breast cancer cells. Sci
Rep. 9:141402019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Miliani de Marval PL, Macias E, Rounbehler
R, Sicinski P, Kiyokawa H, Johnson DG, Conti CJ and
Rodriguez-Puebla ML: Lack of cyclin-dependent kinase 4 inhibits
c-myc tumorigenic activities in epithelial tissues. Mol Cell Biol.
24:7538–7547. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Bertoli C, Skotheim JM and de Bruin RA:
Control of cell cycle transcription during G1 and S phases. Nat Rev
Mol Cell Biol. 14:518–528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Rubin SM: Deciphering the retinoblastoma
protein phosphorylation code. Trends Biochem Sci. 38:12–19. 2013.
View Article : Google Scholar : PubMed/NCBI
|